Evaluation of Marine Brown Algae and Sponges from Brazil as Anticoagulant and Antiplatelet Products


Abstract
:1. Introduction
2. Results and Discussion
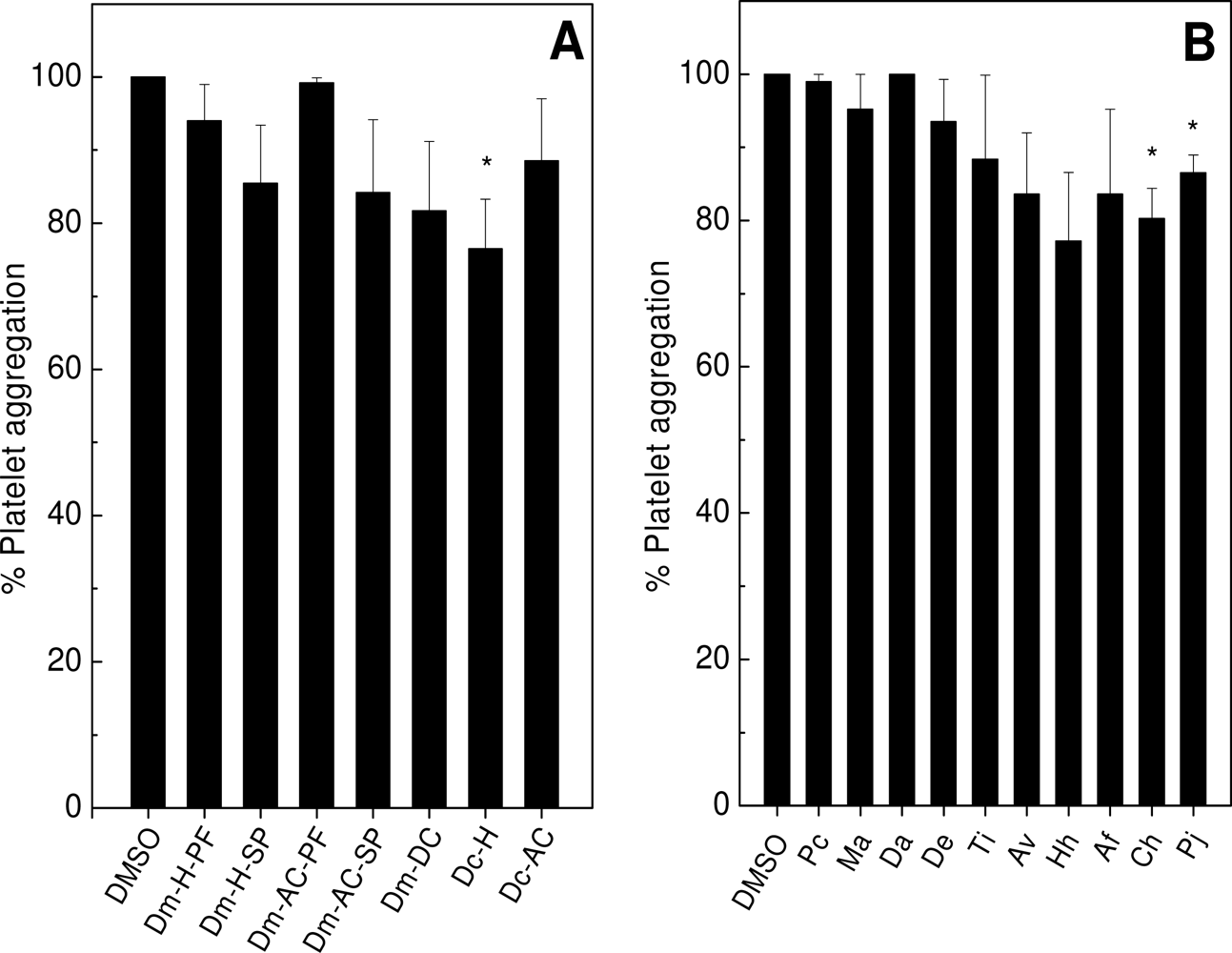
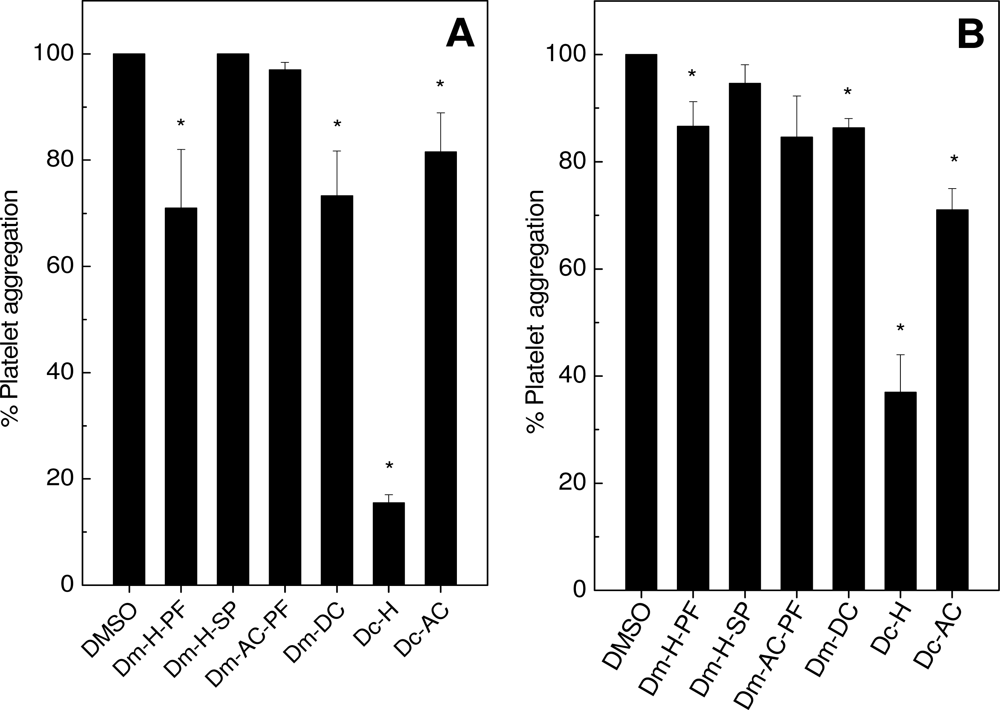
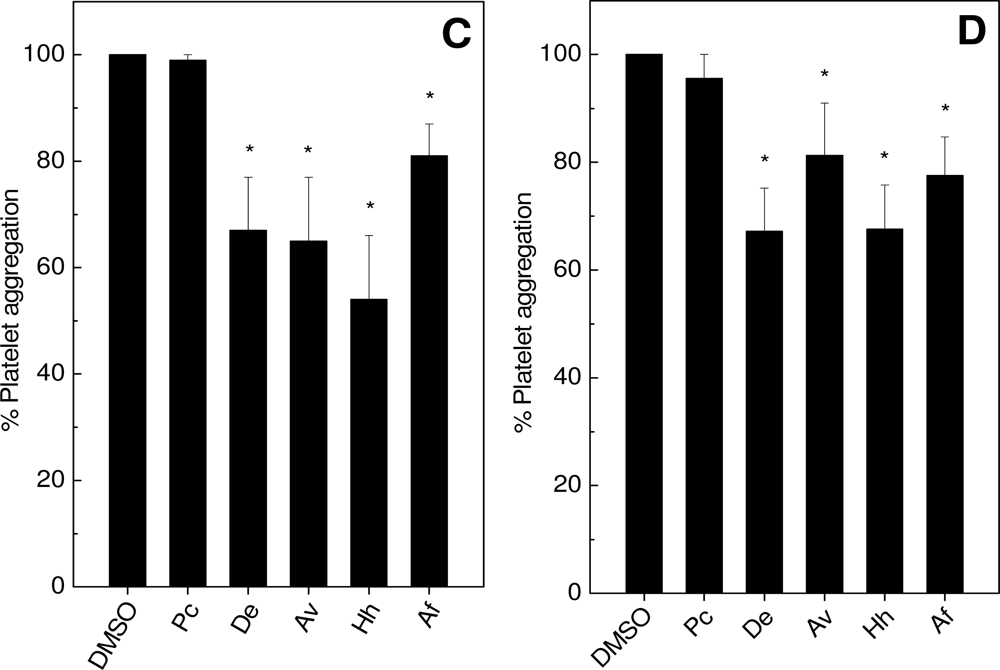
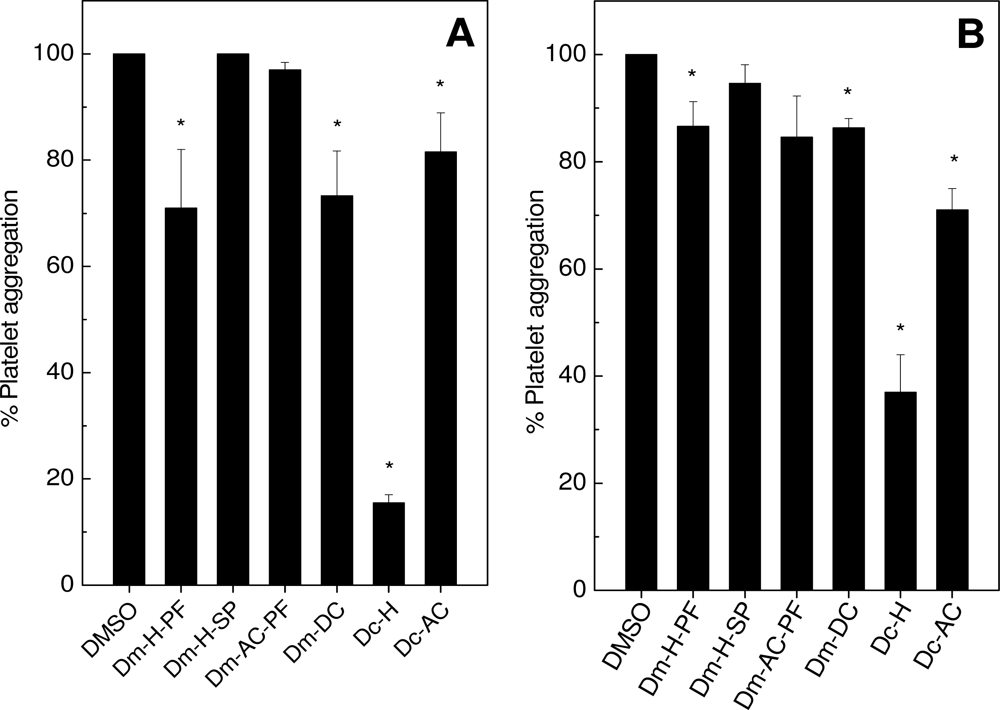
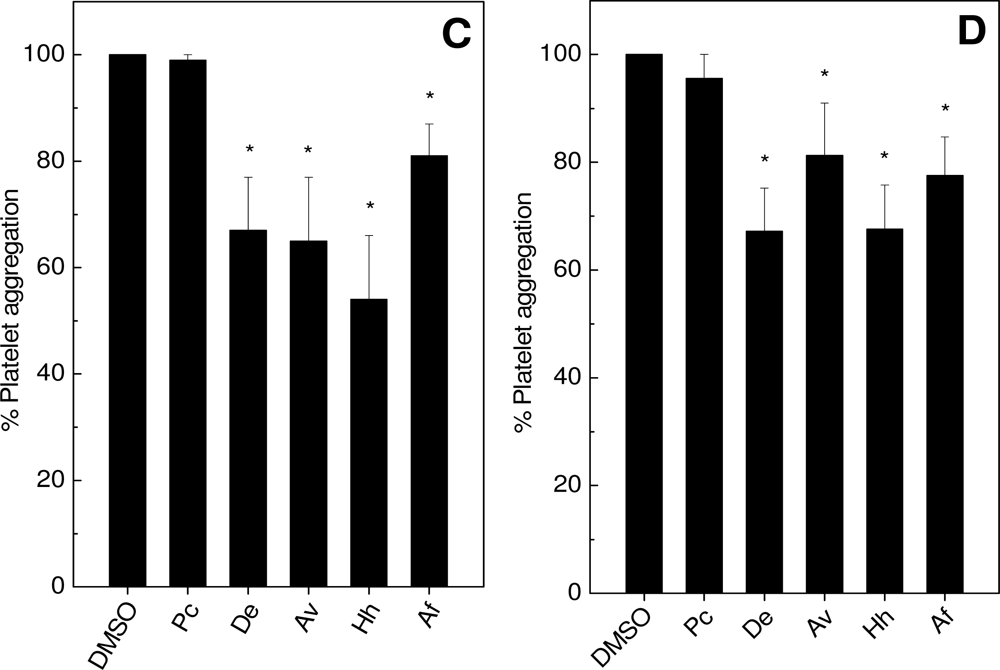
2.1. Effects on Platelet Aggregation
2.2. Effects on Coagulation
3. Experimental Section
3.1. Material
3.2. Marine Organisms
3.3. Platelet Aggregation Assays
3.4. Clotting Assays
3.5. Hydrolytic Activity upon Chromogenic Assay
3.6. Statistical Analyses
4. Conclusions
Acknowledgments
- Samples Availability: Available from the authors.
References
- Esmon, CT. Regulation of blood coagulation. Biochim. Biophys. Acta 2000, 1477, 349–360. [Google Scholar]
- Andrews, RK; Berndt, MC. Platelet physiology and thrombosis. Thromb. Res. 2004, 114, 447–453. [Google Scholar]
- Agnelli, G. Current issues in Anticoagulation. Pathophysiol. Haemost. Thromb. 2005, 34, 2–9. [Google Scholar]
- Sattari, M; Lowenthal, DT. Novel oral anticoagulant in development: Dabigatran, Rivaroxaban and Apixaban. Am. J. Ther. 2011, 18, 332–338. [Google Scholar]
- Reis, AMM; Marques, TC; Opitz, SP; Silva, AEBC; Gimenes, FRE; Teixeira, TCA; Lima, REF; Cassiani, SHB. Errors in medicine administration—profile of medicines: Knowing and preventing. Acta Paul. Enferm. 2010, 23, 181–186. [Google Scholar]
- Pirmohamed, M; James, S; Meakin, S; Green, C; Scott, AK; Walley, TJ; Farrar, K; Park, BK; Breckenridge, AM. Adverse drug reactions as cause of admission to hospital: Prospective analysis of 18,820 patients. Br. Med. J. 2004, 329, 15–19. [Google Scholar]
- Lopes, RD. Antiplatelet agents in cardiovascular disease. J. Thromb. Thrombol. 2011, 31, 306–309. [Google Scholar]
- Moura, LA; Bianco, EM; Pereira, RC; Teixeira, VL; Fuly, AL. Anticoagulation and antiplatelet effects of a dolastane diterpene isolated from the marine brown alga Canistrocarpus cervicornis. J. Thromb. Thrombol. 2001, 31, 235–240. [Google Scholar]
- Nutescu, EA; Shapiro, NL; Chevalier, A; Amin, AN. A pharmacological overview of current and emerging anticoagulants. Clev. Clin. J. Med. 2005, 72, 2–6. [Google Scholar]
- Costello, MJ; Coll, M; Danovaro, R; Halpin, P; Ojaveer, H; Milolasvich, P. A Census of Marine Biodiversity Knowledge, Resources, and Future Challenges. PLoS One 2010, 5, 12110. [Google Scholar]
- Usami, Y. Recent synthetic studies leading to structural revisions of marine natural products. Mar. Drugs 2009, 7, 314–330. [Google Scholar]
- Güven, KC; Percot, A; Sezik, E. Alkaloids in Marine Algae. Mar. Drugs 2010, 8, 269–284. [Google Scholar]
- Clavico, EEG; Muricy, G; Gama, BAP; Batista, D; Ventura, CRR; Pereira, RC. Ecological roles of natural products from the marine sponge Geodia corticostylifera. Mar. Biol. 2006, 148, 479–488. [Google Scholar]
- Teixeira, VL. Produtos Naturais Marinhos. In Biologia Marinha, 2nd ed; Pereira, RC, Soares-Gomes, A, Eds.; Editora Interciência: Rio de Janeiro, Brazil, 2002; pp. 443–472. [Google Scholar]
- Mayer, AMS; Rodrígez, AD; Berlink, RGS; Hamann, MT. Marine pharmacology in 2003–4: Marine compounds with anthelmintic, antibacterial, anticoagulant, antifungal, anti-inflammatory, antimalarial, antiplatelet, antiprotozoal, antituberculosis, and antiviral activities; affecting the cardiovascular, immune and nervous systems, and other miscellaneous mechanisms of action. Comp. Biochem. Physiol. 2007, 145, 553–581. [Google Scholar]
- Molinski, TF; Dalisay, DS; Lievens, SL; Saludes, JP. Drug development from marine natural products. Nat. Rev. Drug Discov. 2009, 8, 69–85. [Google Scholar]
- Cirne-Santos, CC; Teixeira, VL; Castello-Branco, LR; Frugulhetti, ICPP; Bou-Habib, DC. Inhibition of HIV-1 replication ih human primary cells by a dolabellane diterpene isolated from the marine algae Dictyota pfafii. Planta Med. 2006, 72, 295–299. [Google Scholar]
- Soares, AR; Abrantes, JL; Souza, TML; Leite Fontes, CF; Pereira, RC; Frugulhetti, ICPP; Teixeira, VL. In vitro antiviral effect of meroditerpenes isolated from the brazilian seawwed Stypopodium zonale (Dictyotales). Planta Med. 2007, 73, 1–4. [Google Scholar]
- Souza, TM; Abrantes, JL; Epifanio, RA; Leite Fontes, CF; Frugulhetti, ICPP. The alkaloid 4-methylaaptamine isolated from the sponge Aaptos aaptos impairs Herpes simplex virus type 1 penetration and immediate-early protein synthesis. Planta Med. 2007, 73, 200–205. [Google Scholar]
- Domingos, TFS; Carvalho, C; Moura, LA; Teixeira, VL; Pereira, RC; Bianco, EM; Ferreira, WJ; Ramos, CJB; de Miranda, ALP; Melo, PA; et al. Antilonomic effects of brazilian brown seaweeds extracts. Nat. Prod. Commun. 2009, 4, 1075–1078. [Google Scholar]
- Moura, LA; Sanchez, EF; Bianco, EM; Pereira, RC; Teixeira, VL; Fuly, AL. Antiophidian properties of a dolastane diterpene isolated from the marine brown alga Canistrocarpus cervicornis. Biomed. Pharmacother. 2010. [Google Scholar] [CrossRef]
- Azevedo, LG; Muccillo-Baisch, AL; Figueira, DMVB; Boyle, RT; Ramos, DF; Soares, AD; Lerner, C; Silva, PA; Trindade, GS. Comparative cytotoxic and anti-tuberculosis activity of Aplysina caissara marine sponge crude extracts. Comp. Biochem. Physiol. 2008, 147, 36–42. [Google Scholar]
- Cumashi, A; Ushakova, NA; Preobrazhenskaya, ME; D’Incecco, A; Piccoli, A; Totani, L; Tirani, N; Morozevich, GE; Berman, AE; Bilan, MI; et al. A comparative study of the anti-inflammatory, anticoagulant, antiangiogenic, and antiadhesive activities of nine different fucoidans from brown seaweeds. Glycobiology 2007, 17, 541–552. [Google Scholar]
- Albuquerque, IRL; Queiroz, KCS; Alves, LG; Santos, EA; Leite, EL; Rocha, HAO. Heterofucans from Dictyota menstrualis have anticoagulant activity. Braz. J. Med. Biol. Res. 2004, 37, 167–171. [Google Scholar]
- Wijesekara, I; Pangestuti, R; Kim, S. Biological activities and potential health benefits of sulfated polysaccharides derived from marine algae. Carbohydr. Polym. 2011, 84, 14–21. [Google Scholar]
- Matsubara, K. Recent advances in marine algal anticoagulants. Curr. Med. Chem. Cardiovasc. Hematol. Agents 2004, 2, 13–19. [Google Scholar]
- Mourão, PAS. Use of Sulfated fucans as anticoagulant and antithrombotic agents: Future Perspectives. Curr. Pharm. Des. 2004, 10, 967–981. [Google Scholar]
- Camara, RBG; Costa, LS; Fidelis, GP; Nobre, LTDB; Dantas-Santo, N; Cordeiro, SL; Costa, MSSP; Alves, LG; Rocha, HAO. Heterofucans from the brown seaweed Canistrocarpus cervicornis with anticoagulant and antioxidant activities. Mar. Drugs 2011, 9, 124–138. [Google Scholar]
- Lee, K; Kim, M. Antiplatelet and antithrombotic activities of Methanol Extract of Usnea longissima. Phytother. Res. 2005, 19, 1061–1064. [Google Scholar]
- Kim, M; Lee, K. Antithrombotic activity of methanolic extract of Umbilicaria esculenta. J. Ethnopharmacol. 2006, 105, 342–345. [Google Scholar]
- Carvalho, MJ; Pedrosa, TN; Guilhon-Simplicio, F; Nunez, CV; Ohana, DT; Pereira, MM; Lima, ES. Pharmacognostic study and in vitro activity on blood coagulation and platelet aggregation of leaves of Passifora nitida Kunth (Passiforaceae). Acta Amaz. 2010, 40, 199–206. [Google Scholar]
- Melo, FR; Mourão, PA. An algal saulfated galactan has unusual dual effect on venous thrombosis due to activation of factor XII and inhibition of the coagulation proteases. Thromb. Haemostasis 2008, 99, 531–538. [Google Scholar]
- De Azevedo, TCG; Bezerra, MEB; Santos, MGL; Souza, LA; Marques, CT; Benevides, NMB; Leite, EL. Heparinoids algal and their anticoagulant, hemorrhagic activities and platelet aggregation. Biomed. Pharmacother. 2008, 63, 477–483. [Google Scholar]
- Surin, WR; Barthwal, MK; Dikshit, M. Platelet collagen receptors, signaling and antagonism: Emerging approaches for the prevention of intravascular thrombosis. Thromb. Res. 2008, 122, 786–803. [Google Scholar]
- Clemetson, KJ; Clemetson, JM. Collagen receptors as potential targets for novel anti-platelet agents. Curr. Pharm. Des. 2007, 13, 2673–2683. [Google Scholar]
- Rivera, J; Lozano, ML; Navarro-Núñez, L; Vicente, V. Platelet receptors and signaling in the dynamics of thrombus formation. Haematologica 2009, 94, 700–711. [Google Scholar]
- Suzuki-Inoue, K; Fuller, GL; Garcia, A; Eble, JA; Pohlmann, S; Inoue, O; Gartner, TK; Hughan, SC; Pearce, AC; Laing, GD; et al. A novel Syk-dependent mechanism of platelet activation by the C-type lectin receptor CLEC-2. Blood 2006, 107, 542–549. [Google Scholar]
- Maita, N; Nishio, K; Nishimoto, E; Matsui, T; Shikamoto, Y; Morita, T; Sadler, JE; Mizuno, H. Crystal structure of von Willebrand factor A1 domain complexed with snake venom, bitiscetin: Insight into glycoprotein Ibalpha binding mechanism induced by snake venom proteins. J. Biol. Chem. 2003, 278, 37777–37781. [Google Scholar]
- Chen, ZM; Wu, JB; Zhang, Y; Yu, GY; Lee, WH; Lu, QM; Zhang, Y. Jerdonuxin, a novel snaclec (snake C-type lectin) with platelet aggregation activity from Trimeresurus jerdonii venom. Toxicon 2011, 57, 109–116. [Google Scholar]
- Ortiz-Ramirez, FA; Cavalcanti, DN; Villaça, RC; de Paula, JC; Valentin, YY; Teixeira, VL. Chemical variations in the diterpenes from the Brazilian brown alga Dictyota menstrualis (Dictyotaceae, Phaeophyta). Nat. Prod. Commun. 2008, 11, 1879–1884. [Google Scholar]
- Fuly, AL; de Miranda, AL; Zingali, RB; Guimarães, JA. Purification and characterization of a phospholipase A2 isoenzyme isolated from Lachesis muta snake venom. Biochem. Pharmacol 2002, 63, 1589–1597. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
| Samples | Clotting times (s) | |||
|---|---|---|---|---|
| RT | PT | aPTT | FC | |
| NaCl | 122.8 ± 16 | 21.7 ± 0.3 | 32.1 ± 1.2 | 26.4 ± 0.5 |
| DMSO | 149.7 ± 3.3 | 25.6 ± 0.2 | 38.8 ± 0.8 | 34.1 ± 1.4 |
| Dm-H-PF | 158.3 ± 11 | 24.5 ± 0.2 | 38.7 ± 2.0 | 46.8 ± 1.3 * |
| Dm-H-SP | 156.2 ± 8.3 | 25.1 ± 0.4 | 39.3 ± 1.3 | 47.3 ± 1.6 * |
| Dm-AC-PF | 172.3 ± 13 | 29.9 ± 0.6 * | 40.5 ± 1.1 | 86.4 ± 2.6 * |
| Dm-AC-SP | 155.6 ± 3.2 | 24.7 ± 1.0 | 39.1 ± 0.7 | 65.3 ± 3.5 * |
| Dm-DC | 171.6 ± 14 | 23.3 ± 0.2 | 40.2 ± 0.5 | 66.7 ± 0.8 * |
| Dc-H | 177.4 ± 13 | 27.3 ± 0.2 | 38.7 ± 2.4 | 42.7 ± 0.9 * |
| Dc-AC | 166.1 ± 4.1 | 28.9 ± 0.2 | 46.1 ± 2.5 * | 78.4 ± 11 * |
| Samples | Clotting times (s) | |||
|---|---|---|---|---|
| RT | PT | aPTT | FC | |
| NaCl | 124.3 ± 7.2 | 21.7 ± 0.3 | 32.1 ± 1.2 | 25.2 ± 0.1 |
| DMSO | 160.6 ± 11.5 | 22.7 ± 0.2 | 34.8 ± 1.3 | 34.4 ± 0.4 |
| Pc | 203.5 ± 4.7 * | 27.4 ± 1.0 * | 48.5 ± 1.4 * | 86.6 ± 5.8 * |
| Ma | 227.0 ± 8.0 * | 27.1 ± 0.7 * | 54.8 ± 2.0 * | 99.8 ± 9.6 * |
| Da | 210.6 ± 17.1 | 27.9 ± 1.2 * | 52.3 ± 1.7 * | 88.9 ± 6.9 * |
| De | 197.2 ± 10.6 | 26.3 ± 0.4 * | 54.1 ± 1.2 * | 101.6 ± 6.0 * |
| Ti | 246.0 ± 9.3 * | 27.3 ± 0.8 * | 64.2 ± 1.7 * | 176.2 ± 4.7 * |
| Av | 239.3 ± 14.8 * | 27.1 ± 0.6 * | 58.0 ± 1.6 * | 94.4 ± 5.4 * |
| Hh | 185.6 ± 10.1 | 23.9 ± 0.6 | 40.6 ± 1.1 * | 76.6 ± 3.1 * |
| Af | 234.6 ± 13.6 * | 25.7 ± 0.5 * | 45.2 ± 2.2 * | 75.9 ± 5.4 * |
| Ch | 296.3 ± 29.0 * | 27.5 ± 0.4 * | 53.2 ± 2.8 * | 92.6 ± 8.1 * |
| Pj | 278.6 ± 6.5 * | 26.3 ± 0.5 * | 52.2 ± 1.0 * | 107.1 ± 5.0 * |
| Algae | Solvent | Collection Site | Voucher | Code |
|---|---|---|---|---|
| Dictyota menstrualis | Ethyl Acetate | Arquipélago de São Pedro e São Paulo, RN (00°55′N–29°21′W) | 11006 | Dm-AC-SP |
| Dictyota menstrualis | Ethyl Acetate | Praia do Forno Armação de Búzios, RJ (22°45′S–41°52′W) | 84815 | Dm-AC-PF |
| Dictyota menstrualis | Hexane | Arquipélago de São Pedro e São Paulo, RN (00°55′N–29°21′W) | 11006 | Dm-H-SP |
| Dictyota menstrualis | Hexane | Praia do Forno Armação de Búzios, RJ (22°45′S–41°52′W) | 84815 | Dm-H-PF |
| Dictyota menstrualis | Dichloromethane | Praia Rasa Armação de Búzios, RJ (22°45′S–41°52′W) | 10017 | Dm-DC |
| Dictyota ciliolata | Hexane | Praia Vermelha Angra dos Reis, RJ (23°00′S–44°19′W) | 10415 | Dc-H |
| Dictyota ciliolata | Ethyl Acetate | Praia Vermelha Angra dos Reis, RJ (23°00′S–44°19′W) | 10415 | Dc-AC |
| Sponge | Solvent | Collection Site | Voucher | Code |
|---|---|---|---|---|
| Petromica citrina | Dichloromethane | Arquipélago das Cagarras Rio de Janeiro, RJ (23°01′S–43°13′W) | 14537 | Pc |
| Mycale angulosa | Acetone | Praia da Baleia Angra dos Reis, RJ (23°01′S–44°14′W) | 14529 | Ma |
| Chondrosia sp. | Acetone | Praia da Baleia Angra dos Reis, RJ (23°01′S–44°14′W) | 14551 | Ch |
| Dysidea etherea | Acetone | Praia da Baleia Angra dos Reis, RJ (23°01′S–44°14′W) | 14525 | De |
| Desmapsamma anchorata | Acetone | Ilha do Bonfim Angra dos Reis, RJ (23°01′S–44°19′W) | 14520 | Da |
| Amphimedon viridis | Acetone | Ilha do Bonfim. Angra dos Reis, RJ (23°01′S–44°19′W) | 14517 | Av |
| Tedania ignis | Acetone | Praia de Tarituba Paraty, RJ (23°02′S–44°35′W) | 14522 | Ti |
| Hymeniacidon heliophila | Acetone | Praia de Itaipu Niterói, RJ (22°58′S–43°02′W) | 14528 | Hh |
| Aplysina fulva | Acetone | Praia do Forno Arraial do Cabo, RJ (22°57′S–42°00′W) | 13554 | Af |
| Polymastia janeirensis | Acetone | Praia do Forno Arraial do Cabo, RJ (22°57′S–42°00′W) | 14512 | Pj |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Moura, L.d.A.; Ortiz-Ramirez, F.; Cavalcanti, D.N.; Ribeiro, S.M.; Muricy, G.; Teixeira, V.L.; Fuly, A.L. Evaluation of Marine Brown Algae and Sponges from Brazil as Anticoagulant and Antiplatelet Products. Mar. Drugs 2011, 9, 1346-1358. https://doi.org/10.3390/md9081346
Moura LdA, Ortiz-Ramirez F, Cavalcanti DN, Ribeiro SM, Muricy G, Teixeira VL, Fuly AL. Evaluation of Marine Brown Algae and Sponges from Brazil as Anticoagulant and Antiplatelet Products. Marine Drugs. 2011; 9(8):1346-1358. https://doi.org/10.3390/md9081346
Chicago/Turabian StyleMoura, Laura de Andrade, Fredy Ortiz-Ramirez, Diana Negrao Cavalcanti, Suzi Meneses Ribeiro, Guilherme Muricy, Valeria Laneuville Teixeira, and Andre Lopes Fuly. 2011. "Evaluation of Marine Brown Algae and Sponges from Brazil as Anticoagulant and Antiplatelet Products" Marine Drugs 9, no. 8: 1346-1358. https://doi.org/10.3390/md9081346
APA StyleMoura, L. d. A., Ortiz-Ramirez, F., Cavalcanti, D. N., Ribeiro, S. M., Muricy, G., Teixeira, V. L., & Fuly, A. L. (2011). Evaluation of Marine Brown Algae and Sponges from Brazil as Anticoagulant and Antiplatelet Products. Marine Drugs, 9(8), 1346-1358. https://doi.org/10.3390/md9081346