
The Alkaloid Caulerpin Exhibits Potent and Selective Anti-Inflammatory Activity Through Interaction with the Glucocorticoid Receptor

 ,
,  , and
, and
Abstract
1. Introduction
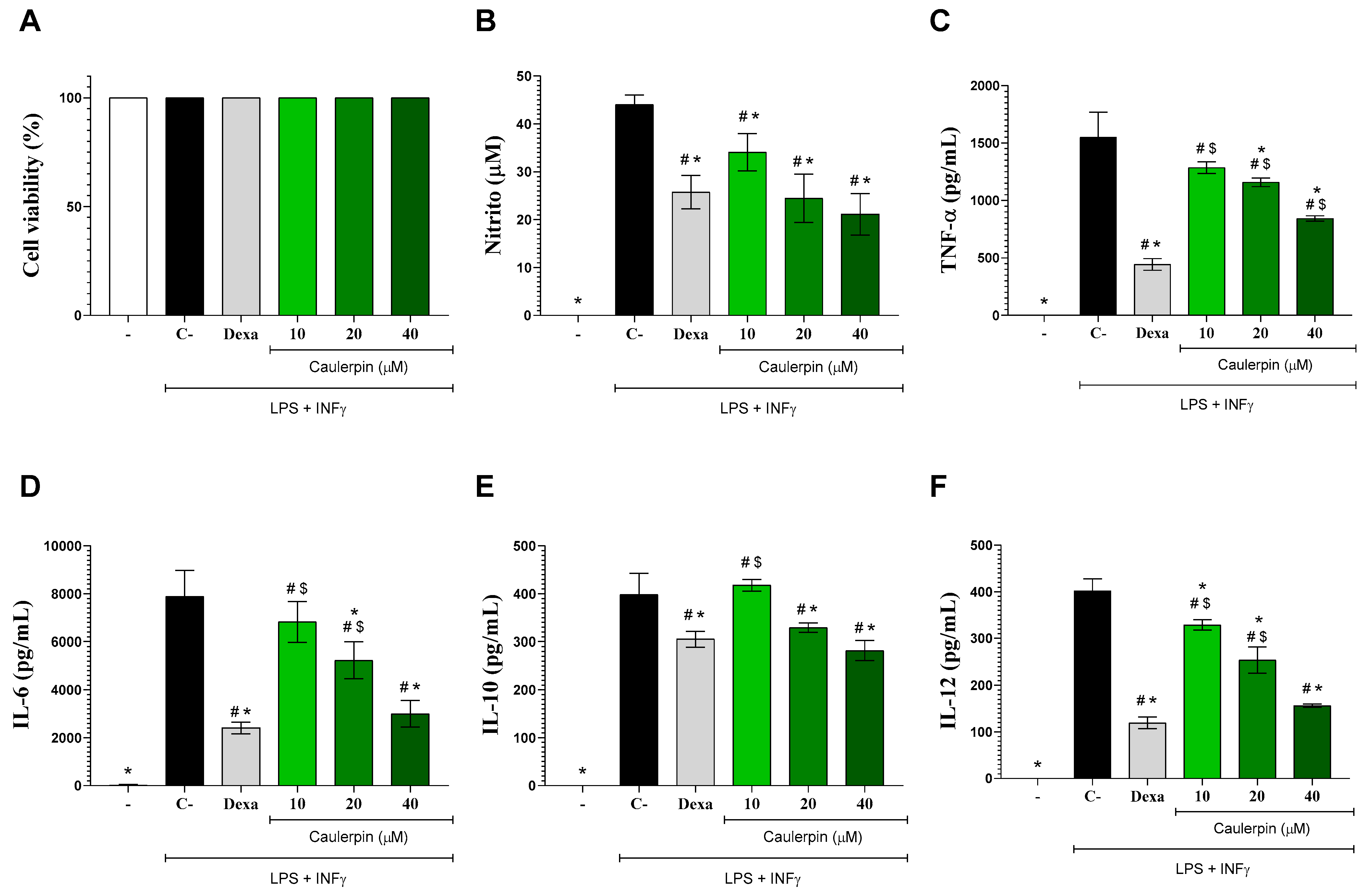
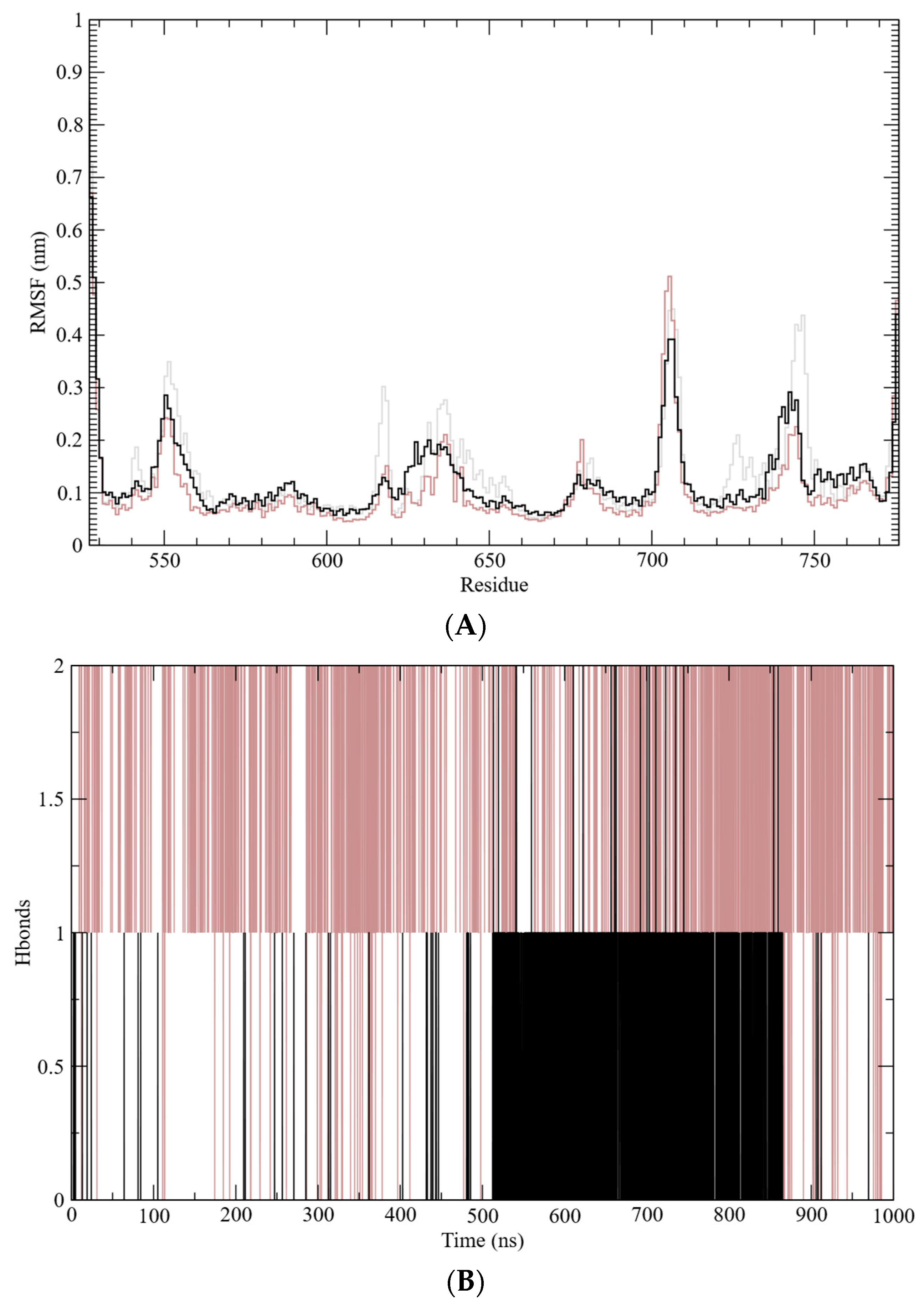
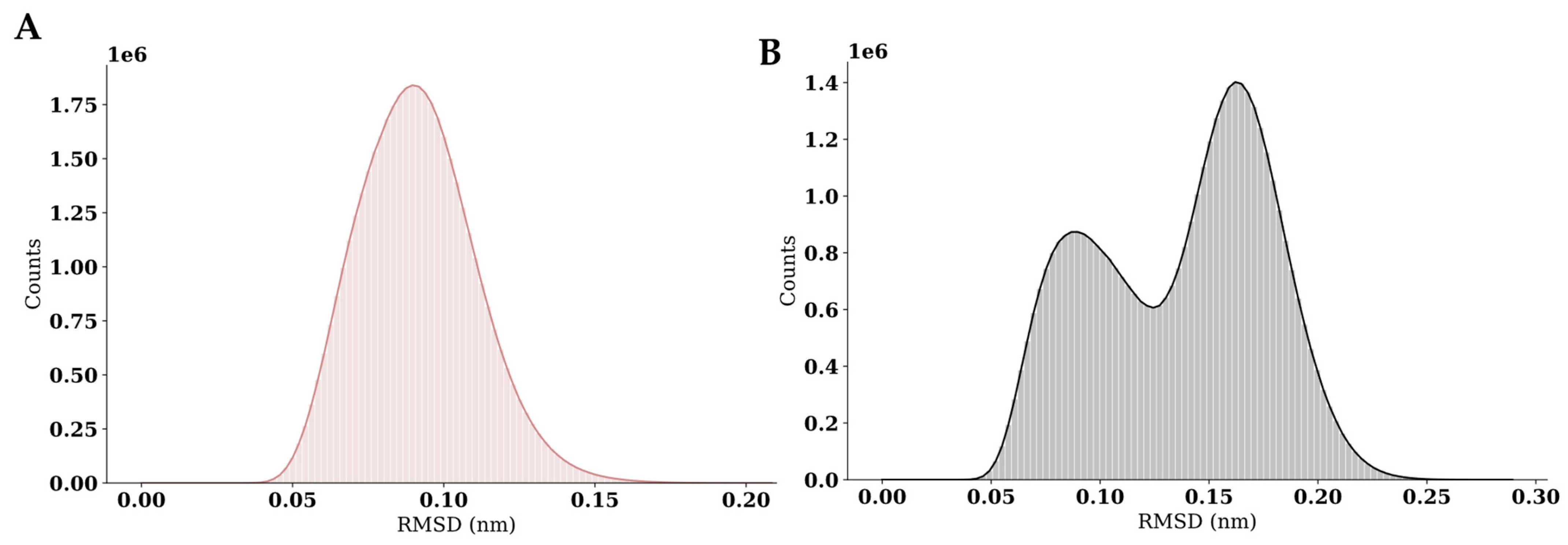
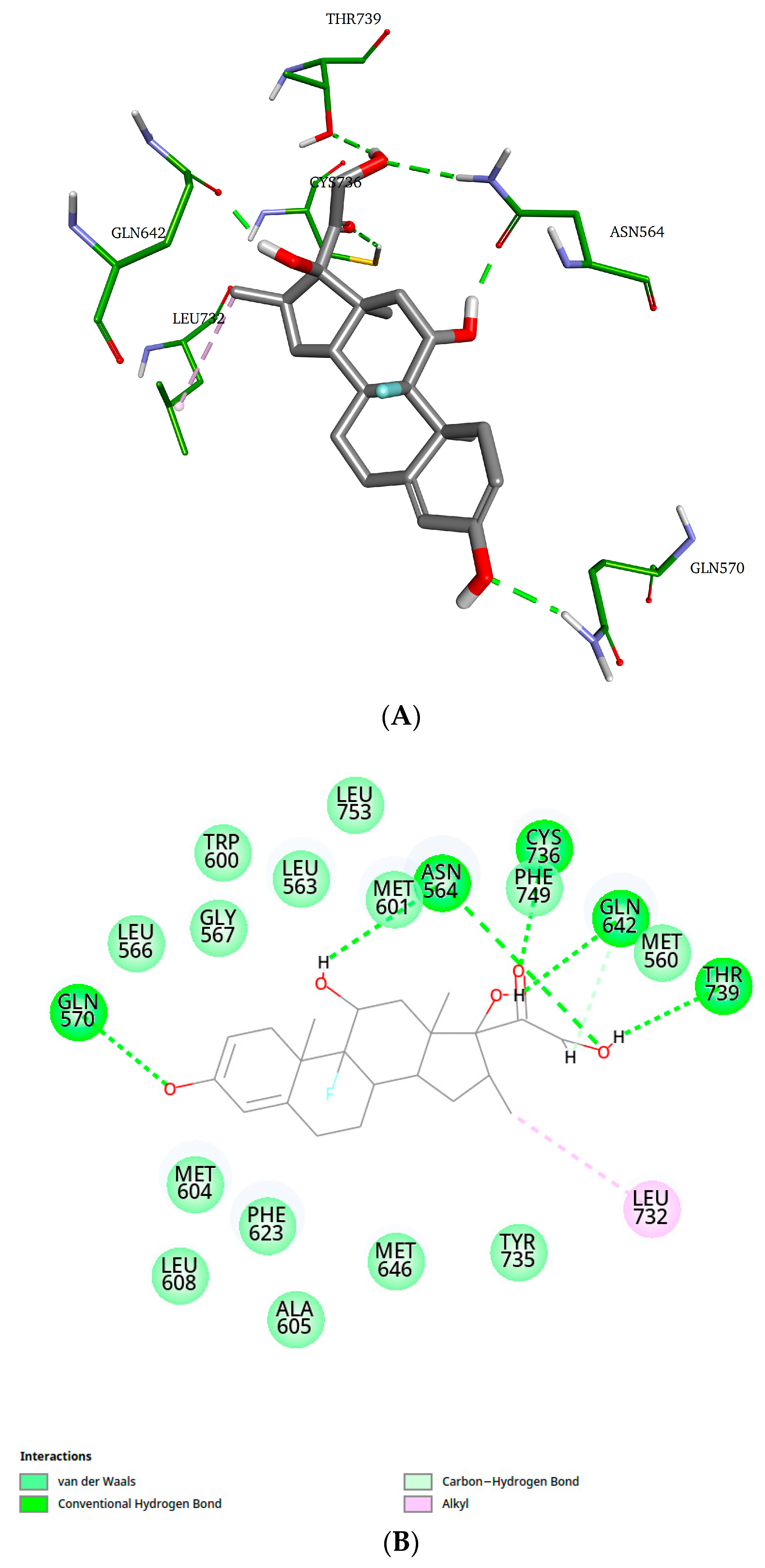
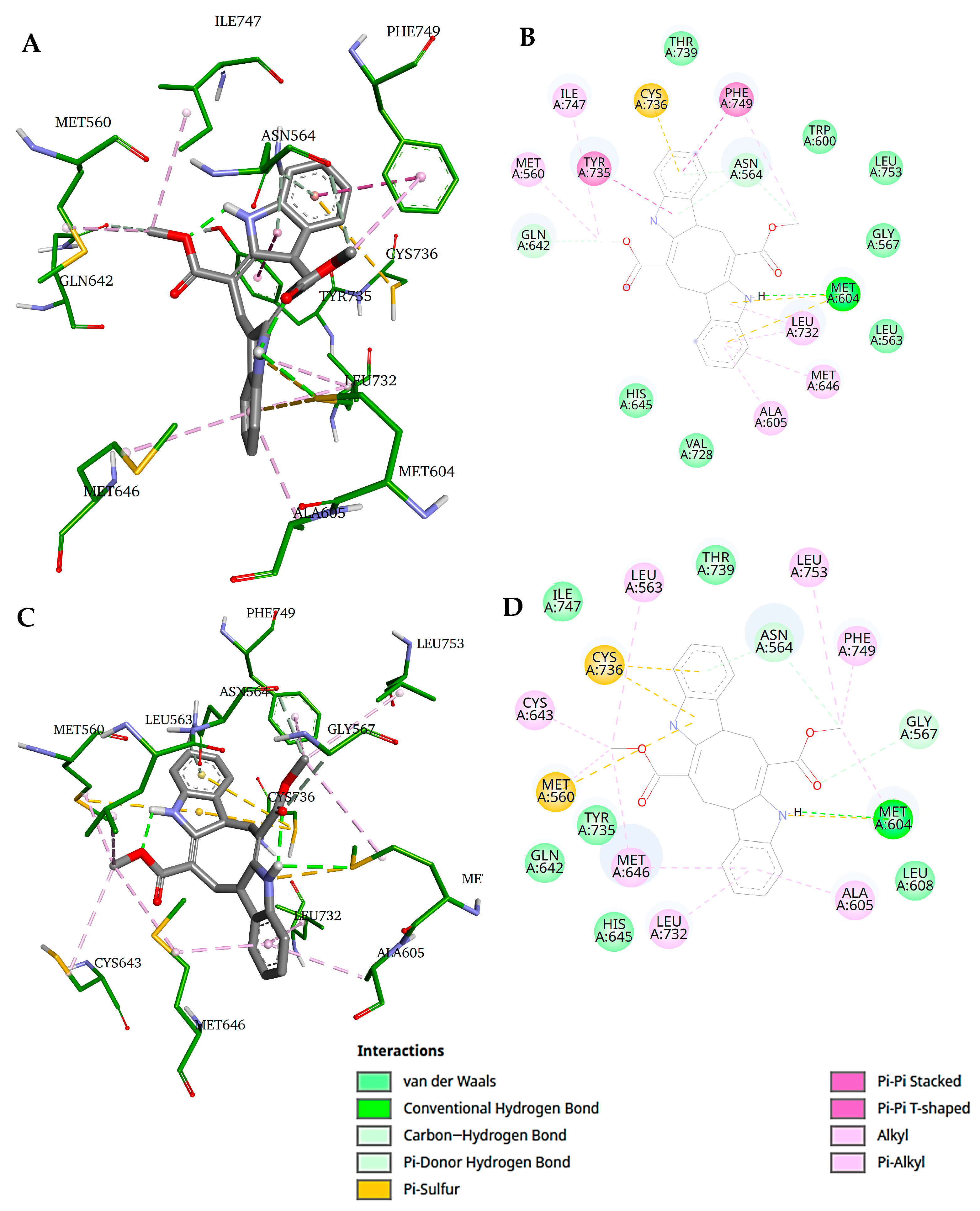
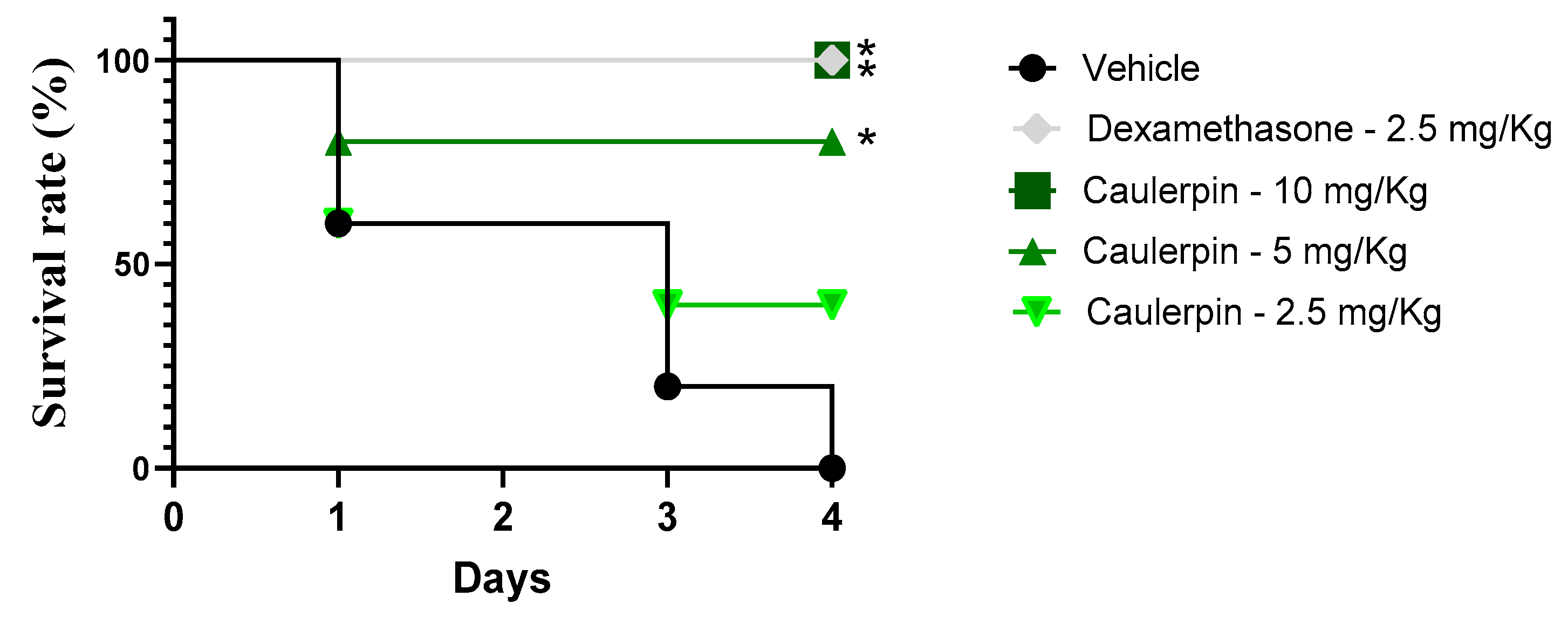
2. Results and Discussion
3. Materials and Methods
3.1. Drugs
3.2. Animals
3.3. In Silico Toxicity Prediction of Caulerpin Using PROTOX 3.0
3.4. Cytotoxicity Assay in Mammalian Cells
3.5. Macrophage Cultures
3.6. Cytokines and Nitric Oxide Production
3.7. Acute Toxicity in Mice
3.8. LPS-Induced Endotoxin Shock
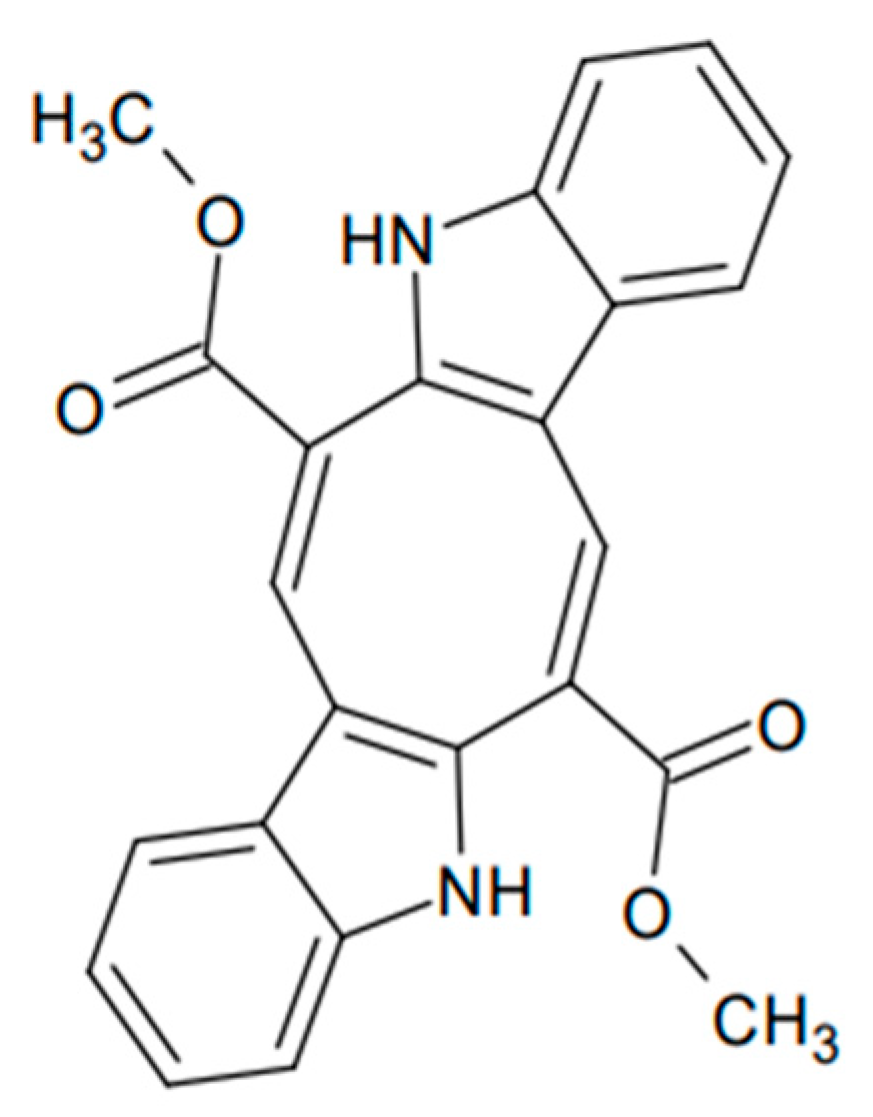
3.9. Dexamethasone and Caulerpin Structures
3.10. Molecular Docking
3.11. Dynamics Simulations
3.12. Visualization Tools and Plots
3.13. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| BBB | blood–brain barrier |
| CYP | cytochrome P450 |
| CGenFF | CHARMM General Force Field |
| DFT | density functional theory |
| Dexa | dexamethasone |
| DMEM | Dulbecco’s Modified Eagle Medium |
| DMSO | dimethyl sulfoxide |
| DSS | dextran sulfate sodium |
| ELISA | enzyme-linked immunosorbent assay |
| FBS | fetal bovine serum |
| GA | genetic algorithm |
| GR | glucocorticoid receptor |
| GR-LBD | glucocorticoid receptor ligand-binding domain |
| IFN-γ | interferon gamma |
| IL | interleukin |
| LD50 | lethal dose 50% |
| LPS | lipopolysaccharide |
| MD | molecular dynamics |
| MM/PBSA | molecular mechanics/Poisson–Boltzmann surface area |
| NF-κB | nuclear factor kappa B |
| NO | nitric oxide |
| NSAIDs | nonsteroidal anti-inflammatory drug |
| NVT | constant number of particles, volume, and temperature ensemble |
| NPT | constant number of particles, pressure, and temperature ensemble |
| ORP | oxidation-reduction potential |
| PBMC | peripheral blood mononuclear cells |
| PME | particle mesh Ewald |
| PROTOX | in silico toxicity prediction tool |
| PPAR-γ | peroxisome proliferator-activated receptor gamma |
| RMSD | root mean square deviation |
| RMSF | root mean square fluctuation |
| ROS | reactive oxygen species |
| TNF-α | tumor necrosis factor alpha |
References
- Singh, N.; Baby, D.; Rajguru, J.; Patil, P.; Thakkannavar, S.; Pujari, V. Inflammation and cancer. Ann. Afr. Med. 2019, 18, 121. [Google Scholar] [CrossRef] [PubMed]
- Kohler, O.; Krogh, J.; Mors, O.; Benros, M.E. Inflammation in Depression and the Potential for Anti-Inflammatory Treatment. Curr. Neuropharmacol. 2016, 14, 732–742. [Google Scholar] [CrossRef] [PubMed]
- Arulselvan, P.; Fard, M.T.; Tan, W.S.; Gothai, S.; Fakurazi, S.; Norhaizan, E.; Kumar, S. Role of Antioxidants and Natural Products in Inflammation. Oxid. Med. Cell. Longev. 2016, 2016, 5276130. [Google Scholar] [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2017, 9, 7204–7218. [Google Scholar] [CrossRef]
- Savin, I.A.; Zenkova, M.A.; Sen’kova, A.V. Pulmonary Fibrosis as a Result of Acute Lung Inflammation: Molecular Mechanisms, Relevant In Vivo Models, Prognostic and Therapeutic Approaches. Int. J. Mol. Sci. 2022, 23, 14959. [Google Scholar] [CrossRef]
- Bindu, S.; Mazumder, S.; Bandyopadhyay, U. Non-steroidal anti-inflammatory drugs (NSAIDs) and organ damage: A current perspective. Biochem. Pharmacol. 2020, 180, 114147. [Google Scholar] [CrossRef]
- Mehra, R.; Bhushan, S.; Bast, F.; Singh, S. Marine macroalga Caulerpa: Role of Its Metabolites in Modulating Cancer Signaling. Mol. Biol. Rep. 2019, 46, 3545–3555. [Google Scholar] [CrossRef]
- Kumar, J.G.S.; Umamaheswari, S.; Kavimani, S.; Ilavarasan, R. Pharmacological Potential of Green Algae Caulerpa: A Review. Int. J. Pharm. Sci. Res. 2019, 10, 1014–1024. [Google Scholar]
- Kurniawan, R.; Nurkolis, F.; Taslim, N.A.; Subali, D.; Surya, R.; Gunawan, W.B.; Alisaputra, D.; Mayulu, N.; Salindeho, N.; Kim, B. Carotenoids Composition of Green Algae Caulerpa racemosa and Their Antidiabetic, Anti-Obesity, Antioxidant, and Anti-Inflammatory Properties. Molecules 2023, 28, 3267. [Google Scholar] [CrossRef]
- Gaillande, C.; Payri, C.; Remoissenet, G.; Zubia, M. Caulerpa Consumption, Nutritional Value and Farming in the Indo-Pacific Region. J. Appl. Phycol. 2017, 29, 2249–2266. [Google Scholar] [CrossRef]
- Mert-Özüpek, N.; Calibaşi-Kocal, G.; Olgun, N.; Başbınar, Y.; Cavas, L.; Ellidokuz, H. An Efficient and Quick Analytical Method for the Quantification of an Algal Alkaloid Caulerpin Showed In-Vitro Anticancer Activity against Colorectal Cancer. Mar. Drugs 2022, 20, 757. [Google Scholar] [CrossRef] [PubMed]
- Ornano, L.; Donno, Y.; Sanna, C.; Ballero, M.; Serafini, M.; Bianco, A. Phytochemical Study of Caulerpa racemosa (Forsk.) J. Agarth, an Invading Alga in the Habitat of La Maddalena Archipelago. Nat. Prod. Res. 2014, 28, 1795–1799. [Google Scholar] [CrossRef] [PubMed]
- Abílio, G.M.F.; Camilo, C.J.; Coutinho, H.D.M.; da Costa, J.G.M.; Pena, L.J.; Silva-Júnior, A.; do Nascimento, Y.M.; Barbosa-Filho, J.M.; de Oliveira Santos, B.V.; de Luna Freire, K.R. Cytotoxic and Anti-HSV-1 Effects of Caulerpin Derivatives. Mar. Drugs 2024, 22, 3859. [Google Scholar] [CrossRef] [PubMed]
- Canché Chay, C.I.; Gómez Cansino, R.; Espitia Pinzón, C.I.; Torres-Ochoa, R.O.; Martínez, R. Synthesis and Anti-Tuberculosis Activity of the Marine Natural Product Caulerpin and Its Analogues. Mar. Drugs 2014, 12, 1757–1772. [Google Scholar] [CrossRef]
- Dissanayake, I.H.; Bandaranayake, U.; Keerthirathna, L.R.; Manawadu, C.; Silva, R.M.; Mohamed, B.; Ali, R.; Peiris, D.C. Integration of in vitro and in-silico analysis of Caulerpa racemosa against antioxidant, antidiabetic, and anticancer activities. Sci. Rep. 2022, 12, 20848. [Google Scholar] [CrossRef]
- Lucena, A.; Souza, C.; Jales, J.; Guedes, P.; Miranda, G.D.; Moura, A.; Araújo-Júnior, J.; Nascimento, G.; Scortecci, K.; Santos, B. The Bisindole Alkaloid Caulerpin, from Seaweeds of the Genus Caulerpa, Attenuated Colon Damage in a Murine Colitis Model. Mar. Drugs 2018, 16, 318. [Google Scholar] [CrossRef]
- Ferreira, D.A.; de Oliveira, G.C.S.L.; de Freitas, M.E.G.; de Araújo, D.A.M.; Scavone, C.; de Souza, T.A.; Villar, J.A.F.P.; Barbosa, L.A.; Mendonça-Junior, F.J.B.; Rodrigues-Junior, V.S.; et al. Evaluation of Anti-Inflammatory Activity of the New Cardiotonic Steroid γ-Benzylidene Digoxin 8 (BD-8) in Mice. Cells 2024, 13, 1568. [Google Scholar] [CrossRef]
- Massart, J.; Zoller, H.; Milosevic, I.; Haag, L.; Müller, T.; Haybaeck, J.; Oberkofler, H.; Stadlbauer, V. Role of Mitochondrial Cytochrome P450 2E1 in Healthy and Diseased Liver. Cells 2022, 11, 288. [Google Scholar] [CrossRef]
- Xu, J.; Ye, Y.; Huang, F.; Chen, G.; Pan, Z.; Yao, P. The Role of Human Cytochrome P450 2E1 in Liver Inflammation and Fibrosis. Hepatol. Commun. 2017, 1, 1043–1057. [Google Scholar] [CrossRef]
- Daly, A.K.; Rettie, A.E.; Fowler, D.M.; Miners, J.O. Pharmacogenomics of CYP2C9: Functional and Clinical Considerations. J. Pers. Med. 2017, 8, 1. [Google Scholar] [CrossRef]
- Yee, D.L.; O’Brien, S.H.; Young, G. Pharmacokinetics and Pharmacodynamics of Anticoagulants in Paediatric Patients. Clin. Pharmacokinet. 2013, 52, 967–980. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-H.; Ko, C.-I.; Ahn, G.; You, S.; Kim, J.-S.; Heu, M.S.; Kim, J.; Jee, Y.; Jeon, Y.-J. Molecular Characteristics and Anti-Inflammatory Activity of the Fucoidan Extracted from Ecklonia cava. Carbohydr. Polym. 2012, 89, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Zanger, U.M.; Schwab, M. Cytochrome P450 Enzymes in Drug Metabolism: Regulation of Gene Expression, Enzyme Activities, and Impact of Genetic Variation. Pharmacol. Ther. 2013, 138, 103–141. [Google Scholar] [CrossRef] [PubMed]
- Rakateli, L.; Huchzermeier, R.; van der Vorst, E.P.C. AhR, PXR and CAR: From Xenobiotic Receptors to Metabolic Sensors. Cells 2023, 12, 2752. [Google Scholar] [CrossRef]
- Dvorak, Z. Involvement of Aryl Hydrocarbon Receptor (AhR) in Polyphenol Inhibition of Benzo[a]pyrene-Induced Oxidative Stress and Neoplastic Transformation. Food Chem. Toxicol. 2017, 107, 523–525. [Google Scholar] [CrossRef]
- Shi, T.; van Soest, D.M.K.; Polderman, P.E.; Burgering, B.M.T.; Dansen, T.B. DNA Damage and Oxidant Stress Activate p53 through Differential Upstream Signaling Pathways. Free. Radic. Biol. Med. 2021, 172, 298–311. [Google Scholar] [CrossRef]
- Villapol, S. Roles of Peroxisome Proliferator-Activated Receptor Gamma on Brain and Peripheral Inflammation. Cell. Mol. Neurobiol. 2018, 38, 121–132. [Google Scholar] [CrossRef]
- Zhang, Y.; Tu, L.; Chen, J.; Zhou, L. Interference Mechanisms of Endocrine System and Other Systems of Endocrine-Disrupting Chemicals in Cosmetics—In Vitro Studies. Cells 2023, 12, 2752. [Google Scholar] [CrossRef]
- Cuomo, P.; Medaglia, C.; Allocca, I.; Montone, A.M.I.; Guerra, F.; Cabaro, S.; Mollo, E.; Eletto, D.; Papaianni, M.; Capparelli, R. Caulerpin Mitigates Helicobacter pylori-Induced Inflammation via Formyl Peptide Receptors. Int. J. Mol. Sci. 2021, 22, 13154. [Google Scholar] [CrossRef]
- Lim, E.H.; Mun, S.-K.; Kim, J.-J.; Chang, D.-J.; Yee, S.-T. Anti-Inflammatory Effects of Phlebia sp. Extract in Lipopolysaccharide-Stimulated RAW 264.7 Macrophages. BioMed Res. Int. 2022, 20, 2717196. [Google Scholar] [CrossRef]
- Sidrônio, T.S.; dos Santos, L.L.; Costa, A.B.; Macêdo, J.R.O.; Silva, M.F.; Freitas, M.L.; Costa, D.M.; Fuly, A.L.; Scotti, L.; Soares, M.B.P.; et al. Host-mediated antimicrobial effects and NLRP3 inflammasome modulation by caulerpin and its derivatives in macrophage models of mycobacterial infections. Microorganisms 2025, 13, 561. [Google Scholar] [CrossRef] [PubMed]
- Bitencourt, M.A.; Silva, H.M..; Abílio, G.M.; Miranda, G.E.; Moura, A.M.; Araújo-Júnior, J.X.D.; Silveira, E.J.; Santos, B.V.; Souto, J.T. Anti-Inflammatory Effects of Methanolic Extract of Green Algae Caulerpa mexicana in a Murine Model of Ulcerative Colitis. Braz. J. Pharmacogn. 2015, 25, 667–682. [Google Scholar] [CrossRef]
- Russo, T.; Coppola, F.; Paris, D.; De Marchi, L.; Meucci, V.; Motta, A.; Carbone, M.; Di Cosmo, A.; Soares, A.M.V.M.; Pretti, C.; et al. Exploring toxicological interactions in a changing sea: The case of the alkaloids caffeine and caulerpin. Sci. Total Environ. 2024, 912, 169190. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, Q.; Li, M.; Lao, J.; Tang, H.; Ming, S.; Wu, M.; Gong, S.; Li, L.; Liu, L.; et al. SLAMF7 Regulates the Inflammatory Response in Macrophages During Polymicrobial Sepsis. J. Clin. Investig. 2023, 133, e150224. [Google Scholar] [CrossRef]
- de Souza, É.T.; Ferreira, E.; Almeida-Lima, J.; Costa, L.S.; Tronchini, E.A.; Lima, M.E.; Rocha, H.A.; Leite, E.L. The Antinociceptive and Anti-Inflammatory Activities of Caulerpin, a Bisindole Alkaloid Isolated from Seaweeds of the Genus Caulerpa. Mar. Drugs 2009, 7, 689–704. [Google Scholar] [CrossRef]
- Cantarino, S.J.; Coutinho, R.; Soares, A.R.; Duarte, H.M.; Martinez, S.T. Microwave irradiation is a suitable method for caulerpin extraction from the green algae Caulerpa racemosa (Chlorophyta, Caulerpaceae). Nat. Prod. Res. 2022, 36, 2149–2153. [Google Scholar] [CrossRef]
- Green, L.C.; Wagner, D.A.; Glogowski, J.; Skipper, P.L.; Wishnok, J.S.; Tannenbaum, S.R. Analysis of Nitrate, Nitrite, and [15N]Nitrate in Biological Fluids. Anal. Biochem. 1982, 121, 431–439. [Google Scholar] [CrossRef]
- Silva, L.P.; Santos, I.P.; Silva, D.K.C.; dos Reis, B.P.Z.C.; Meira, C.S.; de Souza Castro, M.V.B.; dos Santos Filho, J.M.; de Araujo-Neto, J.H.; Ellena, J.A.; da Silveira, R.G.; et al. Molecular Hybridization Strategy on the Design, Synthesis, and Structural Characterization of Ferrocene-N-acyl Hydrazones as Immunomodulatory Agents. Molecules 2022, 27, 8343. [Google Scholar] [CrossRef]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem 2025 Update. Nucleic Acids Res. 2025, 53, D1516–D1525. [Google Scholar] [CrossRef]
- Neese, F. Software Update: The ORCA Program System—Version 5.0. Wiley Interdiscip. Rev. Comput. Mol. Sci. 2022, 12, e1606. [Google Scholar] [CrossRef]
- Jones, G.; Willett, P.; Glen, R.C.; Leach, A.R.; Taylor, R. Development and Validation of a Genetic Algorithm for Flexible Docking. J. Mol. Biol. 1997, 267, 727–748. [Google Scholar] [CrossRef] [PubMed]
- Korb, O.; Stützle, T.; Exner, T.E. Empirical Scoring Functions for Advanced Protein−Ligand Docking with PLANTS. J. Chem. Inf. Model. 2009, 49, 84–96. [Google Scholar] [CrossRef] [PubMed]
- GROMACS Development Team. GROMACS Documentation: Release 2024.2; GROMACS Development Team: 2024. Available online: https://manual.gromacs.org/2024.2/manual-2024.2.pdf (accessed on 1 September 2024).
- Huang, J.; Rauscher, S.; Nawrocki, G.; Ran, T.; Feig, M.; de Groot, B.L.; Grubmüller, H.; MacKerell, A.D. CHARMM36m: An Improved Force Field for Folded and Intrinsically Disordered Proteins. Nat. Methods 2017, 14, 71–73. [Google Scholar] [CrossRef]
- Vanommeslaeghe, K.; Hatcher, E.; Acharya, C.; Kundu, S.; Zhong, S.; Shim, J.; Darian, E.; Guvench, O.; Lopes, P.; Vorobyov, I.; et al. CHARMM General Force Field: A Force Field for Drug-Like Molecules Compatible with the CHARMM All-Atom Additive Biological Force Fields. J. Comput. Chem. 2010, 31, 671–690. [Google Scholar] [CrossRef] [PubMed]
- Berendsen, H.J.C.; Postma, J.P.M.; van Gunsteren, W.F.; DiNola, A.; Haak, J.R. Molecular Dynamics with Coupling to an External Bath. J. Chem. Phys. 1984, 81, 3684–3690. [Google Scholar] [CrossRef]
- Parrinello, M.; Rahman, A. Polymorphic Transitions in Single Crystals: A New Molecular Dynamics Method. J. Appl. Phys. 1981, 52, 7182–7190. [Google Scholar] [CrossRef]
- Essmann, U.; Perera, L.; Berkowitz, M.L.; Darden, T.; Lee, H.; Pedersen, L.G. A Smooth Particle Mesh Ewald Method. J. Chem. Phys. 1995, 103, 8577–8593. [Google Scholar] [CrossRef]
- Valdés-Tresanco, M.S.; Valdés-Tresanco, M.E.; Valiente, P.A.; Moreno, E. gmx_MMPBSA: A New Tool to Perform End-State Free Energy Calculations with GROMACS. J. Chem. Theory Comput. 2021, 17, 6281–6291. [Google Scholar] [CrossRef]
- Daura, X.; Gademann, K.; Jaun, B.; Seebach, D.; Van Gunsteren, W.F.; Mark, A.E. Peptide Folding When Simulation Meets Experiment. Angew. Chem. 1999, 111, 245–249. [Google Scholar] [CrossRef]
- BIOVIA, Dassault Systèmes 21.1.0. Discovery Studio Visualizer. Dassault Systèmes: San Diego, CA, USA, 2021.
- Grace. Available online: https://plasma-gate.weizmann.ac.il/Grace/ (accessed on 2 February 2025).




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organ Toxicity or Toxicity End Points | Prediction | Probability |
|---|---|---|
| Hepatotoxicity | Inactive | 0.59 |
| Neurotoxicity | Inactive | 0.7 |
| Nephrotoxicity | Active | 0.56 |
| Respiratory toxicity | Active | 0.5 |
| Cardiotoxicity | Inactive | 0.84 |
| Carcinogenicity | Inactive | 0.6 |
| Immunotoxicity | Inactive | 0.96 |
| Mutagenicity | Active | 0.53 |
| Cytotoxicity | Inactive | 0.71 |
| BBB-barrier | Active | 0.67 |
| Ecotoxicity | Inactive | 0.57 |
| Clinical toxicity | Inactive | 0.52 |
| Nutritional toxicity | Inactive | 0.52 |
| Behavior and General Appearance | Observations * | |
|---|---|---|
| Vehicle | Caulerpin (100 mg/Kg) | |
| Changes in the eyes | No changes | No changes |
| Changes in the fur | No changes | No changes |
| Changes in the skin | No changes | No changes |
| Coma | Absent | Absent |
| Convulsions | Absent | Absent |
| Diarrhea | Absent | Absent |
| Lethargy | Absent | Absent |
| Salivation | Absent | Absent |
| Sleep | Usual | Usual |
| Tremors | Absent | Absent |
| Days | Vehicle | Caulerpin (100 mg/Kg) |
|---|---|---|
| 0 | 20.2 (± 1.0) | 19.5 (± 1.9) |
| 7 | 22.2 (± 0.8) | 22.7 (± 1.0) |
| 14 | 24.2 (± 1.1) | 24.4 (± 0.8) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
dos Santos, J.S.P.; Silva, D.K.C.; da Silva Oliveira, V.; Junior, S.S.S.; dos Santos Rodrigues, E.; de Souza, C.V.C.; Martinez, S.T.; Santos-Filho, O.A.; Meira, C.S.; Soares, M.B.P. The Alkaloid Caulerpin Exhibits Potent and Selective Anti-Inflammatory Activity Through Interaction with the Glucocorticoid Receptor. Mar. Drugs 2025, 23, 232. https://doi.org/10.3390/md23060232
dos Santos JSP, Silva DKC, da Silva Oliveira V, Junior SSS, dos Santos Rodrigues E, de Souza CVC, Martinez ST, Santos-Filho OA, Meira CS, Soares MBP. The Alkaloid Caulerpin Exhibits Potent and Selective Anti-Inflammatory Activity Through Interaction with the Glucocorticoid Receptor. Marine Drugs. 2025; 23(6):232. https://doi.org/10.3390/md23060232
Chicago/Turabian Styledos Santos, Jônatas Sousa Pires, Dahara Keyse Carvalho Silva, Vanessa da Silva Oliveira, Sergio Santos Silva Junior, Edivaldo dos Santos Rodrigues, Claudia Valeria Campos de Souza, Sabrina Teixeira Martinez, Osvaldo Andrade Santos-Filho, Cássio Santana Meira, and Milena Botelho Pereira Soares. 2025. "The Alkaloid Caulerpin Exhibits Potent and Selective Anti-Inflammatory Activity Through Interaction with the Glucocorticoid Receptor" Marine Drugs 23, no. 6: 232. https://doi.org/10.3390/md23060232
APA Styledos Santos, J. S. P., Silva, D. K. C., da Silva Oliveira, V., Junior, S. S. S., dos Santos Rodrigues, E., de Souza, C. V. C., Martinez, S. T., Santos-Filho, O. A., Meira, C. S., & Soares, M. B. P. (2025). The Alkaloid Caulerpin Exhibits Potent and Selective Anti-Inflammatory Activity Through Interaction with the Glucocorticoid Receptor. Marine Drugs, 23(6), 232. https://doi.org/10.3390/md23060232