

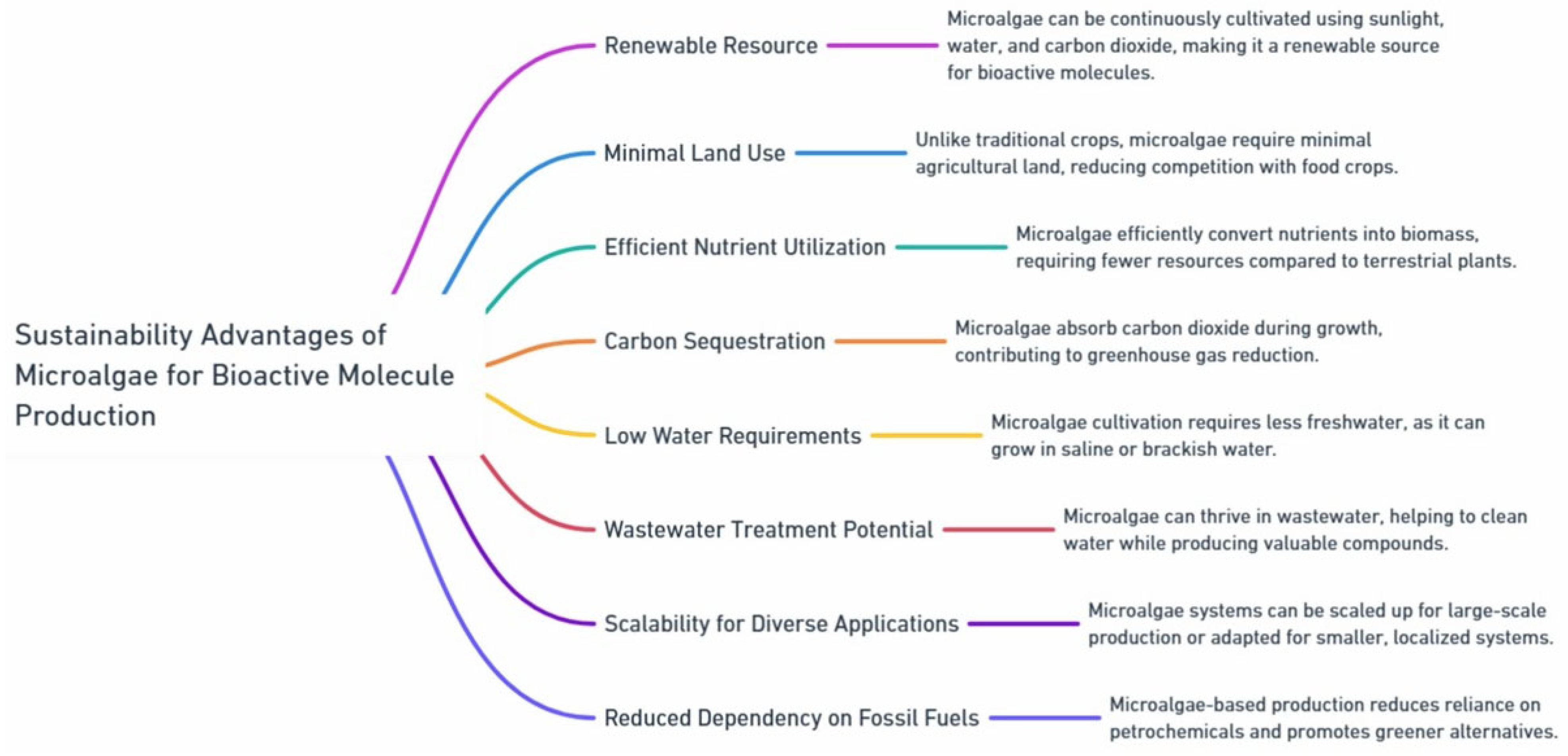
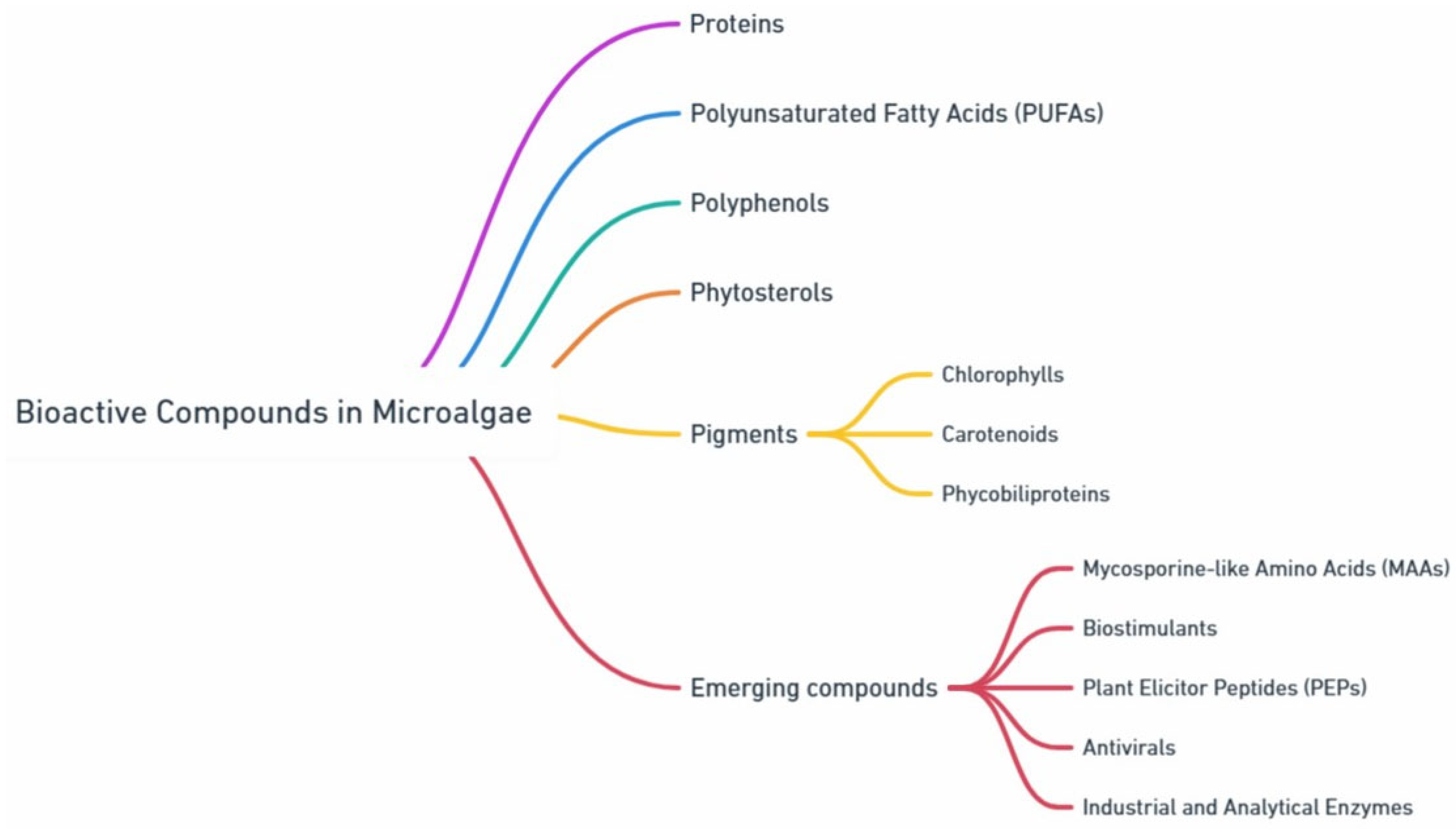
Diversity of Bioactive Compounds in Microalgae: Key Classes and Functional Applications

 , , , , ,
, , , , ,  , , , , , ,
, , , , , ,  ,
,  , , , , ,
, , , , ,  and
and  add
Show full author list
add
Show full author list
Abstract
1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Conditions | ||||
|---|---|---|---|---|---|
| Illumination Period | Temperature (°C) | Nutrient Medium | pH | References | |
| Chlorella vulgaris | 24 h light/day | 25 | BG11 medium | 9 | Deniz, 2020 [27] |
| Limnospira platensis * | 12 h light/day | 32 | Zarrouk’s medium | 9.5 | Soni et al., 2019 [28] |
| Dunaliella salina | 15 h light/day | 25 | f/2 medium | 7.2 | Al-Mhanna et al., 2023 [29] Borovkov et al., 2020 [30] |
| Nannochloropsis oculata | 16 h light/day | 25 | f/2 medium | 7.2 | Maglie et al., 2021 [31] |
| Haematococcus lacustris | 16 h light/day | 30 | Bold’s basal medium | 5 | Hanan et al., 2013 [32] Nahidian et al., 2018 [33] |
| Substance/Microalgal Species | Date of FDA Closure | GRAS Notification No. | Common Applications |
|---|---|---|---|
| Limnospira platensis | 27 July 2011 | GRN 391 | Dietary supplements, protein fortification, natural colorant |
| Chlorella vulgaris | 8 January 2007/23 September 2016 | GRN 224/641 | Smoothies, nutrition bars, fortified beverages |
| Dunaliella salina | 21 May 2002 | GRN 103 | Natural β-carotene source, food colorant, functional ingredient |
| Haematococcus lacustris | 20 October 2010/11 June 2013 | GRN 314/452 | Astaxanthin-rich supplements and functional foods |
| Schizochytrium sp. | 17 February 2004/14 May 2018 | GRN 137/778 | DHA omega-3 for infant formula and nutrition products |
| Euglena gracilis (dried biomass) | 19 July 2017 | GRN 697 | β-1,3-glucans; used in beverages, supplements, and snacks |
| Euglena gracilis | 22 May 2017 | GRN 686 | β-1,3-glucans; used in beverages, supplements, and snacks |
| Aurantiochytrium limacinum TKD-1 (algal oil, ≥45% DHA) | 25 February 2022 | GRN 1008 | DHA-rich algal oil for food and supplements |
| Chlorella protothecoides strain S106 | 12 Jun 2014 | GRN 519 | High-protein flour (40–75%), used in fortified foods |
| Chlorella sorokiniana | 21 December 2021 | GRN 986 | Powder and micro powder for dietary applications |
| Dunaliella bardawil | 19 August 2010 | GRN 351 | β-carotene-rich natural food colorant |
| Haematococcus lacustris | 4 June 2015 | GRN 580 | Astaxanthin esters for supplements and fortified foods |
| Ulkenia sp. SAM2179 (micro-algal oil) | 21 January 2010 | GRN 319 | DHA-rich oil used in nutritional applications |
2. High Added-Value Compounds from Microalgae
2.1. Proteins
2.2. Polyunsaturated Fatty Acids (PUFAs)
2.3. Polyphenols
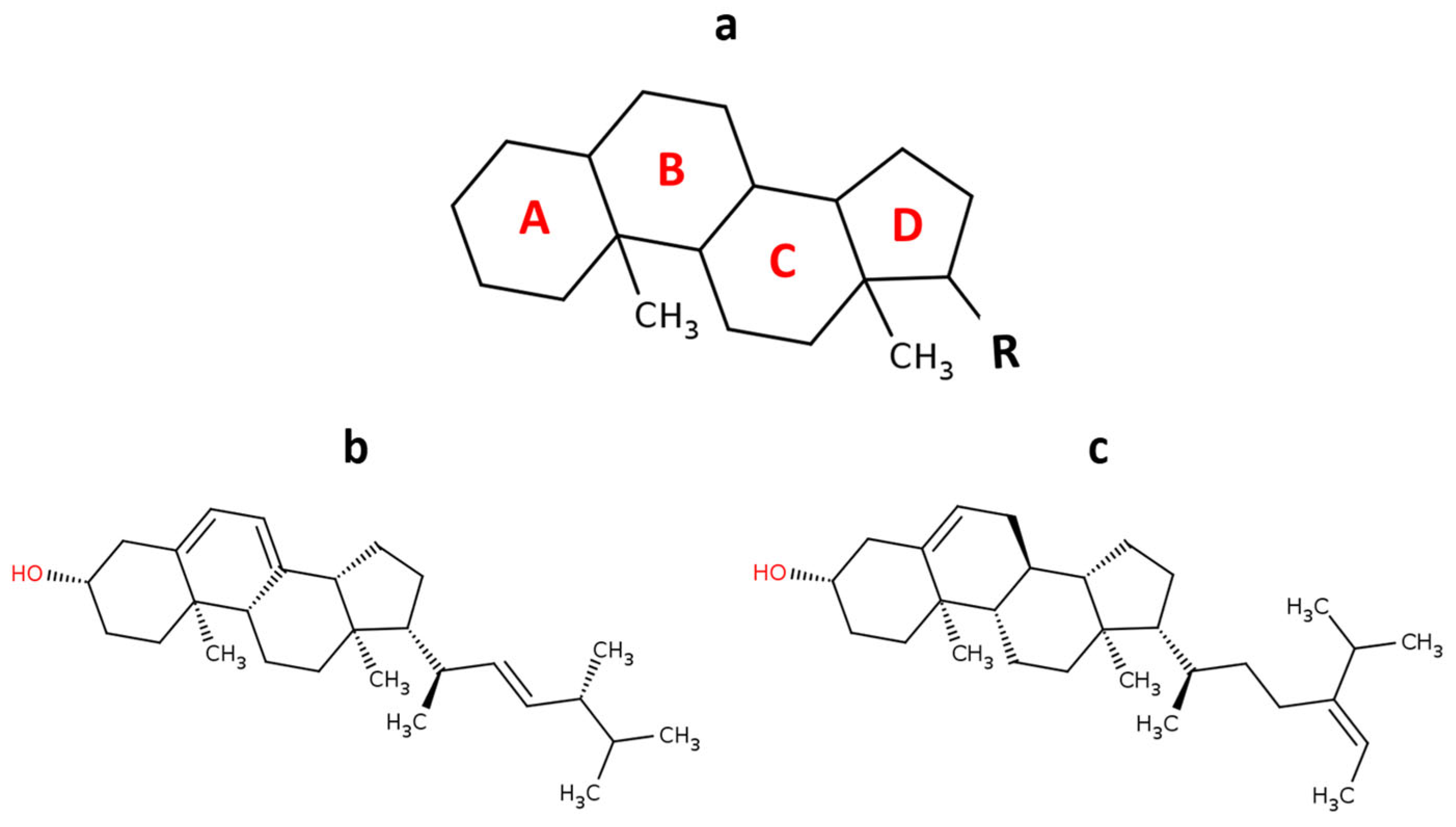
2.4. Phytosterols
2.5. Pigments
2.5.1. Chlorophylls
2.5.2. Carotenoids
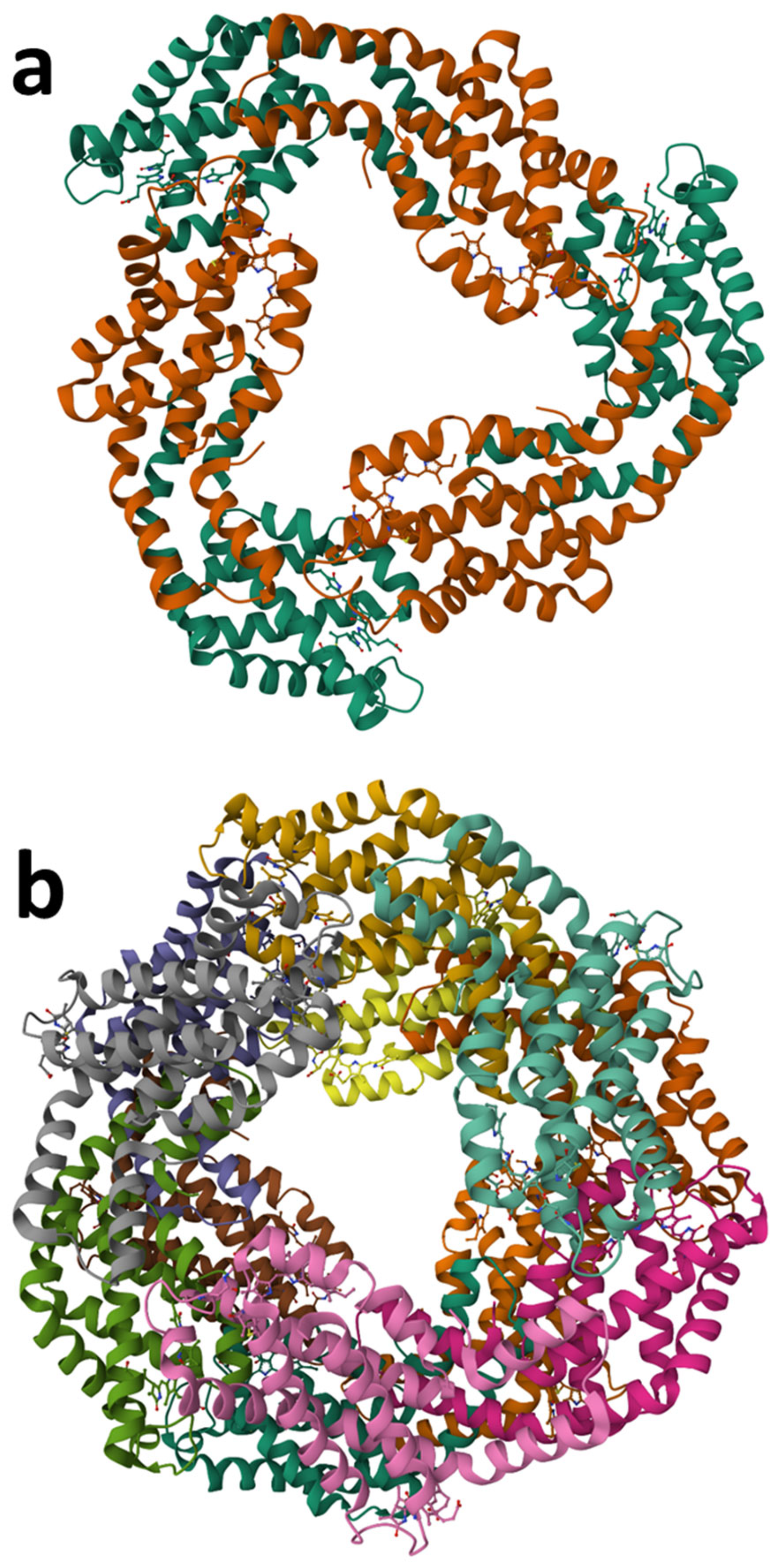
2.5.3. Phycobiliproteins
2.6. Biostimulants
2.7. Plant Elicitor Peptides (PEPs)
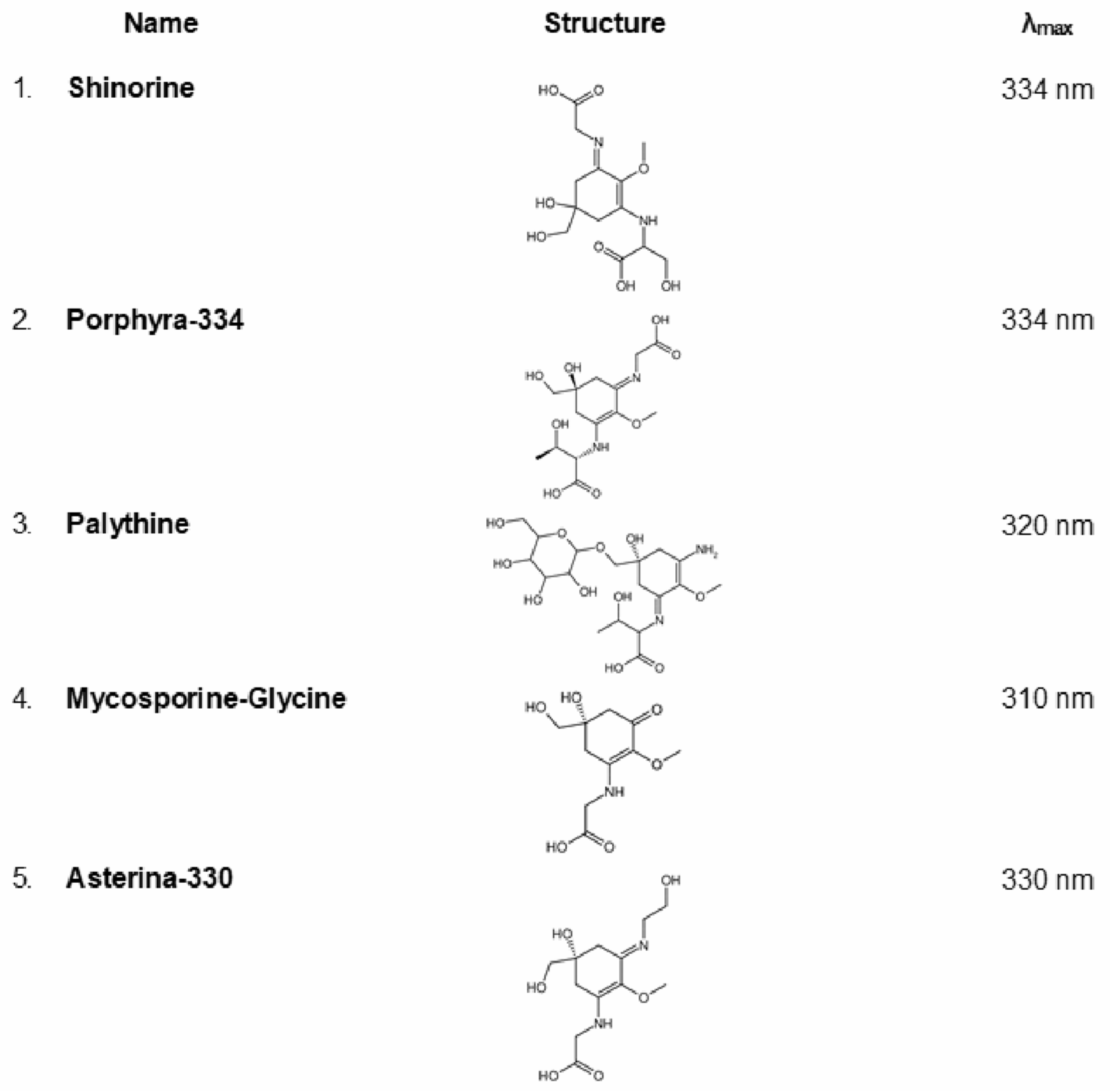
2.8. Mycosporine-like Amino Acids (MAAs)
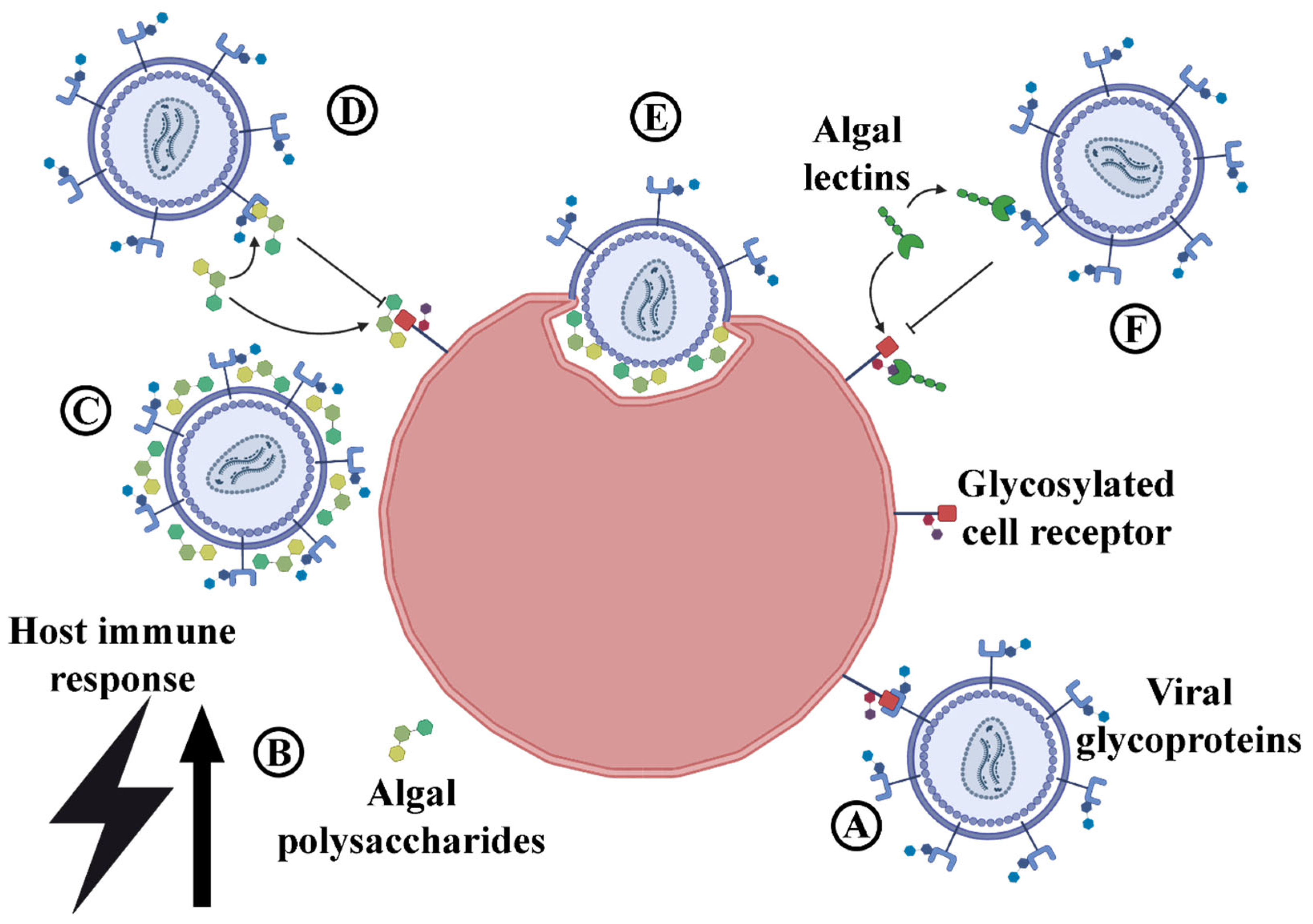
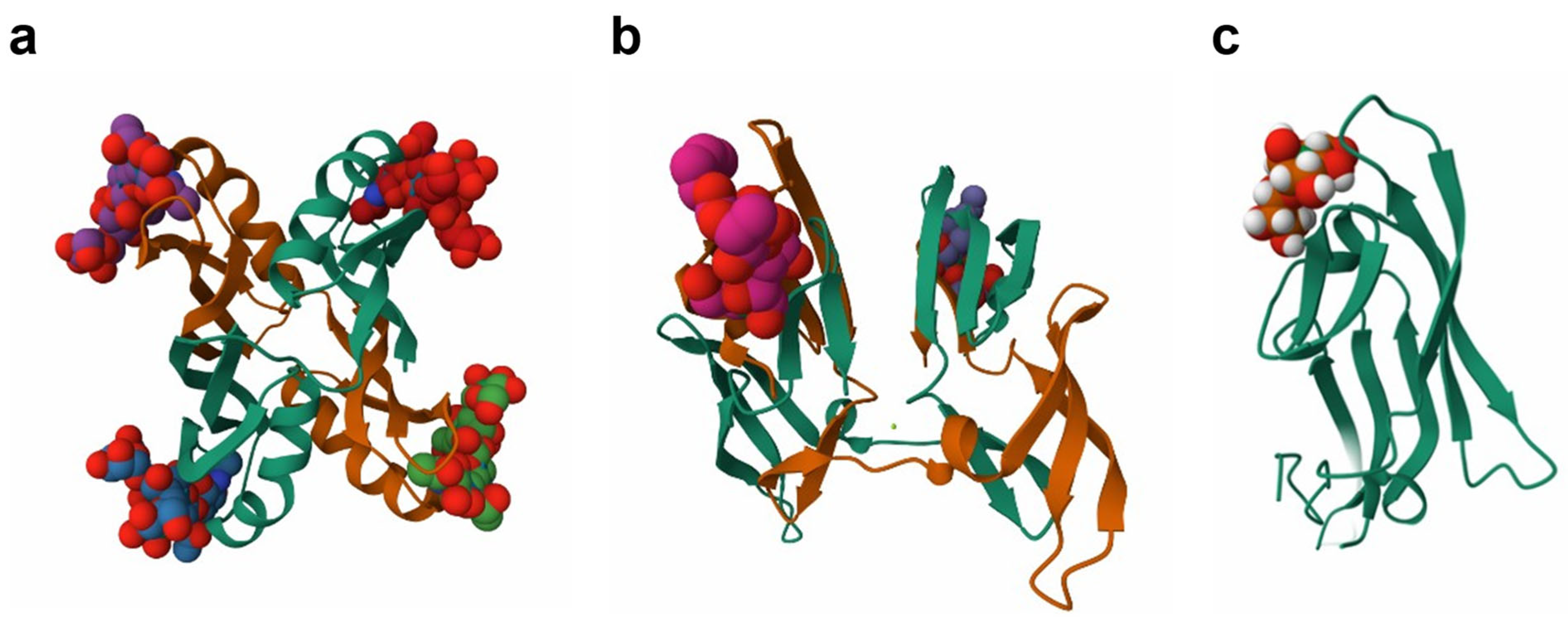
2.9. Antivirals
2.10. Analytical and Industrial Enzymes
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Inam, A.; Oncu-Oner, T.; Deniz, I. Algae in Biomedicine. In Cell Biology and Translational Medicine; Springer: Cham, Switzerland, 2024. [Google Scholar]
- Tan, J.S.; Lee, S.Y.; Chew, K.W.; Lam, M.K.; Lim, J.W.; Ho, S.-H.; Show, P.L. A review on microalgae cultivation and harvesting, and their biomass extraction processing using ionic liquids. Bioengineered 2020, 11, 116–129. [Google Scholar] [CrossRef]
- Najafi, A.; Heidary, M.; Martinez, R.M.; Baby, A.R.; Morowvat, M.H. Microalgae-based sunscreens as green and sustainable cosmetic products. Int. J. Cosmet. Sci. 2024, 47, 213–222. [Google Scholar] [CrossRef]
- Yu, B.S.; Pyo, S.; Lee, J.; Han, K. Microalgae: A multifaceted catalyst for sustainable solutions in renewable energy, food security, and environmental management. Microb. Cell Factories 2024, 23, 308. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.; Hu, Q. Microalgae as feed sources and feed additives for sustainable aquaculture: Prospects and challenges. Rev. Aquac. 2024, 16, 818–835. [Google Scholar] [CrossRef]
- Su, M.; Bastiaens, L.; Verspreet, J.; Hayes, M. Applications of microalgae in foods, pharma and feeds and their use as fertilizers and biostimulants: Legislation and regulatory aspects for consideration. Foods 2023, 12, 3878. [Google Scholar] [CrossRef]
- Saadaoui, I.; Rasheed, R.; Aguilar, A.; Cherif, M.; Al Jabri, H.; Sayadi, S.; Manning, S.R. Microalgal-based feed: Promising alternative feedstocks for livestock and poultry production. J. Anim. Sci. Biotechnol. 2021, 12, 76. [Google Scholar] [CrossRef] [PubMed]
- Jha, D.; Jain, V.; Sharma, B.; Kant, A.; Garlapati, V.K. Microalgae-based pharmaceuticals and nutraceuticals: An emerging field with immense market potential. ChemBioEng Rev. 2017, 4, 257–272. [Google Scholar] [CrossRef]
- Wijffels, R.H.; Kruse, O.; Hellingwerf, K.J. Potential of industrial biotechnology with cyanobacteria and eukaryotic microalgae. Curr. Opin. Biotechnol. 2013, 24, 405–413. [Google Scholar] [CrossRef]
- Pulz, O.; Gross, W. Valuable products from biotechnology of microalgae. Appl. Microbiol. Biotechnol. 2004, 65, 635–648. [Google Scholar] [CrossRef]
- Chisti, Y. Biodiesel from microalgae. Biotechnol. Adv. 2007, 25, 294–306. [Google Scholar] [CrossRef]
- Esteves, A.F.; Gonçalves, A.L.; Vilar, V.J.; Pires, J.C. Is it possible to shape the microalgal biomass composition with operational parameters for target compound accumulation? Biotechnol. Adv. 2024, 79, 108493. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Wang, X.; Zhu, T. Research progress and application of carbon sequestration in industrial flue gas by microalgae: A review. J. Environ. Sci. 2024, 152, 14–28. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Yadav, P.; Sharma, H.; Kumar, A.; Hashem, A.; Abd_Allah, E.F.; Gupta, R.K. Unlocking the adaptation mechanisms of the oleaginous microalga Scenedesmus sp. BHU1 under elevated salt stress: A physiochemical, lipidomics and transcriptomics approach. Front. Microbiol. 2024, 15, 1475410. [Google Scholar] [CrossRef]
- Ievina, B.; Romagnoli, F. Unveiling underlying factors for optimizing light spectrum to enhance microalgae growth. Bioresour. Technol. 2024, 418, 131980. [Google Scholar] [CrossRef] [PubMed]
- Georgieva, Z.; Karcheva, Z.; Toshkova-Yotova, T.; Georgieva, A.; Toshkova, R.; Petrova, D.; Zhiponova, M.; Chaneva, G. Blue Light Enhances the Antioxidant, Antimicrobial, and Antitumor Potential of the Green Microalgae Coelastrella sp. BGV. Plants 2024, 13, 3295. [Google Scholar] [CrossRef]
- Castillo, A.; Finimundy, T.C.; Heleno, S.A.; Rodrigues, P.; Fernandes, F.A.; Pereira, S.; Lores, M.; Barros, L.; Garcia-Jares, C. The generally recognized as safe (GRAS) microalgae Haematococcus pluvialis (wet) as a multifunctional additive for coloring and improving the organoleptic and functional properties of foods. Food Funct. 2023, 14, 6023–6035. [Google Scholar] [CrossRef]
- Masi, A.; Leonelli, F.; Scognamiglio, V.; Gasperuzzo, G.; Antonacci, A.; Terzidis, M.A. Chlamydomonas reinhardtii: A factory of nutraceutical and food supplements for human health. Molecules 2023, 28, 1185. [Google Scholar] [CrossRef]
- Song, I.; Kim, S.; Kim, J.; Oh, H.; Jang, J.; Jeong, S.J.; Baek, K.; Shin, W.-S.; Sim, S.J.; Jin, E. Macular pigment-enriched oil production from genome-edited microalgae. Microb. Cell Factories 2022, 21, 27. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Raven, J.A.; Beardall, J.; Overmans, S.; Xia, J.; Jin, P. The trade-offs associated with the adaptions of marine microalgae to high CO2 and warming. Mar. Environ. Res. 2024, 204, 106853. [Google Scholar] [CrossRef]
- Shitu, A.; Tadda, M.A.; Zhao, J.; Danhassan, U.A.; Ye, Z.; Liu, D.; Chen, W.; Zhu, S. Review of recent advances in utilising aquaculture wastewater for algae cultivation and microalgae-based bioproduct recovery. Environ. Geochem. Health 2024, 46, 485. [Google Scholar] [CrossRef]
- Available online: https://www.eaba-association.org/en/resources?resource_hit=5341&token=397a9021afc6ad2402a1111773c29303 (accessed on 20 May 2025).
- Mikkili, I.; Gaddirala, B.V.S.T.; Borugadda, S.; Davuluri, S.B. Harnessing algal biomass for sustainable energy: Cultivation, strain improvement, and biofuel production. Prep. Biochem. Biotechnol. 2024, 55, 521–534. [Google Scholar] [CrossRef] [PubMed]
- Gamal, R.; Shreadah, M.A. Marine microalgae and their industrial biotechnological applications: A review. J. Genet. Eng. Biotechnol. 2024, 22, 100407. [Google Scholar] [CrossRef]
- Rawindran, H.; Khoo, K.S.; Satpati, G.G.; Maity, S.; Chandran, K.; Lim, J.W.; Tong, W.Y.; Setiabudi, H.D.; Yunus, N.M. Composition of carbohydrate, protein and lipid derived from microalgae using thermally pretreated solid waste. J. Sci. Food Agric. 2024. [CrossRef]
- Lori, M.; Van Verk, M.C.; Hander, T.; Schatowitz, H.; Klauser, D.; Flury, P.; Gehring, C.A.; Boller, T.; Bartels, S. Evolutionary divergence of the plant elicitor peptides (Peps) and their receptors: Interfamily incompatibility of perception but compatibility of downstream signalling. J. Exp. Bot. 2015, 66, 5315–5325. [Google Scholar] [CrossRef]
- Deniz, İ. Determination of growth conditions for Chlorella vulgaris. Mar. Sci. Technol. Bull. 2020, 9, 114–117. [Google Scholar] [CrossRef]
- Soni, R.A.; Sudhakar, K.; Rana, R. Comparative study on the growth performance of Spirulina platensis on modifying culture media. Energy Rep. 2019, 5, 327–336. [Google Scholar] [CrossRef]
- Al-Mhanna, N.; Pistorius, M.; Al Sammarraie, L. Optimization of the Cultivation Conditions of the Green Algae Dunaliella salina by Using Simplex Method. Processes 2023, 11, 292. [Google Scholar] [CrossRef]
- Borovkov, A.B.; Gudvilovich, I.N.; Avsiyan, A.L.; Memetshaeva, N.O.A.; Lelekov, A.S.; Novikova, T.M. Production characteristics of Dunaliella salina at two-phase pilot cultivation (Crimea). Turk. J. Fish. Aquat. Sci. 2019, 20, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Maglie, M.; Baldisserotto, C.; Guerrini, A.; Sabia, A.; Ferroni, L.; Pancaldi, S. A co-cultivation process of Nannochloropsis oculata and Tisochrysis lutea induces morpho-physiological and biochemical variations potentially useful for biotechnological purposes. J. Appl. Phycol. 2021, 33, 2817–2832. [Google Scholar] [CrossRef]
- Hanan, N.A.; Al-Shorgani, N.K.; Shukor, H.; Kalil, M.S. Pre-optimization conditions for Haematococcus pluvialis growth. Int. J. Adv. Sci. Eng. Inf. Technol. 2013, 3, 168–171. [Google Scholar] [CrossRef]
- Nahidian, B.; Ghanati, F.; Shahbazi, M.; Soltani, N. Effect of nutrients on the growth and physiological features of newly isolated Haematococcus pluvialis TMU1. Bioresour. Technol. 2018, 255, 229–237. [Google Scholar] [CrossRef] [PubMed]
- WHO. Protein and amino acid requirements in human nutrition. In World Health Organization Technical Report Series; WHO: Geneva, Switzerland, 2007; Volume 935, p. 1. [Google Scholar]
- Spolaore, P.; Joannis-Cassan, C.; Duran, E.; Isambert, A. Commercial applications of microalgae. J. Biosci. Bioeng. 2006, 101, 87–96. [Google Scholar] [CrossRef]
- Converti, A.; Casazza, A.A.; Ortiz, E.Y.; Perego, P.; Del Borghi, M. Effect of temperature and nitrogen concentration on the growth and lipid content of Nannochloropsis oculata and Chlorella vulgaris for biodiesel production. Chem. Eng. Process. Process Intensif. 2009, 48, 1146–1151. [Google Scholar] [CrossRef]
- Ma, S.; Sun, S.; Li, T.; Yan, Y.; Wang, Z. Application research and progress of microalgae as a novel protein resource in the future. Crit. Rev. Food Sci. Nutr. 2024, 1–24. [Google Scholar] [CrossRef]
- Rather, J.A.; Akhter, N.; Punoo, H.A.; Haddad, M.; Manzoor, N.; Goksen, G.; Dar, B.N. Sustainable algal proteins, novel extraction techniques and applications in the bakery, dairy and pharmaceutical industries: A comprehensive review. Food Chem. 2024, 465, 141828. [Google Scholar] [CrossRef] [PubMed]
- Becker, E.W. Micro-algae as a source of protein. Biotechnol. Adv. 2007, 25, 207–210. [Google Scholar] [CrossRef]
- Podgórska-Kryszczuk, I. Spirulina—An Invaluable Source of Macro-and Micronutrients with Broad Biological Activity and Application Potential. Molecules 2024, 29, 5387. [Google Scholar] [CrossRef]
- Spínola, M.P.; Mendes, A.R.; Prates, J.A. Chemical Composition, Bioactivities, and Applications of Spirulina (Limnospira platensis) in Food, Feed, and Medicine. Foods 2024, 13, 3656. [Google Scholar] [CrossRef]
- Khanra, S.; Mondal, M.; Halder, G.; Tiwari, O.; Gayen, K.; Bhowmick, T.K. Downstream processing of microalgae for pigments, protein and carbohydrate in industrial application: A review. Food Bioprod. Process. 2018, 110, 60–84. [Google Scholar] [CrossRef]
- Siqueira, S.F.; Queiroz, M.I.; Zepka, L.Q.; Jacob-Lopes, E. Introductory chapter: Microalgae biotechnology. A Brief introduction. In Microalgal Biotechnology; IntechOpen: London, UK, 2018; pp. 1–11. [Google Scholar]
- Christaki, E.; Florou-Paneri, P.; Bonos, E. Microalgae: A novel ingredient in nutrition. Int. J. Food Sci. Nutr. 2011, 62, 794–799. [Google Scholar] [CrossRef]
- Apone, F.; Barbulova, A.; Colucci, M.G. Plant and microalgae derived peptides are advantageously employed as bioactive compounds in cosmetics. Front. Plant Sci. 2019, 10, 756. [Google Scholar] [CrossRef]
- Stolz, P.; Obermayer, B. Manufacturing microalgae for skin care. Cosmet. Toilet. 2005, 120, 99–106. [Google Scholar]
- Givens, D.; Gibbs, R. Very long chain n-3 polyunsaturated fatty acids in the food chain in the UK and the potential of animal-derived foods to increase intake. Nutr. Bull. 2006, 31, 104–110. [Google Scholar] [CrossRef]
- Mourelle, M.L.; Gómez, C.P.; Legido, J.L. The potential use of marine microalgae and cyanobacteria in cosmetics and thalassotherapy. Cosmetics 2017, 4, 46. [Google Scholar] [CrossRef]
- Bhalamurugan, G.L.; Valerie, O.; Mark, L. Valuable bioproducts obtained from microalgal biomass and their commercial applications: A review. Environ. Eng. Res. 2018, 23, 229–241. [Google Scholar] [CrossRef]
- Djian, B.; Hornung, E.; Ischebeck, T.; Feussner, I. The green microalga Lobosphaera incisa harbours an arachidonate 15S-lipoxygenase. Plant Biol. 2019, 21, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Galasso, C.; Gentile, A.; Orefice, I.; Ianora, A.; Bruno, A.; Noonan, D.M.; Sansone, C.; Albini, A.; Brunet, C. Microalgal derivatives as potential nutraceutical and food supplements for human health: A focus on cancer prevention and interception. Nutrients 2019, 11, 1226. [Google Scholar] [CrossRef]
- Scaglioni, P.T.; Pagnussatt, F.A.; Lemos, A.C.; Nicolli, C.P.; Del Ponte, E.M.; Badiale-Furlong, E. Nannochloropsis sp. and Spirulina sp. as a source of antifungal compounds to mitigate contamination by Fusarium graminearum species complex. Curr. Microbiol. 2019, 76, 930–938. [Google Scholar] [CrossRef]
- Del Mondo, A.; Smerilli, A.; Ambrosino, L.; Albini, A.; Noonan, D.M.; Sansone, C.; Brunet, C. Insights into phenolic compounds from microalgae: Structural variety and complex beneficial activities from health to nutraceutics. Crit. Rev. Biotechnol. 2021, 41, 155–171. [Google Scholar] [CrossRef]
- Freile-Pelegrín, Y.; Robledo, D. Bioactive phenolic compounds from algae. In Bioactive Compounds from Marine Foods: Plant and Animal Sources; John Wiley and Sons Ltd.: New York, NY, USA, 2013; pp. 113–129. [Google Scholar]
- Nunnery, J.K.; Mevers, E.; Gerwick, W.H. Biologically active secondary metabolites from marine cyanobacteria. Curr. Opin. Biotechnol. 2010, 21, 787–793. [Google Scholar] [CrossRef]
- Boo, Y.C. Can plant phenolic compounds protect the skin from airborne particulate matter? Antioxidants 2019, 8, 379. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Li, X.; Zhang, L.; Xiao, P.; Nie, Y.; Qiu, F.; Cheng, Z.; Li, W.; Zhao, Y. Reactive oxygen species-mediated signal transduction and utilization strategies in microalgae. Bioresour. Technol. 2024, 418, 132004. [Google Scholar] [CrossRef] [PubMed]
- Gullberg, R.C.; Steel, J.J.; Moon, S.L.; Soltani, E.; Geiss, B.J. Oxidative stress influences positive strand RNA virus genome synthesis and capping. Virology 2015, 475, 219–229. [Google Scholar] [CrossRef]
- Karadeniz, F.; Kang, K.-H.; Park, J.W.; Park, S.-J.; Kim, S.-K. Anti-HIV-1 activity of phlorotannin derivative 8,4‴-dieckol from Korean brown alga Ecklonia cava. Biosci. Biotechnol. Biochem. 2014, 78, 1151–1158. [Google Scholar] [CrossRef]
- Zanella, L.; Alam, M.A. Extracts and bioactives from microalgae (sensu stricto): Opportunities and challenges for a new generation of cosmetics. In Microalgae Biotechnology for Food, Health and High Value Products; Springer: Berlin/Heidelberg, Germany, 2020; pp. 295–349. [Google Scholar]
- Coulombier, N.; Jauffrais, T.; Lebouvier, N. Antioxidant compounds from microalgae: A review. Mar. Drugs 2021, 19, 549. [Google Scholar] [CrossRef]
- Miranda, M.; Cintra, R.; Barros, S.B.d.M.; Mancini-Filho, J. Antioxidant activity of the microalga Spirulina maxima. Braz. J. Med. Biol. Res. 1998, 31, 1075–1079. [Google Scholar] [CrossRef]
- El-Baky, H.; El Baz, F.K.; El-Baroty, G.S. Production of phenolic compounds from Spirulina maxima microalgae and its protective effects. Afr. J. Biotechnol. 2009, 8, 7059–7067. [Google Scholar]
- Shalaby, E.A.; Shanab, S.M. Comparison of DPPH and ABTS assays for determining antioxidant potential of water and methanol extracts of Spirulina platensis. Indian J. Geo-Mar. Sci. 2013, 42, 556–564. [Google Scholar]
- Yu, H.; Jia, S.; Dai, Y. Growth characteristics of the cyanobacterium Nostoc flagelliforme in photoautotrophic, mixotrophic and heterotrophic cultivation. J. Appl. Phycol. 2009, 21, 127–133. [Google Scholar] [CrossRef]
- Li, H.-B.; Cheng, K.-W.; Wong, C.-C.; Fan, K.-W.; Chen, F.; Jiang, Y. Evaluation of antioxidant capacity and total phenolic content of different fractions of selected microalgae. Food Chem. 2007, 102, 771–776. [Google Scholar] [CrossRef]
- Hajimahmoodi, M.; Faramarzi, M.A.; Mohammadi, N.; Soltani, N.; Oveisi, M.R.; Nafissi-Varcheh, N. Evaluation of antioxidant properties and total phenolic contents of some strains of microalgae. J. Appl. Phycol. 2010, 22, 43–50. [Google Scholar] [CrossRef]
- Goad, J. Methods in Plant Biochemistry; Charlewood, B.V., Banthorpe, D.V., Eds.; Academic Press: London, UK, 1991; Volume 7. [Google Scholar]
- Fernandes, P.; Cabral, J. Phytosterols: Applications and recovery methods. Bioresour. Technol. 2007, 98, 2335–2350. [Google Scholar] [CrossRef]
- Hernández-Ledesma, B.; Herrero, M. Bioactive Compounds from Marine Foods: Plant and Animal Sources; John Wiley & Sons: Hoboken, NJ, USA, 2013. [Google Scholar]
- Ryckebosch, E.; Bruneel, C.; Termote-Verhalle, R.; Muylaert, K.; Foubert, I. Influence of extraction solvent system on extractability of lipid components from different microalgae species. Algal Res. 2014, 3, 36–43. [Google Scholar] [CrossRef]
- Ahmed, F.; Zhou, W.; Schenk, P.M. Pavlova lutheri is a high-level producer of phytosterols. Algal Res. 2015, 10, 210–217. [Google Scholar] [CrossRef]
- Levasseur, W.; Perré, P.; Pozzobon, V. A review of high value-added molecules production by microalgae in light of the classification. Biotechnol. Adv. 2020, 41, 107545. [Google Scholar] [CrossRef]
- Moreau, R.A.; Whitaker, B.D.; Hicks, K.B. Phytosterols, phytostanols, and their conjugates in foods: Structural diversity, quantitative analysis, and health-promoting uses. Prog. Lipid Res. 2002, 41, 457–500. [Google Scholar] [CrossRef]
- Quilez, J.; Garcia-Lorda, P.; Salas-Salvado, J. Potential uses and benefits of phytosterols in diet: Present situation and future directions. Clin. Nutr. 2003, 22, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Kritchevsky, D.; Chen, S.C. Phytosterols—Health benefits and potential concerns: A review. Nutr. Res. 2005, 25, 413–428. [Google Scholar] [CrossRef]
- Woyengo, T.; Ramprasath, V.; Jones, P. Anticancer effects of phytosterols. Eur. J. Clin. Nutr. 2009, 63, 813–820. [Google Scholar] [CrossRef]
- Kobori, M.; Yoshida, M.; Ohnishi-Kameyama, M.; Shinmoto, H. Ergosterol peroxide from an edible mushroom suppresses inflammatory responses in RAW264. 7 macrophages and growth of HT29 colon adenocarcinoma cells. Br. J. Pharmacol. 2007, 150, 209–219. [Google Scholar] [CrossRef]
- Tang, H.-F.; Yi, Y.-H.; Yao, X.-S.; Xu, Q.-Z.; Zhang, S.-Y.; Lin, H.-W. Bioactive steroids from the brown alga Sargassum carpophyllum. J. Asian Nat. Prod. Res. 2002, 4, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Borowitzka, M.A. Microalgae as sources of pharmaceuticals and other biologically active compounds. J. Appl. Phycol. 1995, 7, 3–15. [Google Scholar] [CrossRef]
- Hamed, I. The evolution and versatility of microalgal biotechnology: A review. Compr. Rev. Food Sci. Food Saf. 2016, 15, 1104–1123. [Google Scholar] [CrossRef] [PubMed]
- García, J.L.; De Vicente, M.; Galán, B. Microalgae, old sustainable food and fashion nutraceuticals. Microb. Biotechnol. 2017, 10, 1017. [Google Scholar] [CrossRef]
- Aizpuru, A.; González-Sánchez, A. Traditional and new trend strategies to enhance pigment contents in microalgae. World J. Microbiol. Biotechnol. 2024, 40, 272. [Google Scholar] [CrossRef]
- Morales-Rivera, F.N.; Torres-Momber, S.; Jacuinde-Ruíz, J.C.; Camacho-Arriaga, J.C.; González-Hernández, J.C. Evaluation of the Effect of Growth Factors on Chlorophylls a and b Production from Microalgae. J. Vis. Exp. 2024, 212, e67208. [Google Scholar]
- Odjadjare, E.C.; Mutanda, T.; Olaniran, A.O. Potential biotechnological application of microalgae: A critical review. Crit. Rev. Biotechnol. 2017, 37, 37–52. [Google Scholar] [CrossRef]
- Yaakob, Z.; Ali, E.; Zainal, A.; Mohamad, M.; Takriff, M.S. An overview: Biomolecules from microalgae for animal feed and aquaculture. J. Biol. Res.-Thessalon. 2014, 21, 1–10. [Google Scholar] [CrossRef]
- Berthon, J.-Y.; Nachat-Kappes, R.; Bey, M.; Cadoret, J.-P.; Renimel, I.; Filaire, E. Marine algae as attractive source to skin care. Free Radic. Res. 2017, 51, 555–567. [Google Scholar] [CrossRef]
- Telfer, A. Too much light? How β-carotene protects the photosystem II reaction centre. Photochem. Photobiol. Sci. 2005, 4, 950–956. [Google Scholar] [CrossRef]
- León, R.; Couso, I.; Fernández, E. Metabolic engineering of ketocarotenoids biosynthesis in the unicelullar microalga Chlamydomonas reinhardtii. J. Biotechnol. 2007, 130, 143–152. [Google Scholar] [CrossRef]
- Shah, F.I.; Imran, H.; Akram, F.; Khalid, T.; Shehzadi, S. Marine Carotenoids: Unlocking Advanced Antioxidant Mechanisms and Therapeutic Applications for Oxidative Stress. Mol. Biotechnol. 2025, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Marcolungo, L.; Bellamoli, F.; Cecchin, M.; Lopatriello, G.; Rossato, M.; Cosentino, E.; Rombauts, S.; Delledonne, M.; Ballottari, M. Haematococcus lacustris genome assembly and annotation reveal diploid genetic traits and stress-induced gene expression patterns. Algal Res. 2024, 80, 103567. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Solovchenko, A.; Neverov, K. Carotenogenic response in photosynthetic organisms: A colorful story. Photosynth. Res. 2017, 133, 31–47. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Ma, T.; Nawaz, M.; Chen, H.; Zheng, H. Advances in Metabolic Engineering for the Accumulation of Astaxanthin Biosynthesis. Mol. Biotechnol. 2024, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.J.; Xie, Q.; Lin, S.; Xu, W.; Cheung, P.C.K. Microalgae-derived astaxanthin: Bioactivities, biotechnological approaches and industrial technologies for its production. Crit. Rev. Food Sci. Nutr. 2025, 1–35. [Google Scholar] [CrossRef] [PubMed]
- Alugoju, P.; Krishna Swamy, V.K.D.; Anthikapalli, N.V.A.; Tencomnao, T. Health benefits of astaxanthin against age-related diseases of multiple organs: A comprehensive review. Crit. Rev. Food Sci. Nutr. 2023, 63, 10709–10774. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Zhu, X.; Yu, X.; Li, S.; Wang, K.; Wei, L.; Li, R.; Qin, S. Advancements of astaxanthin production in Haematococcus pluvialis: Update insight and way forward. Biotechnol. Adv. 2025, 79, 108519. [Google Scholar] [CrossRef] [PubMed]
- Pinto, A.S.; Maia, C.; Sousa, S.A.; Tavares, T.; Pires, J.C.M. Amino Acid and Carotenoid Profiles of Chlorella vulgaris During Two-Stage Cultivation at Different Salinities. Bioengineering 2025, 12, 284. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Li, M.; Gan, Z.; Cui, Y.; Shi, C.; Shi, X. Structure and function characterization of the phytoene desaturase related to the lutein biosynthesis in Chlorella protothecoides CS-41. Mol. Biol. Rep. 2013, 40, 3351–3361. [Google Scholar] [CrossRef]
- Mehariya, S.; Iovine, A.; Di Sanzo, G.; Larocca, V.; Martino, M.; Leone, G.P.; Casella, P.; Karatza, D.; Marino, T.; Musmarra, D.; et al. Supercritical Fluid Extraction of Lutein from Scenedesmus almeriensis. Molecules 2019, 24, 1324. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Rammuni, M.N.; Ariyadasa, T.U.; Nimarshana, P.H.V.; Attalage, R.A. Comparative assessment on the extraction of carotenoids from microalgal sources: Astaxanthin from H. pluvialis and β-carotene from D. salina. Food Chem. 2019, 277, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Budiarso, F.S.; Leong, Y.K.; Chang, J.J.; Chen, C.Y.; Chen, J.H.; Yen, H.W.; Chang, J.S. Current advances in microalgae-based fucoxanthin production and downstream processes. Bioresour. Technol. 2025, 428, 132455. [Google Scholar] [CrossRef] [PubMed]
- Mirza, S.S.; Akbar, S.; Ijaz, M.U.; Morowvat, M.H.; Ishaque, A.; Fatima, K. Nutritional Health Connection of Algae and its Pharmaceutical Value as Anticancer and Antioxidant Constituents of Drugs. Recent Pat. Biotechnol. 2025, 19, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Ye, M.; Zhao, Y. Causal link between dietary antioxidant vitamins intake, oxidative stress injury biomarkers and colorectal cancer: A Mendelian randomization study. Medicine 2025, 104, e41531. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Lee, Z.X.; Guo, H.; Looi, A.D.; Bhuvanendran, S.; Magalingam, K.B.; Lee, W.L.; Radhakrishnan, A.K. Carotenoids Modulate FoxO-Induced Cell Cycle Awrrest in Human Cancer Cell Lines: A Scoping Review. Food Sci. Nutr. 2025, 13, e70100. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Xu, H.; She, P.; Zhao, Z.; Ma, B.; Li, G.; Wang, Y. Duplex Responsive Nanoplatform with Cascade Targeting for Atherosclerosis Photoacoustic Diagnosis and Multichannel Combination Therapy. Adv. Mater. 2023, 35, e2300439. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Lv, M.; Yang, X.; Zhai, S. A Mendelian randomized study of circulating antioxidants in the diet and risk of cardiovascular disease. Sci. Rep. 2025, 15, 10341. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Lu, B.; Ruan, B.; Zhang, J.; Zhang, J.; Liang, Y. Potential Antioxidant and Anti-inflammatory Effects of Astaxanthin Ionic Liquid Liposomes. ACS Appl. Bio. Mater. 2025, 8, 3052–3060. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Geng, X.; Qin, S.; Che, T.; Yan, L.; Yuan, B.; Li, W. Advance on the effects of algal carotenoids on inflammatory signaling pathways. Eur. J. Med. Chem. 2025, 281, 117020. [Google Scholar] [CrossRef] [PubMed]
- Wakuda, N.; Manabe, Y.; Seko, T.; Sugawara, T. Different Suppressive Effects of Canthaxanthin and Fucoxanthin on Cellular Responses of Human Dermal Fibroblasts to Ultraviolet A Irradiation. Biosci. Biotechnol. Biochem. 2025, zbaf056. [Google Scholar] [CrossRef] [PubMed]
- Polamraju, S.M.; Manochkumar, J.; Ganeshbabu, M.; Ramamoorthy, S. Unveiling astaxanthin: Biotechnological advances, delivery systems and versatile applications in nutraceuticals and cosmetics. Arch. Microbiol. 2025, 207, 45. [Google Scholar] [CrossRef] [PubMed]
- Aoki, J.; Koshikawa, R.; Asayama, M. Recent progress in the cyanobacterial products and applications of phycocyanins. World J. Microbiol. Biotechnol. 2025, 41, 84. [Google Scholar] [CrossRef] [PubMed]
- Motamedzadeh, A.; Rahmati-Dehkordi, F.; Heydari, H.; Behnam, M.; Rashidi Noshabad, F.Z.; Tamtaji, Z.; Taheri, A.T.; Nabavizadeh, F.; Aschner, M.; Mirzaei, H.; et al. Therapeutic potential of Phycocyanin in gastrointestinal cancers and related disorders. Mol. Biol. Rep. 2024, 51, 741. [Google Scholar] [CrossRef] [PubMed]
- Kunte, M.; Desai, K. The Inhibitory Effect of C-phycocyanin Containing Protein Extract (C-PC Extract) on Human Matrix Metalloproteinases (MMP-2 and MMP-9) in Hepatocellular Cancer Cell Line (HepG2). Protein J. 2017, 36, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.K.; Li, Y.; Zhai, L.; Tang, Y.; Jiao, Y.; Mei, Y.; Yang, R.; You, R.; Yin, L.; Ni, H.; et al. Natural Phycocyanin/Paclitaxel Micelle Delivery of Therapeutic P53 to Activate Apoptosis for HER2 or ER Positive Breast Cancer Therapy. ACS Biomater. Sci. Eng. 2024, 10, 6995–7004. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, E.; Silva, E.; Figueira, F.S.; Cañedo, A.D.; Machado, K.S.; Salgado, M.T.S.F.; Silva, T.K.; Wagner, E.F.; Mattozo, F.H.; Lima, É.A.; et al. C-phycocyanin to overcome the multidrug resistance phenotype in human erythroleukemias with or without interaction with ABC transporters. Biomed. Pharmacother. 2018, 106, 532–542. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Gao, M.-H.; Chu, X.-M.; Teng, L.; Lv, C.-Y.; Yang, P.; Yin, Q.-F. The synergistic antitumor effects of all-trans retinoic acid and C-phycocyanin on the lung cancer A549 cells in vitro and in vivo. Eur. J. Pharmacol. 2015, 749, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Gardeva, E.; Toshkova, R.; Yossifova, L.; Minkova, K.; Ivanova, N.; Gigova, L. Antitumor activity of C-phycocyanin from Arthronema africanum (Cyanophyceae). Braz. Arch. Biol. Technol. 2014, 57, 675–684. [Google Scholar] [CrossRef]
- de Amarante, M.C.A.; Braga, A.R.C.; Sala, L.; Kalil, S.J. Colour stability and antioxidant activity of C-phycocyanin-added ice creams after in vitro digestion. Food Res. Int. 2020, 137, 109602. [Google Scholar] [CrossRef]
- Mohammadi-Gouraji, E.; Soleimanian-Zad, S.; Ghiaci, M. Phycocyanin-enriched yogurt and its antibacterial and physicochemical properties during 21 days of storage. LWT 2019, 102, 230–236. [Google Scholar] [CrossRef]
- Galetović, A.; Seura, F.; Gallardo, V.; Graves, R.; Cortés, J.; Valdivia, C.; Núñez, J.; Tapia, C.; Neira, I.; Sanzana, S. Use of phycobiliproteins from atacama cyanobacteria as food colorants in a dairy beverage prototype. Foods 2020, 9, 244. [Google Scholar] [CrossRef] [PubMed]
- Bermejo, P.; Piñero, E.; Villar, Á.M. Iron-chelating ability and antioxidant properties of phycocyanin isolated from a protean extract of Spirulina platensis. Food Chem. 2008, 110, 436–445. [Google Scholar] [CrossRef]
- Chaubey, M.G.; Patel, S.N.; Rastogi, R.P.; Srivastava, P.L.; Singh, A.K.; Madamwar, D.; Singh, N.K. Therapeutic potential of cyanobacterial pigment protein phycoerythrin: In silico and in vitro study of BACE1 interaction and in vivo Aβ reduction. Int. J. Biol. Macromol. 2019, 134, 368–378. [Google Scholar] [CrossRef] [PubMed]
- Milledge, J.J. Commercial application of microalgae other than as biofuels: A brief review. Rev. Environ. Sci. Bio./Technol. 2011, 10, 31–41. [Google Scholar] [CrossRef]
- Yu, Z.; Zhao, W.; Sun, H.; Mou, H.; Liu, J.; Yu, H.; Dai, L.; Kong, Q.; Yang, S. Phycocyanin from microalgae: A comprehensive review covering microalgal culture, phycocyanin sources and stability. Food Res. Int. 2024, 186, 114362. [Google Scholar] [CrossRef]
- Miranda, A.M.; Hernandez-Tenorio, F.; Villalta, F.; Vargas, G.J.; Sáez, A.A. Advances in the Development of Biofertilizers and Biostimulants from Microalgae. Biology 2024, 13, 199. [Google Scholar] [CrossRef]
- Sánchez-Quintero, Á.; Fernandes, S.C.; Beigbeder, J.-B. Overview of microalgae and cyanobacteria-based biostimulants produced from wastewater and CO2 streams towards sustainable agriculture: A review. Microbiol. Res. 2023, 277, 127505. [Google Scholar] [CrossRef]
- Chabili, A.; Minaoui, F.; Hakkoum, Z.; Douma, M.; Meddich, A.; Loudiki, M. A comprehensive review of microalgae and cyanobacteria-based biostimulants for agriculture uses. Plants 2024, 13, 159. [Google Scholar] [CrossRef]
- Prisa, D.; Spagnuolo, D. Plant Production with Microalgal Biostimulants. Horticulturae 2023, 9, 829. [Google Scholar] [CrossRef]
- Zelman, A.K.; Berkowitz, G.A. Plant elicitor peptide (Pep) signaling and pathogen defense in tomato. Plants 2023, 12, 2856. [Google Scholar] [CrossRef] [PubMed]
- Kusvuran, S. Microalgae (Chlorella vulgaris Beijerinck) alleviates drought stress of broccoli plants by improving nutrient uptake, secondary metabolites, and antioxidative defense system. Hortic. Plant J. 2021, 7, 221–231. [Google Scholar] [CrossRef]
- Rachidi, F.; Benhima, R.; Kasmi, Y.; Sbabou, L.; Arroussi, H.E. Evaluation of microalgae polysaccharides as biostimulants of tomato plant defense using metabolomics and biochemical approaches. Sci. Rep. 2021, 11, 930. [Google Scholar] [CrossRef]
- Drira, M.; Elleuch, J.; Ben Hlima, H.; Hentati, F.; Gardarin, C.; Rihouey, C.; Le Cerf, D.; Michaud, P.; Abdelkafi, S.; Fendri, I. Optimization of exopolysaccharides production by Porphyridium sordidum and their potential to induce defense responses in Arabidopsis thaliana against Fusarium oxysporum. Biomolecules 2021, 11, 282. [Google Scholar] [CrossRef]
- Augustiniene, E.; Valanciene, E.; Matulis, P.; Syrpas, M.; Jonuskiene, I.; Malys, N. Bioproduction of L-and D-lactic acids: Advances and trends in microbial strain application and engineering. Crit. Rev. Biotechnol. 2022, 42, 342–360. [Google Scholar] [CrossRef]
- Poveda, J.; Díez-Méndez, A. Use of elicitors from macroalgae and microalgae in the management of pests and diseases in agriculture. Phytoparasitica 2023, 51, 667–701. [Google Scholar] [CrossRef]
- Rao, S.R.; Tripathi, U.; Suresh, B.; Ravishankar, G. Enhancement of secondary metabolite production in hairy root cultures of Beta vulgaris and Tagetes patula under the influence of microalgal elicitors. Food Biotechnol. 2001, 15, 35–46. [Google Scholar] [CrossRef]
- Tsintzou, G.; Madesis, P. Peptide Elicitors for Defense Against Abiotic Stress. In Plant Elicitor Peptides: New Tool for Sustainable Agriculture; Springer: Berlin/Heidelberg, Germany, 2024; pp. 19–47. [Google Scholar]
- Mishra, R.; Kaur, P.; Soni, R.; Madan, A.; Agarwal, P.; Singh, G. Decoding the photoprotection strategies and manipulating cyanobacterial photoprotective metabolites, mycosporine-like amino acids, for next-generation sunscreens. Plant Physiol. Biochem. 2024, 212, 108744. [Google Scholar] [CrossRef]
- Cagney, M.H.; O’Neill, E.C. Strategies for producing high value small molecules in microalgae. Plant Physiol. Biochem. 2024, 214, 108942. [Google Scholar] [CrossRef]
- Rosic, N.; Thornber, C. Biotechnological Potential of Macroalgae during Seasonal Blooms for Sustainable Production of UV-Absorbing Compounds. Mar. Drugs 2023, 21, 633. [Google Scholar] [CrossRef]
- Chen, M.; Jiang, Y.; Ding, Y. Recent progress in unraveling the biosynthesis of natural sunscreens mycosporine-like amino acids. J. Ind. Microbiol. Biotechnol. 2023, 50, kuad038. [Google Scholar] [CrossRef] [PubMed]
- Punchakara, A.; Prajapat, G.; Bairwa, H.K.; Jain, S.; Agrawal, A. Applications of mycosporine-like amino acids beyond photoprotection. Appl. Environ. Microbiol. 2023, 89, e0074023. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Guo, F.; Liu, S.; Fang, H.; Xu, Z.; Wang, T. Recent advances and future prospects of mycosporine-like amino acids. Molecules 2023, 28, 5588. [Google Scholar] [CrossRef]
- Schmid, D.; Schürch, C.; Zülli, F.; Nissen, H.-P.; Prieur, H. Mycosporine-like amino acids: Natural UV-screening compounds from red algae to protect the skin against photoaging. SÖFW-J. 2003, 129, 38–42. [Google Scholar]
- Gao, Q.; Garcia-Pichel, F. Microbial ultraviolet sunscreens. Nat. Rev. Microbiol. 2011, 9, 791–802. [Google Scholar] [CrossRef]
- Conde, F.R.; Churio, M.S.; Previtali, C.M. The deactivation pathways of the excited-states of the mycosporine-like amino acids shinorine and porphyra-334 in aqueous solution. Photochem. Photobiol. Sci. 2004, 3, 960–967. [Google Scholar] [CrossRef]
- Llewellyn, C.A.; Greig, C.; Silkina, A.; Kultschar, B.; Hitchings, M.D.; Farnham, G. Mycosporine-like amino acid and aromatic amino acid transcriptome response to UV and far-red light in the cyanobacterium Chlorogloeopsis fritschii PCC 6912. Sci. Rep. 2020, 10, 20638. [Google Scholar] [CrossRef] [PubMed]
- Chaves-Peña, P.; de la Coba, F.; Figueroa, F.L.; Korbee, N. Quantitative and qualitative HPLC analysis of mycosporine-like amino acids extracted in distilled water for cosmetical uses in four Rhodophyta. Mar. Drugs 2019, 18, 27. [Google Scholar] [CrossRef]
- Hartmann, A.; Glaser, K.; Holzinger, A.; Ganzera, M.; Karsten, U. Klebsormidin A and B, two new UV-sunscreen compounds in green microalgal Interfilum and Klebsormidium species (Streptophyta) from terrestrial habitats. Front. Microbiol. 2020, 11, 499. [Google Scholar] [CrossRef]
- Orfanoudaki, M.; Alilou, M.; Hartmann, A.; Mayr, J.; Karsten, U.; Nguyen-Ngoc, H.; Ganzera, M. Isolation and structure elucidation of novel mycosporine-like amino acids from the two intertidal red macroalgae Bostrychia scorpioides and Catenella caespitosa. Mar. Drugs 2023, 21, 543. [Google Scholar] [CrossRef]
- Orfanoudaki, M.; Hartmann, A.; Miladinovic, H.; Nguyen Ngoc, H.; Karsten, U.; Ganzera, M. Bostrychines A–F, six novel mycosporine-like amino-acids and a novel betaine from the red alga Bostrychia scorpioides. Mar. Drugs 2019, 17, 356. [Google Scholar] [CrossRef] [PubMed]
- Raj, S.; Kuniyil, A.M.; Sreenikethanam, A.; Gugulothu, P.; Jeyakumar, R.B.; Bajhaiya, A.K. Microalgae as a source of mycosporine-like amino acids (MAAs); advances and future prospects. Int. J. Environ. Res. Public Health 2021, 18, 12402. [Google Scholar] [CrossRef] [PubMed]
- Salehian, S.; Saadatbakht, M.; Tabarzad, M.; Hosseinabadi, T. Culture optimization to produce high yields of mycosporine-like amino acids by Fischerella sp. F5. Mol. Biotechnol. 2023, 1–11. [Google Scholar] [CrossRef]
- Bloom, D.E.; Cadarette, D. Infectious disease threats in the twenty-first century: Strengthening the global response. Front. Immunol. 2019, 10, 549. [Google Scholar] [CrossRef]
- Roychoudhury, S.; Das, A.; Sengupta, P.; Dutta, S.; Roychoudhury, S.; Choudhury, A.P.; Ahmed, A.F.; Bhattacharjee, S.; Slama, P. Viral pandemics of the last four decades: Pathophysiology, health impacts and perspectives. Int. J. Environ. Res. Public Health 2020, 17, 9411. [Google Scholar] [CrossRef]
- Claus-Desbonnet, H.; Nikly, E.; Nalbantova, V.; Karcheva-Bahchevanska, D.; Ivanova, S.; Pierre, G.; Benbassat, N.; Katsarov, P.; Michaud, P.; Lukova, P. Polysaccharides and their derivatives as potential antiviral molecules. Viruses 2022, 14, 426. [Google Scholar] [CrossRef] [PubMed]
- Carbone, D.A.; Pellone, P.; Lubritto, C.; Ciniglia, C. Evaluation of microalgae antiviral activity and their bioactive compounds. Antibiotics 2021, 10, 746. [Google Scholar] [CrossRef]
- Panggabean, J.A.; Adiguna, S.b.P.; Rahmawati, S.I.; Ahmadi, P.; Zainuddin, E.N.; Bayu, A.; Putra, M.Y. Antiviral activities of algal-based sulfated polysaccharides. Molecules 2022, 27, 1178. [Google Scholar] [CrossRef]
- Zappa, A.; Amendola, A.; Romanò, L.; Zanetti, A. Emerging and re-emerging viruses in the era of globalisation. Blood Transfus. 2009, 7, 167. [Google Scholar]
- Sharon, N. Lectins: Carbohydrate-specific reagents and biological recognition molecules. J. Biol. Chem. 2007, 282, 2753–2764. [Google Scholar] [CrossRef]
- Praseptiangga, D.; Hirayama, M.; Hori, K. Purification, characterization, and cDNA cloning of a novel lectin from the green alga, Codium barbatum. Biosci. Biotechnol. Biochem. 2012, 76, 805–811. [Google Scholar] [CrossRef] [PubMed]
- Hirayama, M.; Shibata, H.; Imamura, K.; Sakaguchi, T.; Hori, K. High-mannose specific lectin and its recombinants from a carrageenophyta Kappaphycus alvarezii represent a potent anti-HIV activity through high-affinity binding to the viral envelope glycoprotein gp120. Mar. Biotechnol. 2016, 18, 144–160. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Morimoto, K.; Kubo, T.; Sakaguchi, T.; Nishizono, A.; Hirayama, M.; Hori, K. Entry inhibition of influenza viruses with high mannose binding lectin ESA-2 from the red alga Eucheuma serra through the recognition of viral hemagglutinin. Mar. Drugs 2015, 13, 3454–3465. [Google Scholar] [CrossRef] [PubMed]
- Huskens, D.; Férir, G.; Vermeire, K.; Kehr, J.-C.; Balzarini, J.; Dittmann, E.; Schols, D. Microvirin, a novel α (1,2)-mannose-specific lectin isolated from Microcystis aeruginosa, has anti-HIV-1 activity comparable with that of cyanovirin-N but a much higher safety profile. J. Biol. Chem. 2010, 285, 24845–24854. [Google Scholar] [CrossRef] [PubMed]
- Bewley, C.A.; Cai, M.; Ray, S.; Ghirlando, R.; Yamaguchi, M.; Muramoto, K. New carbohydrate specificity and HIV-1 fusion blocking activity of the cyanobacterial protein MVL: NMR, ITC and sedimentation equilibrium studies. J. Mol. Biol. 2004, 339, 901–914. [Google Scholar] [CrossRef]
- Boyd, M.R.; Gustafson, K.R.; McMahon, J.B.; Shoemaker, R.H.; O’Keefe, B.R.; Mori, T.; Gulakowski, R.J.; Wu, L.; Rivera, M.I.; Laurencot, C.M. Discovery of cyanovirin-N, a novel human immunodeficiency virus-inactivating protein that binds viral surface envelope glycoprotein gp120: Potential applications to microbicide development. Antimicrob. Agents Chemother. 1997, 41, 1521–1530. [Google Scholar] [CrossRef]
- Bokesch, H.R.; O’Keefe, B.R.; McKee, T.C.; Pannell, L.K.; Patterson, G.M.; Gardella, R.S.; Sowder, R.C.; Turpin, J.; Watson, K.; Buckheit, R.W. A potent novel anti-HIV protein from the cultured cyanobacterium Scytonema varium. Biochemistry 2003, 42, 2578–2584. [Google Scholar] [CrossRef]
- O’Keefe, B.R.; Shenoy, S.R.; Xie, D.; Zhang, W.; Muschik, J.M.; Currens, M.J.; Chaiken, I.; Boyd, M.R. Analysis of the interaction between the HIV-inactivating protein cyanovirin-N and soluble forms of the envelope glycoproteins gp120 and gp41. Mol. Pharmacol. 2000, 58, 982–992. [Google Scholar] [CrossRef]
- Saad, M.H.; El-Fakharany, E.M.; Salem, M.S.; Sidkey, N.M. The use of cyanobacterial metabolites as natural medical and biotechnological tools. J. Biomol. Struct. Dyn. 2022, 40, 2828–2850. [Google Scholar] [CrossRef]
- Mu, J.; Hirayama, M.; Sato, Y.; Morimoto, K.; Hori, K. A novel high-mannose specific lectin from the green alga Halimeda renschii exhibits a potent anti-influenza virus activity through high-affinity binding to the viral hemagglutinin. Mar. Drugs 2017, 15, 255. [Google Scholar] [CrossRef]
- Saad, M.H.; Sidkey, N.M.; Khan, R.H.; El-Fakharany, E.M. Nostoc muscorum is a novel source of microalgal lectins with potent antiviral activity against herpes simplex type-1. Int. J. Biol. Macromol. 2022, 210, 415–429. [Google Scholar] [CrossRef] [PubMed]
- Saad, M.H.; El-Fakharany, E.M.; Salem, M.S.; Sidkey, N.M. In vitro assessment of dual (antiviral and antitumor) activity of a novel lectin produced by the newly cyanobacterium isolate, Oscillatoria acuminate MHM-632 MK014210. 1. J. Biomol. Struct. Dyn. 2022, 40, 3560–3580. [Google Scholar] [CrossRef]
- El-Fakharany, E.M.; Saad, M.H.; Salem, M.S.; Sidkey, N.M. Biochemical characterization and application of a novel lectin from the cyanobacterium Lyngabya confervoides MK012409 as an antiviral and anticancer agent. Int. J. Biol. Macromol. 2020, 161, 417–430. [Google Scholar] [CrossRef] [PubMed]
- Santoyo, S.; Jaime, L.; Plaza, M.; Herrero, M.; Rodriguez-Meizoso, I.; Ibañez, E.; Reglero, G. Antiviral compounds obtained from microalgae commonly used as carotenoid sources. J. Appl. Phycol. 2012, 24, 731–741. [Google Scholar] [CrossRef]
- Fritzsche, S.; Blenk, P.; Christian, J.; Castiglione, K.; Becker, A.M. Inhibitory properties of crude microalgal extracts on the in vitro replication of cyprinid herpesvirus 3. Sci. Rep. 2021, 11, 23134. [Google Scholar] [CrossRef]
- Silva, T.; Salomon, P.S.; Hamerski, L.; Walter, J.; Menezes, R.B.; Siqueira, J.E.; Santos, A.; Santos, J.A.M.; Ferme, N.; Guimarães, T. Inhibitory effect of microalgae and cyanobacteria extracts on influenza virus replication and neuraminidase activity. PeerJ 2018, 6, e5716. [Google Scholar] [CrossRef] [PubMed]
- Baldisserotto, C.; Gentili, V.; Rizzo, R.; Di Donna, C.; Ardondi, L.; Maietti, A.; Pancaldi, S. Characterization of Neochloris oleoabundans under different cultivation modes and first results on bioactivity of its extracts against HCoV-229E virus. Plants 2022, 12, 26. [Google Scholar] [CrossRef]
- Cao, Y.-g.; Hao, Y.; Li, Z.-h.; Liu, S.-t.; Wang, L.-x. Antiviral activity of polysaccharide extract from Laminaria japonica against respiratory syncytial virus. Biomed. Pharmacother. 2016, 84, 1705–1710. [Google Scholar] [CrossRef]
- Elizondo-Gonzalez, R.; Cruz-Suarez, L.E.; Ricque-Marie, D.; Mendoza-Gamboa, E.; Rodriguez-Padilla, C.; Trejo-Avila, L.M. In vitro characterization of the antiviral activity of fucoidan from Cladosiphon okamuranus against Newcastle Disease Virus. Virol. J. 2012, 9, 307. [Google Scholar] [CrossRef]
- Kim, M.; Yim, J.H.; Kim, S.-Y.; Kim, H.S.; Lee, W.G.; Kim, S.J.; Kang, P.-S.; Lee, C.-K. In vitro inhibition of influenza A virus infection by marine microalga-derived sulfated polysaccharide p-KG03. Antivir. Res. 2012, 93, 253–259. [Google Scholar] [CrossRef]
- El-Fakharany, E.M.; Saleh, A.K.; El-Maradny, Y.A.; El-Sayed, M.H.; Alali, I.; Alsirhani, A.M.; Alalawy, A.I.; Alhawiti, A.S.; Alatawi, I.S.; Mazi, W. Comprehensive insight into recent algal enzymes production and purification advances: Toward effective commercial applications: A review. Int. J. Biol. Macromol. 2024, 283, 137783. [Google Scholar] [CrossRef]
- Bozkurt, E.U.; Ørsted, E.C.; Volke, D.C.; Nikel, P.I. Accelerating enzyme discovery and engineering with high-throughput screening. Nat. Prod. Rep. 2024. [Google Scholar] [CrossRef] [PubMed]
- Reisenbauer, J.C.; Sicinski, K.M.; Arnold, F.H. Catalyzing the future: Recent advances in chemical synthesis using enzymes. Curr. Opin. Chem. Biol. 2024, 83, 102536. [Google Scholar] [CrossRef] [PubMed]
- Patil, P.D.; Gargate, N.; Dongarsane, K.; Jagtap, H.; Phirke, A.N.; Tiwari, M.S.; Nadar, S.S. Revolutionizing biocatalysis: A review on innovative design and applications of enzyme-immobilized microfluidic devices. Int. J. Biol. Macromol. 2024, 281, 136193. [Google Scholar] [CrossRef] [PubMed]
- Schober, L.; Schiefer, A.; Winkler, M.; Rudroff, F. Harnessing Nature’s Catalysts: Advances in Enzymatic Alkene Cleavage. J. Biotechnol. 2024, 395, 189–204. [Google Scholar] [CrossRef]
- Mavrommatis, A.; Tsiplakou, E.; Zerva, A.; Pantiora, P.D.; Georgakis, N.D.; Tsintzou, G.P.; Madesis, P.; Labrou, N.E. Microalgae as a sustainable source of antioxidants in animal nutrition, health and livestock development. Antioxidants 2023, 12, 1882. [Google Scholar] [CrossRef]
- Vingiani, G.M.; De Luca, P.; Ianora, A.; Dobson, A.D.W.; Lauritano, C. Microalgal Enzymes with Biotechnological Applications. Mar. Drugs 2019, 17, 459. [Google Scholar] [CrossRef]
- Brasil, B.D.S.A.F.; de Siqueira, F.G.; Salum, T.F.C.; Zanette, C.M.; Spier, M.R. Microalgae and cyanobacteria as enzyme biofactories. Algal Res. 2017, 25, 76–89. [Google Scholar] [CrossRef]
- Navvabi, A.; Homaei, A.; Pletschke, B.I.; Navvabi, N.; Kim, S.K. Marine Cellulases and their Biotechnological Significance from Industrial Perspectives. Curr. Pharm. Des. 2022, 28, 3325–3336. [Google Scholar] [CrossRef]
- Oumer, O. Pectinase: Substrate, Production and their Biotechnological Applications. Int. J. Environ. Agric. Biotechnol. 2017, 2, 1007–1014. [Google Scholar] [CrossRef]
- Amin, F.; Anam Asad, S.; Bilal, M. Chapter 20—Microbial exo-polygalacturonase—A versatile enzyme with multiindustrial applications. In Micro and Nano Technologies, Nanomaterials for Biocatalysis; Castro, G.R., Nadda, A.K., Nguyen, T.A., Qi, X., Yasin, G., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 595–621. [Google Scholar]
- Parte, S.; Sirisha, V.L.; D’Souza, J.S. Chapter Four—Biotechnological Applications of Marine Enzymes From Algae, Bacteria, Fungi, and Sponges. In Advances in Food and Nutrition Research; Kim, S.-K., Toldrá, F., Eds.; Academic Press: Cambridge, MA, USA, 2017; Volume 80, pp. 75–106. [Google Scholar] [CrossRef]
- Spier, M.R.; Schlosser, B.P.; Paludo, L.C.; Gallo-García, A.; Zanette, C.M. Chapter 25—Microalgae as enzymes biofactories. In Handbook of Microalgae-Based Processes and Products; Jacob-Lopes, E., Maroneze, M.M., Queiroz, M.I., Zepka, L.Q., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 687–706. [Google Scholar]
- Sonali; Arora, R. Industrial potential of microbial enzymes. In Microbial Diversity, Interventions and Scope; Springer: Singapore, 2020; pp. 301–318. [Google Scholar]
- Mogharabi, M.; Faramarzi, M.A. Are algae the future source of enzymes? Trends Pept. Protein Sci. 2016, 1, 1–6. [Google Scholar]
- Sikandar, S.; Afzal, I.; Riaz, N.; Ashiq, A. Production of Industrially Important Cellulases and Pectinases using Lignocellulosic Biomass. LGU J. Life Sci. 2023, 7, 537–551. [Google Scholar] [CrossRef]
- Yang, C.-T.; Kristiani, E.; Leong, Y.K.; Chang, J.-S. Machine learning in microalgae biotechnology for sustainable biofuel production: Advancements, applications, and prospects. Bioresour. Technol. 2024, 413, 131549. [Google Scholar] [CrossRef]
- Dalmaso, G.Z.L.; Ferreira, D.; Vermelho, A.B. Marine extremophiles: A source of hydrolases for biotechnological applications. Mar. Drugs 2015, 13, 1925–1965. [Google Scholar] [CrossRef]
- Barzkar, N.; Sukhikh, S.; Babich, O. Study of marine microorganism metabolites: New resources for bioactive natural products. Front. Microbiol. 2024, 14, 1285902. [Google Scholar] [CrossRef]
- Barzkar, N.; Khan, Z.; Jahromi, S.T.; Pourmozaffar, S.; Gozari, M.; Nahavandi, R. A critical review on marine serine protease and its inhibitors: A new wave of drugs? Int. J. Biol. Macromol. 2021, 170, 674–687. [Google Scholar] [CrossRef]
- García, M.M.; de Llasera, M.P.G. A review on the enzymes and metabolites identified by mass spectrometry from bacteria and microalgae involved in the degradation of high molecular weight PAHs. Sci. Total Environ. 2021, 797, 149035. [Google Scholar] [CrossRef] [PubMed]
- Anwar, M.; Lou, S.; Chen, L.; Li, H.; Hu, Z. Recent advancement and strategy on bio-hydrogen production from photosynthetic microalgae. Bioresour. Technol. 2019, 292, 121972. [Google Scholar] [CrossRef]
- González-Fernández, C.; Ballesteros, M. Linking microalgae and cyanobacteria culture conditions and key-enzymes for carbohydrate accumulation. Biotechnol. Adv. 2012, 30, 1655–1661. [Google Scholar] [CrossRef]
- Narang, P.K.; Dey, J.; Mahapatra, S.R.; Roy, R.; Kushwaha, G.S.; Misra, N.; Suar, M.; Raina, V. Genome-based identification and comparative analysis of enzymes for carotenoid biosynthesis in microalgae. World J. Microbiol. Biotechnol. 2022, 38, 8. [Google Scholar] [CrossRef]
- Alves, P.L.M.; Nieri, V.; Moreli, F.d.C.; Constantino, E.; de Souza, J.; Oshima-Franco, Y.; Grotto, D. Unveiling New Horizons: Advancing Technologies in Cosmeceuticals for Anti-Aging Solutions. Molecules 2024, 29, 4890. [Google Scholar] [CrossRef]
- Zhang, K.; Han, Y. Thermostable Bacterial Collagenolytic Proteases: A Review. J. Microbiol. Biotechnol. 2024, 34, 1385. [Google Scholar] [CrossRef] [PubMed]
- Koroleva, V.; Lavlinskaya, M.; Holyavka, M.; Penkov, N.; Zuev, Y.; Artyukhov, V. Thermal Inactivation, Denaturation and Aggregation Processes of Papain Like Proteases. Chem. Biodivers. 2024, 21, e202401038. [Google Scholar] [CrossRef]
- Mousavi Ghahfarrokhi, S.S.; Mahdigholi, F.S.; Amin, M. Collateral beauty in the damages: An overview of cosmetics and therapeutic applications of microbial proteases. Arch. Microbiol. 2023, 205, 375. [Google Scholar] [CrossRef] [PubMed]
- Del Rosso, J.Q. Application of protease technology in dermatology: Rationale for incorporation into skin care with initial observations on formulations designed for skin cleansing, maintenance of hydration, and restoration of the epidermal permeability barrier. J. Clin. Aesthetic Dermatol. 2013, 6, 14. [Google Scholar]
- Trevisol, T.C.; Henriques, R.O.; Souza, A.J.A.; Furigo, A., Jr. An overview of the use of proteolytic enzymes as exfoliating agents. J. Cosmet. Dermatol. 2022, 21, 3300–3307. [Google Scholar] [CrossRef]
- Trevisol, T.C.; Henriques, R.O.; Cesca, K.; Souza, A.J.A.; Furigo, A., Jr. In vitro effect on the proteolytic activity of papain with proteins of the skin as substrate. Int. J. Cosmet. Sci. 2022, 44, 542–554. [Google Scholar] [CrossRef]
- Ataide, J.A.; de Carvalho, N.M.; Rebelo, M.d.A.; Chaud, M.V.; Grotto, D.; Gerenutti, M.; Rai, M.; Mazzola, P.G.; Jozala, A.F. Bacterial nanocellulose loaded with bromelain: Assessment of antimicrobial, antioxidant and physical-chemical properties. Sci. Rep. 2017, 7, 18031. [Google Scholar] [CrossRef] [PubMed]
- Artem Ataide, J.; Caramori Cefali, L.; Machado Croisfelt, F.; Arruda Martins Shimojo, A.; Oliveira Nascimento, L.; Gava Mazzola, P. Natural actives for wound healing: A review. Phytother. Res. 2018, 32, 1664–1674. [Google Scholar] [CrossRef]
- Ioannou, E.; Labrou, N.E. Development of enzyme-based cosmeceuticals: Studies on the proteolytic activity of Arthrospira platensis and its efficient incorporation in a hydrogel formulation. Cosmetics 2022, 9, 106. [Google Scholar] [CrossRef]
- Cirulis, J.T.; Scott, J.A.; Ross, G.M. Management of oxidative stress by microalgae. Can. J. Physiol. Pharmacol. 2013, 91, 15–21. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Osathanunkul, M.; Thanaporn, S.; Karapetsi, L.; Nteve, G.M.; Pratsinakis, E.; Stefanidou, E.; Lagiotis, G.; Avramidou, E.; Zorxzobokou, L.; Tsintzou, G.; et al. Diversity of Bioactive Compounds in Microalgae: Key Classes and Functional Applications. Mar. Drugs 2025, 23, 222. https://doi.org/10.3390/md23060222
Osathanunkul M, Thanaporn S, Karapetsi L, Nteve GM, Pratsinakis E, Stefanidou E, Lagiotis G, Avramidou E, Zorxzobokou L, Tsintzou G, et al. Diversity of Bioactive Compounds in Microalgae: Key Classes and Functional Applications. Marine Drugs. 2025; 23(6):222. https://doi.org/10.3390/md23060222
Chicago/Turabian StyleOsathanunkul, Maslin, Suebsuya Thanaporn, Lefkothea Karapetsi, Georgia Maria Nteve, Emmanouil Pratsinakis, Eleni Stefanidou, Giorgos Lagiotis, Eleni Avramidou, Lydia Zorxzobokou, Georgia Tsintzou, and et al. 2025. "Diversity of Bioactive Compounds in Microalgae: Key Classes and Functional Applications" Marine Drugs 23, no. 6: 222. https://doi.org/10.3390/md23060222
APA StyleOsathanunkul, M., Thanaporn, S., Karapetsi, L., Nteve, G. M., Pratsinakis, E., Stefanidou, E., Lagiotis, G., Avramidou, E., Zorxzobokou, L., Tsintzou, G., Athanasiou, A., Mpelai, S., Constandinidis, C., Pantiora, P., Merino, M., Mullor, J. L., Dobrovic, L., Cerasino, L., Ogawa, T., ... Madesis, P. (2025). Diversity of Bioactive Compounds in Microalgae: Key Classes and Functional Applications. Marine Drugs, 23(6), 222. https://doi.org/10.3390/md23060222