West Nile Virus Transmission in Sentinel Chickens and Potential Mosquito Vectors, Senegal River Delta, 2008–2009

Abstract
:1. Introduction
2. Material and Methods
3. Results
3.1. Sentinel Serology

{kind=link}
{kind=link}
| Cohort | I1 | I2 | I3 | I4 | I5 | I6 | IT |
|---|---|---|---|---|---|---|---|
| P1 | 0.0 (0/51) | 0.0 (0/47) | 0.0 (0/42) | 0.0 (0/41) | 0.0 (0/40) | 0.0 (0/40) | 0.0 (0/43) |
| P2 | 0.0 (0/73) | 1.4 (1/69) | 3.2 (2/63) | 0.0 (0/60) | 0.0 (0/57) | 0.0 (0/52) | 4.8 (3/62) |
| P3 | 0.0 (0/55) | 5.7 (3/53) | 2.1 (1/48) | 0.0 (0/44) | 0.0 (0/42) | 0.0 (0/42) | 8.5 (4/47) |
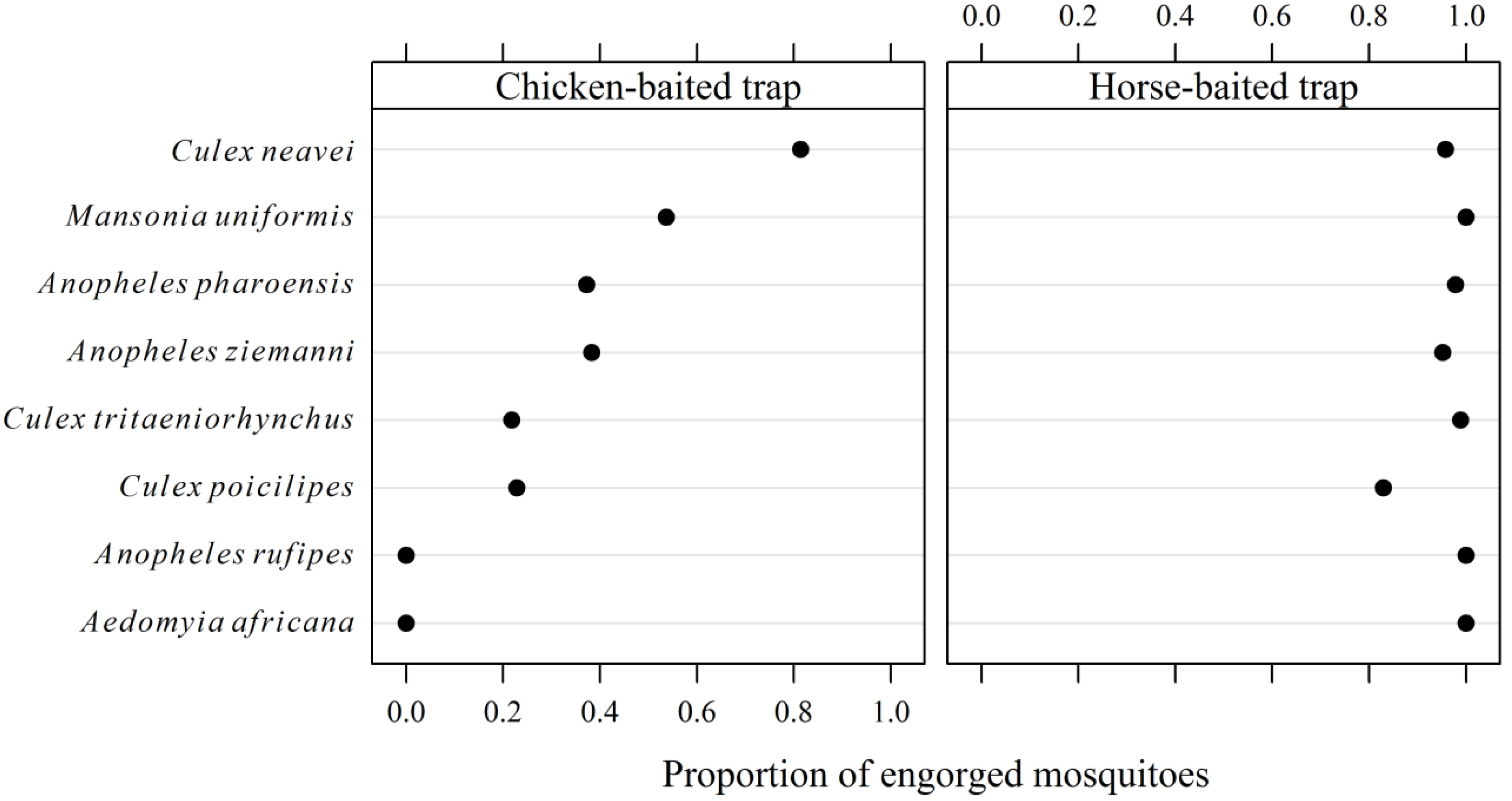
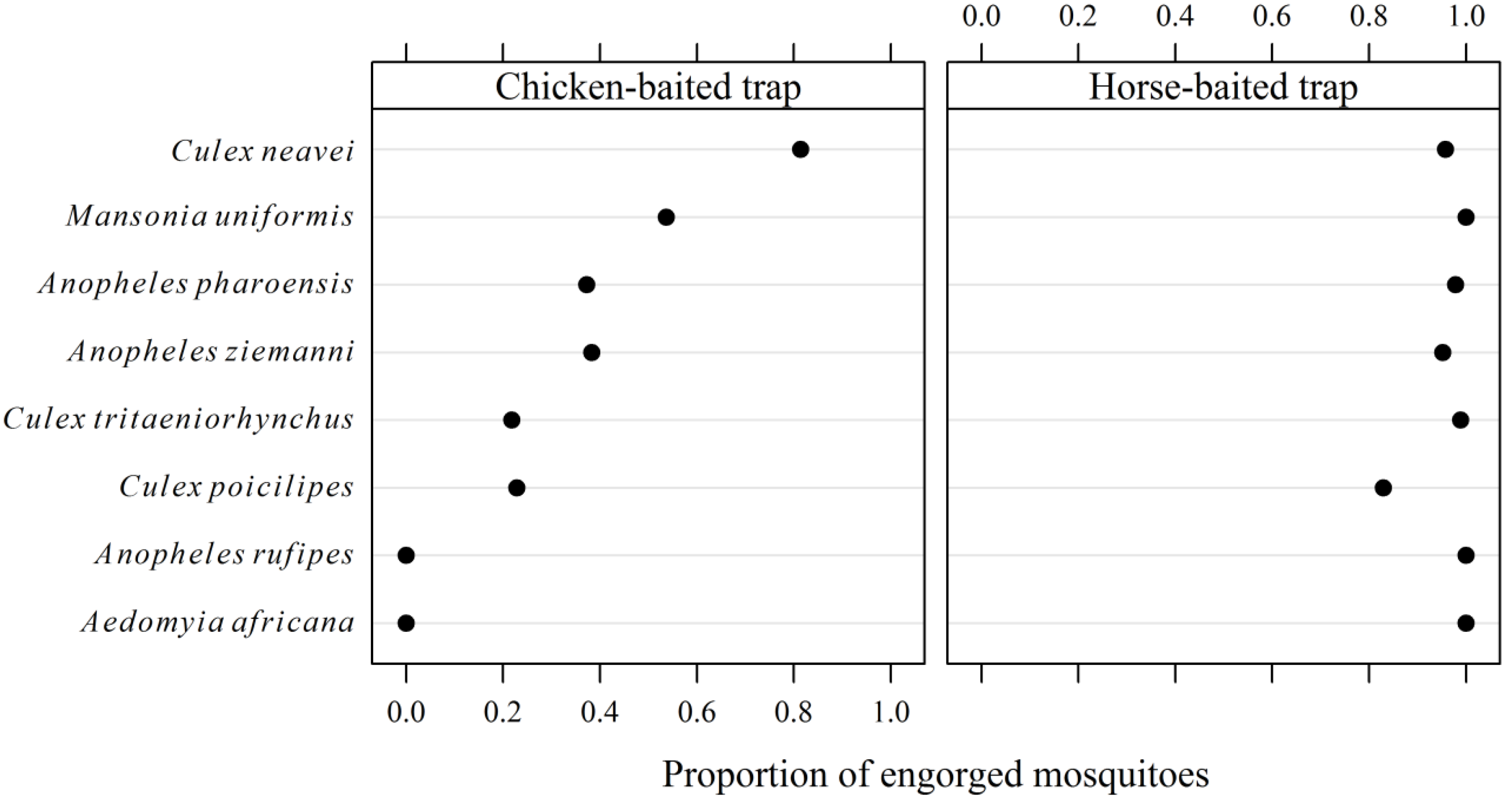
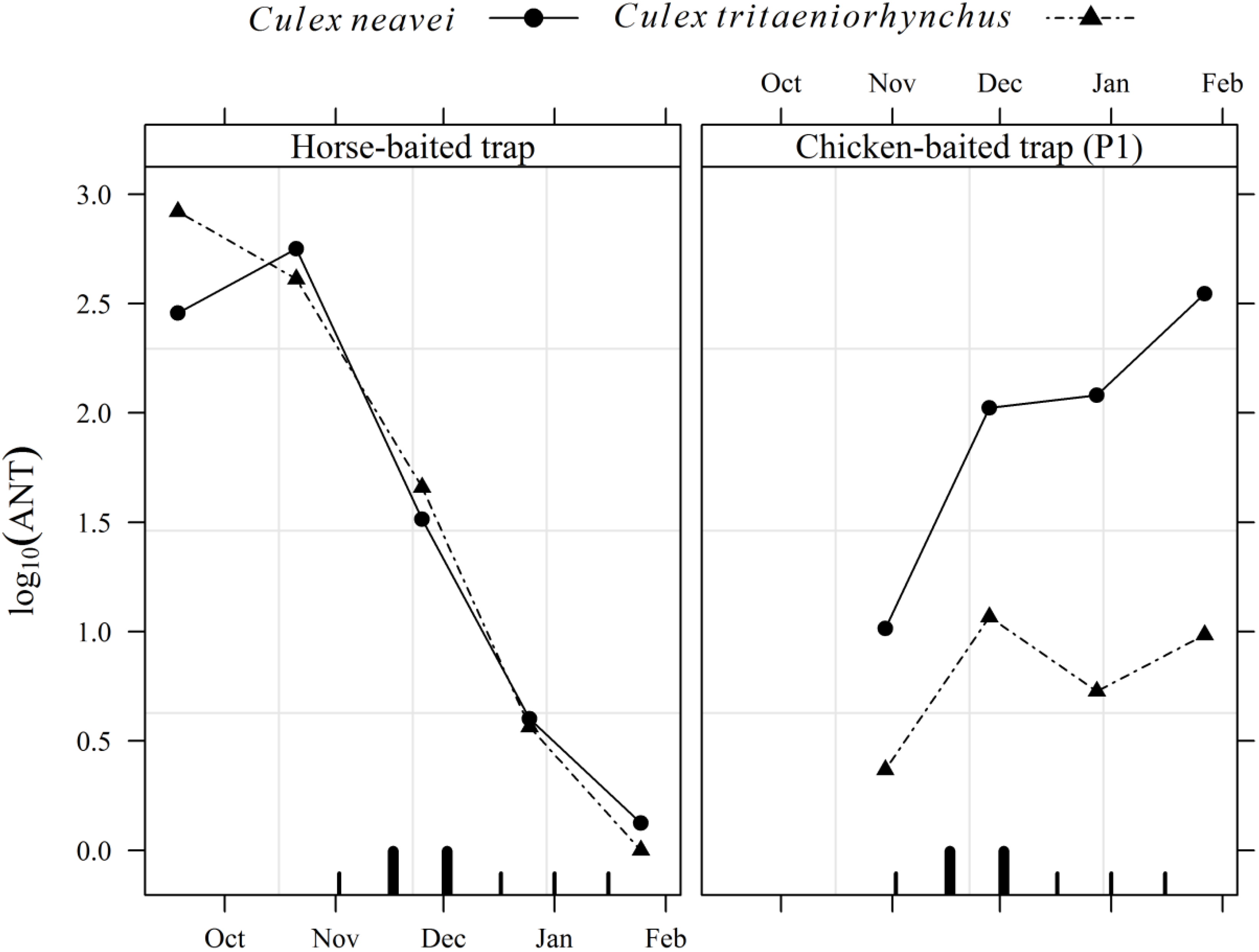
3.2. Entomological Survey
| Mosquito species | Horse-baited trap (n = 15) | Chicken-baited traps | Total (%) | ||
|---|---|---|---|---|---|
| P1 (n = 12) | P2 (n = 12) | P3 (n = 12) | |||
| Culex neavei | 176.7 ± 275.5 | 23.7 ± 15.1 | 44.4 ± 29.9 | 145.9 ± 133.1 | 5,218 (51.20) |
| Culex tritaeniorhynchus | 257.4 ± 485.0 | 4.8 ± 8.3 | 2.5 ± 2.7 | 6.3 ± 4.3 | 4,023 (39.47) |
| Mansonia uniformis | 2.2 ± 3.2 | 0.0 | 0.1 ± 0.3 | 1.0 ± 1.2 | 46 (0.45) |
| Aedomyia africana | 0.1 ± 0.3 | 0.0 | 0.0 | 0.0 | 1 (0.01) |
| Anopheles rufipes | 0.1 ± 0.3 | 0.0 | 0.0 | 0.0 | 1 (0.01) |
| Anopheles pharoensis | 29.8 ± 44.3 | 0.4 ± 0.9 | 0.1 ± 0.3 | 1.7 ± 2.1 | 473 (4.64) |
| Anopheles ziemanni | 7.1 ± 8.7 | 0.2 ± 0.4 | 0.0 | 0.5 ± 1.2 | 115 (1.13) |
| Culex poicilipes | 20.1 ± 26.3 | 0.1 ± 0.3 | 0.1 ± 0.3 | 0.9 ± 1.3 | 315 (3.09) |
| Total | 10,192 | ||||
4. Discussion
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Kramer, L.D.; Styer, L.M.; Ebel, G.D. A global perspective on the epidemiology of West Nile virus. Annu. Rev. Entomol. 2008, 53, 61–81. [Google Scholar] [CrossRef]
- Fontenille, D.; Traoré-Lamizana, M.; Diallo, M.; Thonnon, J.; Digoutte, J.P.; Zeller, H.G. New vectors of Rift Valley fever in West Africa. Emerg. Infect. Dis. 1998, 4, 289–93. [Google Scholar] [CrossRef]
- Chevalier, V.; Lancelot, R.; Diaïté, A.; Mondet, B.; Sall, B.; Lamballerie, X.D. Serological assessment of West Nile fever virus activity in the pastoral system of Ferlo, Senegal. Ann. N. Y. Acad. Sci. 2006, 1081, 216–225. [Google Scholar]
- Chevalier, V.; Reynaud, P.; Lefrançois, T.; Durand, B.; Baillon, F.; Balança, G.; Gaidet, N.; Mondet, B.; Lancelot, R. Predicting West Nile virus seroprevalence in wild birds in Senegal. Vector Borne Zoonotic Dis. 2009, 9, 589–596. [Google Scholar] [CrossRef]
- Bayly, N.J.; Atkinson, P.W.; Rumsey, S.J. Fuelling for the Sahara crossing: variation in site use and the onset and rate of spring mass gain by 38 Palearctic migrants in the western Sahel. Journal of Ornithology 2012, 153, 931–945. [Google Scholar] [CrossRef]
- Jourdain, E.; Gauthier-Clerc, M.; Bicout, D.J.; Sabatier, P. Bird migration routes and risk for pathogen dispersion into western Mediterranean wetlands. Emerg. Infect. Dis. 2007, 13, 365–372. [Google Scholar] [CrossRef]
- Fall, A.G.; Diaïté, A.; Lancelot, R.; Tran, A.; Soti, V.; Etter, É.; Konaté, L.; Faye, O.; Bouyer, J. Feeding behaviour of potential vectors of West Nile virus in Senegal. Parasit. Vectors 2011, 4, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Morel, G.J.; Roux, F. Les migrateurs paléarctiques au Sénégal. II. Passereaux et synthèse générale. La Terre et la Vie 1966, 2, 143–176. [Google Scholar]
- Triplet, P.; Yésou, P. Controlling the flood in the Senegal Delta: do waterfowl populations adapt to their new environment? Ostrich 2000, 71, 106–111. [Google Scholar] [CrossRef]
- Langevin, S.A.; Bunning, M.; Davis, B.; Komar, N. Experimental infection of chickens as candidate sentinels for West Nile virus. Emerg. Infect. Dis. 2001, 7, 726–729. [Google Scholar]
- Blitvich, B.J.; Bowen, R.A.; Marlenee, N.L.; Hall, R.A.; Bunning, M.L.; Beaty, B.J. Epitope-blocking enzyme-linked immunosorbent assays for detection of West Nile virus antibodies in domestic mammals. J. Clin. Microbiol. 2003, 41, 2676–2679. [Google Scholar] [CrossRef]
- Blitvich, B.J.; Marlenee, N.L.; Hall, R.A.; Calisher, C.H.; Bowen, R.A.; Roehrig, J.T.; Komar, N.; Langevin, S.A.; Beaty, B.J. Epitope-blocking enzyme-linked immunosorbent assays for the detection of serum antibodies to West Nile virus in multiple avian species. J. Clin. Microbiol. 2003, 41, 1041–1047. [Google Scholar] [CrossRef]
- Lefrançois, T.; Blitvich, B.J.; Pradel, J.; Molia, S.; Vachi’ery, N.; Martinez, D. West Nile virus in Guadeloupe: introduction, spread, and decrease in circulation level: 2002-2005. Ann. N. Y. Acad. Sci. 2006, 1081, 206–215. [Google Scholar]
- Lefrançois, T.; Blitvich, B.; Pradel, J.; Molia, S.; Vachiéry, N.; Pallavicini, P.; Marlenee, N.; Zientara, S.; Petitclerc, M.; Martinez, D. West Nile virus surveillance, Guadeloupe, 2003–2004. Emerg. Infect. Dis. 2005, 11, 1100–1103. [Google Scholar] [CrossRef]
- Bargaoui, R.; Lecollinet, S.; Lancelot, R. Mapping the serological prevalence rate of West Nile fever in equids, Tunisia. Transbound. Emerg. Dis. 2013, 1–12. [Google Scholar] [CrossRef]
- Lesnoff, M.; Lancelot, R.; Moulin, C.H.; Messad, S.; Juanès, X.; Sahut, C. Calculation of demographic parameters in tropical livestock herds. A discrete time approach with LASER animal-based monitoring data; QUAE: Montpellier, France, 2011; p. 99. [Google Scholar]
- Balenghien, T.; Fouque, F.; Sabatier, P.; Bicout, D.J. Horse-, bird-, and human-seeking behavior and seasonal abundance of mosquitoes in a West Nile virus focus of southern France. J. Med. Entomol. 2006, 43, 936–946. [Google Scholar] [CrossRef]
- Edwards, F.W. Mosquitoes of the Ethiopian Region. III.—Culicine Adults and Pupae; Brit. Mus. (NH): London, UK, 1941; p. 499. [Google Scholar]
- Diagne, N.; Fontenille, D.; Konate, L.; Faye, O.; Lamizana, M.; Legros, F.; Molez, J.F.; Trape, J.F. Les anophèles du Sénégal: liste commentée et illustrée. Bull. Soc. Pathol. Exot. 1994, 87, 267–277. [Google Scholar]
- Diallo, M.; Nabeth, P.; Ba, K.; Sall, A.A.; Ba, Y.; Mondo, M.; Girault, L.; Abdalah, M.O.; Mathiot, C. Mosquito vectors of the 1998–1999 outbreak of Rift Valley fever and other arboviruses (Bagaza, Sanar, Wesselsbron and West Nile) in Mauritania and Senegal. Med. Vet. Entomol. 2005, 19, 119–126. [Google Scholar] [CrossRef]
- Traoré-Lamizana, M.; Fontenille, D.; Diallo, M.; Bâ, Y.; Zeller, H.G.; Mondo, M.; Adam, F.; Thonon, J.; Maïga, A. Arbovirus surveillance from 1990 to 1995 in the Barkedji area (Ferlo) of Senegal, a possible natural focus of Rift Valley fever virus. J. Med. Entomol. 2001, 38, 480–492. [Google Scholar] [CrossRef]
- Traoré-Lamizana, M.; Zeller, H.G.; Mondo, M.; Hervy, J.P.; Adam, F.; Digoutte, J.P. Isolations of West Nile and Bagaza viruses from mosquitoes (Diptera: Culicidae) in central Senegal (Ferlo). J. Med. Entomol. 1994, 31, 934–938. [Google Scholar]
- Chevalier, V.; Lancelot, R.; Diaïté, A.; Mondet, B.; De Lamballerie, X. Use of sentinel chickens to study the transmission dynamics of West Nile virus in a Sahelian ecosystem. Epidemiol. Infect. 2008, 136, 525–528. [Google Scholar]
- Cabre, O.; Grandadam, M.; Marié, J.L.; Gravier, P.; Prangé, A.; Santinelli, Y.; Rous, V.; Bourry, O.; Durand, J.P.; Tolou, H.; Davoust, B. West Nile Virus in horses, sub-Saharan Africa. Emerg. Infect. Dis. 2006, 12, 1958–1960. [Google Scholar] [CrossRef]
- Chevalier, V.; Dupressoir, A.; Tran, A.; Diop, O.M.; Gottland, C.; Diallo, M.; Etter, E.; Ndiaye, M.; Grosbois, V.; Dia, M.; Gaidet, N.; Sall, A.A.; Soti, V.; Niang, M. Environmental risk factors of West Nile virus infection of horses in the Senegal River basin. Epidemiol. Infect. 2010, 138, 1601–1609. [Google Scholar] [CrossRef] [Green Version]
- Murgue, B.; Murri, S.; Triki, H.; Deubel, V.; Zeller, H.G. West Nile in the Mediterranean basin: 1950-2000. Ann. N. Y. Acad. Sci. 2001, 951, 117–126. [Google Scholar]
- Diouf, N. Potential epidemiological role of the Senegal River Delta and arthropod-borne-virus impacts within the horse population: case of West Nile fever (WNF) and African horse sickness (AHS). PhD Thesis, Ecole Doctorale Sciences de la Vie, de la Santé et de l’Environnement—Université Cheikh Anta Diop, Dakar, Senegal, 28 June 2013; p. 188. [Google Scholar]
- Gordon, S.W.; Tammariello, R.F.; Linthicum, K.J.; Dohm, D.J.; Digoutte, J. Arbovirus isolations from mosquitoes collected during 1988 in the Senegal River basin. Am. J. Trop. Med. Hyg. 1992, 47, 742–748. [Google Scholar]
- Chaskopoulou, A.; Dovas, C.I.; Chaintoutis, S.C.; Bouzalas, I.; Ara, G.; Papanastassopoulou, M. Evidence of enzootic circulation of West Nile virus (Nea Santa-Greece-2010,lineage 2), Greece, May to July 2011. Euro Surveill. 2011, 16. Available online: http://www.eurosurveillance.org/ViewArticle.aspx?ArticleId=19933 (accessed on 29 September 2013).
- Komar, N. West Nile virus surveillance using sentinel birds. Ann. N. Y. Acad. Sci. 2001, 951, 58–73. [Google Scholar] [CrossRef]
- Migliani, R.; Tehindrazanarivelo, A.; Rasamoelisoa, J.; Raobijaona, H.; Rakotonirina, G.; Ramamonjisoa, J.; Ratsitorahina, M.; Ramarokoto, C.; Grosjean, P.; Rakoto-Andrianarivelo, M.; Rousset, D. Proceedings of Epidemiology of acute encephalitis in Antananarivo. Colloque Scientifique International: “Santé, Environnement et Développement”., Antananarivo, Madagascar, 24–25 July 2002.
- Jentes, E.S.; Robinson, J.; Johnson, B.W.; Conde, I.; Sakouvougui, Y.; Iverson, J.; Beecher, S.; Bah, M.A.; Diakite, F.; Coulibaly, M.; Bausch, D.G.; Bryan, J. Acute arboviral infections in Guinea, West Africa, 2006. Am. J. Trop. Med. Hyg. 2010, 83, 388–394. [Google Scholar] [CrossRef]
- Estival, J.L.; Skowron, F.; Dupin, M.; Combemale, P. Primo-infection à virus West-Nile [Primary infection with West-Nile virus]. Ann. Dermatol. Venereol. 2001, 128, 656–658. [Google Scholar]
- Anderson, R.C.; Horn, K.B.; Hoang, M.P.; Gottlieb, E.; Bennin, B. Punctate exanthem of West Nile Virus infection: report of 3 cases. J. Am. Acad. Dermatol. 2004, 51, 820–823. [Google Scholar] [CrossRef]
- Larrieu, S.; Cardinale, E.; Ocquidant, P.; Roger, M.; Lepec, R.; Delatte, H.; Camuset, G.; Desprès, P.; Brottet, E.; Charlin, C.; Michault, A. A fatal neuroinvasive West Nile virus infection in a traveler returning from Madagascar: clinical, epidemiological and veterinary investigations. Am. J. Trop. Med. Hyg. 2013, 89, 211–213. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Fall, A.G.; Diaïté, A.; Seck, M.T.; Bouyer, J.; Lefrançois, T.; Vachiéry, N.; Aprelon, R.; Faye, O.; Konaté, L.; Lancelot, R. West Nile Virus Transmission in Sentinel Chickens and Potential Mosquito Vectors, Senegal River Delta, 2008–2009. Int. J. Environ. Res. Public Health 2013, 10, 4718-4727. https://doi.org/10.3390/ijerph10104718
Fall AG, Diaïté A, Seck MT, Bouyer J, Lefrançois T, Vachiéry N, Aprelon R, Faye O, Konaté L, Lancelot R. West Nile Virus Transmission in Sentinel Chickens and Potential Mosquito Vectors, Senegal River Delta, 2008–2009. International Journal of Environmental Research and Public Health. 2013; 10(10):4718-4727. https://doi.org/10.3390/ijerph10104718
Chicago/Turabian StyleFall, Assane Gueye, Amadou Diaïté, Momar Talla Seck, Jérémy Bouyer, Thierry Lefrançois, Nathalie Vachiéry, Rosalie Aprelon, Ousmane Faye, Lassana Konaté, and Renaud Lancelot. 2013. "West Nile Virus Transmission in Sentinel Chickens and Potential Mosquito Vectors, Senegal River Delta, 2008–2009" International Journal of Environmental Research and Public Health 10, no. 10: 4718-4727. https://doi.org/10.3390/ijerph10104718
APA StyleFall, A. G., Diaïté, A., Seck, M. T., Bouyer, J., Lefrançois, T., Vachiéry, N., Aprelon, R., Faye, O., Konaté, L., & Lancelot, R. (2013). West Nile Virus Transmission in Sentinel Chickens and Potential Mosquito Vectors, Senegal River Delta, 2008–2009. International Journal of Environmental Research and Public Health, 10(10), 4718-4727. https://doi.org/10.3390/ijerph10104718