Physiological and Psychological Effects of Forest and Urban Sounds Using High-Resolution Sound Sources


 ,
,  ,
, 
 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Auditory Stimulation
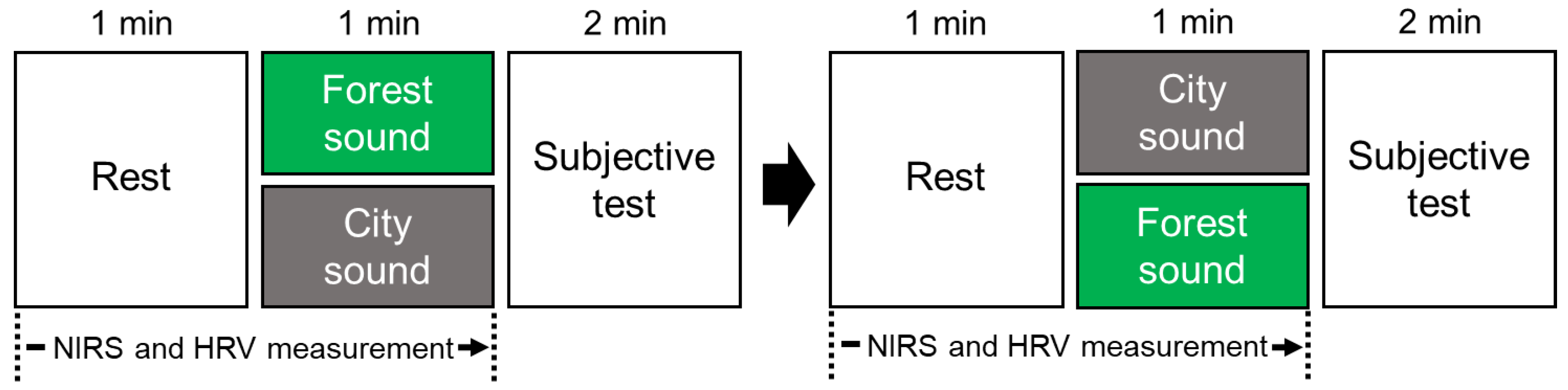
2.3. Study Protocol
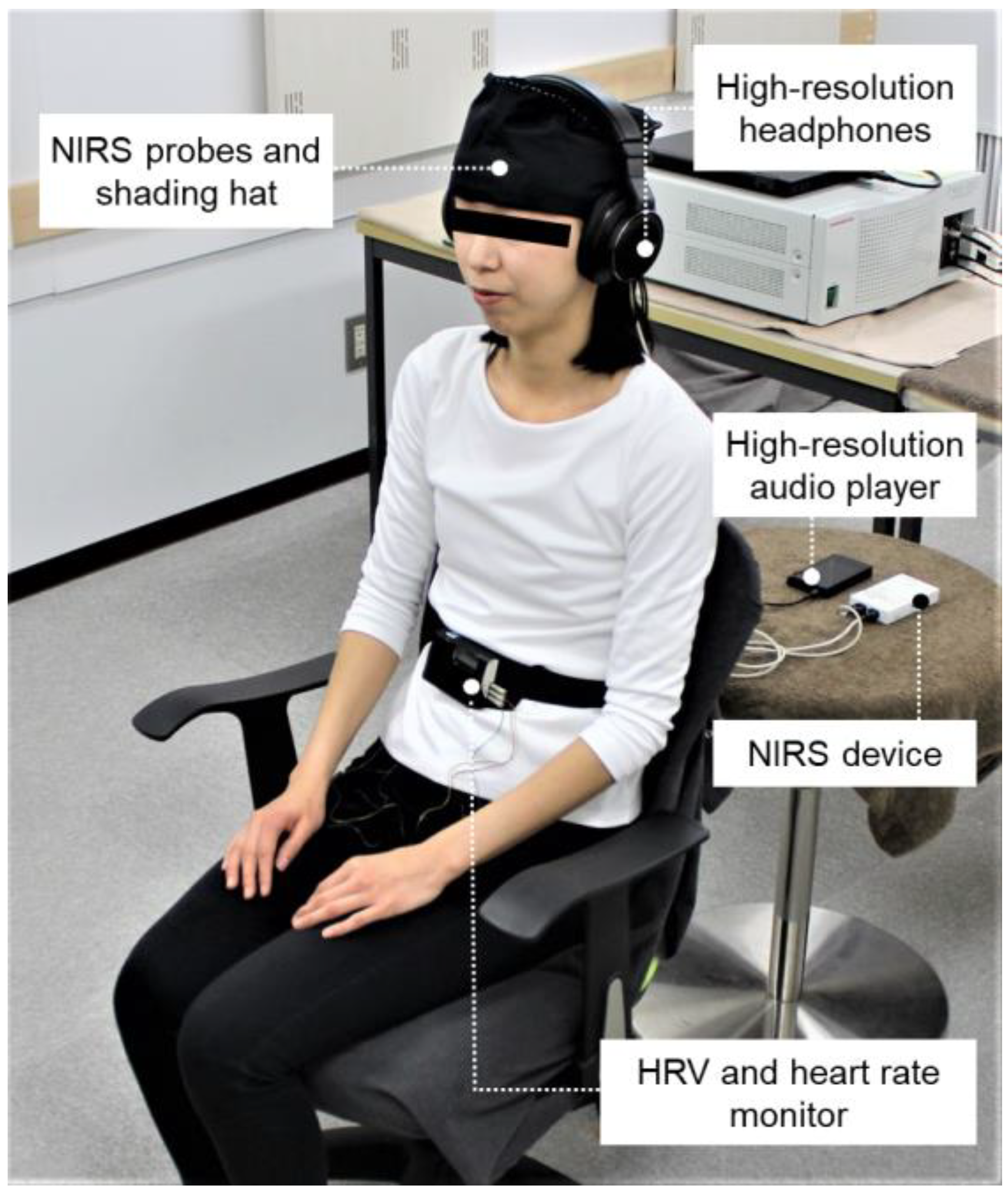
2.4. Physiological Measurements
2.4.1. Near-Infrared Spectroscopy
2.4.2. Heart Rate Variability and Heart Rate
2.5. Psychological Measurements
2.6. Statistical Analysis
3. Results
3.1. Physiological Effects
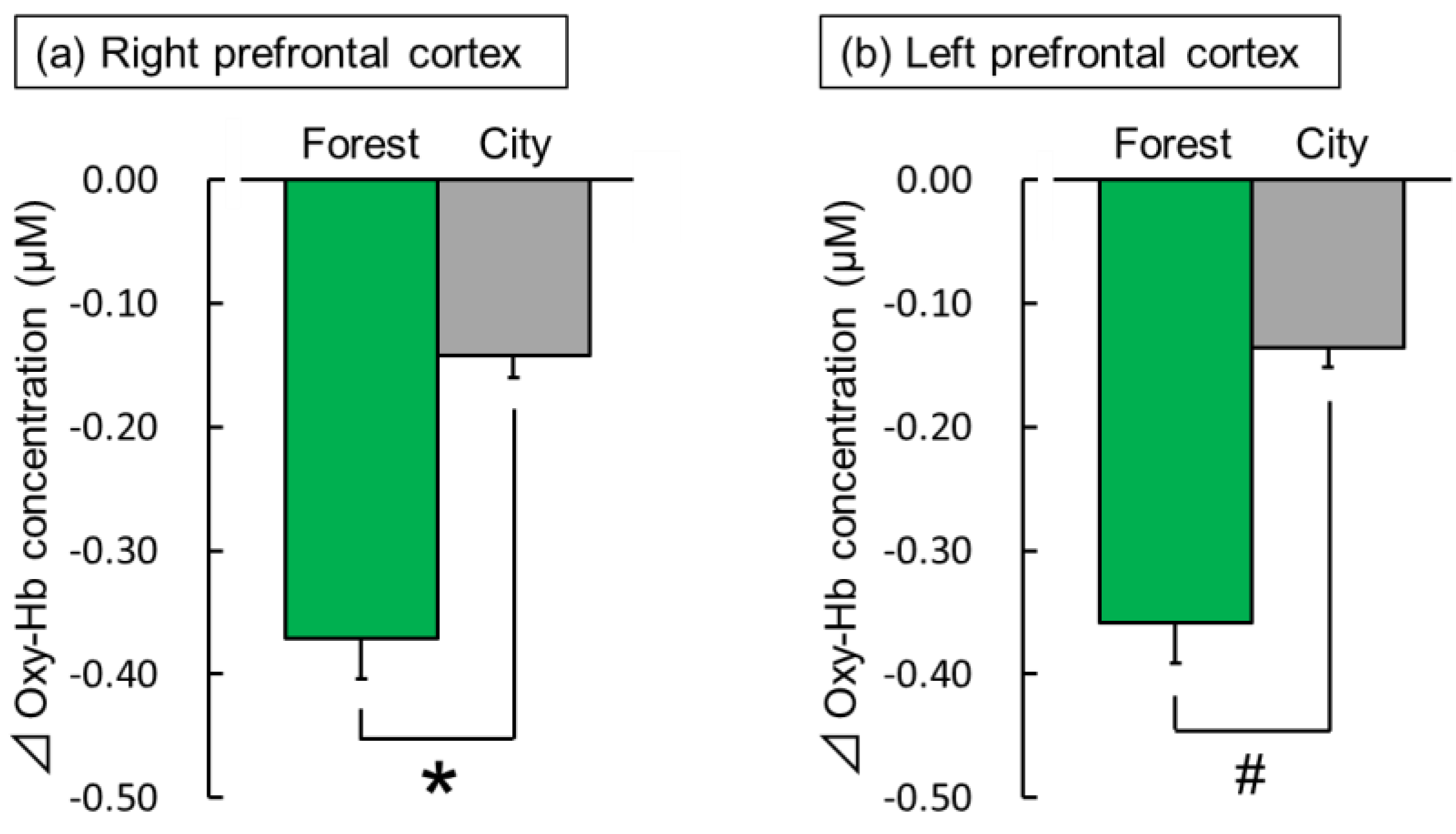
3.1.1. Near-Infrared Spectroscopy (NIRS)
3.1.2. Heart Rate Variability (HRV) and Heart Rate
3.2. Psychological Effects
3.2.1. Modified Semantic Differential (SD) Method
3.2.2. Profile of Mood States (POMS)
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Brunet, M.; Guy, F.; Pilbeam, D.; Mackaye, H.T.; Likius, A.; Ahounta, D.; Beauvilain, A.; Blondel, C.; Bocherens, H.; Boisserie, J.R.; et al. A new hominid from the Upper Miocene of Chad, Central Africa. Nature 2002, 418, 141–151. [Google Scholar] [CrossRef]
- Miyazaki, Y.; Park, B.J.; Lee, J. Nature therapy. In Designing our Future: Local Perspectives on Bioproduction, Ecosystems and Humanity; Osaki, M., Braimoh, A., Nakagami, K., Eds.; United Nations University Press: New York, NY, USA, 2011; pp. 407–412. [Google Scholar]
- United Nations, Department of Economic and Social Affairs, Population Division. World Urbanization Prospects: The 2014 Revision; United Nations: New York, NY, USA, 2014; pp. 1–27. [Google Scholar]
- Song, C.; Ikei, H.; Miyazaki, Y. Physiological effects of nature therapy: A review of the research in Japan. Int. J. Environ. Res. Public Health 2016, 13, 781. [Google Scholar] [CrossRef]
- Miyazaki, Y. Shinrin-Yoku: The Japanese Way of Forest Bathing for Health and Relaxation; Aster: London, UK, 2018. [Google Scholar]
- James, P.; Banay, R.F.; Hart, J.E.; Laden, F. A review of the health benefits of greenness. Curr. Epidemiol. Rep. 2015, 2, 131–142. [Google Scholar] [CrossRef]
- Frumkin, H. Beyond toxicity: Human health and the natural environment. Am. J. Prev. Med. 2001, 20, 234–240. [Google Scholar] [CrossRef]
- Tsunetsugu, Y.; Park, B.J.; Miyazaki, Y. Trends in research related to “Shinrin-yoku” (taking in the forest atmosphere or forest bathing) in Japan. Environ. Health Prev. Med. 2010, 15, 27–37. [Google Scholar] [CrossRef]
- Tsunetsugu, Y.; Park, B.J.; Ishii, H.; Hirano, H.; Kagawa, T.; Miyazaki, Y. Physiological effects of “Shinrin-yoku” (taking in the atmosphere of the forest) in an old-growth broadleaf forest in Yamagata prefecture, Japan. J. Physiol. Anthropol. 2007, 26, 135–142. [Google Scholar] [CrossRef]
- Park, B.J.; Tsunetsugu, Y.; Kasetani, T.; Hirano, H.; Kagawa, T.; Sato, M.; Miyazaki, Y. Physiological effects of Shinrin-yoku (taking in the atmosphere of the forest)—Using salivary cortisol and cerebral activity as indicators. J. Physiol. Anthropol. 2007, 26, 123–128. [Google Scholar] [CrossRef]
- Park, B.J.; Tsunetsugu, Y.; Kasetani, T.; Kagawa, T.; Miyazaki, Y. The physiological effects of Shinrin-yoku (taking in the forest atmosphere or forest bathing): Evidence from field experiments in 24 forests across Japan. Environ. Health Prev. Med. 2010, 15, 18–26. [Google Scholar] [CrossRef]
- Park, B.J.; Tsunetsugu, Y.; Lee, J.; Kagawa, T.; Miyazaki, Y. Effect of the forest environment on physiological relaxation-the results of field tests at 35 sites throughout Japan. In Forest Medicine; Li, Q., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2012; pp. 55–65. [Google Scholar]
- Lee, J.; Park, B.J.; Tsunetsugu, Y.; Ohira, T.; Kagawa, T.; Miyazaki, Y. Effect of forest bathing on physiological and psychological responses in young Japanese male subjects. Public Health 2011, 125, 93–100. [Google Scholar] [CrossRef]
- Lee, J.; Tsunetsugu, Y.; Takayama, N.; Park, B.J.; Li, Q.; Song, C.; Komatsu, M.; Ikei, H.; Tyrväinen, L.; Kagawa, T.; et al. Influence of forest therapy on cardiovascular relaxation in young adults. Evid. Based Complement. Altern. Med. 2014, 2014, 834360. [Google Scholar] [CrossRef]
- Ochiai, H.; Ikei, H.; Song, C.; Kobayashi, M.; Takamatsu, A.; Miura, T.; Kagawa, T.; Li, Q.; Kumeda, S.; Imai, M.; et al. Physiological and psychological effects of forest therapy on middle-age males with high-normal blood pressure. Int. J. Environ. Res. Public Health 2015, 12, 2521–2531. [Google Scholar] [CrossRef]
- Song, C.; Ikei, H.; Kobayashi, M.; Miura, T.; Taue, M.; Kagawa, T.; Li, Q.; Kumeda, S.; Imai, M.; Miyazaki, Y. Effect of forest walking on autonomic nervous system activity in middle-aged hypertensive individuals. Int. J. Environ. Res. Public Health 2015, 12, 2687–2699. [Google Scholar] [CrossRef]
- Nilsson, M.E.; Berglund, B. Soundscape quality in suburban green areas and city parks. Acta Acust. United Acust. 2006, 92, 903–911. [Google Scholar]
- Ward Thompson, C.; Roe, J.; Aspinall, P.; Mitchell, R.; Clow, A.; Miller, D. More green space is linked to less stress in deprived communities: Evidence from salivary cortisol patterns. Landsc. Urban Plan. 2012, 105, 221–229. [Google Scholar] [CrossRef]
- Song, C.; Ikei, H.; Miyazaki, Y. Physiological effects of visual stimulation with forest imagery. Int. J. Environ. Res. Public Health 2018, 15, E213. [Google Scholar] [CrossRef]
- Sakuragawa, S.; Miyazaki, Y.; Kaneko, T.; Makita, T. Influence of wood wall panels on physiological and psychological responses. J. Wood Sci. 2005, 51, 136–140. [Google Scholar] [CrossRef]
- Song, C.; Ikei, H.; Nara, M.; Takayama, D.; Miyazaki, Y. Physiological effects of viewing bonsai in elderly patients undergoing rehabilitation. Int. J. Environ. Res. Public Health 2018, 14, 2635. [Google Scholar] [CrossRef]
- Park, S.A.; Song, C.; Choi, J.Y.; Son, K.C.; Miyazaki, Y. Foliage Plants cause physiological and psychological relaxation as evidenced by measurements of prefrontal cortex activity and profile of mood states. HortScience 2016, 51, 1308–1312. [Google Scholar] [CrossRef]
- Ochiai, H.; Song, C.; Ikei, H.; Imai, M.; Miyazaki, Y. Effects of visual stimulation with bonsai trees on adult male patients with spinal cord injury. Int. J. Environ. Res. Public Health 2017, 14, 1017. [Google Scholar] [CrossRef]
- Ikei, H.; Song, C.; Igarashi, M.; Namekawa, T.; Miyazaki, Y. Physiological and psychological relaxing effects of visual stimulation with foliage plants in high school students. Adv. Hort. Sci. 2014, 28, 111–116. [Google Scholar] [CrossRef]
- Ikei, H.; Komatsu, M.; Song, C.; Himoro, E.; Miyazaki, Y. The physiological and psychological relaxing effects of viewing rose flowers in office workers. J. Physiol. Anthropol. 2014, 33, 6. [Google Scholar] [CrossRef]
- Song, C.; Igarashi, M.; Ikei, H.; Miyazaki, Y. Physiological effects of viewing fresh red roses. Complement. Ther. Med. 2017, 35, 78–84. [Google Scholar] [CrossRef]
- Ulrich, R.S.; Simons, R.F.; Losito, B.D.; Fiorito, E.; Miles, M.A.; Zelson, M. Stress recovery during exposure to natural and urban environments. J. Environ. Psychol. 1991, 11, 201–230. [Google Scholar] [CrossRef]
- de Kort, Y.A.W.; Meijnders, A.L.; Sponselee, A.A.G.; IJsselsteijn, W.A. What’s wrong with virtual trees? Restoring from stress in a mediated environment. J. Environ. Psychol. 2006, 26, 309–320. [Google Scholar] [CrossRef]
- Jiang, B.; Chang, C.Y.; Sullivan, W.C. A dose of nature: Tree cover, stress reduction, and gender differences. Landsc. Urban Plan. 2014, 132, 26–36. [Google Scholar] [CrossRef]
- Felnhofer, A.; Kothgassner, O.D.; Schmidt, M.; Heinzle, A.K.; Beutl, L.; Hlavacs, H.; Kryspin-Exner, I. Is virtual reality emotionally arousing? Investigating five emotion inducing virtual park scenarios. Int. J. Hum. Comput. Stud. 2015, 82, 48–56. [Google Scholar] [CrossRef]
- Ikei, H.; Song, C.; Miyazaki, Y. Physiological effect of olfactory stimulation by Hinoki cypress (Chamaecyparis obtusa) leaf oil. J. Physiol. Anthropol. 2015, 34, 44. [Google Scholar] [CrossRef]
- Ikei, H.; Song, C.; Miyazaki, Y. Effect of olfactory stimulation by α-pinene on autonomic nervous activity. J. Wood Sci. 2016, 62, 568–572. [Google Scholar] [CrossRef]
- Igarashi, M.; Song, C.; Ikei, H.; Ohira, T.; Miyazaki, Y. Effect of olfactory stimulation by fresh rose flowers on autonomic nervous activity. J. Altern. Complement. Med. 2014, 20, 727–731. [Google Scholar] [CrossRef]
- Igarashi, M.; Ikei, H.; Song, C.; Miyazaki, Y. Effects of olfactory stimulation with rose and orange oil on prefrontal cortex activity. Complement. Ther. Med. 2014, 22, 1027–1031. [Google Scholar] [CrossRef]
- Ikei, H.; Song, C.; Miyazaki, Y. Physiological effects of touching hinoki cypress (Chamaecyparis obtusa). J. Wood Sci. 2018, 64, 226–236. [Google Scholar] [CrossRef]
- Ikei, H.; Song, C.; Miyazaki, Y. Physiological effects of touching wood. Int. J. Environ. Res. Public Health 2017, 14, 801. [Google Scholar] [CrossRef]
- Ikei, H.; Song, C.; Miyazaki, Y. Physiological effects of touching coated wood. Int. J. Environ. Res. Public Health 2017, 14, 773. [Google Scholar] [CrossRef]
- Alvarsson, J.J.; Wiens, S.; Nilsson, M.E. Stress recovery during exposure to nature sound and environmental noise. Int. J. Environ. Res. Public Health 2010, 7, 1036–1046. [Google Scholar] [CrossRef]
- Annerstedt, M.; Jönsson, P.; Wallergård, M.; Johansson, G.; Karlson, B.; Grahn, P.; Hansen, A.M.; Währborg, P. Inducing physiological stress recovery with sounds of nature in a virtual reality forest—Results from a pilot study. Physiol. Behav. 2013, 118, 240–250. [Google Scholar] [CrossRef]
- Oohashi, T.; Nishina, E.; Honda, M.; Yonekura, Y.; Fuwamoto, Y.; Kawai, N.; Maekawa, T.; Nakamura, S.; Fukuyama, H.; Shibasaki, H. Inaudible high-frequency sounds affect brain activity: Hypersonic effect. J. Neurophysiol. 2000, 83, 3548–3558. [Google Scholar] [CrossRef]
- Fox, P.T.; Raichle, M.E. Focal physiological uncoupling of cerebral blood flow and oxidative metabolism during somatosensory stimulation in human subjects. Proc. Natl. Acad. Sci. USA 1986, 83, 1140–1144. [Google Scholar] [CrossRef]
- Hoshi, Y.; Kobayashi, N.; Tamura, M. Interpretation of near infrared spectroscopy signals: A study with a newly developed perfused rat brain model. J. Appl. Physiol. 2001, 90, 1657–1662. [Google Scholar] [CrossRef]
- Watanabe, T.; Mizuno, T.; Shikayama, T.; Miwa, M. Development of a wireless near infrared tissue oxygen monitor system with high sampling rate. In Digital Holography and Three-Dimensional Imaging; Optical Society of America: Miami, FL, USA, 2012. [Google Scholar]
- Task Force of the European Society of Cardiology and the North American Society of Pacing and Electrophysiology. Heart rate variability: Standards of measurement, physiological interpretation and clinical use. Circulation 1996, 93, 1043–1065. [Google Scholar] [CrossRef]
- Kobayashi, H.; Ishibashi, K.; Noguchi, H. Heart rate variability; an index for monitoring and analyzing human autonomic activities. Appl. Hum. Sci. 1999, 18, 53–59. [Google Scholar] [CrossRef]
- Kanaya, N.; Hirata, N.; Kurosawa, S.; Nakayama, M.; Namiki, A. Differential effects of propofol and sevoflurane on heart rate variability. Anesthesiology 2003, 98, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Sawada, Y.; Ohtomo, N.; Tanaka, Y.; Tanaka, G.; Yamakoshi, K.; Terachi, S.; Shimamoto, K.; Nakagawa, M.; Satoh, S.; Kuroda, S.; et al. New technique for time series analysis combining the maximum entropy method and non-linear least squares method: Its value in heart rate variability analysis. Med. Biol. Eng. Comput. 1997, 35, 318–322. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, H.; Park, B.J.; Miyazaki, Y. Normative references of heart rate variability and salivary alpha-amylase in a healthy young male population. J. Physiol. Anthropol. 2012, 31, 9. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, A.; Kratky, K.W. Estimation of breathing rate from respiratory sinus arrhythmia: Comparison of various methods. Ann. Biomed. Eng. 2008, 36, 476–485. [Google Scholar] [CrossRef] [PubMed]
- McCrady, J.D.; Vallbona, C.; Hoff, H.E. Neural origin of the respiratory-heart rate response. Am. J. Physiol. 1966, 211, 323–328. [Google Scholar] [CrossRef]
- Kobayashi, H. Normalization of respiratory sinus arrhythmia by factoring in tidal volume. Appl. Hum. Sci. 1998, 17, 207–213. [Google Scholar] [CrossRef]
- Osgood, C.E.; Suci, G.J.; Tannenbaum, P. The Measurement of Meaning; University of Illinois Press: Urbana, IL, USA, 1957. [Google Scholar]
- Mcnair, D.M.; Lorr, M. An analysis of mood in neurotics. J. Abnorm. Psychol. 1964, 69, 620–627. [Google Scholar] [CrossRef]
- McNair, D.M.; Lorr, M.; Droppleman, L.F. Profile of Mood States Manual; Educational and Industrial Testing Services: San Diego, CA, USA, 1992. [Google Scholar]
- Yokoyama, K. POMS Shortened Version—Manual and Commentary on Cases; Kaneko Syoboh: Tokyo, Japan, 2005. (In Japanese) [Google Scholar]
- Igarashi, M.; Aga, M.; Ikei, H.; Namekawa, T.; Miyazaki, T. Physiological and psychological effects on high school students of viewing real and artificial pansies. Int. J. Environ. Res. Public Health 2015, 12, 2521–2531. [Google Scholar] [CrossRef]
- Igarashi, M.; Yamamoto, T.; Lee, J.; Song, C.; Ikei, H.; Miyazaki, Y. Effects of stimulation by three-dimensional natural images on prefrontal cortex and autonomic nerve activity: A comparison with stimulation using two-dimensional images. Cogn. Process. 2014, 15, 551–556. [Google Scholar] [CrossRef]
- Ghezeljeh, T.N.; Nasari, M.; Haghani, H.; Loieh, H.R. The effect of nature sounds on physiological indicators among patients in the cardiac care unit. Complement. Ther. Clin. Pract. 2017, 29, 147–152. [Google Scholar] [CrossRef]
- Goldstein, D.S.; Bentho, O.; Park, M.Y.; Sharabi, Y. Low-frequency power of heart rate variability is not a measure of cardiac sympathetic tone but may be a measure of modulation of cardiac autonomic outflows by baroreflexes. Exp. Physiol. 2011, 96, 1255–1261. [Google Scholar] [CrossRef]
- Heathers, J.A. Everything Hertz: Methodological issues in short-term frequency-domain HRV. Front. Physiol. 2014, 5, 177. [Google Scholar] [CrossRef] [PubMed]
- Pagani, M.; Lucini, D.; Porta, A. Sympathovagal balance from heart rate variability: Time for a second round? Exp. Physiol. 2012, 97, 1141–1142. [Google Scholar] [CrossRef] [PubMed]
- Jo, H.; Rodiek, S.; Fujii, E.; Miayazaki, Y.; Park, B.J.; Ann, S.W. Physiological and psychological response of floral scent. Am. Soc. Hortsci. 2013, 48, 82–88. [Google Scholar] [CrossRef]
- Lee, W.P.; Wu, P.Y.; Lee, M.Y.; Ho, L.H.; Shih, W.M. Music listening alleviates anxiety and physiological responses inpatients receiving spinal anesthesia. Complement. Ther. Med. 2017, 31, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Magee, W.L.; Davidson, J.W. The effect of music therapy on mood states in neurological patients: A pilot study. J. Music Ther. 2002, 1, 20–29. [Google Scholar] [CrossRef]
- Ratcliffe, E.; Gatersleben, B.; Sowden, P.T. Bird sounds and their contributions to perceived attention restoration and stress recovery. J. Environ. Psychol. 2013, 36, 221–228. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jo, H.; Song, C.; Ikei, H.; Enomoto, S.; Kobayashi, H.; Miyazaki, Y. Physiological and Psychological Effects of Forest and Urban Sounds Using High-Resolution Sound Sources. Int. J. Environ. Res. Public Health 2019, 16, 2649. https://doi.org/10.3390/ijerph16152649
Jo H, Song C, Ikei H, Enomoto S, Kobayashi H, Miyazaki Y. Physiological and Psychological Effects of Forest and Urban Sounds Using High-Resolution Sound Sources. International Journal of Environmental Research and Public Health. 2019; 16(15):2649. https://doi.org/10.3390/ijerph16152649
Chicago/Turabian StyleJo, Hyunju, Chorong Song, Harumi Ikei, Seiya Enomoto, Hiromitsu Kobayashi, and Yoshifumi Miyazaki. 2019. "Physiological and Psychological Effects of Forest and Urban Sounds Using High-Resolution Sound Sources" International Journal of Environmental Research and Public Health 16, no. 15: 2649. https://doi.org/10.3390/ijerph16152649
APA StyleJo, H., Song, C., Ikei, H., Enomoto, S., Kobayashi, H., & Miyazaki, Y. (2019). Physiological and Psychological Effects of Forest and Urban Sounds Using High-Resolution Sound Sources. International Journal of Environmental Research and Public Health, 16(15), 2649. https://doi.org/10.3390/ijerph16152649