

Role of Enzyme Technologies and Applied Enzymology in Valorising Seaweed Bioproducts

 ,
,  ,
, 
Abstract
1. Introduction
2. Background to Seaweeds
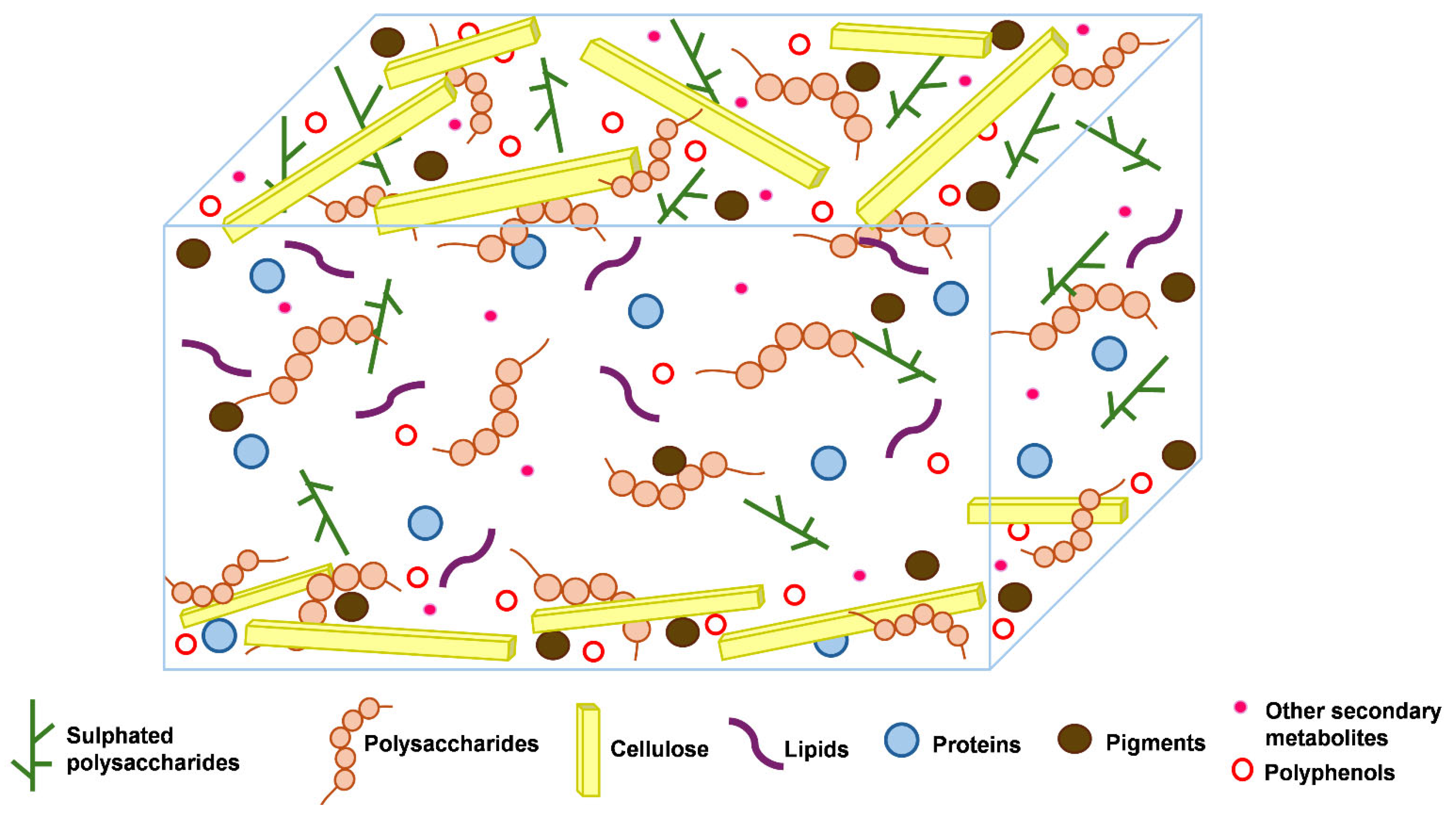
2.1. Structures of Seaweeds
2.2. Applications of Seaweeds and Their Bioproducts

{kind=link}
| Seaweed Type | Sector | Bioproducts | Applications | References |
|---|---|---|---|---|
| Brown | Pharmaceutical | Fucoidan, polyphenols/phlorotannins | Antidiabetes, anticancer, antioxidant, antiviral | [27,28,29] |
| Nutraceutical | Fucoxanthin & alginates | Antioxidants, weight management & immune boosters | [25] | |
| Agricultural | Alginates, growth hormones | Biostimulants & soil conditioners | [26] | |
| Cosmetic | Alginates & phlorotannins | Anti-ageing creams, moisturisers & sunscreens | [30] | |
| Red | Pharmaceutical | Carrageenan & agar | Drug delivery systems, anticoagulants & antiviral agents | [31,32] |
| Nutraceutical | Agar & protein extracts | Dietary fibre, prebiotics & functional foods | [33] | |
| Agricultural | Carrageenan & whole extracts | Plant growth promoters | [34] | |
| Green | Pharmaceutical | Ulvan, polyphenols | Antioxidants, immunomodulatory, & antibacterial agents | [35,36] |
| Nutraceutical | Polyphenols and protein extracts | Functional foods, anti-inflammatory supplements | [37] | |
| Agricultural | Whole extracts | Biofertilisers, plant defense enhancers | [38] |
3. Marine Bioproducts Inhibiting Enzymes Involved in Human Diseases
| Seaweed Bioproduct | Source (Seaweed Type) | Target Enzyme | Disease Association | Mechanism of Action | Biological Effect | Reference |
|---|---|---|---|---|---|---|
| Fucoidan | Ecklonia radiata | α-Amylase, α-glucosidase | Type 2 Diabetes, Obesity | Inhibits carbohydrate hydrolysis, reducing postprandial glucose levels | α-Glucosidase: IC50 19 μg/mL. | [52] |
| Sargassum binderi | α-Glucosidase, pancreatic lipase | Type 2 Diabetes, Obesity | Prevents fat digestion and glucose absorption | α-Glucosidase (IC50 = 174.63 ± 23.94 μg/mL) | [57] | |
| Padina tetrastromatica | 3C-like protease (3CLpro), | Coronavirus | Disrupts coronavirus replication and blocks viral entry, collectively hindering infection and propagation | 3CLpro: IC50 0.37 mg/mL | [58] | |
| Laminarin | Laminaria digitata and Fucus vesiculosus | Cyclooxygenase-1 (COX-1) and cyclooxygenase-2 (COX-2) | Inflammation | Preventing tissue damage, modulating immune responses, and alleviating inflammatory conditions | Crude: IC50 = 48.96 µg/mL (COX-1) and IC50 = 42.74 µg/mL (COX-2) | [59] |
| Laminaria digitata and Fucus vesiculosus | Dipeptidyl Peptidase-IV (DPP-IV) | Type 2 Diabetes | Enhances insulin secretion by inhibiting DPP-IV | Crude laminarin: 46.84% inhibition at 1 mg/mL | [59] | |
| Carrageenan Oligosaccharides | Eucheuma cottonii | α-Amylase | Type 2 Diabetes | Lowers glucose absorption | α-amylase: 59.33% inhibition at 1000 ppm | [60] |
| Ulvan | Ulva lactuca | Angiotensin-converting enzyme (ACE) | Obesity and Hypertension | Reduce vasoconstriction and lower blood pressure and prevent fat breakdown and absorption for obesity | Ulvan oligosaccharides: 50.18% inhibition | [61] |
| Ulva species | α-Glucosidase | Type 2 Diabetes | Lowers glucose absorption | Completely hydrolysed: IC50 = 2.51 ± 0.19 mg/mL | ||
| Polyphenols | Phyllospora comosa | α-Amylase, ACE, and pancreatic lipase | Diabetes, Obesity and Hypertension | Lowers glucose absorption. Reduces vasoconstriction and lowers blood pressure and prevents fat breakdown and absorption for obesity | ACE: IC50 = 583.76 ± 9.42 α-Amylase: IC50 = 58.31 ± 1.41 μg/mL Pancreatic lipase: IC50 = 52.14 ± 2.77 μg/mL | [55] |
| Bioactive Peptides (Hydrolyzed seaweed proteins) | Phyllospora comosa and Ulva ohnoi | α-Amylase, ACE, and pancreatic lipase | Diabetes, Obesity and Hypertension | Lowers glucose absorption. Reduces vasoconstriction, lowers blood pressure and prevents fat breakdown and absorption for obesity | ACE: IC50 = 167.52 ± 3.17 μg/mL (U. ohnoi) α-Amylase: IC50 = 423.39 ± 18.60 μg/mL (U. ohnoi) Pancreatic lipase: IC50 = 742.48 ± 30.37 μg/mL (P. comosa) | [55] |
| Sterols | Sargassum horridum | Acetylcholinesterase (AChE) | Alzheimer’s Disease | Prevents acetylcholine breakdown, improving cognitive function | Potent non-competitive inhibition of AChE | [62] |
4. Enzyme-Assisted Extractions (EAEs)
| Seaweed Species | Bioactive Compound | Extraction Conditions | Enzyme | Yield (Dry Weight) | Bioactivity | Reference |
|---|---|---|---|---|---|---|
| Kjellmaniella crassifolia | Fucoidan | 50 mM citrate buffer (pH 4.8) at 50 °C for 10 h | Cellulase and β-glucosidase (ratio: 1:4.29) | 4.74% | Antioxidant activity | [69] |
| Cystoseira myrica | Fucoidan | Digestion with papain for 24 h at 60 °C | Papain (100 mg) | 3.07% | Antioxidant and anticoagulant activities | [70] |
| Padina arborescens | Fucoidan | Deionized water, pH (4.5) at 50 °C, shaking for 24 h | Celluclast (1%, mg/g of biomass) | 26% | Anti-inflammatory | [71] |
| Fucus vesiculosus | Plant growth regulators, neutral and reducing sugars and phenolics | Deionized water, enzymes 5% (enzyme/seaweed ratio (w/w)) at 50 °C, shaking for 17 h | β-glucosidase, β-1,3-glucanase, Botrytis glucanase, and protease (neutral, endo) | Reducing sugars (21%), neutral sugars (10%) | Antioxidant activity | [72] |
| Gracilaria dura | Protein extracts | Citrate buffer solution (0.1 M, pH 4.5), shaking at 35 °C for 20 h | Hemicellulase | 212.57 mg/g (protein/extract) | Emulsifying and antioxidant activities | [73] |
| Porphyra dioica | Peptides | Deionized water, pH (8.0) at 50 °C, shaking for 120 min | Prolyve 1000® and Flavourzyme® | Protein content (25.65%) | Antioxidant activity | [74] |
| Ulva rigida | Ulvan | Deionized water, pH (8.0) at 50 for 4 h | Alcalase | Not specified | Food preservative | [75] |
| Ulva fenestrata | Ulvan | 0.1 M sodium acetate buffer (pH 5.0) and 50 °C (Viscozyme L) or 40 °C (Cellulysin), 0.1 M Tris HCl buffer (pH 7.0) and 60 °C (Neutrase) and pH 5.0 at 50 °C (Flavourzyme) for 20 h | Viscozyme L, Cellulysin, Neutrase and Flavourzyme | 14.1% with Cellulysin | Not specified | [76] |
| Fucus vesiculosus | Fucoxanthin | 0.1 M Sodium acetate buffer (pH 4.5), 50 °C, shaking for 12 h | Viscozyme | 0.657 mg/g | Not specified | [77] |
| Turbinaria conoides and Padina tetrastromatica | Phenolic compounds | Viscozyme in acetate buffer (pH 4.5), flavourzyme in phosphate buffer (pH 7.0), alcalase in phosphate buffer (pH 8.0), 50 °C for 20 h | Carbohydrases and proteases | 73.3% yield and 3.15 mg gallic acid equivalents/g extract | Antioxidant activities | [78] |
5. Challenges in the Enzyme-Assisted Extractions of Seaweed Bioproducts
6. Advancements Mitigating Enzyme-Assisted Challenges
6.1. Metagenomics
6.2. Enzyme Engineering
6.3. Enzyme Immobilisation
6.4. Dual Valorisation Impact of Immobilised Enzymes and Seaweed Bioproducts
7. Circular Bioeconomy
Enzyme-Assisted Biorefineries for Seaweeds
8. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Accelerated solvent extraction | ASE |
| Angiotensin-converting enzyme | ACE |
| Cyclooxygenase | COX |
| Dipeptidyl-peptidase-4 | DPP-IV |
| Enzyme-assisted extraction | EAE |
| Microwave-assisted extraction | MAE |
| Protein tyrosine phosphatase 1B | PTP1B |
| Reactive oxygen species | ROS |
| Supercritical fluid extraction | SFE |
| Sustainable Development Goals | SDGs |
| Type 2 diabetes Mellitus | T2DM |
| Ultrasound-assisted extraction | UAE |
References
- Ashkenazi, D.Y.; Figueroa, F.L.; Korbee, N.; García-Sánchez, M.; Vega, J.; Ben-Valid, S.; Paz, G.; Salomon, E.; Israel, Á.; Abelson, A. Enhancing Bioproducts in Seaweeds via Sustainable Aquaculture: Antioxidant and Sun-Protection Compounds. Mar. Drugs. 2022, 20, 767. [Google Scholar] [CrossRef]
- Layn, J.; Farkan, M.; Sektiana, S. Strategy for Developing a Sustainable Seaweed Cultivation Industry in The Kotania Bay Area, Maluku. Int. J. Progress. Sci. Technol. 2023, 41, 254–268. [Google Scholar] [CrossRef]
- Sharma, U.; Jadaun, S.; Khapudaang, R.; Siddiqui, S. Seaweed- A Sustainable Food Source in the Food Industry. In Sustainable Food Systems (Volume II); Thakur, M., Ed.; World Sustainability Series; Springer: Cham, Switzerland, 2024. [Google Scholar] [CrossRef]
- Nesic, A.; Meseldzija, S.; Benavides, S.; Figueroa, F.A.; Cabrera-Barjas, G. Seaweed as a Valuable and Sustainable Resource for Food Packaging Materials. Foods 2024, 13, 3212. [Google Scholar] [CrossRef]
- Daub, C.D.; Michaels, A.L.; Mabate, B.; Mkabayi, L.; Edkins, A.L.; Pletschke, B.I. Exploring the Inhibitory Potential of Sodium Alginate Against Digestive Enzymes Linked to Obesity and Type 2 Diabetes. Molecules 2025, 30, 1155. [Google Scholar] [CrossRef]
- Cho, C.H.; Lu, Y.A.; Kim, M.Y.; Jeon, Y.J.; Lee, S.H. Therapeutic Potential of Seaweed-Derived Bioactive Compounds for Cardiovascular Disease Treatment. Appl. Sci. 2022, 12, 1025. [Google Scholar] [CrossRef]
- Khairinisa, M.A.; Latarissa, I.R.; Athaya, N.S.; Charlie, V.; Musyaffa, H.A.; Prasedya, E.S.; Puspitasari, I.M. Potential Application of Marine Algae and Their Bioactive Metabolites in Brain Disease Treatment: Pharmacognosy and Pharmacology Insights for Therapeutic Advances. Brain Sci. 2023, 13, 1686. [Google Scholar] [CrossRef]
- Mabate, B.; Pletschke, B.I. Sequential and enzyme-assisted extraction of algal bioproducts from Ecklonia maxima. Enzym. Microb. Technol. 2024, 173, 110364. [Google Scholar] [CrossRef]
- Gao, K.; McKinley, K.R. Use of macroalgae for marine biomass production and CO2 remediation: A review. J. Appl. Phycol. 1994, 60, 45–60. [Google Scholar] [CrossRef]
- Zhang, L.; Liao, W.; Huang, Y. Global seaweed farming and processing in the past 20 years. Food Prod. Process Nutr. 2022, 4, 23. [Google Scholar] [CrossRef]
- Ahmad, T.B.S. Methods for Quantification and Extraction of Fucoidan, and Quantification of the Release of Total Carbohydrate and Fucoidan from the Brown Algae Laminaria hyperborea. Master’s Thesis, NTNU, Ålesund, Norway, 2015. [Google Scholar]
- Begum, R.H.S.; Mamun-Or-Rashid, A.N.M.; Rafiquzzaman, S.M.; Ashraf, G.M.; Albadrani, G.M.; Sayed, A.A.; Peluso, I.; Abdel-Daim, M.M.; Uddin, M.S. Antioxidant and Signal-Modulating Effects of Brown Seaweed-Derived Compounds against Oxidative Stress-Associated Pathology. Oxid. Med. Cell. Longev. 2021, 2021, 9974890. [Google Scholar] [CrossRef]
- Eladl, S.N.; Elnabawy, A.M.; Eltanahy, E.G. Recent biotechnological applications of value-added bioactive compounds from microalgae and seaweeds. Bot. Stud. 2024, 65, 28. [Google Scholar] [CrossRef]
- Menaa, F.; Wijesinghe, P.A.U.I.; Thiripuranathar, G.; Uzair, B.; Iqbal, H.; Khan, B.A.; Menaa, B. Ecological and Industrial Implications of Dynamic Seaweed-Associated Microbiota Interactions. Mar. Drugs. 2020, 18, 641. [Google Scholar] [CrossRef] [PubMed]
- Shao, Z.; Duan, D. The Cell Wall Polysaccharides Biosynthesis in Seaweeds: A Molecular Perspective. Front. Plant Sci. 2022, 13, 902823. [Google Scholar] [CrossRef] [PubMed]
- Deniaud-Bouët, E.; Kervarec, N.; Michel, G.; Tonon, T.; Kloareg, B.; Hervé, C. Chemical and enzymatic fractionation of cell walls from Fucales: Insights into the structure of the extracellular matrix of brown algae. Ann. Bot. 2014, 114, 1203–1216. [Google Scholar] [CrossRef]
- Salmeán, A.; Duffieux, D.; Harholt, J.; Qin, F.; Michel, G.; Czjzek, M.; Willats, W.G.T.; Hervé, C. Insoluble (1 → 3), (1 → 4)-β-D-glucan is a component of cell walls in brown algae (Phaeophyceae) and is masked by alginates in tissues. Sci. Rep. 2017, 7, 2880. [Google Scholar] [CrossRef]
- Synytsya, A.; Čopíková, J.; Kim, W.J.; Park, Y.I. Cell Wall Polysaccharides of Marine Algae. In Springer Handbook of Marine Biotechnology; Kim, S.K., Ed.; Springer Handbooks; Springer: Berlin/Heidelberg, Germany, 2015; pp. 543–590. [Google Scholar] [CrossRef]
- Demir-Yilmaz, I.; Schiavone, M.; Esvan, J.; Guiraud, P.; Formosa-Dague, C. Combining AFM, XPS and chemical hydrolysis to understand the complexity and dynamics of C. vulgaris cell wall composition and architecture. Algal Res. 2023, 72, 103102. [Google Scholar] [CrossRef]
- Mukhia, D.J.; Chatterjee, R.; Biswal, S.; Rai, S.; Rai, U.; Kumar, R.; Rai, S. Sea Vegetables: An Alternative Food Source: A Review. Agric. Rev. 2024, 2665, 1–9. [Google Scholar] [CrossRef]
- Abbott, I.A. The Uses of Seaweed as Food in Hawaii. Econ. Bot. 1978, 32, 409–412. [Google Scholar] [CrossRef]
- Pérez-Lloréns, J.L.; Critchley, A.T.; Cornish, M.L. Traditional knowledge, home remedies, medicine, surgery, and pharmacopoeia. J. Appl. Phycol. 2023, 35, 2049–2068. [Google Scholar] [CrossRef]
- Bennett, J.P.; Robinson, L.F.; Gomez, L.D. Valorisation strategies for brown seaweed biomass production in a European context. Algal Res. 2023, 75, 103248. [Google Scholar] [CrossRef]
- Mooney, D.E. Charred Fucus-Type Seaweed in the North Atlantic: A Survey of Finds and Potential Uses. Environ. Archaeol. 2018, 26, 238–250. [Google Scholar] [CrossRef]
- Morparia, S.; Suvarna, V. Recent Advancements in Applications of Alginates in Drug Delivery, Tissue Engineering, and Biomedical Field. J. Nat. Prod. 2024, 14, e260124226231. [Google Scholar] [CrossRef]
- Zabihi, M. Applications of Alginate in the Fields of Research Medicine, Industry and Agriculture. In Alginate—Applications and Future Perspectives; Severo, I.A., Mariano, A.B., Vargas, J.V.C., Eds.; IntechOpen: London, UK, 2024. [Google Scholar] [CrossRef]
- Mabate, B.; Daub, C.D.; Malgas, S.; Pletschke, B.I. Characterisation of Sargassum elegans fucoidans extracted using different technologies: Linking their structure to α-glucosidase inhibition. Algal Res. 2025, 85, 103885. [Google Scholar] [CrossRef]
- Mabate, B.; Daub, C.D.; Pletschke, B.I.; Edkins, A.L. Comparative Analyses of Fucoidans from South African Brown Seaweeds That Inhibit Adhesion, Migration, and Long-Term Survival of Colorectal Cancer Cells. Mar. Drugs. 2023, 21, 203. [Google Scholar] [CrossRef]
- Ghahtan, N.; Dehghan, N.; Ullah, M.; Khoradmehr, A.; Habibi, H.; Nabipour, I.; Baghban, N. From seaweed to healing: The potential of fucoidan in wound therapy. Nat. Prod. Res. 2024, 39, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, P.F.; Murillo-González, L.; Rodríguez, E.; Pérez, A.M. Marine phenolic compounds: Sources, commercial value, and biological activities. In Marine Phenolic Compounds; Pérez-Correa, J.R., Mateos, R., Domínguez, H., Eds.; Elsevier: New York, NY, USA, 2023; pp. 47–86. [Google Scholar] [CrossRef]
- Cotas, J.; Leandro, A.; Pacheco, D.; Gonçalves, A.M.M.; Pereira, L. A Comprehensive Review of the Nutraceutical and Therapeutic Applications of Red Seaweeds (Rhodophyta). Life 2020, 10, 19. [Google Scholar] [CrossRef]
- Ismail, M.M.; Alotaibi, B.S.; EL-Sheekh, M.M. Therapeutic Uses of Red Macroalgae. Molecules 2020, 25, 4411. [Google Scholar] [CrossRef]
- Jiménez-González, A.C.; Torrado, A.M.; Federico, M.; Rúa, M.L.; Clara, F. Red seaweed proteins: Valuable marine-origin compounds with encouraging applications. Algal Res. 2023, 75, 103262. [Google Scholar] [CrossRef]
- Nivetha, N.; Shukla, P.S.; Nori, S.S.; Kumar, S.; Suryanarayan, S. A red seaweed Kappaphycus alvarezii-based biostimulant (AgroGain®) improves the growth of Zea mays and impacts agricultural sustainability by beneficially priming rhizosphere soil microbial community. Front. Microbiol. 2024, 15, 1330237. [Google Scholar] [CrossRef] [PubMed]
- Amlanı, M.; Yetgin, S. Seaweeds: Bioactive Components and Properties, Potential Risk Factors, Uses, Extraction and Purification Methods. Mar. Sci. Bull. 2022, 11, 9–31. [Google Scholar] [CrossRef]
- Sivaramakrishnan, T.; Biswas, L.; Baby Shalini, K.S.; Sankar, R.K.; Bharathi, M.P.G.; Roy, D. Analysis of Proximate Composition and In-Vitro Antibacterial Activity of Selected Green Seweeds from South Andaman Coast of India. Int. J. Curr. Microbiol. App. Sci. 2017, 12, 1739–1749. [Google Scholar] [CrossRef]
- Das, S.; Behera, M.; Das, S.R.; Behera, K.C.; Singh, L. Green Seaweeds as a Potential Source of Biomolecules and Bioactive Peptides: Recent Progress and Applications—A Review. Chem. Biodivers. 2024, 20, e202401695. [Google Scholar] [CrossRef]
- van der Weide, R.Y.; Elissen, H.J.H.; Hol, S.J.E.; Evenhuis, A.; de Vos, R.C.H.; van der Meer, I.M.; van Geel, W.C.A. Agricultural Applications of Seaweed Extracts: Seaweeds for Plant Care: Review and Experiments in The Netherlands; Report Wageningen Plant Research, OT Team Schimmels Onkr. en PlagenBioscienceEPSOT Team Bedrijfssyst.onderz./Bodemkwaliteit: Wageningen, The Netherlands, 2022. [Google Scholar]
- Daub, C.D.; Mabate, B.; Malgas, S.; Pletschke, B.I. Fucoidan from Ecklonia maxima is a powerful inhibitor of the diabetes-related enzyme, α-glucosidase. Int. J. Biol. Macromol. 2020, 151, 412–420. [Google Scholar] [CrossRef]
- Duan, X.; Agar, O.T.; Barrow, C.J.; Dunshea, F.R.; Suleria, H.A.R. Improving potential strategies for biological activities of phlorotannins derived from seaweeds. Crit. Rev. Food Sci. Nutr. 2023, 65, 833–835. [Google Scholar] [CrossRef]
- Pradhan, B.; Ki, S. Antioxidant and chemotherapeutic efficacies of seaweed-derived phlorotannins in cancer treatment: A review regarding novel anticancer drugs. Phytother. Res. 2023, 35, 2067–2091. [Google Scholar] [CrossRef] [PubMed]
- Maheswari, V.; Babu, P.A. Phlorotannin and its Derivatives, a Potential Antiviral Molecule from Brown Seaweeds, an Overview. Russ. J. Mar. Biol. 2022, 48, 309–324. [Google Scholar] [CrossRef] [PubMed]
- Pangestuti, R.; Ridwanudin, A.; Putra, Y.; Prathama, I.S.; Rahmadi, P.; Siahaan, E.A.; Kim, S.K. Valuable bioproducts from seaweeds obtained by green extraction technologies: Potential health benefits and applications in pharmacological industries. In Algae and Aquatic Macrophytes in Cities; Pandey, V.C., Ed.; Elsevier: New York, NY, USA, 2022; pp. 315–347. [Google Scholar] [CrossRef]
- Wang, X.; He, L.; Ma, Y.; Huan, L.; Wang, Y.; Xia, B.; Wang, G. Economically important red algae resources along the Chinese coast: History, status, and prospects for their utilization. Algal Res. 2020, 46, 101817. [Google Scholar] [CrossRef]
- Fradinho, P.; Raymundo, A.; Sousa, I.; Domínguez, H.; Torres, M.D. Edible Brown Seaweed in Gluten-Free Pasta: Technological and Nutritional Evaluation. Foods 2019, 8, 622. [Google Scholar] [CrossRef]
- Shan, X.; Liu, X.; Hao, J.; Cai, C.; Fan, F.; Dun, Y.; Zhao, X.; Liu, X.; Li, C.; Yu, G. In vitro and in vivo hypoglycemic effects of brown algal fucoidans. Int. J. Biol. Macromol. 2016, 82, 249–255. [Google Scholar] [CrossRef]
- Snarska, J.; Cieślińska, A.; Fiedorowicz, E.; Jarmołowska, B.; Sienkiewicz-Szłapka, E.; Matysiewicz, M.; Kiper, K.; Kostyra, E. Polymorphism in DPPIV Gene in Acute Pancreatitis. Pancreas 2017, 49, e71–e72. [Google Scholar] [CrossRef]
- Calderwood, D.; Rafferty, E.; Fitzgerald, C.; Stoilova, V.; Wylie, A.; Gilmore, B.F.; Castaneda, F.; Israel, A.; Maggs, C.A.; Green, B.D. Profiling the activity of edible European macroalgae towards pharmacological targets for type 2 diabetes mellitus. Appl. Phycol. 2021, 2, 10–21. [Google Scholar] [CrossRef]
- Kaur, K.K.; Allahbadia, G.; Singh, M. Development of protein tyrosine phosphatase 1B (PTPIB) Inhibitors from marine sources and other natural products-Future of Antidiabetic Therapy: A Systematic Review. Korean J. Food Health Converg. 2019, 5, 21–33. [Google Scholar] [CrossRef]
- Dhara, S.; Chakraborty, K. Turbinafuranone A–C, new 2-furanone analogues from marine macroalga Turbinaria ornata as prospective anti-hyperglycemic agents attenuate tyrosine phosphatase-1B. Med. Chem. Res. 2021, 30, 1635–1648. [Google Scholar] [CrossRef]
- Truong, T.P.T.; Tran, T.M.; Dai, T.X.T.; Tran, C.L. Antihyperglycemic and anti-type 2 diabetic activity of marine hydroquinone isolated from brown algae (Dictyopteris polypodioides). J. Tradit. Complement. Med. 2023, 13, 408–416. [Google Scholar] [CrossRef]
- Mabate, B.; Daub, C.D.; Malgas, S.; Edkins, A.L.; Pletschke, B.I. A Combination Approach in Inhibiting Type 2 Diabetes-Related Enzymes Using Ecklonia radiata Fucoidan and Acarbose. Pharmaceutics 2021, 13, 1979. [Google Scholar] [CrossRef]
- Garza, A.L.; de la, M.F.I.; Boque, N.; Campión, J.; Martínez, J.A. Natural inhibitors of pancreatic lipase as new players in obesity treatment. Planta Medica 2011, 77, 773–785. [Google Scholar] [CrossRef]
- Ferrari, R.; Fox, K. Insight into the Mode of Action of ACE Inhibition in Coronary Artery Disease. Drugs 2009, 69, 265–277. [Google Scholar] [CrossRef]
- Shannon, E.; Conlon, M.M. In vitro enzyme inhibitory effects of green and brown Australian seaweeds and potential impact on metabolic syndrome. J. Appl. Phycol. 2023, 35, 893–910. [Google Scholar] [CrossRef]
- Murakami, S.; Yoshikawa, R.; Ohya, T.; Ma, N.; Owaki, Y.; Owaki, T.; Ito, T.; Matsuzaki, C. The anti-obesity and anti-diabetic effects of the edible seaweed Gloiopeltis furcata (Postels et Ruprecht) J. Agardh in mice fed a high-fat diet. Food Sci. Nutr. 2023, 11, 599–610. [Google Scholar] [CrossRef]
- Din, N.A.S.; Ishak, A.A.; Maarof, S.; Abdul Rahman, H.; Sofian-Seng, N.S.; Mohd Razali, N.S.; Lim, S.J.; Wan Mustapha, W.A. Potential anti-obesity effects of fucoidan from Malaysian brown seaweed (Sargassum binderi) by in vivo study. Int. J. Food Sci. Technol. 2024, 59, 3402–3411. [Google Scholar] [CrossRef]
- Hans, N.; Gupta, S.; Patel, A.K.; Naik, S.; Malik, A. Deciphering the role of fucoidan from brown macroalgae in inhibiting SARS-CoV-2 by targeting its main protease and receptor binding domain: Invitro and insilico approach. Int. J. Biol. Macromol. 2023, 248, 125950. [Google Scholar] [CrossRef] [PubMed]
- Karuppusamy, S.; Wanigasekara, J.; Fitzpatrick, S.; Lyons, H.; Curtin, J.; Rajauria, G.; Tiwari, B.K.; O’Donnell, C. Investigation of Biological Activity of Fucoidan and Laminarin as Bioactive Polysaccharides from Irish Brown Macroalgae. Cells 2024, 13, 1938. [Google Scholar] [CrossRef] [PubMed]
- Prasasty, V.D.; Haryani, B.; Hutagalung, R.A.; Mulyono, N.; Yazid, F.; Rosmalena, R.; Sinaga, E. Evaluation of antioxidant and antidiabetic activities from red seaweed (Eucheuma cottonii). Sys. Rev. Pharm. 2019, 10, 276–288. [Google Scholar] [CrossRef]
- Hung, Y.H.R.; Chen, G.W.; Pan, C.L.; Lin, H.T.V. Production of Ulvan Oligosaccharides with Antioxidant and Angiotensin-Converting Enzyme-Inhibitory Activities by Microbial Enzymatic Hydrolysis. Fermentation 2021, 7, 160. [Google Scholar] [CrossRef]
- Castro-Silva, E.S.; Bello, M.; Hernández-Rodríguez, M.; Correa-Basurto, J.; Murillo-Álvarez, J.I.; Rosales-Hernández, M.C.; Muñoz-Ochoa, M. In vitro and in silico evaluation of fucosterol from Sargassum horridum as potential human acetylcholinesterase inhibitor. J. Biomol. Struct. Dyn. 2019, 37, 3259–3268. [Google Scholar] [CrossRef]
- Sanjeewa, K.K.A.; Herath, K.H.I.N.M.; Kim, Y.; Jeon, Y.; Kim, S. Enzyme-assisted extraction of bioactive compounds from seaweeds and microalgae. TrAC Trends Anal. Chem. 2023, 167, 117266. [Google Scholar] [CrossRef]
- Olivares-Molina, A.; Fernández, K. Comparison of different extraction techniques for obtaining extracts from brown seaweeds and their potential effects as angiotensin I-converting enzyme (ACE) inhibitors. J. Appl. Phycol. 2016, 28, 1295–1302. [Google Scholar] [CrossRef]
- Choulot, M.; Michalak, I.; Jing, L.; Szymczycha-Madeja, A.; Wełna, M.; Bourgougnon, N.; Le Guillard, C. The Enzyme-assisted extraction of compounds of interest in agriculture: Case study of the red seaweed Solieria chordalis (C. agardh) J. Agard. Algal Res. 2023, 75, 103239. [Google Scholar] [CrossRef]
- Nguyen, H.C.; Ngo, K.N.; Tran, H.K.; Barrow, C.J. Enzyme-Assisted Coextraction of Phenolics and Polysaccharides from Padina gymnospora. Mar. Drugs 2024, 22, 42. [Google Scholar] [CrossRef]
- Rhein-Knudsen, N.; Reyes-Weiss, D.; Horn, S.J. Extraction of high purity fucoidans from brown seaweeds using cellulases and alginate lyases. Int. J. Biol. Macromol. 2023, 229, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Karabulut, G.; Purkiewicz, A.; Goksen, G. Recent developments and challenges in algal protein and peptide extraction strategies, functional and technological properties, bioaccessibility, and commercial applications. Compr. Rev. Food Sci. Food Saf. 2024, 23, e13372. [Google Scholar] [CrossRef]
- Tang, S.; Ma, Y.; Dong, X.; Zhou, H.; He, Y.; Ren, D.; Wang, Q.; Yang, H.; Liu, S.; Wu, L. Enzyme-assisted extraction of fucoidan from Kjellmaniella crassifolia based on kinetic study of enzymatic hydrolysis of algal cellulose. Algal Res. 2022, 66, 102795. [Google Scholar] [CrossRef]
- Dhahri, M. Cystoseira myrica: From beach-cast seaweed to fucoidan with antioxidant and anticoagulant capacity. Front. Mar. Sci. 2023, 10, 1327408. [Google Scholar] [CrossRef]
- Lee, H.G.; Liyanage, N.M.; Yang, F.; Kim, Y.S.; Lee, S.H.; Ko, S.C.; Yang, H.W.; Jeon, Y.J. Investigation of physical characteristics and in vitro anti-inflammatory effects of fucoidan from Padina arborescens A comprehensive assessment against lipopolysaccharide-induced inflammation. Mar. Drugs 2024, 22, 109. [Google Scholar] [CrossRef]
- Choulot, M.; Jabbour, C.; Burlot, A.S.; Jing, L.; Welna, M.; Szymczycha-Madeja, A.; Le Guillard, C.; Michalak, I.; Bourgougnon, N. Application of enzyme-assisted extraction on the brown seaweed Fucus vesiculosus Linnaeus (Ochrophyta, Fucaceae) to produce extracts for a sustainable agriculture. J. Appl. Phycol. 2024, 37, 1325–1340. [Google Scholar] [CrossRef]
- Bozdemir, A.; Şensu, E.; Okudan, E.Ş.; Özçelik, B.; Yücetepe, A. Ultrasound-assisted enzymatic extraction of proteins from Gracilaria dura: Investigation of antioxidant activity and techno-functional properties. J. Food Process. Preserv. 2022, 46, e16803. [Google Scholar] [CrossRef]
- Pimentel, F.B.; Machado, M.; Cermeño, M.; Kleekayai, T.; Machado, S.; Rego, A.M.; Abreu, M.H.; Alves, R.C.; Oliveira, M.B.P.; FitzGerald, R.J. Enzyme-assisted release of antioxidant peptides from Porphyra dioica Conchocelis. Antioxidants 2021, 10, 249. [Google Scholar] [CrossRef]
- Alboofetileh, M.; Jeddi, S.; Mohammadzadeh, B.; Noghani, F.; Kamali, S. Exploring the impact of enzyme-extracted Ulvan from Ulva rigida on quality characteristics and oxidative stability of beef sausage during refrigerated storage. J. Food Sci. Technol. 2024, 1–12. [Google Scholar] [CrossRef]
- Malvis Romero, A.; Picado Morales, J.J.; Klose, L.; Liese, A. Enzyme-assisted extraction of ulvan from the green macroalgae Ulva fenestrata. Molecules 2023, 28, 6781. [Google Scholar] [CrossRef]
- Shannon, E.; Abu-Ghannam, N. Enzymatic extraction of fucoxanthin from brown seaweeds. Jordan Sinnott Found. Trust 2018, 53, 2195–2204. [Google Scholar] [CrossRef]
- Paul, P.T.; Joy, J.M.; Akshay, P.; Rosemol Jacob, M.; Amruth, P.; Chatterjee, N.S.; Mathew, S. Assessment of total phenolic content and in vitro antioxidant activity in brown seaweeds Turbinaria conoides and Padina tetrastromatica from the the Mandapam Coast. Fishery Technol. 2024, 61. [Google Scholar] [CrossRef]
- Chadwick, M.; Carvalho, L.G.; Vanegas, C.; Dimartino, S. A Comparative Review of Alternative Fucoidan Extraction Techniques from Seaweed. Mar. Drugs 2025, 23, 27. [Google Scholar] [CrossRef]
- Gu, X.; Cao, Z.; Zhao, L.; Seswita-Zilda, D.; Zhang, Q.; Fu, L.; Li, J. Metagenomic insights reveal the microbial diversity and associated algal-polysaccharide-degrading enzymes on the surface of red algae among remote regions. Int. J. Mol. Sci. 2023, 24, 11019. [Google Scholar] [CrossRef]
- Mathew, D.E.; Soni, A.; Dhimmar, A.; Gajjar, A.; Parab, A.S.; Phakatkar, S.S.; Sahastrabudhe, H.; Manohar, C.S.; Shinde, P.B.; Mantri, V.A. Characterization, bio-prospection, and comparative metagenomics of bacterial communities revealing the predictive functionalities in wild and cultured samples of industrially important red seaweed Gracilaria dura. Curr. Microbiol. 2025, 82, 85. [Google Scholar] [CrossRef]
- Bhattacharyya, P.N.; Tanti, B.; Barman, P.; Jha, D.K. Culture-independent metagenomic approach to characterize the surface and subsurface soil bacterial community in the Brahmaputra valley, Assam, North-East India, an Indo-Burma mega-biodiversity hotspot. World J. Microbiol. Biotechnol. 2014, 30, 519–528. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, F.; Zeng, Z.; Xu, M.; Sun, F.; Yang, L.; Bi, X.; Lin, Y.; Gao, Y.; Hao, H.; et al. Advances in Metagenomics and Its Application in Environmental Microorganisms. Front. Microbiol. 2021, 12, 766364. [Google Scholar] [CrossRef]
- Anderson, L.A.; Islam, M.A.; Prather, K.L.J. Synthetic biology strategies for improving microbial synthesis of “green” biopolymers. J. Biol. Chem. 2018, 293, 5053–5061. [Google Scholar] [CrossRef]
- Buchholz, K.; Kasche, V.; Bornscheuer, U.T. Biocatalysts and Enzyme Technology, 2nd ed.; KGaA, Ed.; Wiley-VCH Verlag & Co.: Weinheim, Germany, 2012. [Google Scholar]
- Barzkar, N.; Sheng, R.; Sohail, M.; Jahromi, S.T.; Babich, O.; Sukhikh, S.; Nahavandi, R. Alginate lyases from marine bacteria: An enzyme ocean for sustainable future. Molecules 2022, 27, 3375. [Google Scholar] [CrossRef]
- Xu, F.; Wang, P.; Zhang, Y.; Chen, X.; Zhou, N. Diversity of three-dimensional structures and catalytic mechanisms of alginate lyases. Appl. Environ. Microbiol. 2018, 84, 1–12. [Google Scholar] [CrossRef]
- Zheng, K.; Zhu, Y.; An, Z.; Lin, J.; Shan, S.; Zhang, H. Cloning, Expression and Characterization of an Alginate Lyase in Bacillus subtilis WB600. Fermentation 2023, 9, 144. [Google Scholar] [CrossRef]
- Rhein-Knudsen, N.; Guan, C.; Mathiesen, G.; Horn, S.J. Expression and production of thermophilic alginate lyases in Bacillus and direct application of culture supernatant for seaweed saccharification. Algal Res. 2021, 60, 102512. [Google Scholar] [CrossRef]
- Dharani, S.R.; Srinivasan, R.; Sarath, R.; Ramyam, M. Recent progress on engineering microbial alginate lyases towards their versatile role in biotechnological applications. Folia Microbiol. 2020, 65, 937–954. [Google Scholar] [CrossRef]
- Grobler, C.E.; Mabate, B.; Prins, A.; Le Roes-Hill, M.; Pletschke, B.I. Expression, purification, and characterisation of recombinant alginate lyase (Flammeovirga AL2) for the bioconversion of alginate into alginate oligosaccharides. Molecules 2024, 29, 5578. [Google Scholar] [CrossRef]
- Yang, M.; Yang, S.; Liu, Z.; Li, N.; Li, L.; Mou, H. Rational design of alginate lyase from Microbulbifer sp. Q7 to improve thermal stability. Mar. Drugs 2019, 17, 378. [Google Scholar] [CrossRef]
- Ndochinwa, O.G.; Wang, Q.; Amadi, O.C.; Nwagu, T.N.; Nnamchi, C.I.; Okeke, E.S.; Moneke, A.N. Current status and emerging frontiers in enzyme engineering: An industrial perspective. Heliyon 2024, 10, e32673. [Google Scholar] [CrossRef] [PubMed]
- Bilal, M.; Iqbal, H.M.N. Naturally-derived biopolymers: Potential platforms for enzyme immobilization. Int. J. Biol. Macromol. 2019, 30, 462–482. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.E.; Yang, Q.; Xiao, Z. Covalent immobilization of glucoamylase enzyme onto chemically activated surface of κ-carrageenan. Bull. Natl. Res. Cent. 2019, 23, 1–11. [Google Scholar] [CrossRef]
- Zayed, A.; Attia, E.Z.; Abdelmohsen, U.R. Adopting biorefinery and a circular bioeconomy for extracting and isolating natural products from marine algae. Front. Nat. Prod. 2024, 3, 1425242. [Google Scholar] [CrossRef]
- Orejuela-Escobar, L.M.; Landázuri, A.C.; Goodell, B. Second generation biorefining in Ecuador: Circular bioeconomy, zero waste technology, environment and sustainable development: The nexus. J. Bioresour. Bioprod. 2021, 6, 83–107. [Google Scholar] [CrossRef]
- El-Shamy, S.; Farag, M.A. Novel trends in extraction and optimization methods of bioactives recovery from pomegranate fruit biowastes: Valorization purposes for industrial applications. Food Chem. 2021, 365, 130465. [Google Scholar] [CrossRef]
- Álvarez-Viñas, M.; Flórez-Fernández, N.; Torres, M.D.; Domínguez, H. Successful approaches for a red seaweed biorefinery. Mar. Drugs 2019, 17, 620. [Google Scholar] [CrossRef] [PubMed]
- Spain, O.; Hardouin, K. Bourgougnon Nea Enzyme-assisted extraction of red seaweed Solieria chordalis (C.Agardh) J. Agardh 1842—The starting point for the production of biostimulants of plant growth and biosorbents of metal ions. Biomass Conv. Bioref. 2024, 14, 1621–1635. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mabate, B.; Mkabayi, L.; Goddard, D.R.; Grobler, C.E.; Pletschke, B.I. Role of Enzyme Technologies and Applied Enzymology in Valorising Seaweed Bioproducts. Mar. Drugs 2025, 23, 303. https://doi.org/10.3390/md23080303
Mabate B, Mkabayi L, Goddard DR, Grobler CE, Pletschke BI. Role of Enzyme Technologies and Applied Enzymology in Valorising Seaweed Bioproducts. Marine Drugs. 2025; 23(8):303. https://doi.org/10.3390/md23080303
Chicago/Turabian StyleMabate, Blessing, Lithalethu Mkabayi, Deandra Rochelle Goddard, Coleen Elizabeth Grobler, and Brett Ivan Pletschke. 2025. "Role of Enzyme Technologies and Applied Enzymology in Valorising Seaweed Bioproducts" Marine Drugs 23, no. 8: 303. https://doi.org/10.3390/md23080303
APA StyleMabate, B., Mkabayi, L., Goddard, D. R., Grobler, C. E., & Pletschke, B. I. (2025). Role of Enzyme Technologies and Applied Enzymology in Valorising Seaweed Bioproducts. Marine Drugs, 23(8), 303. https://doi.org/10.3390/md23080303