
Indoor Heating Drives Water Bacterial Growth and Community Metabolic Profile Changes in Building Tap Pipes during the Winter Season

Abstract
:1. Introduction
2. Experimental Section
2.1. Sampling Description
2.2. Water Quality Determination
2.3. Flow Cytometry Examination
2.4. Bacterial Community Metabolic Determination

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Carbohydrates | Carboxylic Acids | Amino Acids | Polymers | Phenolic Compounds | Amines |
|---|---|---|---|---|---|
| D,L-α-Glycerol phosphate | Pyruvic acid methyl ester | Arginine | α-Cyclodextrin Glycogen | 4-Hydroxybenzoic acid | Phenylethyl-amine Putrescine |
| β-Methyl-D-glucoside | γ-Hydroxybutyric acid | Threonine | Tween40 | 2-Hydroxybenzoic acid | |
| i-Erythritol | D-Galacturonic acid | Serine | Tween80 | ||
| D-Cellobiose | α-Ketobutyric acid | Phenylalanine | |||
| D-Mannitol | D-Glucosaminic acid | Asparagine | |||
| D-Xylose | D-Malic acid | Glycyl-L-glutamic acid | |||
| Glucose-1-phosphate | Itaconic acid | ||||
| N-Acetyl-D-glucosamine | |||||
| D-Galactonic acid γ-lactone |
2.5. Data Analyses
3. Results and Discussion
3.1. Water Temperature and Quality
| Water Quality Parameters | “Before” Water Samples | “Flushed” Water Samples | T-Text |
|---|---|---|---|
| pH | 7.3 ± 0.02 | 7.2 ± 0.03 | NS |
| Dissolved organic carbon (mg/L) | 1.8 ± 0.1 | 1.9 ± 0.3 | NS |
| UV254 | 0.08 ± 0.0 | 0.09 ± 0.0 | NS |
| Turbidity (NTU) | 0.5 ± 0.0 | 0.4 ± 0.0 | NS |
| Residual chlorine (mg/L) | 0.2 ± 0.0 | 0.3 ± 0.0 | NS |
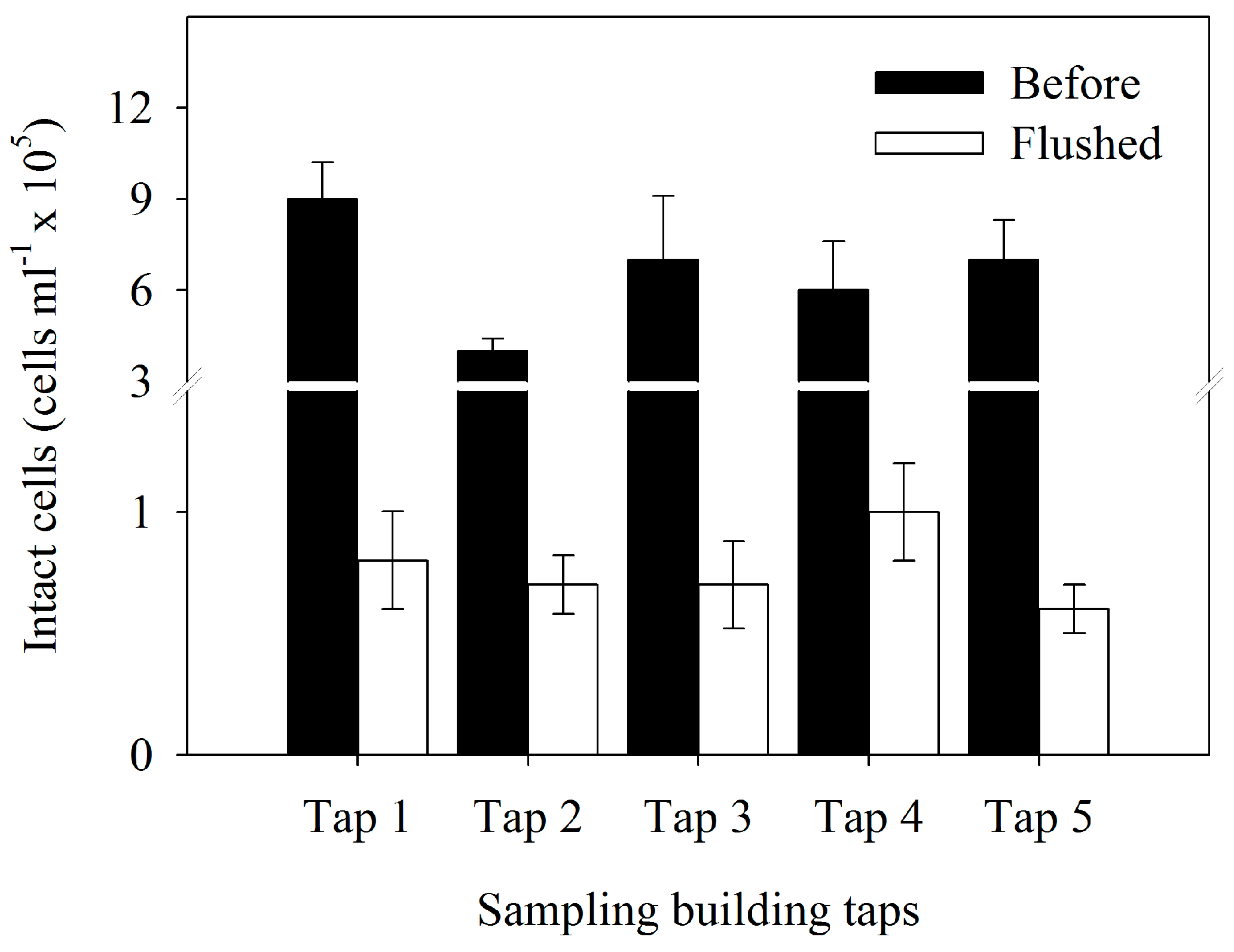
3.2. Bacterial Cell Concentration
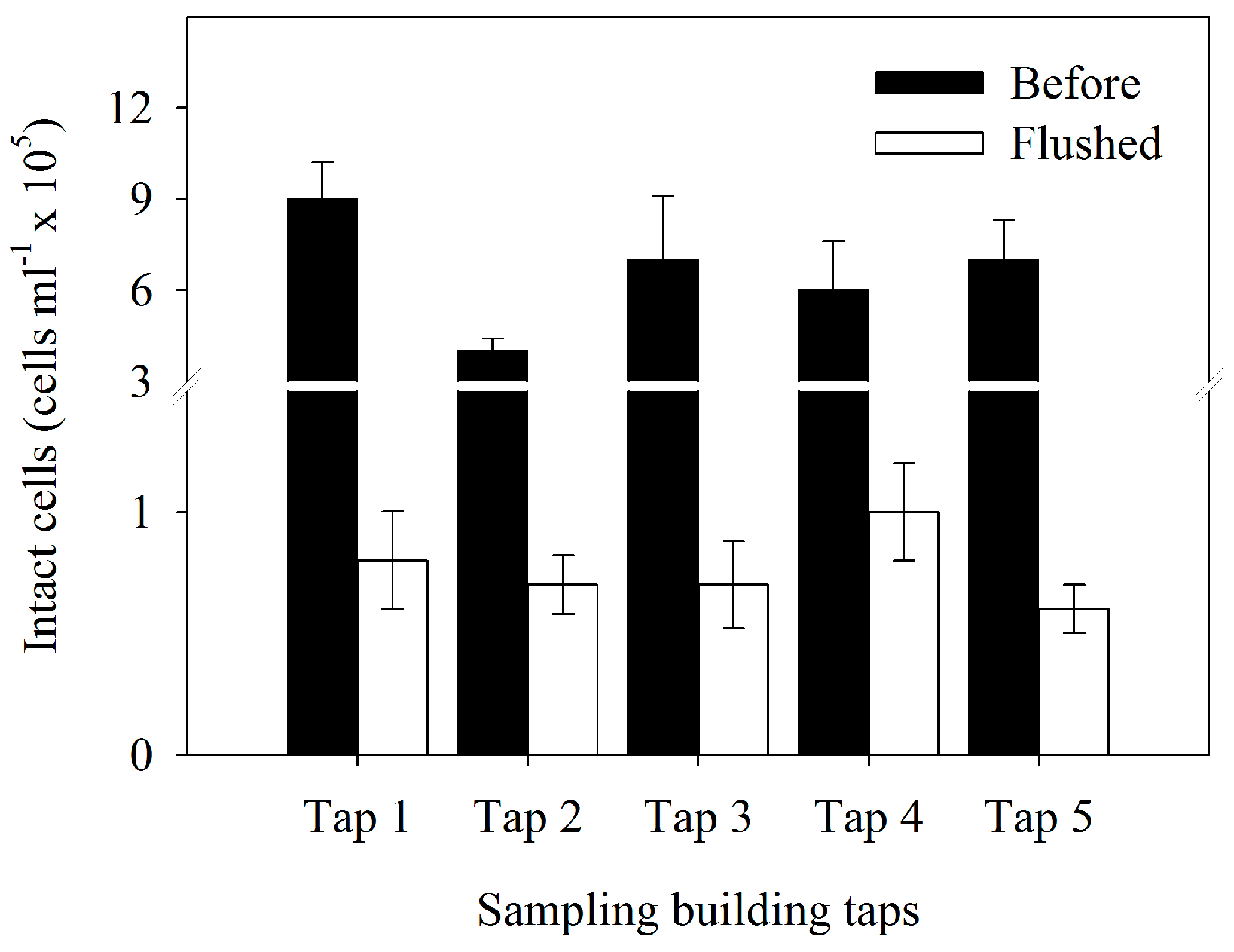
| Bacterial Parameters | Flushed (F-Value) | Tap (F-Value) | Interaction (F-Value) |
|---|---|---|---|
| Bacterial cell number | 252.65 *** | 12.56 ** | 6.83 * |
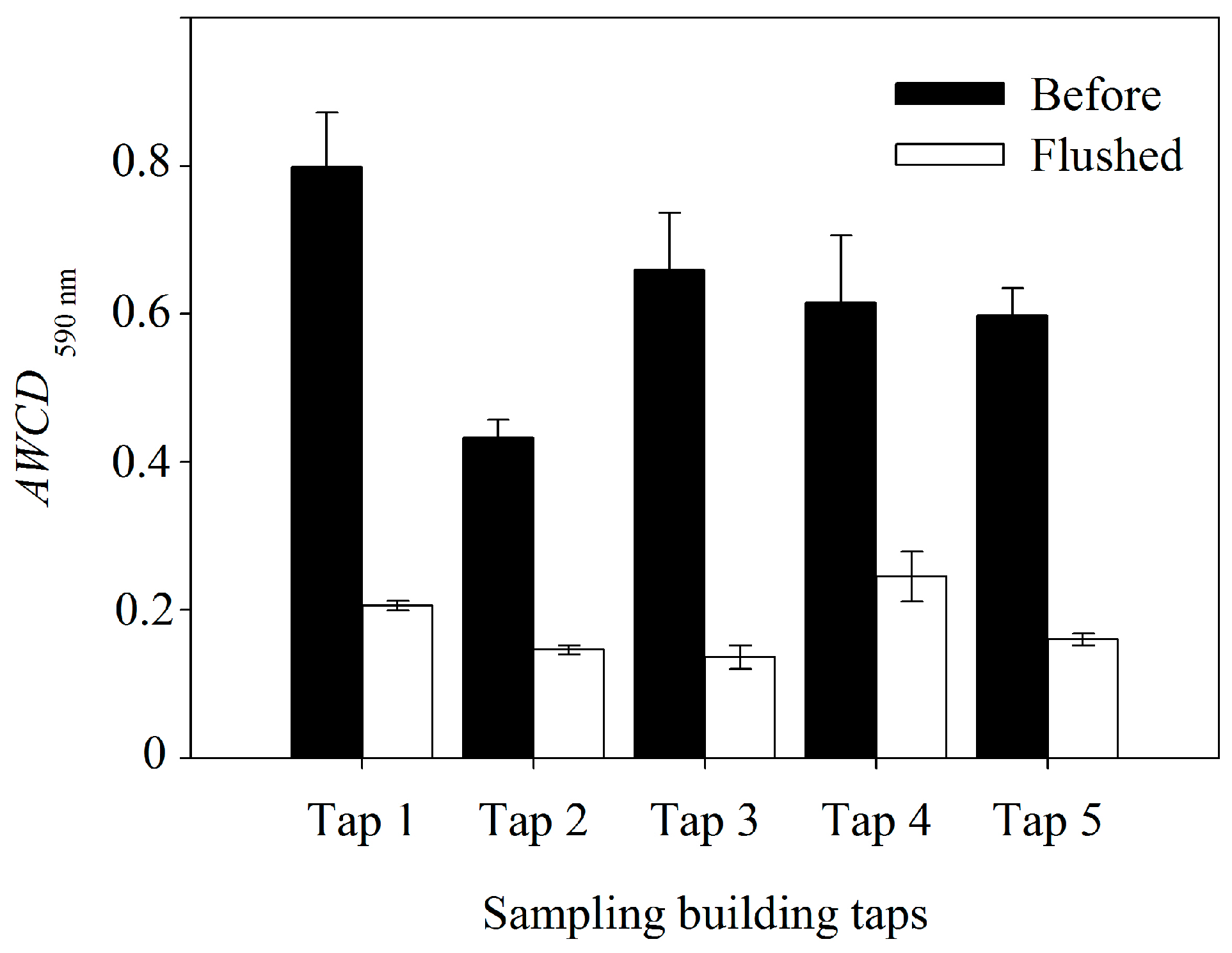
| AWCD590nm | 156.21 *** | 16.92 ** | 5.61 * |
| Carbohydrates | 58.61 ** | 1.04NS | 0.84NS |
| Carboxylic acids | 35.47 ** | 6.32 * | 5.04 * |
| Amino acids | 14.28 ** | 4.59 * | 1.09NS |
| Polymers | 1.53NS | 0.96NS | 0.08NS |
| Phenolic compounds | 19.21 ** | 8.62 ** | 1.68NS |
| Amines | 66.37 ** | 0.43NS | 1.35NS |
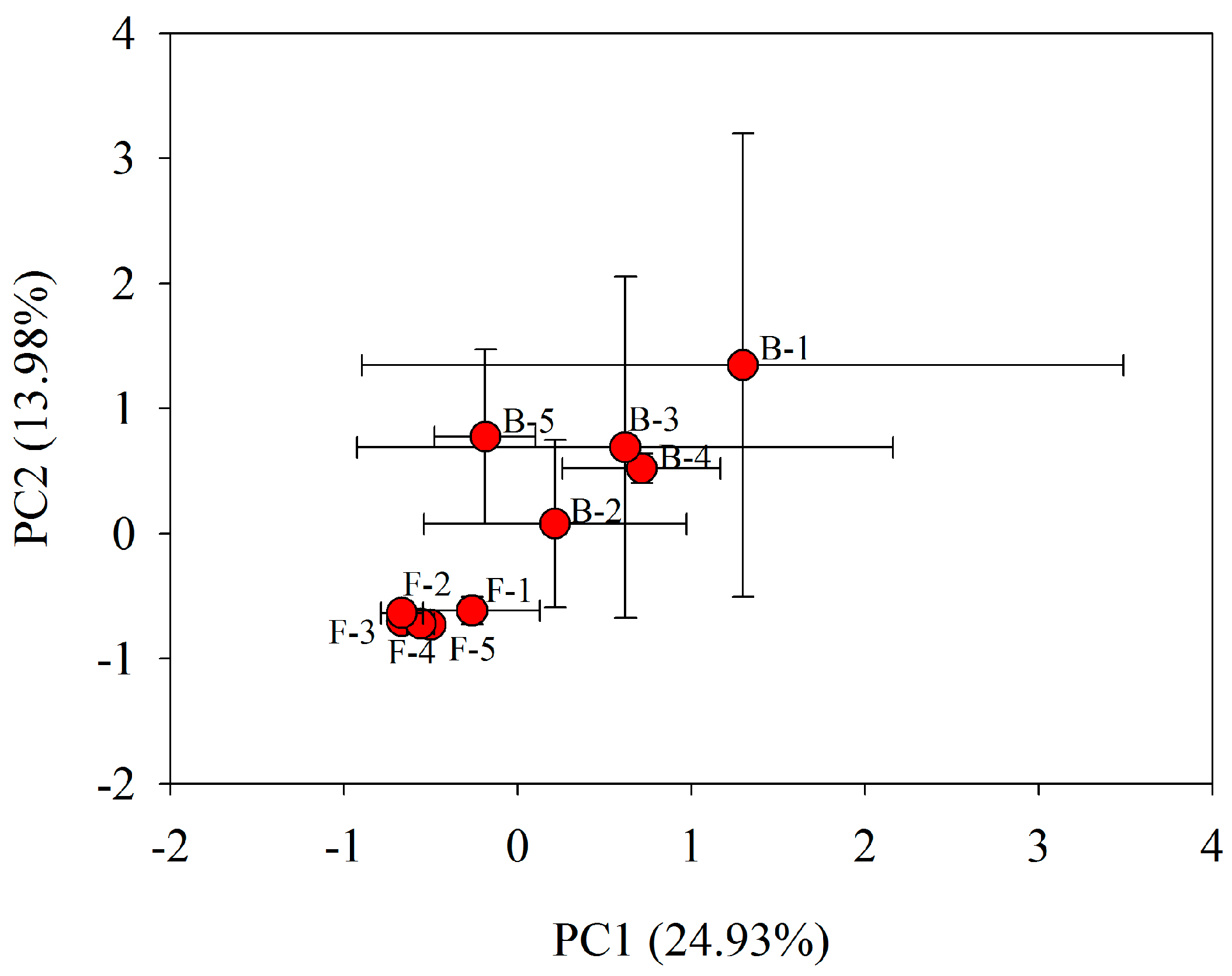
3.3. Bacterial Community Metabolic Profiles
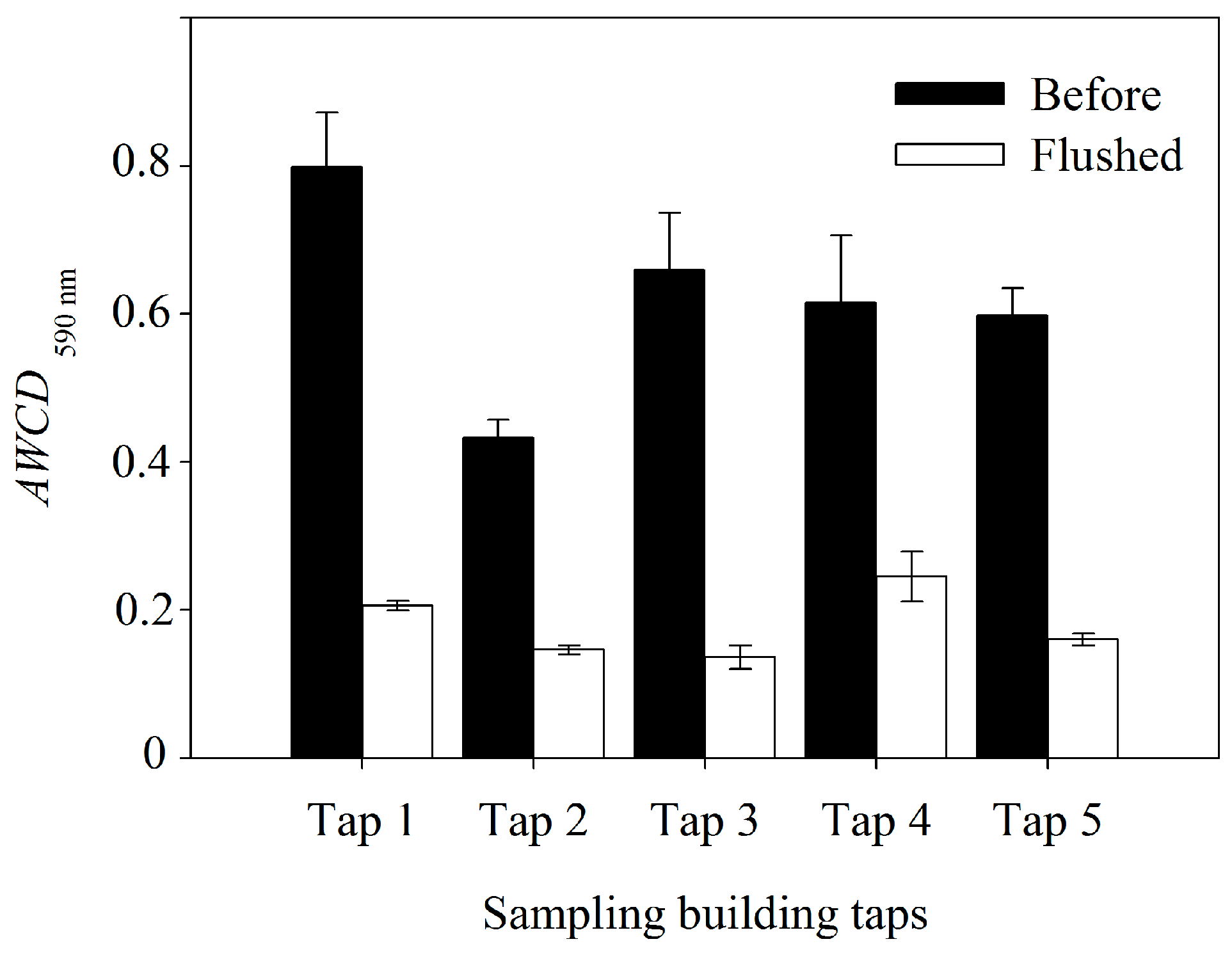

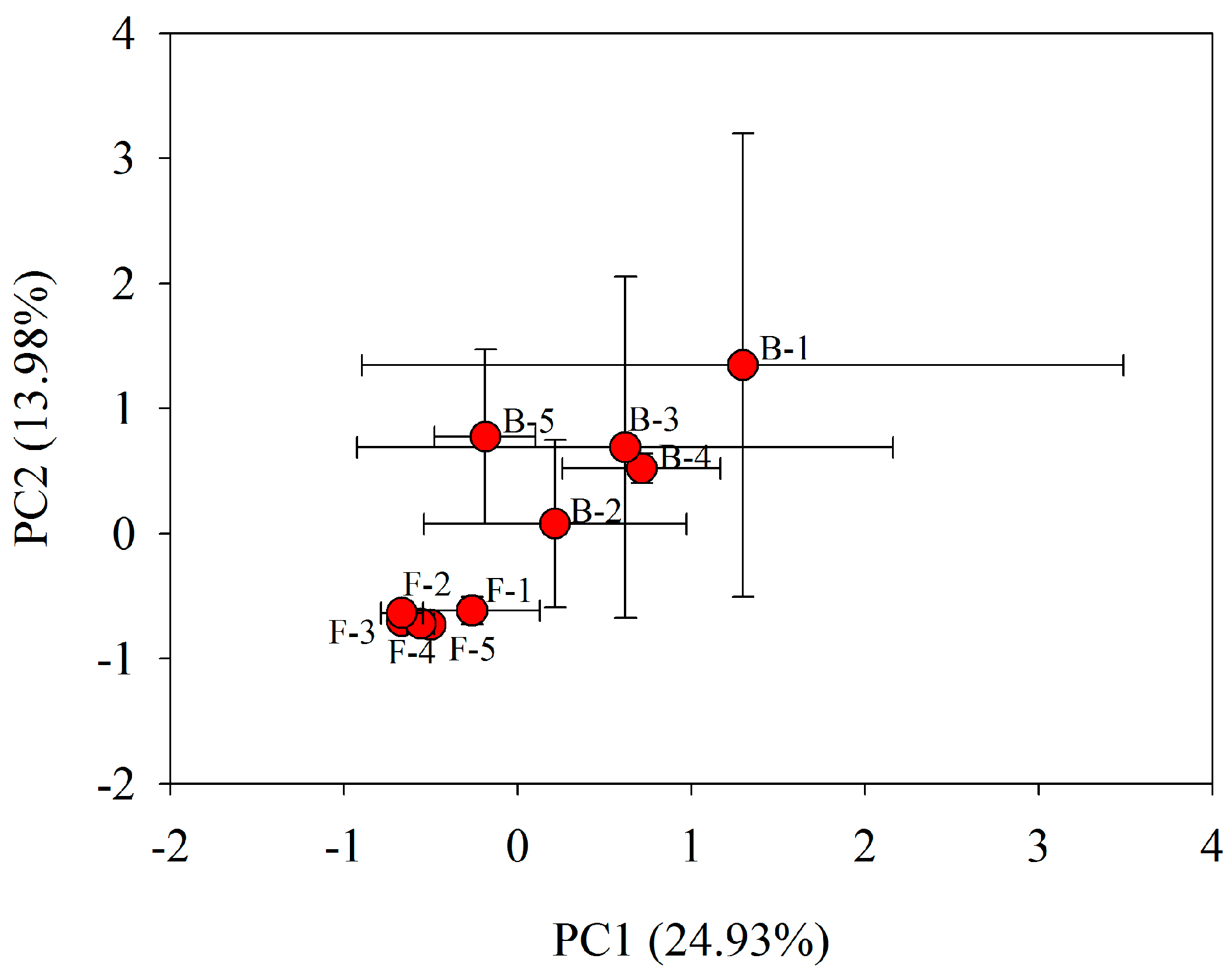
4. Conclusions
Acknowledgements
Author Contributions
Conflicts of Interest
References
- Pepper, I.L.; Rusin, P.; Quintanar, D.R.; Haney, C.; Josephson, K.L.; Gerba, C.P. Tracking the concentration of heterotrophic plate count bacteria from the source to the consumer’s tap. Int. J. Food Microbiol. 2004, 92, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Rintala, H.; Pitkäranta, M.; Toivola, M.; Paulin, L.; Nevalainen, A. Diversity and seasonal dynamics of bacterial community in indoor environment. BMC Microbiol. 2008, 8. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y. An evaluation of microbial and chemical contamination sources related to the deterioration of tap water quality in the household water supply system. Int. J. Environ. Res. Public Health 2013, 10, 4143–4160. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.H.; Huang, T.L.; Chen, S.N.; Guo, L.; Yang, X.; Liu, T.L. Spatial pattern of bacterial community functional diversity in a drinking water reservoir, Shaanxi Province, Northwest China. J. Pure Appl. Microb. 2013, 7, 1647–1654. [Google Scholar]
- Zhang, H.H.; Huang, T.L.; Chen, S.N.; Sekar, R.; Yang, X.; Lv, K. Abundance and diversity of bacteria in oxygen minimum drinking water reservoir sediments studied by quantitative PCR and pyrosequencing. Microb. Ecol. 2015, 69, 618–629. [Google Scholar] [CrossRef] [PubMed]
- Liao, X.; Chen, C.; Wang, Z.; Chang, C.H.; Zhang, X.; Xie, S. Bacterial community change through drinking water treatment processes. Int. J. Environ. Sci. Tech. 2014, 1–8. [Google Scholar] [CrossRef]
- Enkiri, F.; Legrand, J.Y.; Squinazi, F.; Ponelle Leroy, P. Assessment of microbial support potential of six materials used in drinking water distribution systems. Eur. J. Water Qual. 2006, 37, 175–188. [Google Scholar] [CrossRef]
- Lehtola, M.J.; Miettinen, I.T.; Keinänen, M.M.; Kekki, T.K.; Laine, O.; Hirvonen, A.; Vartiainen, T.; Martikainen, P.J. Microbiology, chemistry and biofilm development in a pilot drinking water distribution system with copper and plastic pipes. Water Res. 2004, 38, 3769–3779. [Google Scholar] [CrossRef] [PubMed]
- Lehtola, M.J.; Miettinen, I.T.; Lampola, T.; Hirvonen, A.; Vartiainen, T.; Martikainen, P.J. Pipeline materials modify the effectiveness of disinfectants in drinking water distribution systems. Water Res. 2005, 39, 1962–1971. [Google Scholar] [CrossRef] [PubMed]
- Lautenschlager, K.; Boon, N.; Wang, Y.Y.; Egli, T.; Hammes, F. Overnight stagnation of drinking water in household taps induces microbial growth and changes in community composition. Water Res. 2010, 44, 4868–4877. [Google Scholar] [CrossRef] [PubMed]
- Feng, T.; Li, X.; Ding, D.P. Numerical simulation of the heating system for the office building in Beijing area. J. Civil Archit. Environ. Eng. 2013, 35, 181–185. [Google Scholar]
- Yoshino, H.; Yoshino, Y.; Zhang, Q.; Mochida, A.; Li, N.; Li, Z.; Miyasaka, H. Indoor thermal environment and energy saving for urban residential buildings in China. Energ. Build. 2006, 38, 1308–1319. [Google Scholar] [CrossRef]
- Nam, S.C.; Lee, Y.J. Investigation of nutrients controlling bacterial regrowth in a PEX and stainless steel plumbing water system. Asian J. Chem. 2010, 22, 2427–2441. [Google Scholar]
- Berry, D.; Xi, C.; Raskin, L. Microbial ecology of drinking water distribution systems. Curr. Opin. Biotech. 2006, 17, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Jiang, Z. Changing of energy consumption patterns from rural households to urban households in China: An example from Shaanxi Province, China. Renew. Sust. Energ. Rev. 2008, 12, 1667–1680. [Google Scholar] [CrossRef]
- Huang, T.; Li, X.; Rijnaarts, H.; Grotenhuis, T.; Ma, W.; Sun, X.; Xu, J. Effects of storm runoff on the thermal regime and water quality of a deep, stratified reservoir in a temperate monsoon zone, in Northwest China. Sci. Total Environ. 2014, 485, 820–827. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Singer, P.C. Factors influencing the formation and relative distribution of haloacetic acids and trihalomethanes in drinking water. Environ. Sci. Technol. 2003, 37, 2920–2928. [Google Scholar] [CrossRef] [PubMed]
- Nescerecka, A.; Rubulis, J.; Vital, M.; Juhna, T.; Hammes, F. Biological instability in a chlorinated drinking water distribution network. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Van der Mark, E.J.; Verberk, J.Q.J.C.; Van Dijk, J.C. Flow cytometry total cell counts: A field study assessing microbiological water quality and growth in unchlorinated drinking water distribution systems. Biomed Res. Int. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Hammes, F.; Berney, M.; Wang, Y.; Vital, M.; Koster, O.; Egli, T. Flow-cytometric total bacterial cell counts as a descriptive microbiological parameter for drinking water treatment processes. Water Res. 2008, 42, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Garland, J.L. Analytical approaches to the characterization of samples of microbial communities using patterns of potential C utilization. Soil Biol. Biochem. 1996, 28, 223–231. [Google Scholar] [CrossRef]
- Classen, A.T.; Boyle, S.I.; Haskins, K.E.; Overby, S.T.; Hart, S.C. Community-level physiological profiles of bacteria and fungi: Plate type and incubation temperature influences on contrasting soils. FEMS Microbiol. Ecol. 2003, 44, 319–328. [Google Scholar] [CrossRef]
- Inkinen, J.; Kaunisto, T.; Pursiainen, A.; Miettinen, I.T.; Kusnetsov, J.; Riihinen, K.; Keinänen-Toivola, M.M. Drinking water quality and formation of biofilms in an office building during its first year of operation, a full scale study. Water Res. 2014, 49, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Bagh, L.K.; Albrechtsen, H.J.; Arvin, E.; Ovesen, K. Distribution of bacteria in a domestic hot water system in a Danish apartment building. Water Res. 2004, 38, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Lipphaus, P.; Hammes, F.; Kötzsch, S.; Green, J.; Gillespie, S.; Nocker, A. Microbiological tap water profile of a medium-sized building and effect of water stagnation. Environ. Technol. 2014, 35, 620–628. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S. Heterotrophic bacteria in drinking water distribution system: A review. Environ. Monit. Assess. 2012, 184, 6087–6137. [Google Scholar] [CrossRef] [PubMed]
- Charisiadis, P.; Andra, S.S.; Makris, K.C.; Christophi, C.A.; Skarlatos, D.; Vamvakousis, V.; Stephanou, E.G. Spatial and seasonal variability of tap water disinfection by-products within distribution pipe networks. Sci Total Environ. 2015, 506, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Pachepsky, Y.; Morrow, J.; Guber, A.; Shelton, D.; Rowland, R.; Davies, G. Effect of biofilm in irrigation pipes on microbial quality of irrigation water. Lett. Appl. Microbial. 2012, 54, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Rakic, A.; Peric, J.; Foglar, L. Influence of temperature, chlorine residual and heavy metals on the presence of Legionella pneumophila in hot water distribution systems. Ann. Agric. Environ. Med. 2012, 19, 431–436. [Google Scholar] [PubMed]
- Lin, W.; Yu, Z.; Chen, X.; Liu, R.; Zhang, H. Molecular characterization of natural biofilms from household taps with different materials: PVC, stainless steel, and cast iron in drinking water distribution system. Appl. Microbiol. Biotechnol. 2013, 97, 8393–8401. [Google Scholar] [CrossRef] [PubMed]
- Heinrichs, G.; Hübner, I.; Schmidt, C.K.; de Hoog, G.S.; Haase, G. Analysis of black fungal biofilms occurring at domestic water taps (II): Potential routes of entry. Mycopathologia 2013, 175, 399–412. [Google Scholar] [CrossRef] [PubMed]
- Heinrichs, G.; Hübner, I.; Schmidt, C.K.; de Hoog, G.S.; Haase, G. Analysis of black fungal biofilms occurring at domestic water taps. I: Compositional analysis using Tag-encoded FLX amplicon pyrosequencing. Mycopathologia 2013, 175, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Kelley, K.M.; Stenson, A.C.; Dey, R.; Whelton, A.J. Release of drinking water contaminants and odor impacts caused by green building cross-linked polyethylene (PEX) plumbing systems. Water Res. 2014, 67, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.M.; Cupples, A.M. Diversity of five anaerobic toluene-degrading microbial communities investigated using stable isotope probing. Appl. Environ. Microbiol. 2012, 78, 972–980. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.M.; Sun, X.X.; Cupples, A.M. Anaerobic methyl tert-butyl ether-degrading microorganisms identified in wastewater treatment plant samples by stable isotope probing. Appl. Environ. Microbiol. 2012, 78, 2973–2980. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.M.; Xiao, T.; Sun, M.; Dong, Y.; Ning, Z.; Xiao, E.; Li, J. Diversity of the sediment microbial community in the Aha watershed (Southwest China) in response to acid mine drainage pollution gradients. Appl. Environ. Microbiol. 2015, 81, 4874–4884. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, S.; Kleinsteuber, S.; Chatzinotas, A.; Kuppardt, S.; Lueders, T.; Richnow, H.H.; Vogt, C. Functional characterization of an anaerobic benzene-degrading enrichment culture by DNA stable isotope probing. Environ. Microbiol. 2010, 12, 401–411. [Google Scholar] [CrossRef] [PubMed]
- Kasai, Y.; Takahata, Y.; Manefield, M.; Watanabe, K. RNA-based stable isotope probing and isolation of anaerobic benzene-degrading bacteria from gasoline-contaminated groundwater. Appl. Environ. Microbiol. 2006, 72, 3586–3592. [Google Scholar] [CrossRef] [PubMed]
- Kunapuli, U.; Lueders, T.; Meckenstock, R.U. The use of stable isotope probing to identify key iron-reducing microorganisms involved in anaerobic benzene degradation. ISME J. 2007, 1, 643–653. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.-H.; Chen, S.-N.; Huang, T.-L.; Shang, P.-L.; Yang, X.; Ma, W.-X. Indoor Heating Drives Water Bacterial Growth and Community Metabolic Profile Changes in Building Tap Pipes during the Winter Season. Int. J. Environ. Res. Public Health 2015, 12, 13649-13661. https://doi.org/10.3390/ijerph121013649
Zhang H-H, Chen S-N, Huang T-L, Shang P-L, Yang X, Ma W-X. Indoor Heating Drives Water Bacterial Growth and Community Metabolic Profile Changes in Building Tap Pipes during the Winter Season. International Journal of Environmental Research and Public Health. 2015; 12(10):13649-13661. https://doi.org/10.3390/ijerph121013649
Chicago/Turabian StyleZhang, Hai-Han, Sheng-Nan Chen, Ting-Lin Huang, Pan-Lu Shang, Xiao Yang, and Wei-Xing Ma. 2015. "Indoor Heating Drives Water Bacterial Growth and Community Metabolic Profile Changes in Building Tap Pipes during the Winter Season" International Journal of Environmental Research and Public Health 12, no. 10: 13649-13661. https://doi.org/10.3390/ijerph121013649
APA StyleZhang, H.-H., Chen, S.-N., Huang, T.-L., Shang, P.-L., Yang, X., & Ma, W.-X. (2015). Indoor Heating Drives Water Bacterial Growth and Community Metabolic Profile Changes in Building Tap Pipes during the Winter Season. International Journal of Environmental Research and Public Health, 12(10), 13649-13661. https://doi.org/10.3390/ijerph121013649