E-cigarette Aerosol Condensate Enhances Metabolism of Benzo(a)pyrene to Genotoxic Products, and Induces CYP1A1 and CYP1B1, Likely by Activation of the Aryl Hydrocarbon Receptor


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Line and Culture Conditions
2.2. E-Cig Aerosol Condensate Preparation
2.3. E-Cigarette Liquid
2.4. Preparation of Tobacco Smoke Extract (TSE)
2.5. Metabolism of BaP by MSK Cells
2.6. Gene Expression
2.7. Analysis of BaP Tetrols
2.8. Analysis of Nicotine
2.9. mRNA Expression
2.10. Western Blot Analysis
3. Results
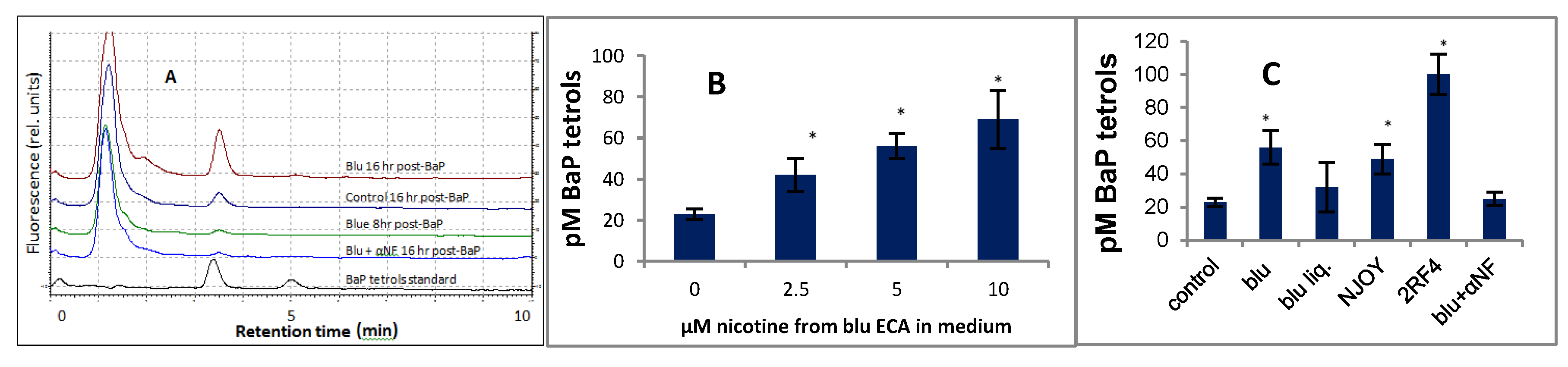
3.1. Effect of α-Napthoflavone on the Metabolism of BaP
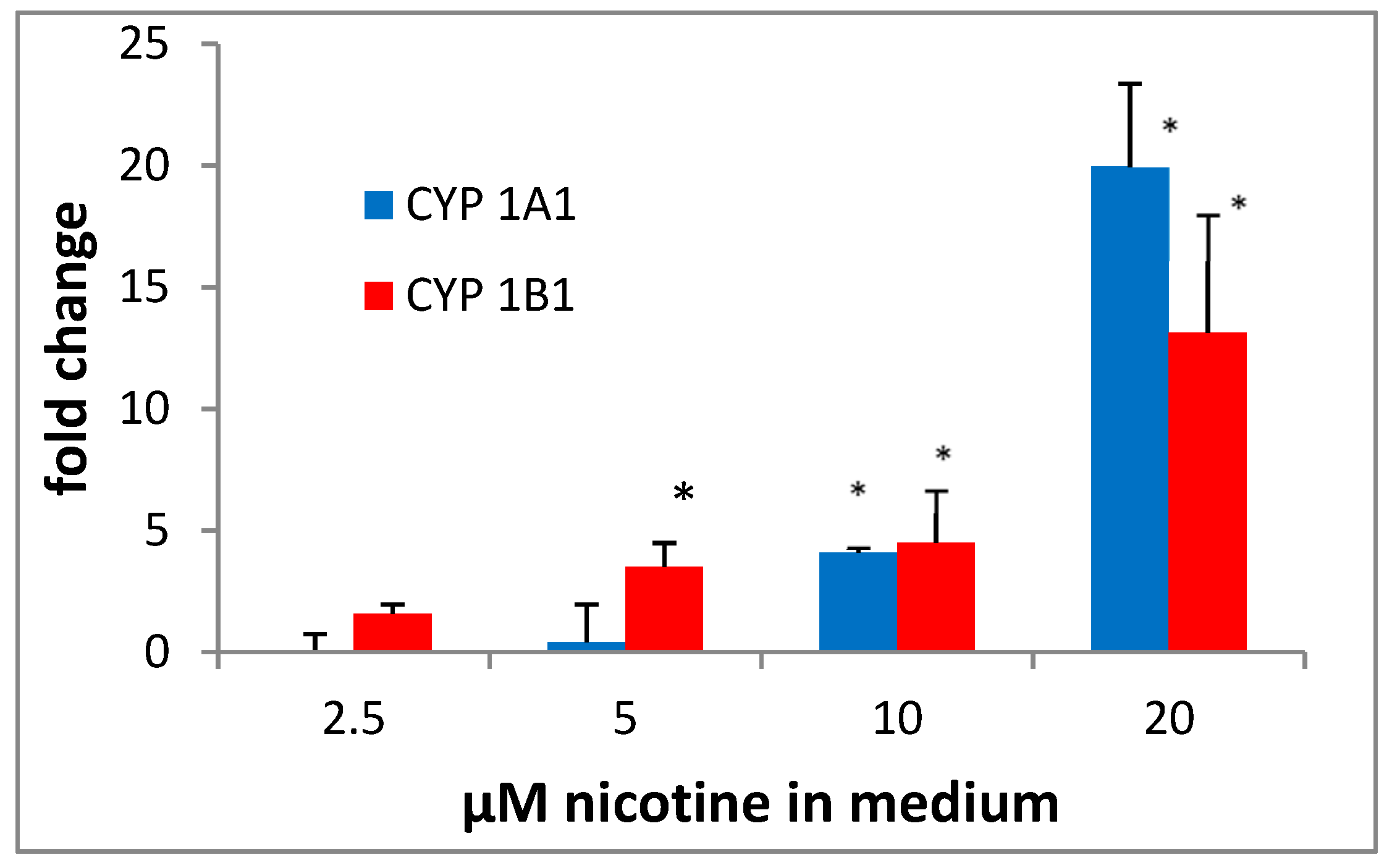
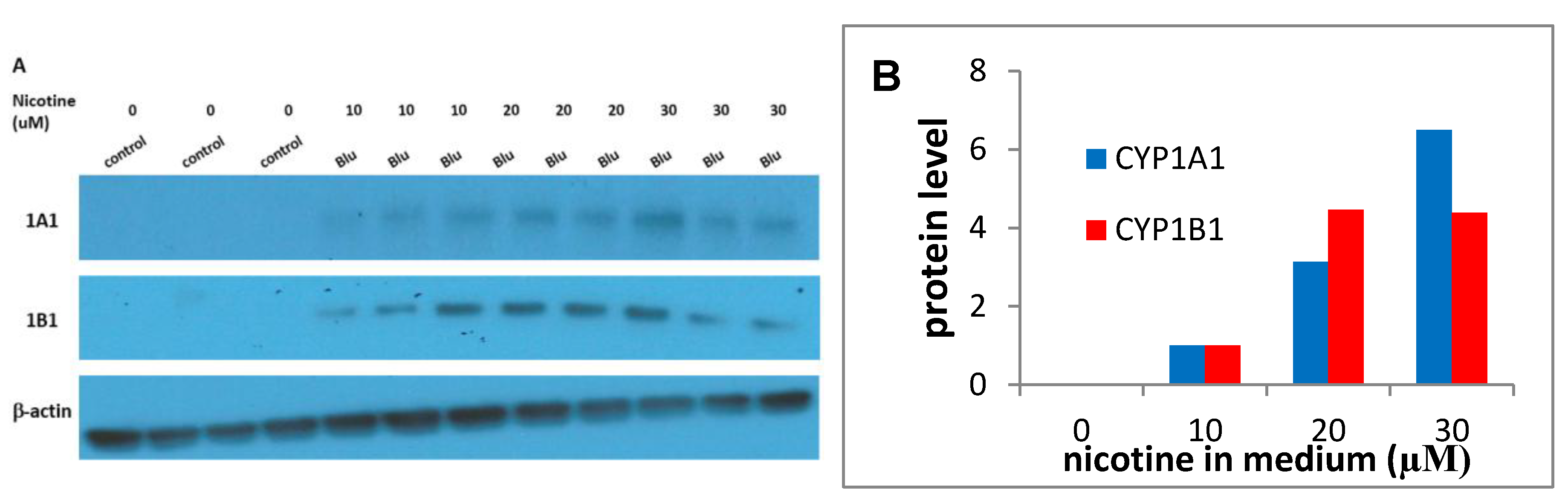
3.2. Cyp1a1 and 1B1
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Fairchild, A.L.; Bayer, R. Public health. Smoke and fire over e-cigarettes. Science 2015, 347, 375–376. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.S.; Homa, D.M.; Wang, T.; Gomez, Y.; Walton, K.; Lu, H.; Neff, L. State-Specific Patterns of Cigarette Smoking, Smokeless Tobacco Use, and E-Cigarette Use among Adults—United States, 2016. Prev. Chronic Dis. 2019, 16, E17. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T. Chemical evaluation of electronic cigarettes. Tob. Control 2014, 23 (Suppl. 2), ii11–ii17. [Google Scholar] [CrossRef]
- Mirbolouk, M.; Charkhchi, P.; Kianoush, S.; Uddin, S.M.I.; Orimoloye, O.A.; Jaber, R.; Bhatnagar, A.; Benjamin, E.J.; Hall, M.E.; DeFilippis, A.P.; et al. Prevalence and Distribution of E-Cigarette Use among U.S. Adults: Behavioral Risk Factor Surveillance System, 2016. Ann. Intern. Med. 2018, 169, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Mirbolouk, M.; Charkhchi, P.; Orimoloye, O.A.; Uddin, S.M.I.; Kianoush, S.; Jaber, R.; Bhatnagar, A.; Benjamin, E.J.; Hall, M.E.; DeFilippis, A.P.; et al. E-Cigarette Use without a History of Combustible Cigarette Smoking among U.S. Adults: Behavioral Risk Factor Surveillance System, 2016. Ann. Intern. Med. 2019, 170, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Gillison, M.L. Current topics in the epidemiology of oral cavity and oropharyngeal cancers. Head Neck 2007, 29, 779–792. [Google Scholar] [CrossRef] [PubMed]
- Salamanca, J.C.; Meehan-Atrash, J.; Vreeke, S.; Escobedo, J.O.; Peyton, D.H.; Strongin, R.M. E-cigarettes can emit formaldehyde at high levels under conditions that have been reported to be non-averse to users. Sci. Rep. 2018, 8, 7559. [Google Scholar] [CrossRef]
- Shahab, L.; Goniewicz, M.L.; Blount, B.C.; Brown, J.; West, R. E-Cigarettes and Toxin Exposure. Ann. Intern. Med. 2017, 167, 525–526. [Google Scholar] [CrossRef]
- Baird, W.M.; Hooven, L.A.; Mahadevan, B. Carcinogenic polycyclic aromatic hydrocarbon-DNA adducts and mechanism of action. Environ. Mol. Mutagenes. 2005, 45, 106–114. [Google Scholar] [CrossRef]
- Xue, W.; Warshawsky, D. Metabolic activation of polycyclic and heterocyclic aromatic hydrocarbons and DNA damage: A review. Toxicol. Appl. Pharmacol. 2005, 206, 73–93. [Google Scholar] [CrossRef]
- Conney, A.H.; Chang, R.L.; Jerina, D.M.; Wei, S.J. Studies on the metabolism of benzo[a]pyrene and dose-dependent differences in the mutagenic profile of its ultimate carcinogenic metabolite. Drug Metab. Rev. 1994, 26, 125–163. [Google Scholar] [CrossRef] [PubMed]
- Sacks, P.G.; Parnes, S.M.; Gallick, G.E.; Mansouri, Z.; Lichtner, R.; Satya-Prakash, K.L.; Pathak, S.; Parsons, D.F. Establishment and characterization of two new squamous cell carcinoma cell lines derived from tumors of the head and neck. Cancer Res. 1988, 48, 2858–2866. [Google Scholar] [PubMed]
- Kochhar, A.; Kopelovich, L.; Sue, E.; Guttenplan, J.B.; Herbert, B.S.; Dannenberg, A.J.; Subbaramaiah, K. p53 modulates Hsp90 ATPase activity and regulates aryl hydrocarbon receptor signaling. Cancer Prev. Res. 2014, 7, 596–606. [Google Scholar] [CrossRef] [PubMed]
- Sacks, P.G.; Zhao, Z.L.; Kosinska, W.; Fleisher, K.E.; Gordon, T.; Guttenplan, J.B. Concentration dependent effects of tobacco particulates from different types of cigarettes on expression of drug metabolizing proteins, and benzo(a)pyrene metabolism in primary normal human oral epithelial cells. Food Chem. Toxicol. 2011, 49, 2348–2355. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Alexandrov, K.; Rojas, M.; Geneste, O.; Castegnaro, M.; Camus, A.M.; Petruzzelli, S.; Giuntini, C.; Bartsch, H. An improved fluorometric assay for dosimetry of benzo(a)pyrene diol-epoxide-DNA adducts in smokers’ lung: Comparisons with total bulky adducts and aryl hydrocarbon hydroxylase activity. Cancer Res. 1992, 52, 6248–6253. [Google Scholar] [PubMed]
- Etter, J.F. A longitudinal study of cotinine in long-term daily users of e-cigarettes. Drug Alcohol Depend. 2016, 160, 218–221. [Google Scholar] [CrossRef] [PubMed]
- Papaseit, E.; Farre, M.; Graziano, S.; Pacifici, R.; Perez-Mana, C.; Garcia-Algar, O.; Pichini, S. Monitoring nicotine intake from e-cigarettes: Measurement of parent drug and metabolites in oral fluid and plasma. Clin. Chem. Lab. Med. 2016, 55, 415–423. [Google Scholar] [CrossRef]
- Nebert, D.W. Aryl hydrocarbon receptor (AHR): “pioneer member” of the basic-helix/loop/helix per-Arnt-sim (bHLH/PAS) family of “sensors” of foreign and endogenous signals. Prog. Lipid Res. 2017, 67, 38–57. [Google Scholar] [CrossRef]
- Nagao, M.; Ushijima, T.; Okonogi, H.; Ochiai, M.; Wakabayashi, K.; Suzuki, T.; Sofuni, T.; Fujita, H.; Nagano, K.; Matsushima, T.; et al. Relationship of DNA Adduct Levels, Cell Proliferation, Mutation Frequencing and Tumor Development in Big BlueR Mouse. Proc. Am. Assoc. Cancer Res. 1995, 36, 159. [Google Scholar]
- Hughes, D.; Guttenplan, J.B.; Marcus, C.B.; Subbaramaiah, K.; Dannenberg, A.J. Heat shock protein 90 inhibitors suppress aryl hydrocarbon receptor-mediated activation of CYP1A1 and CYP1B1 transcription and DNA adduct formation. Cancer Prev. Res. 2008, 1, 485–493. [Google Scholar] [CrossRef]
- Gumus, Z.H.; Du, B.; Kacker, A.; Boyle, J.O.; Bocker, J.M.; Mukherjee, P.; Subbaramaiah, K.; Dannenberg, A.J.; Weinstein, H. Effects of tobacco smoke on gene expression and cellular pathways in a cellular model of oral leukoplakia. Cancer Prev. Res. 2008, 1, 100–111. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Wolkowicz, M.J.; Kotova, T.; Fan, L.; Timko, M.P. Transcriptome sequencing reveals e-cigarette vapor and mainstream-smoke from tobacco cigarettes activate different gene expression profiles in human bronchial epithelial cells. Sci. Rep. 2016, 6, 23984. [Google Scholar] [CrossRef] [PubMed]
- Canistro, D.; Vivarelli, F.; Cirillo, S.; Babot Marquillas, C.; Buschini, A.; Lazzaretti, M.; Marchi, L.; Cardenia, V.; Rodriguez-Estrada, M.T.; Lodovici, M.; et al. E-cigarettes induce toxicological effects that can raise the cancer risk. Sci. Rep. 2017, 7, 2028. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, A.L.; Morse, M.A.; Fields, H.W.; Rothas, D.A.; Pei, P.; Rodrigo, K.A.; Renner, R.J.; Mallery, S.R. Curcumin activates the aryl hydrocarbon receptor yet significantly inhibits (-)-benzo(a)pyrene-7R-trans-7,8-dihydrodiol bioactivation in oral squamous cell carcinoma cells and oral mucosa. Cancer Res. 2002, 62, 5451–5456. [Google Scholar] [PubMed]
- Murray, I.A.; Patterson, A.D.; Perdew, G.H. Aryl hydrocarbon receptor ligands in cancer: Friend and foe. Nat. Rev. Cancer 2014, 14, 801–814. [Google Scholar] [CrossRef] [PubMed]
- Soshilov, A.A.; Denison, M.S. Ligand promiscuity of aryl hydrocarbon receptor agonists and antagonists revealed by site-directed mutagenesis. Mol. Cell. Biol. 2014, 34, 1707–1719. [Google Scholar] [CrossRef] [PubMed]
- Pankow, J.F.; Strongin, R.M.; Peyton, D.H. Formaldehyde from e-cigarettes—It’s not as simple as some suggest. Addiction 2015, 110, 1687–1688. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Erythropel, H.C.; Jabba, S.V.; DeWinter, T.M.; Mendizabal, M.; Anastas, P.T.; Jordt, S.E.; Zimmerman, J.B. Formation of Flavorant-Propylene Glycol Adducts with Novel Toxicological Properties in Chemically Unstable E-Cigarette Liquids. Nicotine Tob. Res. 2018. [Google Scholar] [CrossRef] [PubMed]
- Bitzer, Z.T.; Goel, R.; Reilly, S.M.; Foulds, J.; Muscat, J.; Elias, R.J.; Richie, J.P. Solvent and Temperature Effects on Free Radical Formation in Electronic Cigarette Aerosols. Chem. Res. Toxicol. 2017, 31, 4–12. [Google Scholar] [CrossRef] [PubMed]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.-W.; Kosinska, W.; Guttenplan, J.B. E-cigarette Aerosol Condensate Enhances Metabolism of Benzo(a)pyrene to Genotoxic Products, and Induces CYP1A1 and CYP1B1, Likely by Activation of the Aryl Hydrocarbon Receptor. Int. J. Environ. Res. Public Health 2019, 16, 2468. https://doi.org/10.3390/ijerph16142468
Sun Y-W, Kosinska W, Guttenplan JB. E-cigarette Aerosol Condensate Enhances Metabolism of Benzo(a)pyrene to Genotoxic Products, and Induces CYP1A1 and CYP1B1, Likely by Activation of the Aryl Hydrocarbon Receptor. International Journal of Environmental Research and Public Health. 2019; 16(14):2468. https://doi.org/10.3390/ijerph16142468
Chicago/Turabian StyleSun, Yuan-Wan, Wieslawa Kosinska, and Joseph B. Guttenplan. 2019. "E-cigarette Aerosol Condensate Enhances Metabolism of Benzo(a)pyrene to Genotoxic Products, and Induces CYP1A1 and CYP1B1, Likely by Activation of the Aryl Hydrocarbon Receptor" International Journal of Environmental Research and Public Health 16, no. 14: 2468. https://doi.org/10.3390/ijerph16142468
APA StyleSun, Y.-W., Kosinska, W., & Guttenplan, J. B. (2019). E-cigarette Aerosol Condensate Enhances Metabolism of Benzo(a)pyrene to Genotoxic Products, and Induces CYP1A1 and CYP1B1, Likely by Activation of the Aryl Hydrocarbon Receptor. International Journal of Environmental Research and Public Health, 16(14), 2468. https://doi.org/10.3390/ijerph16142468