
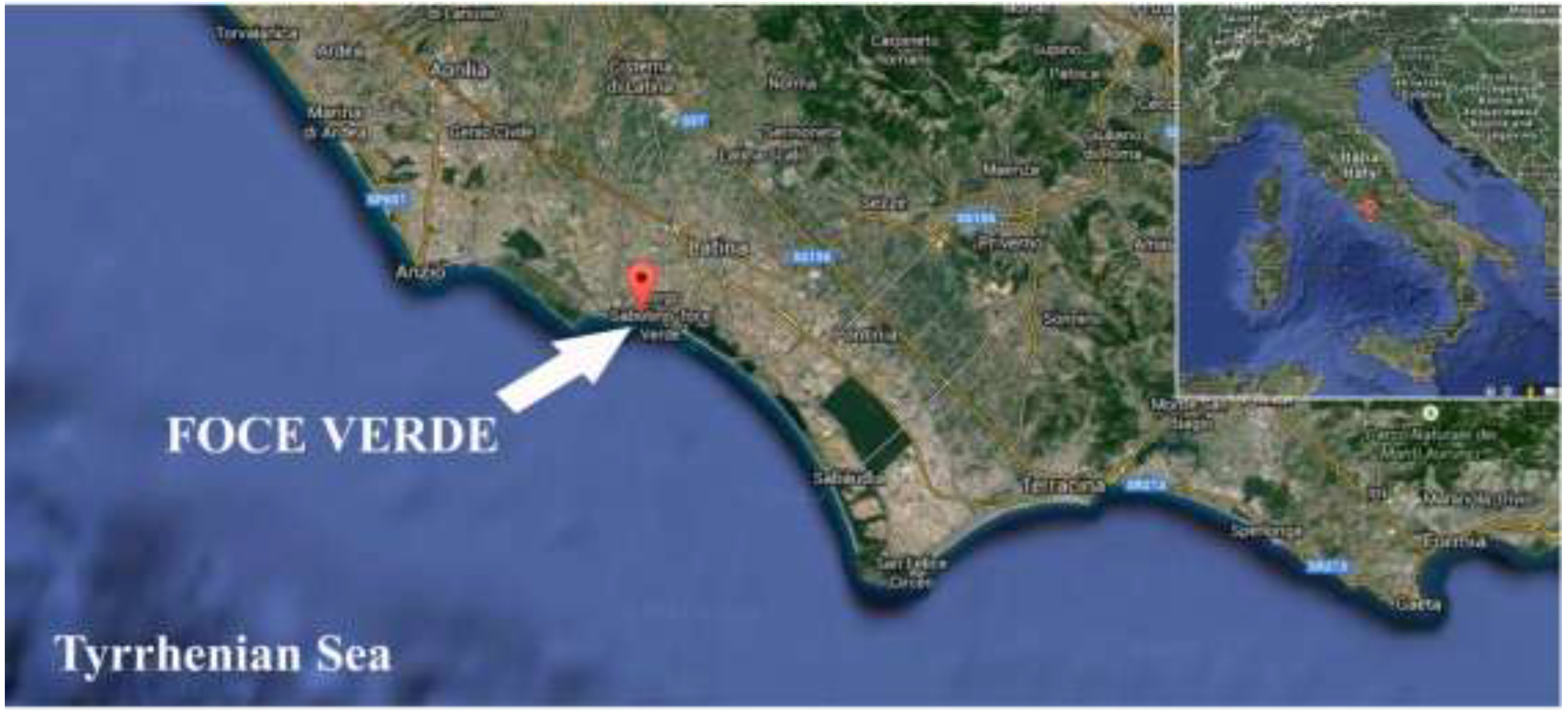
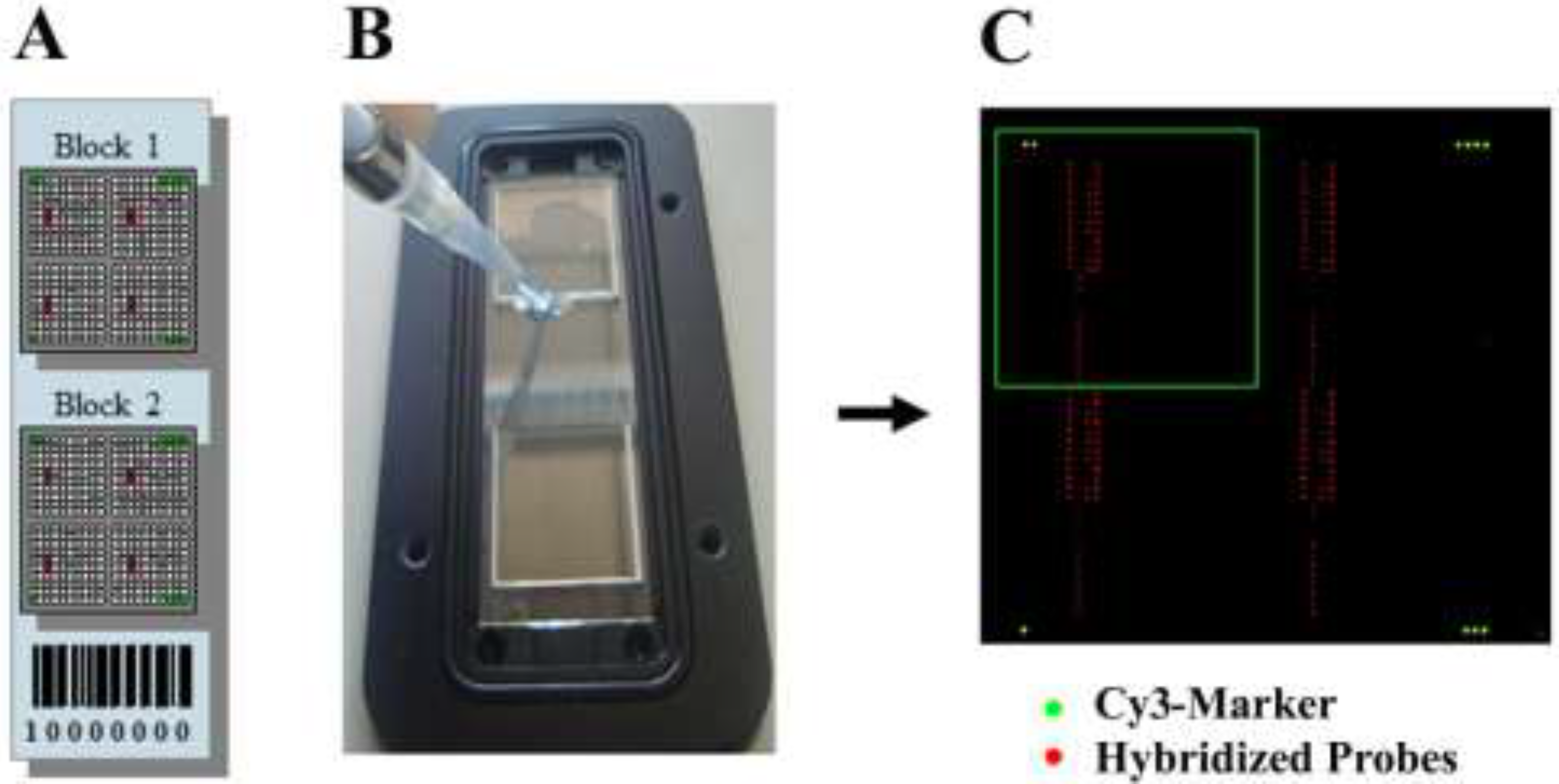
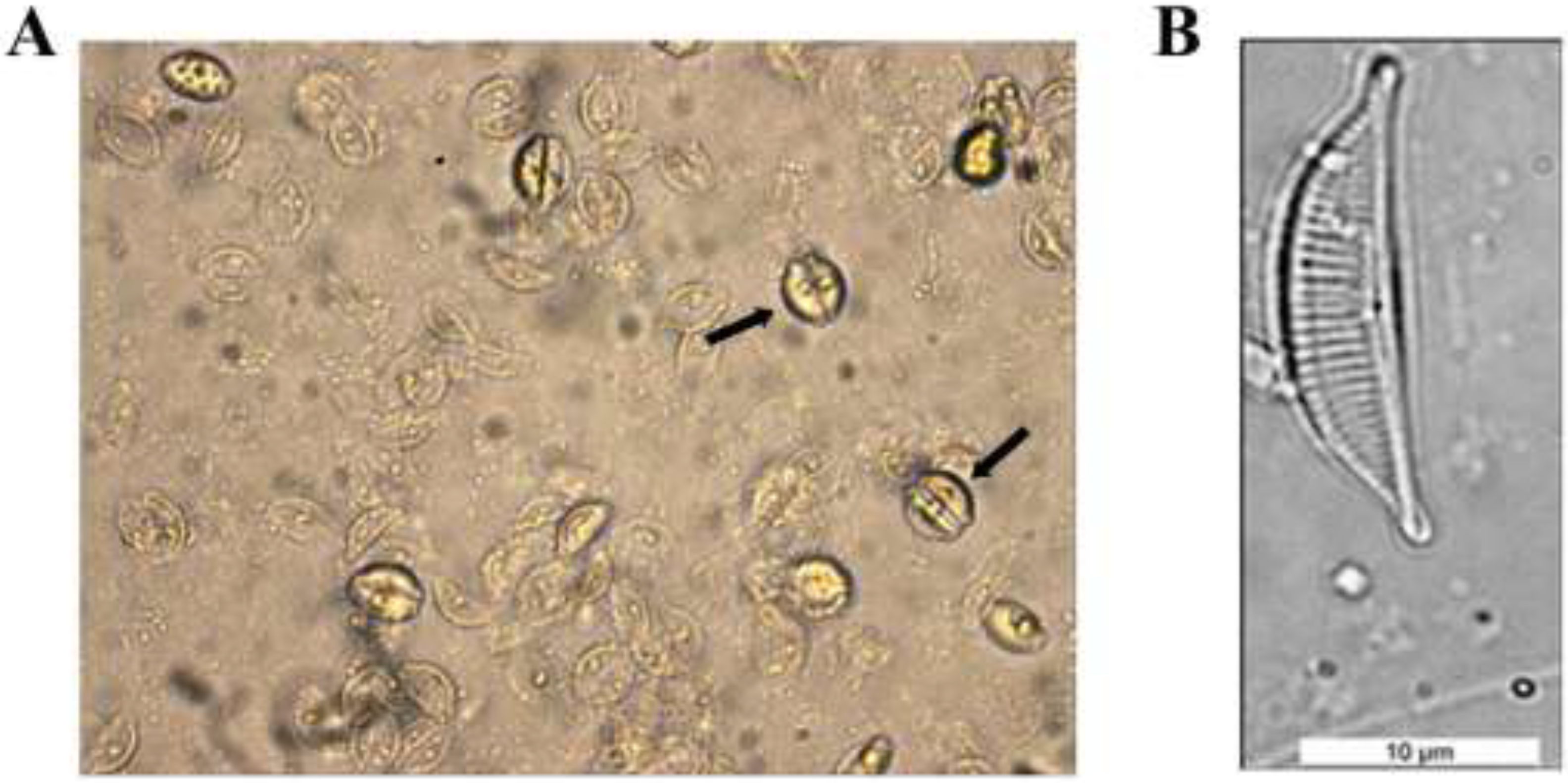
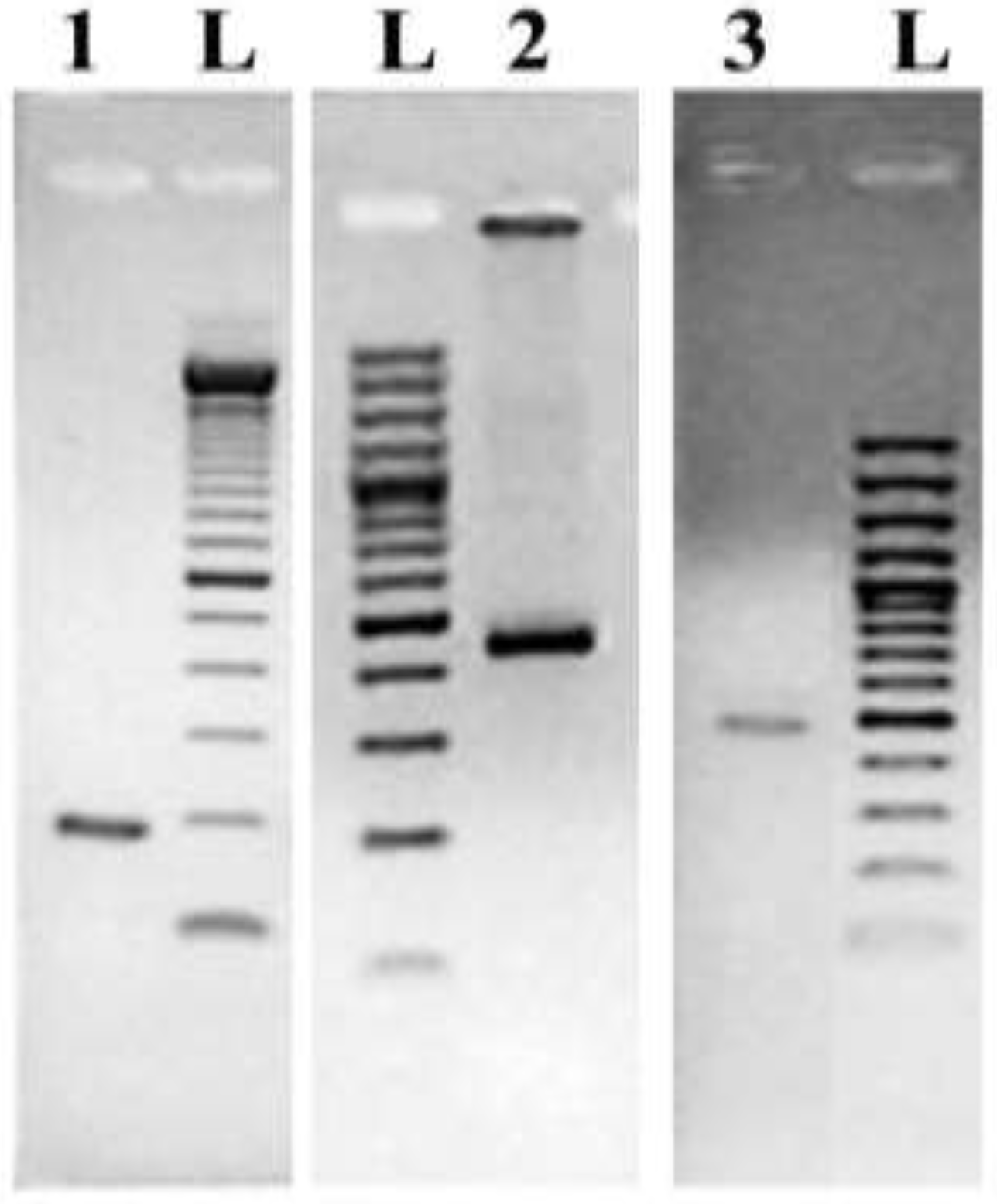
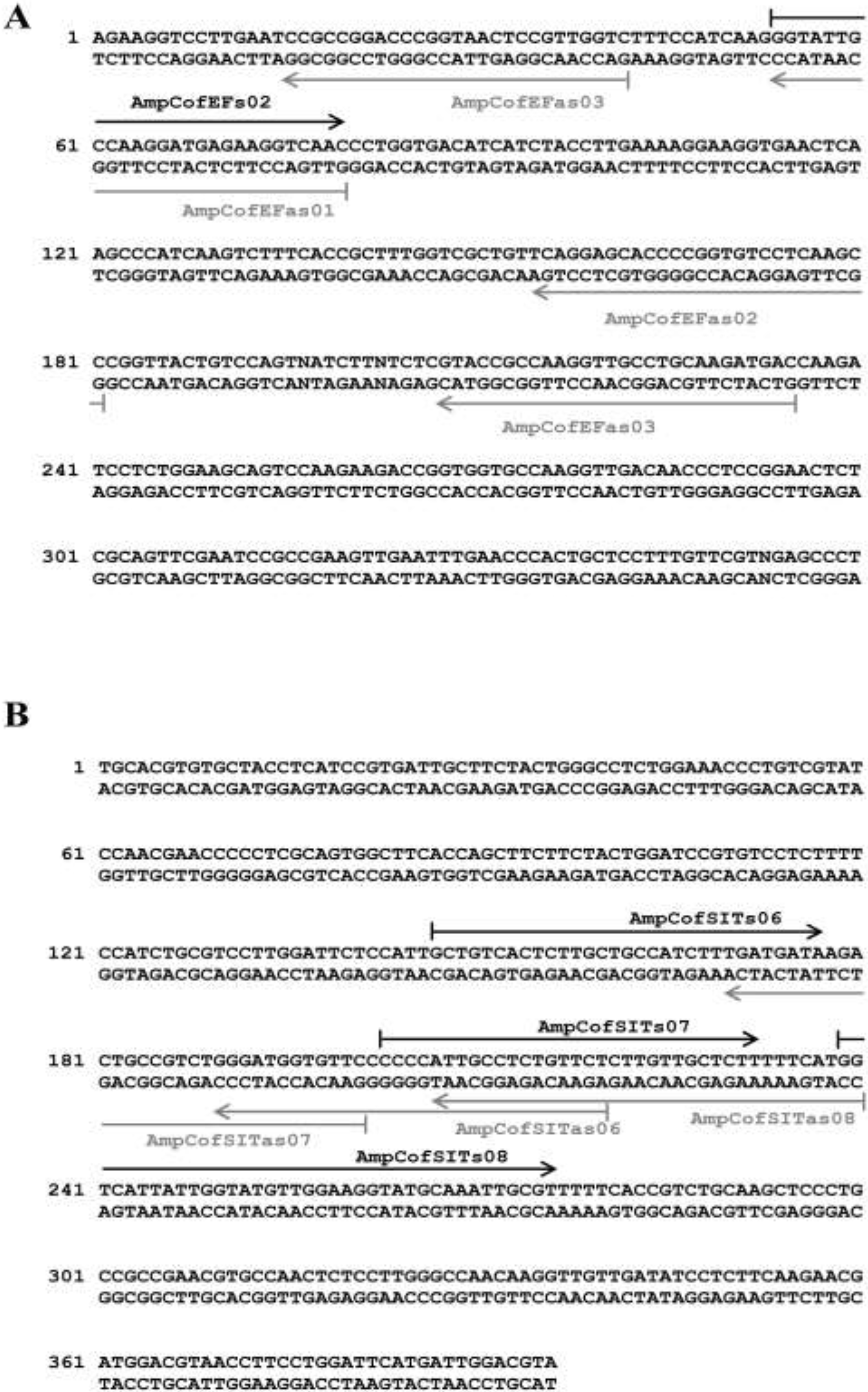
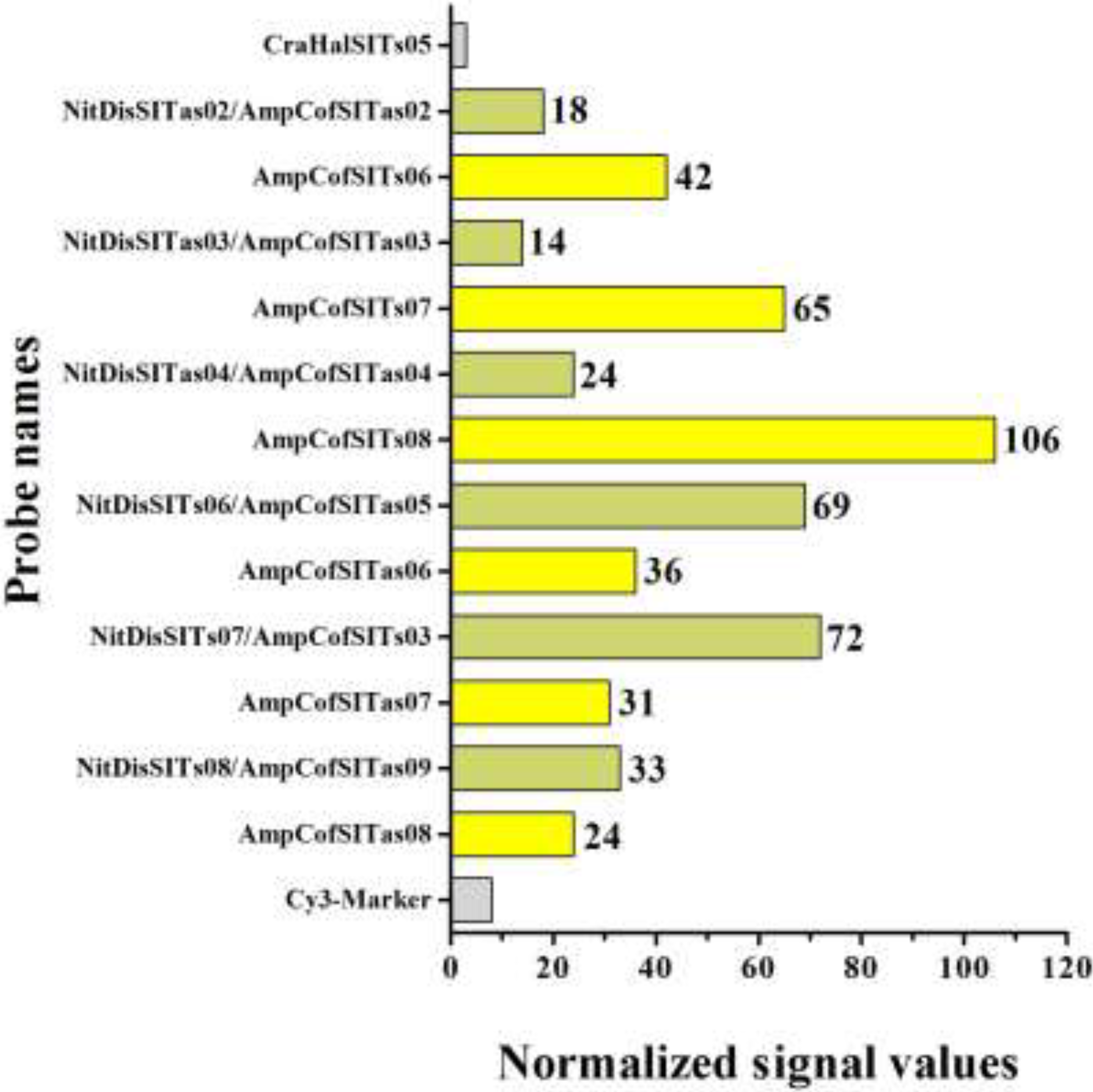
Molecular Detection of a Potentially Toxic Diatom Species


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Share and Cite
Dhar, B.C.; Cimarelli, L.; Singh, K.S.; Brandi, L.; Brandi, A.; Puccinelli, C.; Marcheggiani, S.; Spurio, R. Molecular Detection of a Potentially Toxic Diatom Species. Int. J. Environ. Res. Public Health 2015, 12, 4921-4941. https://doi.org/10.3390/ijerph120504921
Dhar BC, Cimarelli L, Singh KS, Brandi L, Brandi A, Puccinelli C, Marcheggiani S, Spurio R. Molecular Detection of a Potentially Toxic Diatom Species. International Journal of Environmental Research and Public Health. 2015; 12(5):4921-4941. https://doi.org/10.3390/ijerph120504921
Chicago/Turabian StyleDhar, Bidhan Chandra, Lucia Cimarelli, Kumar Saurabh Singh, Letizia Brandi, Anna Brandi, Camilla Puccinelli, Stefania Marcheggiani, and Roberto Spurio. 2015. "Molecular Detection of a Potentially Toxic Diatom Species" International Journal of Environmental Research and Public Health 12, no. 5: 4921-4941. https://doi.org/10.3390/ijerph120504921
APA StyleDhar, B. C., Cimarelli, L., Singh, K. S., Brandi, L., Brandi, A., Puccinelli, C., Marcheggiani, S., & Spurio, R. (2015). Molecular Detection of a Potentially Toxic Diatom Species. International Journal of Environmental Research and Public Health, 12(5), 4921-4941. https://doi.org/10.3390/ijerph120504921