The Role of Matrix Metalloproteinases in Periodontal Disease

 ,
, 
 ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Matrix Metalloproteinases (MMPs)
3. The Role of MMPs in Periodontal Disease
4. MMP Inhibitors
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chapple, I.; Mealey, B.L.; Van Dyke, T.E.; Bartold, P.M.; Dommisch, H.; Eickholz, P.; Geisinger, M.L.; Genco, R.J.; Glogauer, M.; Goldstein, M.; et al. Periodontal health and gingival diseases and conditions on an intact and a reduced periodontium: Consensus report of workgroup 1 of the 2017 World Workshop on the Classification of Periodontal and Peri-Implant Diseases and Conditions. J. Periodontol. 2018, 89, 74–84. [Google Scholar] [CrossRef]
- Checchi, L.; Gatto, M.R.; Checchi, V.; Carinci, F. Bacteria prevalence in a large Italian population sample: A clinical and microbiological study. J. Biol. Regul. Homeost. Agents 2016, 30, 199–208. [Google Scholar] [PubMed]
- Checchi, L.; Montevecchi, M.; Gatto, M.R.; Trombelli, L. Retrospective study of tooth loss in 92 treated periodontal patients. J. Clin. Periodontol. 2002, 29, 651–656. [Google Scholar] [CrossRef]
- Trombelli, L.; Minenna, L.; Toselli, L.; Zaetta, A.; Checchi, L.; Checchi, V.; Nieri, M.; Farina, R. Prognostic value of a simplified method for periodontal risk assessment during supportive periodontal therapy. J. Clin. Periodontol. 2017, 44, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Leppilahti, J.M.; Ahonen, M.; Hernández, M.; Munjal, S.; Netuschil, L.; Uitto, V.J.; Sorsa, T.; Mäntylä, P. Oral rinse MMP-8 point-of-care immuno test identifies patients with strong periodontal inflammatory burden. Oral Dis. 2011, 17, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Golub, L.M.; Lee, H.M. Periodontal therapeutics: Current host-modulation agents and future directions. Periodontol. 2000 2020, 82, 186–204. [Google Scholar] [CrossRef] [PubMed]
- Letra, A.; Silva, R.M.; Rylands, R.J.; Silveira, E.M.; de Souza, A.P.; Wendell, S.K.; Garlet, G.P.; Vieira, A.R. MMP3 and TIMP1 variants contribute to chronic periodontitis and may be implicated in disease progression. J. Clin. Periodontol. 2012, 39, 707–716. [Google Scholar] [CrossRef]
- Franco, C.; Patricia, H.R.; Timo, S.; Claudia, B.; Marcela, H. Matrix metalloproteinases as regulators of periodontal inflammation. Int. J. Mol. Sci. 2017, 18, 1–12. [Google Scholar] [CrossRef]
- Preshaw, P.M. Host modulation therapy with anti-inflammatory agents. Periodontol. 2000 2018, 76, 131–149. [Google Scholar] [CrossRef]
- Visse, R.; Nagase, H. Matrix Metalloproteinases and Tissue Inhibitors of Metalloproteinases. Circ. Res. 2003, 92, 827–839. [Google Scholar] [CrossRef]
- Sternlicht, M.D.; Werb, Z. How matrix metalloproteinases regulate cell behavior. Annu. Rev. Cell Dev. Biol. 2001, 17, 463–516. [Google Scholar] [CrossRef]
- Mazzoni, A.; Breschi, L.; Carrilho, M.; Nascimento, F.D.; Orsini, G.; Ruggeri, A., Jr.; Gobbi, P.; Manzoli, L.; Tay, F.R.; Pashley, D.H.; et al. A review of the nature, role, and function of dentin non-collagenous proteins. Part II: Enzymes, serum proteins, and growth factors. Endod. Top. 2012, 21, 19–40. [Google Scholar] [CrossRef]
- Hannas, A.R.; Pereira, J.C.; Granjeiro, J.M.; Tjäderhane, L. The role of matrix metalloproteinases in the oral environment. Acta Odontol. Scand. 2007, 65, 1–13. [Google Scholar] [CrossRef]
- Van Wart, H.E.; Birkedal-Hansen, H. The cysteine switch: A principle of regulation of metalloproteinase activity with potential applicability to the entire matrix metalloproteinase gene family. Proc. Natl. Acad. Sci. USA 1990, 87, 5578–5582. [Google Scholar] [CrossRef] [PubMed]
- Gioia, M.; Monaco, S.; Fasciglione, G.F.; Coletti, A.; Modesti, A.; Marini, S.; Coletta, M. Characterization of the Mechanisms by which Gelatinase A, Neutrophil Collagenase, and Membrane-Type Metalloproteinase MMP-14 Recognize Collagen I and Enzymatically Process the Two α-Chains. J. Mol. Biol. 2007, 368, 1101–1113. [Google Scholar] [CrossRef]
- Chung, L.; Dinakarpandian, D.; Yoshida, N.; Lauer-Fields, J.L.; Fields, G.B.; Visse, R.; Nagase, H. Collagenase unwinds triple-helical collagen prior to peptide bond hydrolysis. EMBO J. 2004, 23, 3020–3030. [Google Scholar] [CrossRef]
- Lauer-Fields, J.L.; Juska, D.; Fields, G.B. Matrix metalloproteinases and collagen catabolism. Biopolymers 2002, 66, 19–32. [Google Scholar] [CrossRef]
- Garnero, P.; Borel, O.; Byrjalsen, I.; Ferreras, M.; Drake, F.H.; McQueney, M.S.; Foged, N.T.; Delmas, P.D.; Delaissé, J.M. The collagenolytic activity of cathepsin K is unique among mammalian proteinases. J. Biol. Chem. 1999, 273, 32347–32352. [Google Scholar] [CrossRef]
- Perumal, S.; Antipova, O.; Orgel, J.P. Collagen fibril architecture, domain organization, and triple-helical conformation govern its proteolysis. Proc. Natl. Acad. Sci. USA 2008, 105, 2824–2829. [Google Scholar] [CrossRef]
- Mirigian, L.S.; Makareeva, E.; Koistinen, H.; Itkonen, O.; Sorsa, T.; Stenman, U.H.; Salo, T.; Leikin, L. Collagen degradation by tumor-associated trypsins. Arch. Biochem. Biophys. 2013, 535, 111–114. [Google Scholar] [CrossRef][Green Version]
- Gomez, D.E.; Alonso, D.F.; Yoshiji, H.; Thorgeirsson, U.P. Tissue inhibitors of metalloproteinases: Structure, regulation and biological functions. Eur. J. Cell Biol. 1997, 74, 111–122. [Google Scholar]
- Woessner, J.F. Role of matrix proteases in processing enamel proteins. Connect. Tissue Res. 1998, 39, 69–73. [Google Scholar] [CrossRef]
- Lambert, E.; Dassé, E.; Haye, B.; Petitfrère, E. TIMPs as multifacial proteins. Crit. Rev. Oncol. Hematol. 2004, 49, 187–198. [Google Scholar] [CrossRef]
- Brew, K.; Dinakarpandian, D.; Nagase, H. Tissue inhibitors of metalloproteinases: Evolution, structure and function. Biochim. Biophys. Acta 2000, 1477, 267–283. [Google Scholar] [CrossRef]
- Wojtowicz-Praga, S.M.; Dickson, R.B.; Hawkins, M.J. Matrix metalloproteinase inhibitors. Investig. New Drugs 1997, 15, 61–75. [Google Scholar] [CrossRef]
- Tervahartiala, T.; Pirilä, E.; Ceponis, A.; Maisi, P.; Salo, T.; Tuter, G.; Kallio, P.; Törnwall, J.; Srinivas, R.; Konttinen, Y.T.; et al. The in vivo expression of the collagenolytic matrix metalloproteinases (MMP-2, -8, -13, and -14) and matrilysin (MMP-7) in adult and localized juvenile periodontitis. J. Dent. Res. 2000, 79, 1969–1977. [Google Scholar] [CrossRef]
- Choi, D.H.; Moon, I.S.; Choi, B.K.; Paik, J.W.; Kim, Y.S.; Choi, S.H.; Kim, C.K. Effects of sub-antimicrobial dose doxycycline therapy on crevicular fluid MMP-8, and gingival tissue MMP-9, TIMP-1 and IL-6 levels in chronic periodontitis. J. Periodontal Res. 2004, 39, 20–26. [Google Scholar] [CrossRef]
- Beklen, A.; Tüter, G.; Sorsa, T.; Hanemaaijer, R.; Virtanen, I.; Tervahartiala, T.; Konttinen, Y.T. Gingival tissue and crevicular fluid co-operation in adult periodontitis. J. Dent. Res. 2006, 85, 59–63. [Google Scholar] [CrossRef]
- Soder, B.; Airila Mansson, S.; Soder, P.O.; Kari, K.; Meurman, J. Levels of matrix metalloproteinases-8 and -9 with simultaneous presence of periodontal pathogens in gingival crevicular fluid as well as matrix metalloproteinase-9 and cholesterol in blood. J. Periodontal Res. 2006, 41, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.S.; Vamsi, G.; Sripriya, R.; Sehgal, P.K. Expression of matrix metalloproteinases (MMP-8 and -9) in chronic periodontitis patients with and without diabetes mellitus. J. Periodontol. 2006, 77, 1803–1808. [Google Scholar] [CrossRef]
- Marcaccini, A.M.; Meschiari, C.A.; Zuardi, L.R.; de Sousa, T.S.; Taba, M., Jr.; Teofilo, J.M.; Jacob-Ferreira, A.L.B.; Tanus-Santos, J.E.; Novaes, A.B., Jr.; Gerlach, R.F. Gingival crevicular fluid levels of MMP-8, MMP-9, TIMP-2, and MPO decrease after periodontal therapy. J. Clin. Periodontol. 2010, 37, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Kraft-Neumärker, M.; Lorenz, K.; Koch, R.; Hoffmann, T.; Mäntylä, P.; Sorsa, T.; Netuschil, L. Full-mouth profile of active MMP-8 in periodontitis patients. J. Periodontal Res. 2012, 47, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Charles, K.; Honibald, E.N.; Raghavendra, R.N.; Palani, A.K.P.; Ramamurthy, R.D.; Sankaralingam, T. Role of matrix metalloproteinases (MMPS) in periodontitis and its management. J. Indian Acad. Dent. Spec. Res. 2014, 1, 65–69. [Google Scholar]
- Rai, B.; Kharb, S.; Jain, R.; Anand, S.C. Biomarkers of periodontitis in oral fluids. J. Oral Sci. 2008, 50, 53–56. [Google Scholar] [CrossRef]
- Xu, L.; Yu, Z.; Lee, H.M.; Wolff, M.S.; Golub, L.M.; Sorsa, T.; Kuula, H. Characteristics of collagenase-2 from gingival crevicular fluid and peri-implant sulcular fluid in periodontitis andperi-impalntitis patients: Pilot study. Acta Odontol. Scand. 2008, 66, 219–224. [Google Scholar] [CrossRef]
- Schure, R.; Costa, K.D.; Rezaei, R.; Lee, W.; Laschinger, C.; Tenenbaum, H.C.; McCulloch, C.A. Impact of matrix metalloproteinases on inhibition of mineralization by fetuin. J. Periodontal Res. 2013, 48, 357–366. [Google Scholar] [CrossRef]
- Gursoy, U.K.; Könönen, E.; Pradhan-Palikhe, P.; Tervahartiala, T.; Pussinen, P.J.; Suominen-Taipale, L.; Sorsa, T. Salivary MMP-8, TIMP-1, and ICTP as markers of advanced periodontitis. J. Clin. Periodontol. 2010, 37, 487–493. [Google Scholar] [CrossRef]
- Emingil, G.; Tervahartiala, T.; Mãntylã, P.; Määttä, M.; Sorsa, T.; Atilla, G. Gingival Crevicular Fluid Matrix Metalloproteinase (MMP)-7, Extracellular MMP Inducer, and Tissue Inhibitor of MMP-1 Levels in Periodontal Disease. J. Periodontol. 2006, 77, 2040–2050. [Google Scholar] [CrossRef]
- Emingil, G.; Kuula, H.; Pirilä, E.; Atilla, G.; Sorsa, T. Gingival crevicular fluid laminin-5 γ2-chain levels in periodontal disease. J. Clin. Periodontol. 2006, 33, 462–468. [Google Scholar] [CrossRef]
- Emingil, G.; Kuula, H.; Sorsa, T.; Atilla, G. Gingival Crevicular Fluid Matrix Metalloproteinase-25 and -26 Levels in Periodontal Disease. J. Periodontol. 2006, 77, 664–671. [Google Scholar] [CrossRef]
- Ramseier, C.A.; Kinney, J.S.; Herr, A.E.; Braun, T.; Sugai, J.V.; Shelburne, C.A.; Rayburn, L.A.; Tran, H.M.; Singh, A.K.; Giannobile, W.V. Identification of Pathogen and Host-Response Markers Correlated with Periodontal Disease. J. Periodontol. 2009, 80, 436–446. [Google Scholar] [CrossRef] [PubMed]
- Hernández Ríos, M.; Sorsa, T.; Obregón, F.; Tervahartiala, T.; Valenzuela, M.A.; Pozo, P.; Dutzan, N.; Lesaffre, E.; Molas, M.; Gamonal, J. Proteolytic roles of matrix metalloproteinase (MMP)-13 during progression of chronic periodontitis: Initial evidence for MMP-13/MMP-9 activation cascade. J. Clin. Periodontol. 2009, 36, 1011–1017. [Google Scholar] [CrossRef] [PubMed]
- Alpagot, T.; Bell, C.; Lundergan, W.; Chambers, D.W.; Rudin, R. Longitudinal evaluation of GCF MMP-3 and TIMP-1 levels as prognostic factors for progression of periodontitis. J. Clin. Periodontol. 2001, 28, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Marcaccini, A.M.; Novaes, A.B.; Meschiari, C.A.; Souza, S.L.; Palioto, D.B.; Sorgi, C.A.; Faccioli, L.H.; Tanus-Santos, J.E.; Gerlach, R.F. Circulating matrix metalloproteinase-8 (MMP-8) and MMP-9 are increased in chronic periodontal disease and decrease after non-surgical periodontal therapy. Clin. Chim. Acta 2009, 409, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Kinane, D.F.; Darby, I.B.; Said, S.; Luoto, H.; Sorsa, T.; Tikanoja, S.; Mäntylä, P. Changes in gingival crevicular fluid matrix metalloproteinase-8 levels during periodontal treatment and maintenance. J. Periodontal Res. 2003, 38, 400–404. [Google Scholar] [CrossRef]
- Mäntylä, P.; Stenman, M.; Kinane, D.; Salo, T.; Suomalainen, K.; Tikanoja, S.; Sorsa, T. Monitoring periodontal disease status in smokers and nonsmokers using a gingival crevicular fluid matrix metalloproteinase-8-specific chair-side test. J. Periodontal Res. 2006, 41, 503–512. [Google Scholar] [CrossRef]
- Pozo, P.; Valenzuela, M.A.; Melej, C.; Zaldívar, M.; Puente, J.; Martínez, B.; Gamonal, J. Longitudinal analysis of metalloproteinases, tissue inhibitors of metalloproteinases and clinical parameters in gingival crevicular fluid from periodontitis-affected patients. J. Periodontal Res. 2005, 40, 199–207. [Google Scholar] [CrossRef]
- Mäntylä, P.; Stenman, M.; Kinane, D.F.; Salo, T.; Suomalainen, K.; Tikanoja, S.; Sorsa, T. Gingival crevicular fluid collagenase-2 (MMP-8) test stick for chair-side monitoring of periodontitis. J. Periodontal Res. 2003, 38, 436–439. [Google Scholar] [CrossRef]
- Somerville, R.P.T.; Oblander, S.A.; Apte, S.S. Matrix metalloproteinases: Old dogs with new tricks. Genome Biol. 2003, 4, 216. [Google Scholar] [CrossRef][Green Version]
- Delaissé, J.M.; Andersen, T.L.; Engsig, M.T.; Henriksen, K.; Troen, T.; Blavier, L. Matrix metalloproteinases (MMP) and cathepsin K contribute differently to osteoclastic activities. Microsc. Res. Tech. 2003, 61, 504–513. [Google Scholar] [CrossRef]
- Fuller, K.; Kirstein, B.; Chambers, T.J. Regulation and enzymatic basis of bone resorption by human osteoclasts. Clin. Sci. 2007, 112, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Huang, H.Z. The role of RANKL and MMP-9 in the bone resorption caused by ameloblastoma. J. Oral Pathol. Med. 2010, 39, 592–598. [Google Scholar] [CrossRef]
- Kanno, S.; Hirano, S.; Kayama, F. Effects of the phytoestrogen coumestrol on RANK-ligand-induced differentiation of osteoclasts. Toxicology 2004, 203, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Loo, W.T.Y.; Wang, M.; Jin, L.J.; Cheung, M.N.B.; Li, G.R. Association of matrix metalloproteinase (MMP-1, MMP-3 and MMP-9) and cyclooxygenase-2 gene polymorphisms and their proteins with chronic periodontitis. Arch. Oral Biol. 2011, 56, 1081–1090. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.; Campos, M.; Souza, A.; Trevilatto, P.; Line, S. Analysis of MMP-1 and MMP-9 promoter polymorphisms in early osseointegrated implant failure. Int. J. Oral Maxillofac. Implant. 2004, 19, 38–43. [Google Scholar]
- Leite, M.F.; Santos, M.C.; de Souza, A.P.; Line, S.R. Osseintegrated implant failure associeted with MMP-1 promoter polymorphisms (-1607 and -519). Int. J. Oral Maxillofac. Implant. 2008, 23, 653–658. [Google Scholar]
- Costa-Junior, F.R.; Alvim-Pereira, C.C.; Alvim-Pereira, F.; Trevilatto, P.C.; de Souza, A.P.; Santos, M.C. Influence of MMP-8 promoter polymorphism in early osseointegrated implant failure. Clin. Oral Investig. 2013, 17, 311–316. [Google Scholar] [CrossRef]
- Sapna, G.; Gokul, S.; Bagri-Manjrekar, K. Matrix metalloproteinases and periodontal diseases. Oral Dis. 2014, 20, 538–550. [Google Scholar] [CrossRef]
- Tu, C.; Ortega-Cava, C.F.; Chen, G.; Fernandes, N.D.; Cavallo-Medved, D.; Sloane, B.F.; Band, V.; Band, H. Lysosomal cathepsin B participates in the podosome-mediated extracellular matrix degradation and invasion via secreted lysosomes in v-Src fibroblasts. Cancer Res. 2008, 68, 9147–9156. [Google Scholar] [CrossRef]
- Li, X.; Wu, J. Recent developments in patent anti-cancer agents targeting the matrix metalloproteinases (MMPs). Recent Pat. Anticancer Drug Discov. 2010, 5, 109–141. [Google Scholar] [CrossRef]
- Georgiadis, D.; Yiotakis, A. Specific targeting of metzincin family members with small-molecule inhibitors: Progress toward a multifarious challenge. Bioorgan. Med. Chem. 2008, 16, 8781–8794. [Google Scholar] [CrossRef] [PubMed]
- Reiss, K.; Saftig, P. The “A Disintegrin And Metalloprotease” (ADAM) family of sheddases: Physiological and cellular functions. Semin. Cell Dev. Biol. 2009, 20, 126–137. [Google Scholar] [CrossRef] [PubMed]
- Tallant, C.; Marrero, A.; Gomis-Rüth, F.X. Matrix metalloproteinases: Fold and function of their catalytic domains. Biochim. Biophys. Acta. Mol. Cell Res. 2010, 1803, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Golub, L.; Goodson, J.; Lee, H.; Vidal, A.; McNamara, T.; Ramamurthy, N.S. Tetracyclines Inhibit Tissue Collagenases: Effects of Ingested Low-Dose and Local Delivery Systems. J. Periodontol. 1985, 56, 93–97. [Google Scholar] [CrossRef]
- Steinsvoll, S. Periodontal disease, matrix metalloproteinases and chemically modified tetracyclines. Microb. Ecol. Health Dis. 2004, 16, 1–7. [Google Scholar]
- Golub, L.; Lee, H.; Ryan, M.; Giannobile, W.; Payne, P.; Sorsa, T. Tetracyclines inhibit connective tissue breakdown by multiple non-antimicrobial mechanisms. Adv. Dent. Res. 1998, 12, 12–26. [Google Scholar] [CrossRef]
- Ramamurthy, N.; Vernillo, A.; Greenwald, R.; Lee, H.M.; Sorsa, T.; Golub, L.M.; Rifkin, B.R. Reactive oxygen species activate and tetracyclines inhibit rat osteoblast collagenase. J. Bone Miner. Res. 1993, 8, 1247–1253. [Google Scholar] [CrossRef]
- Uitto, V.; Firth, J.; Nip, L.; Golub, L. Doxycycline and chemically modified tetracyclines inhibit gelatinase A (MMP-2) gene expression in human skin keratinocytes. Ann. N. Y. Acad. Sci. 1994, 6, 140–151. [Google Scholar] [CrossRef]
- Hanemaaijer, R.; Visser, H.; Koolwijk, P.; Sorsa, T.; Salo, T.; Golub, L.M.; van Hinsbergh, V.W. Inhibition of MMP synthesis by doxycycline and chemically modified tetracyclines (CMTs) in human endothelial cells. Adv. Dent. Res. 1998, 12, 114–118. [Google Scholar] [CrossRef]
- Sadowski, T.; Steinmeyer, J. Effects of tetracyclines on the production of matrix metalloproteinases and plasminogen activators as well as of their natural inhibitors, tissue inhibitor of metalloproteinases-1 and plasminogen activator inhibitor-1. Inflamm. Res. 2001, 50, 175–182. [Google Scholar] [CrossRef]
- Golub, L.; McNamara, T.; D’Angelo, G.; Greenwald, R.; Ramamurthy, N. A non-antibacterial chemically-modified tetracycline inhibits mammalian collagenase activity. J. Dent. Res. 1987, 66, 1310–1314. [Google Scholar] [CrossRef] [PubMed]
- Greenwald, R.; Golub, L.; Ramamurthy, N.; Chowdhury, M.; Moak, S.; Sorsa, T. In vitro sensitivity of the three mammalian collagenases to tetracycline inhibition: Relationship to bone and cartilage degradation. Bone 1998, 22, 33–38. [Google Scholar] [CrossRef]
- Sorsa, T.; Ramamurthy, N.; Vernillo, A.; Zhang, X.; Konttinen, Y.T.; Rifkin, B.R.; Golub, L.M. Functional sites of chemically modified tetracyclines: Inhibition of the oxidative activation of human neutrophil and chicken osteoclast pro-matrix metalloproteinases. J. Rheumatol. 1998, 25, 975–982. [Google Scholar] [PubMed]
- Collins, J.R.; Olsen, J.; Cuesta, A.; Silva-Vetri, M.; Hernández, M.; Romanos, G.; Rajendra Santosh, A.B.; Palma, P. In vitro microbiological analysis on antibacterial, anti-inflammatory, and inhibitory action on matrix metalloproteinases-8 of commercially available chlorhexidine digluconate mouth rinses. Indian J. Dent. Res. 2018, 29, 799–807. [Google Scholar] [CrossRef]
- Gendron, R.; Grenier, D.; Sorsa, T.; Mayrand, D. Inhibition of the activities of matrix metalloproteinases 2, 8, and 9 by chlorhexidine. Clin. Diagn. Lab. Immunol. 1999, 6, 437–439. [Google Scholar] [CrossRef] [PubMed]
- Scaffa, P.M.C.; Vidal, C.M.P.; Barros, N.; Gesteira, T.F.; Carmona, A.K.; Breschi, L.; Pashley, D.H.; Tjäderhane, L.; Tersariol, I.L.; Nascimento, F.D.; et al. Chlorhexidine inhibits the activity of dental cysteine cathepsins. J. Dent. Res. 2012, 91, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Pashley, D.; Tay, F.; Yiu, C.; Hashimoto, M.; Breschi, L.; Carvalho, R. Collagen degradation by host-derived enzymes during aging. J. Dent. Res. 2004, 83, 216–221. [Google Scholar] [CrossRef] [PubMed]
- Tjäderhane, L.; Nascimento, F.D.; Breschi, L.; Mazzoni, A.; Tersariol, I.L.S.; Geraldeli, S.; Tezvergil-Mutluay, A.; Carrilho, M.; Carvalho, R.M. Strategies to prevent hydrolytic degradation of the hybrid layer-A review. Dent. Mater. 2014, 29, 999–1011. [Google Scholar] [CrossRef]
- Mazzoni, A.; Apolonio, F.M.; Saboia, V.P.A.; Santi, S.; Angeloni, V.; Checchi, V.; Curci, R.; Di Lenarda, R.; Tay, F.R.; Pashley, D.H.; et al. Carbodiimide inactivation of MMPs and effect on dentin bonding. J. Dent. Res. 2014, 93, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Widler, L.; Jaeggi, K.; Glatt, M.; Müller, K.; Bachmann, R.; Bisping, M.; Born, A.R.; Cortesi, R.; Guiglia, G.; Jeker, H.; et al. Highly potent geminal bisphosphonates. From pamidronate disodium (Aredia) to zoledronic acid (Zometa). J. Med. Chem. 2002, 45, 3721–3738. [Google Scholar] [CrossRef]
- Boissier, S.; Ferreras, M.; Peyruchaud, O.; Magnetto, S.; Ebetino, F.H.; Colombel, M.; Delmas, P.; Delaissé, J.M.; Clézardin, P. Bisphosphonates Inhibit Breast and Prostate Carcinoma Cell Invasion, an Early Event in the Formation of Bone Metastases. Cancer Res. 2000, 60, 2949–2954. [Google Scholar]
- Evans, C.; Braidman, I. Effects of two novel bisphosphonates on bone cells in vitro. J. Bone Miner. Res. 1994, 26, 95–107. [Google Scholar] [CrossRef]
- Heikkilä, P.; Teronen, O.; Moilanen, M.; Konttinen, Y.T.; Hanemaaijer, R.; Laitinen, M.; Maisi, P.; van der Pluijm, G.; Bartlett, J.D.; Salo, T.; et al. Bisphosphonates inhibit stromelysin-1 (MMP-3), matrix metalloelastase (MMP-12), collagenase-3 (MMP-13) and enamelysin (MMP-20), but not urokinase-type plasminogen activator, and diminish invasion and migration of human malignant and endothelial cell lines. Anticancer Drugs 2002, 13, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Heikkilä, P.; Teronen, O.; Hirn, M.; Sorsa, T.; Tervahartiala, T.; Salo, T.; Konttinen, Y.T.; Halttunen, T.; Moilanen, M.; Hanemaaijer, R.; et al. Inhibition of matrix metalloproteinase-14 in osteosarcoma cells by clodronate. J. Surg. Res. 2003, 111, 45–52. [Google Scholar] [CrossRef]
- Teronen, O.; Konttinen, Y.; Lindqvist, C.; Salo, T.; Ingman, T.; Lauhio, A.; Ding, Y.; Santavirta, S.; Valleala, H.; Sorsa, T. Inhibition of matrix metalloproteinase-1 by dichloromethylene bisphosphonate (clodronate). Calcif. Tissue Int. 1997, 61, 59–61. [Google Scholar] [CrossRef] [PubMed]
- Akram, Z.; Abduljabbar, T.; Kellesarian, S.V.; Abu Hassan, M.I.; Javed, F.; Vohra, F. Efficacy of Bisphosphonate as an Adjunct to Nonsurgical Periodontal Therapy in the Management of Periodontal Disease: A Systematic Review. Br. J. Clin. Pharmacol. 2017, 83, 444–454. [Google Scholar] [CrossRef]
- Koivunen, E.; Arap, W.; Valtanen, H.; Rainisalo, A.; Medina, O.P.; Heikkilä, P.; Kantor, C.; Gahmberg, C.G.; Salo, T.; Konttinen, Y.T.; et al. Tumor targeting with a selective gelatinase inhibitor. Nat. Biotechnol. 1999, 17, 768–774. [Google Scholar] [CrossRef] [PubMed]


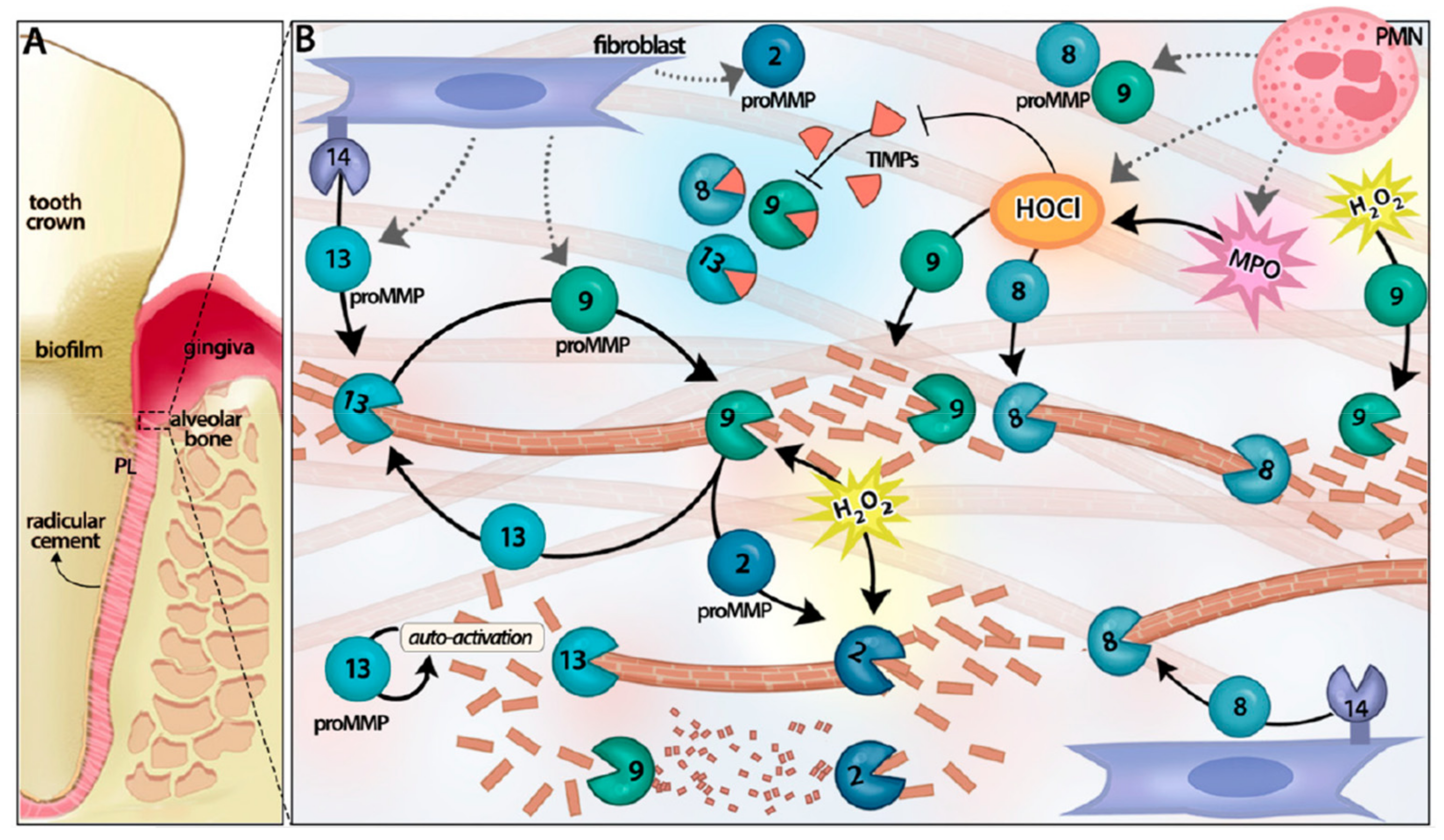{kind=link}
| MMP | Other Names | Group | Substrates in Vitro |
|---|---|---|---|
| MMP-1 | Collagenase-1, Interstitial collagenase, Fibroblast collagenase | Collagenases | Type I, II, III, VII, VIII, X, XI collagens, gelatin, fibronectin, laminin, tenascin, a2-macroglobulin, IL-1b, pro-TNF-a, pro-MMP-1, -2, -9 |
| MMP-2 | Gelatinase A, 72 kDa gelatinase/type IV collagenase | Gelatinases | Type I, II, III, IV, V, VII, X, XI collagens, gelatin, laminin, elastin, fibronectin, a2-macroglobulin, IL-1b, pro-TNF-a, latent TGF-b, pro-MMP-1, -2, -9, -13 |
| MMP-3 | Stromelysin-1, Transin, Proteoglycanase, Collagenase activating protein (CAP) | Stromelysins | Type III, IV, V, VII, IX, X, XI collagens, collagen telopeptides, gelatin, elastin, fibronectin, laminin, aggrecan, decorin, perlecan, versican, a2-macroglobulin, IL-1b, pro-TNF-a, fibrinogen, pro-MMP-1, -3, -7, -8, -9, -13 |
| MMP-7 | Matrilysin-1, Putative Metalloprotease (PUMP-1), Matrin | Matrilysins | Type I, IV collagens, gelatin, elastin, fibronectin, laminin, aggrecan, a2-macroglobulin, pro-TNF-a, pro-MMP-1, -2, -7, -9 |
| MMP-8 | Collagenase-2, Neutrophil collagenase | Collagenases | Type I, II, III collagens, aggrecan, fibrinogen, a2-macroglobulin, bradykinin |
| MMP-9 | Gelatinase B, 92 kDa gelatinase/type IV collagenase, type V collagenase | Gelatinases | Type IV, V, XI, XIV collagens, gelatin, elastin, laminin, aggrecan, a2-macroglobulin, IL-1b, pro-TNF-a |
| MMP-10 | Stromelysin-2, Transin-2 | Stromelysins | Type III, IV, V, VII, IX, X, XI collagens, collagen telopeptides, gelatin, elastin, fibronectin, laminin, aggrecan, decorin, perlecan, versican, a2-macroglobulin, IL-1b, pro-TNF-a, fibrinogen, pro-MMP-1, -3, -7, -8, -9, -13 |
| MMP-11 | Stromelysin-3 | Stromelysins | Type IV collagen, gelatin, fibronectin, a2-proteinase inhibitor |
| MMP-12 | Macrophage elastase, Metalloelastase | Other MMPs | Type I, IV, V collagens, elastin, gelatin, fibronectin, laminin, aggrecan, a2-macroglobulin, pro-TNF-a, fibrinogen |
| MMP-13 | Collagenase-3 | Collagenases | Type I, II, III, IV, VI, IX, X, XIV collagens, collagen telopeptides, gelatin, fibronectin, tenascin-C, aggrecan, fibrinogen, a2-macroglobulin, pro-MMP-9 |
| MMP-14 | MT1-MMP | MT-MMPs | Type I, II, III collagens, gelatin, fibronectin, laminin, aggrecan, a2-macroglobulin, pro-TNF-a, fibrinogen, pro-MMP-2, -13, -20 |
| MMP-15 | MT2-MMP | MT-MMPs | Fibronectin, tenascin, laminin, aggrecan, pro-TNF-a, pro-MMP-2 |
| MMP-16 | MT3-MMP | MT-MMPs | Type III collagen, gelatin, fibronectin, laminin, a2-macroglobulin, pro-MMP-2 |
| MMP-17 | MT4-MMP | MT-MMPs | Gelatin, fibrinogen, fibrin, pro-TNF-a |
| MMP-19 | Matrix metalloproteinase RASI-1 | Other MMPs | Type IV collagen, gelatin, laminin, fibronectin, fibrinogen, fibrin |
| MMP-20 | Enamelysin | Other MMPs | Amelogenin, type IV collagen, aggrecan, fibronectin, laminin, tenascin-C |
| MMP-21 | - | Other MMPs | a1-antitrypsin |
| MMP-23 | Cysteine array (CA)-MMP | Other MMPs | Gelatin |
| MMP-24 | MT5-MMP | MT-MMPs | Fibronectin, gelatin, chondroitin sulfate proteoglycan, pro-MMP-2 |
| MMP-25 | MT6-MMP, Leukolysin | MT-MMPs | Type IV collagen, gelatin, fibronectin, fibrinogen, fibrin, pro-MMP-2 |
| MMP-26 | Matrilysin-2 | Matrilysins | Gelatin, fibronectin, a2-macroglobulin, fibrinogen, pro-MMP-9 |
| MMP-27 | - | Other MMPs | - |
| MMP-28 | Epilysin | Other MMPs | Casein |
| Author (Year) | MMP Involved Into Periodontal Tissues Degradation |
|---|---|
| Choi D.H. et al. (2004) [27] | MMP-8, MMP-9 |
| Beklen A. et al. (2006) [28] | MMP-8, MMP-9 |
| Soder B. et al. (2006) [29] | MMP-8, MMP-9 |
| Kumar M.S. et al. (2006) [30] | MMP-8, MMP-9 |
| Marcaccini A.M. et al. (2010) [31] | MMP-1, MMP-13 |
| Kraft-Naumarker M. et al. (2012) [32] | MMP-8 |
| Rai B. et al. (2008) [34] | MMP-8, MMP-9 |
| Xu L. et al. (2008) [35] | MMP-8 |
| Schure R. et al. (2013) [36] | MMP-3, MMP-7, MMP-8 |
| Gursoy U.K. et al. (2010) [37] | MMP-8 |
| Eminigil G. et al. (2006) [39] | MMP-7 |
| Eminigil G. et al. (2006) [40] | MMP-25, MMP-26 |
| Ramseier C.A. et al. (2009) [41] | MMP-8, MMP-9 |
| Hernandez Rios M. et al. (2009) [42] | MMP-9, MMP-13 |
| Alpagot T. et al. (2001) [43] | MMP-3 |
| Marcaccini A.M. et al. (2009) [44] | MMP-3, MMP-8, MMP-9 |
| Kinane D.F. et al. (2003) [45] | MMP-8 |
| Mantyla P. et al. (2006) [46] | MMP-8 |
| Pozo P. et al. (2005) [47] | MMP-8, MMP-9 |
| Mantyla P. et al. (2003) [48] | MMP-8 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Checchi, V.; Maravic, T.; Bellini, P.; Generali, L.; Consolo, U.; Breschi, L.; Mazzoni, A. The Role of Matrix Metalloproteinases in Periodontal Disease. Int. J. Environ. Res. Public Health 2020, 17, 4923. https://doi.org/10.3390/ijerph17144923
Checchi V, Maravic T, Bellini P, Generali L, Consolo U, Breschi L, Mazzoni A. The Role of Matrix Metalloproteinases in Periodontal Disease. International Journal of Environmental Research and Public Health. 2020; 17(14):4923. https://doi.org/10.3390/ijerph17144923
Chicago/Turabian StyleChecchi, Vittorio, Tatjana Maravic, Pierantonio Bellini, Luigi Generali, Ugo Consolo, Lorenzo Breschi, and Annalisa Mazzoni. 2020. "The Role of Matrix Metalloproteinases in Periodontal Disease" International Journal of Environmental Research and Public Health 17, no. 14: 4923. https://doi.org/10.3390/ijerph17144923
APA StyleChecchi, V., Maravic, T., Bellini, P., Generali, L., Consolo, U., Breschi, L., & Mazzoni, A. (2020). The Role of Matrix Metalloproteinases in Periodontal Disease. International Journal of Environmental Research and Public Health, 17(14), 4923. https://doi.org/10.3390/ijerph17144923