

Codium fragile Extract Ameliorates Respiratory Function by Controlling Allergic Inflammation in Ovalbumin-Induced Bronchial Disorders in Mice

Abstract

1. Introduction
2. Results
2.1. High-Performance Liquid Chromatography with Photodiode Array Detection (HPLC-PDA) Analysis
2.1.1. Quantitative Analysis of Oleamide in WCF
2.1.2. Method Validation
2.2. Effect of WCF on Activation of T Cells in Whole Blood of OVA-Induced Mice
2.3. Effect of WCF on Allergic Inflammatory Cytokines in Lung Tissues of OVA-Induced Mice
2.4. Effect of WCF on Number of Leukocytes in Bronchoalveolar Lavage Fluid (BALF) of OVA-Induced Mice
2.5. Effect of WCF on OVA-Specific IgE Levels in BALF and Serum of OVA-Induced Mice
2.6. Effect of WCF on Histopathological Changes in Lung Tissue of OVA-Induced Mice
2.7. Effect of WCF on Levels of Antioxidant System in Lung Tissues of OVA-Induced Mice
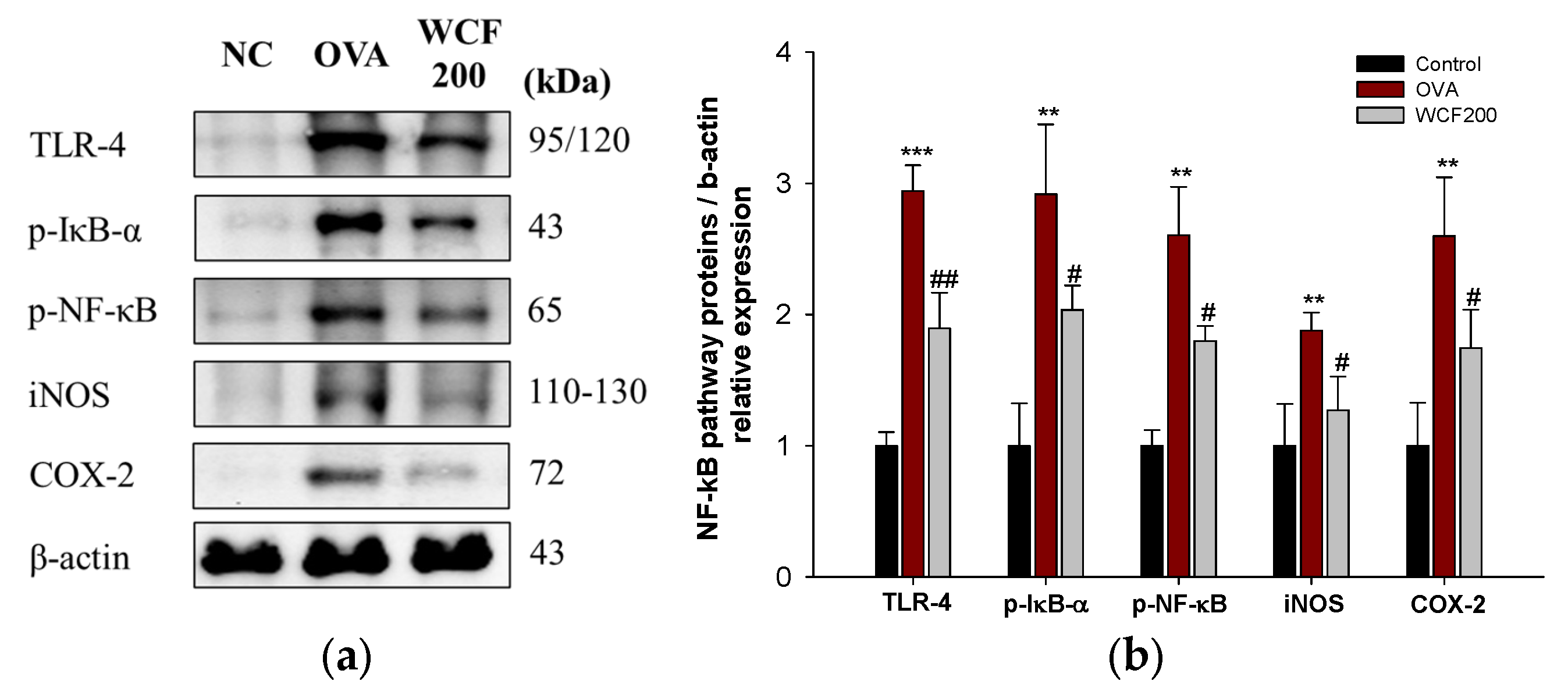
2.8. Effect of WCF on Activation of Nuclear Factor kB (NF-κB) Pathway in Lung Tissues of OVA-Induced Mice
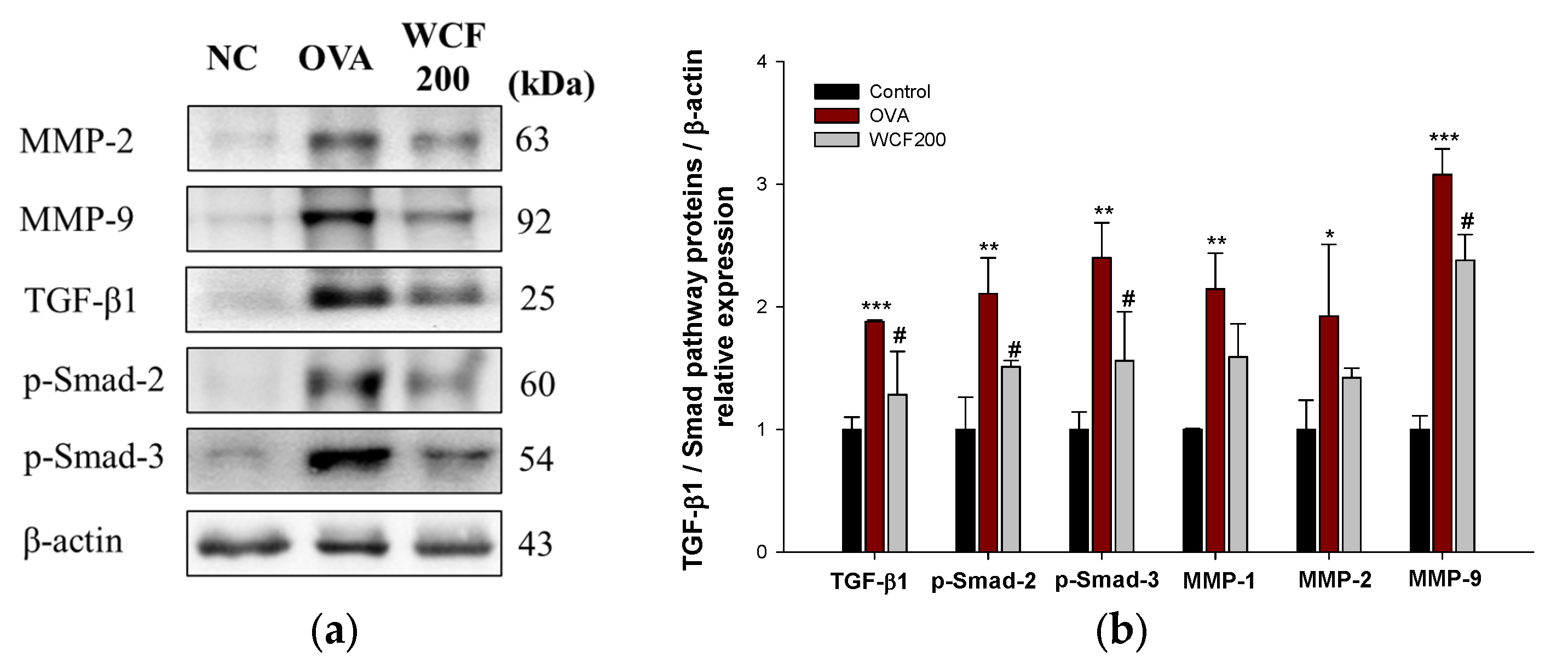
2.9. Effect of WCF on Activation of Transforming Growth Factor β (TGF-β)/Smad Pathway in Lung Tissues of OVA-Induced Mice
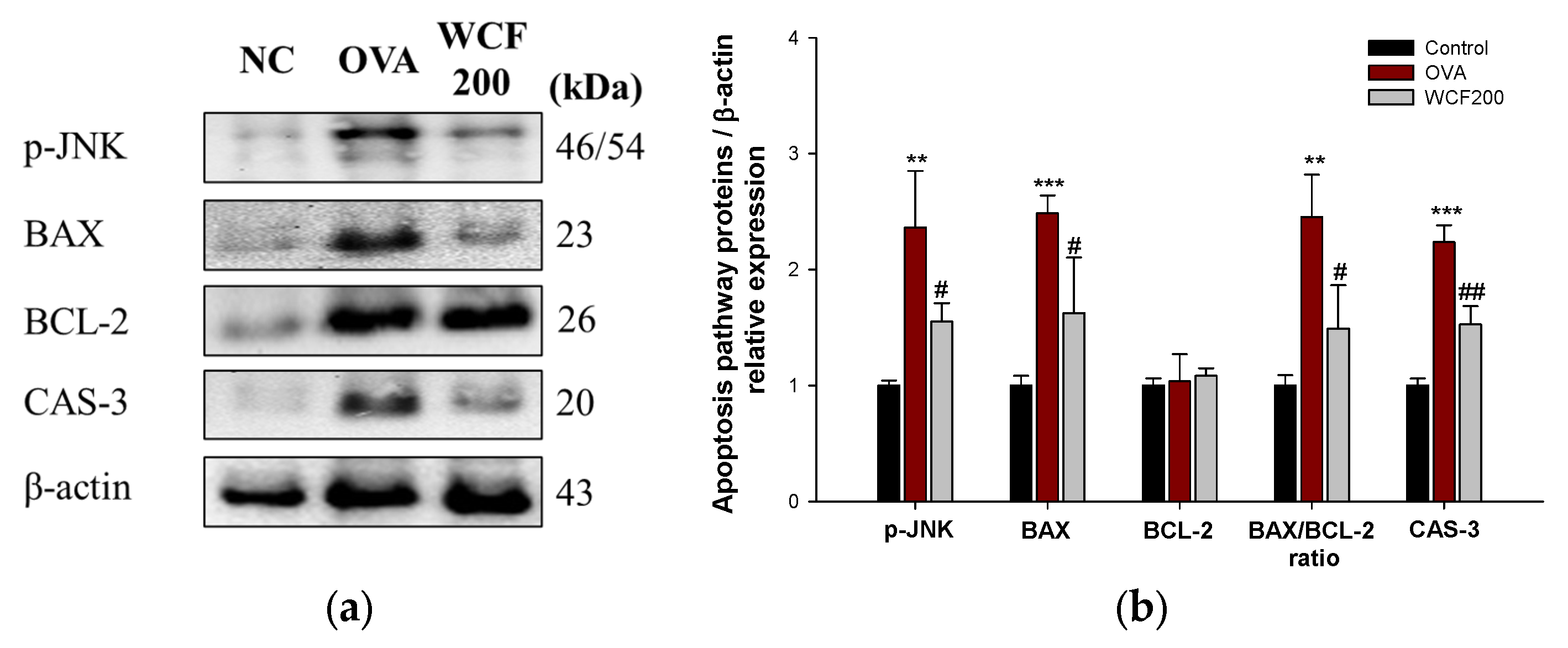
2.10. Effect of WCF on Activation of Apoptosis Pathway in Lung Tissues of OVA-Induced Mice
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Preparation of WCF
4.3. HPLC with Photodiode Array Detection (HPLC-PDA) Analysis
4.4. Method Validation
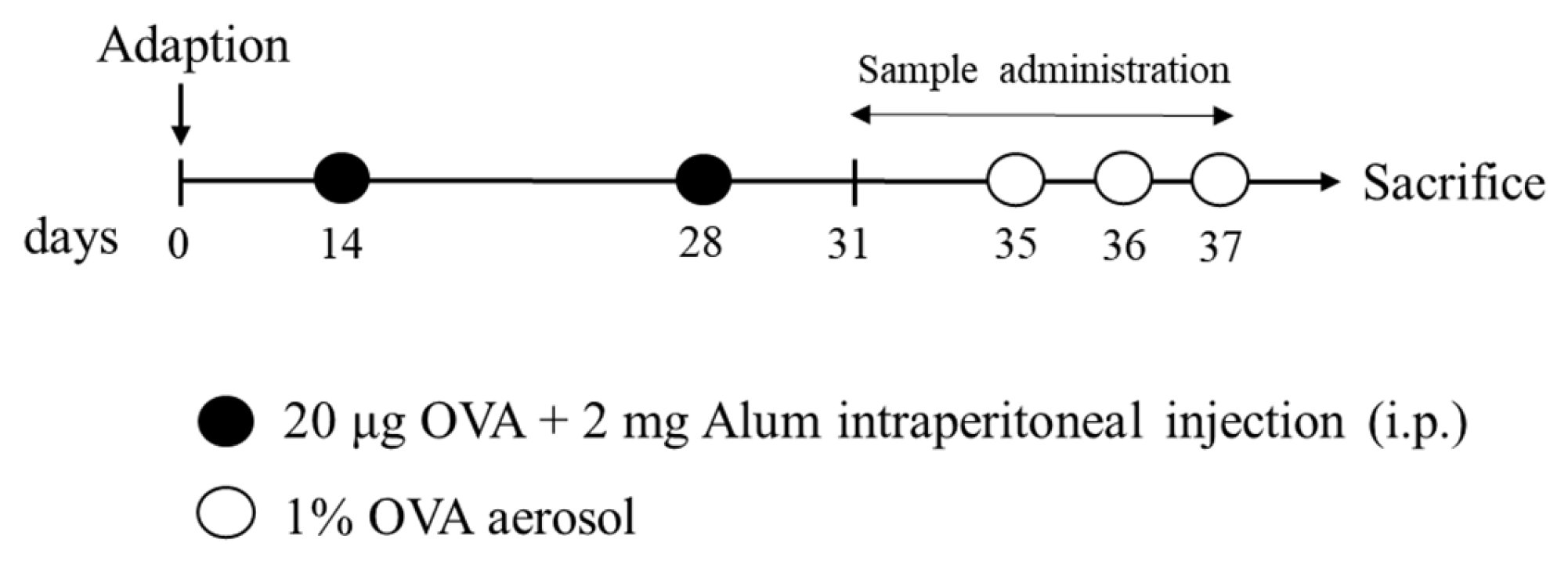
4.5. Animal Experiment Design
4.5.1. Animals
4.5.2. Immunization and Treatment
4.6. Flow Cytometry Fluorescence-Activated Cell Sorting (FACS) Analysis
4.7. Cell Counting in BALF
4.8. OVA-Specific IgE Level
4.9. Histopathological Analysis
4.10. Measurement of SOD, Reduced GSH, and MDA Levels
4.11. Western Blotting
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Labaki, W.W.; Han, M.K. Chronic respiratory diseases: A global view. Lancet Respir. Med. 2020, 8, 531–533. [Google Scholar] [CrossRef]
- Dey, S.; Eapen, M.S.; Chia, C.; Gaikwad, A.V.; Wark, P.A.; Sohal, S.S. Pathogenesis, clinical features of asthma COPD overlap, and therapeutic modalities. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2022, 322, L64–L83. [Google Scholar] [CrossRef]
- Witt, A.; Douglass, J.A.; Harun, N.S. Overview of recent advancements in asthma management. Intern. Med. J. 2022, 52, 1478–1487. [Google Scholar] [CrossRef]
- Hammad, H.; Lambrecht, B.N. The basic immunology of asthma. Cell 2021, 184, 1469–1485. [Google Scholar] [CrossRef]
- Wang, M.-C.; Huang, W.-C.; Chen, L.-C.; Yeh, K.-W.; Lin, C.-F.; Liou, C.-J. Sophoraflavanone G from Sophora flavescens ameliorates allergic airway inflammation by suppressing Th2 response and oxidative stress in a murine asthma model. Int. J. Mol. Sci. 2022, 23, 6104. [Google Scholar] [CrossRef]
- Timalsina, D.; Pokhrel, K.P.; Bhusal, D. Pharmacologic activities of plant-derived natural products on respiratory diseases and inflammations. BioMed Res. Int. 2021, 2021, 1636816. [Google Scholar] [CrossRef]
- Jasemi, S.V.; Khazaei, H.; Morovati, M.R.; Joshi, T.; Aneva, I.Y.; Farzaei, M.H.; Echeverría, J. Phytochemicals as treatment for allergic asthma: Therapeutic effects and mechanisms of action. Phytomedicine 2024, 122, 155149. [Google Scholar] [CrossRef]
- Agache, I.; Rocha, C.; Beltran, J.; Song, Y.; Posso, M.; Solà, I.; Jutel, M. Efficacy and safety of treatment with biologicals (benralizumab, dupilumab and omalizumab) for severe allergic asthma: A systematic review for the EAACI Guidelines-recommendations on the use of biologicals in severe asthma. Allergy 2020, 75, 1043–1057. [Google Scholar] [CrossRef]
- Ramírez-Jiménez, F.; Pavón-Romero, G.F.; Velásquez-Rodríguez, J.M.; López-Garza, M.I.; Lazarini-Ruiz, J.F.; Gutiérrez-Quiroz, K.V.; Teran, L.M. Biologic therapies for asthma and allergic disease: Past, present, and future. Pharmaceuticals 2023, 16, 270. [Google Scholar] [CrossRef]
- Amaral-Machado, L.; Oliveira, W.N.; Moreira-Oliveira, S.S.; Pereira, D.T.; Alencar, E.N.; Tsapis, N.; Egito, E.S.T. Use of natural products in asthma treatment. Evid.-Based Complement. Altern. Med. 2020, 2020, 1021258. [Google Scholar] [CrossRef]
- Karthikeyan, A.; Joseph, A.; Nair, B.G. Promising bioactive compounds from the marine environment and their potential effects on various diseases. J. Genet. Eng. Biotechnol. 2022, 20, 14. [Google Scholar] [CrossRef]
- Kim, T.Y.; Kim, J.M.; Lee, H.L.; Go, M.J.; Joo, S.G.; Kim, J.H.; Lee, H.S.; Jeong, W.M.; Lee, D.Y.; Kim, H.-J. Codium fragile suppressed chronic pm2.5-exposed pulmonary dysfunction via TLR/TGF-β pathway in BALB/c mice. Antioxidants 2023, 12, 1743. [Google Scholar] [CrossRef]
- Jang, A.-y.; Choi, J.; Rod-In, W.; Choi, K.Y.; Lee, D.-H.; Park, W.J. In vitro anti-inflammatory and skin protective effects of Codium fragile extract on macrophages and human keratinocytes in atopic dermatitis. J. Microbiol. Biotechnol. 2024, 34, 940. [Google Scholar] [CrossRef]
- Lee, C.; Park, G.H.; Ahn, E.M.; Kim, B.-A.; Park, C.-I.; Jang, J.-H. Protective effect of Codium fragile against UVB-induced pro-inflammatory and oxidative damages in HaCaT cells and BALB/c mice. Fitoterapia 2013, 86, 54–63. [Google Scholar] [CrossRef]
- Akarsha, B.; Krishnakumar, G.; Khandige, P.S. Hepatoprotective potency of Lagenandra toxicaria and Ariopsis peltata against CCl4 induced liver fibrosis in Wistar rats. Plant Sci. Today 2022, 9, 855–866. [Google Scholar] [CrossRef]
- Meinita, M.D.N.; Harwanto, D.; Choi, J.-S. A concise review of the bioactivity and pharmacological properties of the genus Codium (Bryopsidales, Chlorophyta). J. Appl. Phycol. 2022, 34, 2827–2845. [Google Scholar] [CrossRef]
- Moon, S.-M.; Lee, S.A.; Hong, J.H.; Kim, J.-S.; Kim, D.K.; Kim, C.S. Oleamide suppresses inflammatory responses in LPS-induced RAW264.7 murine macrophages and alleviates paw edema in a carrageenan-induced inflammatory rat model. Int. Immunopharmacol. 2018, 56, 179–185. [Google Scholar] [CrossRef]
- AOAC INTERNATIONAL. AOAC Official Methods of Analysis; Appendix F: Guidelines for Standard Method Performance Requirements; AOAC INTERNATIONAL: Rockville, MD, USA, 2016; pp. 1–17. [Google Scholar]
- Shen, Q.-Y.; Wu, L.; Wei, C.-S.; Zhou, Y.-N.; Wu, H.-M. Sevoflurane prevents airway remodeling via downregulation of VEGF and TGF-β1 in mice with OVA-induced chronic airway inflammation. Inflammation 2019, 42, 1015–1022. [Google Scholar] [CrossRef]
- Lee, S.A.; Moon, S.-M.; Choi, Y.H.; Han, S.H.; Park, B.-R.; Choi, M.S.; Kim, J.-S.; Kim, Y.H.; Kim, D.K.; Kim, C.S. Aqueous extract of Codium fragile suppressed inflammatory responses in lipopolysaccharide-stimulated RAW264.7 cells and carrageenan-induced rats. Biomed. Pharmacother. 2017, 93, 1055–1064. [Google Scholar] [CrossRef]
- Awakan, O.J.; Malomo, S.O.; Adejare, A.A.; Igunnu, A.; Atolani, O.; Adebayo, A.H.; Owoyele, B.V. Anti-inflammatory and bronchodilatory constituents of leaf extracts of Anacardium occidentale L. in animal models. J. Integr. Med. 2018, 16, 62–70. [Google Scholar] [CrossRef]
- Yi, L.; Zhou, Y.; Song, J.; Tang, W.; Yu, H.; Huang, X.; Shi, H.; Chen, M.; Sun, J.; Wei, Y. A novel iridoid glycoside leonuride (ajugol) attenuates airway inflammation and remodeling through inhibiting type-2 high cytokine/chemokine activity in OVA-induced asthmatic mice. Phytomedicine 2022, 105, 154345. [Google Scholar] [CrossRef] [PubMed]
- Gieseck III, R.L.; Wilson, M.S.; Wynn, T.A. Type 2 immunity in tissue repair and fibrosis. Nat. Rev. Immunol. 2018, 18, 62–76. [Google Scholar] [CrossRef]
- Dominguez-Flores, A.; López, G.M.R.; Soria-Castro, R.; López-Santiago, R.; Rodríguez-Cortés, O.; Pérez-Tapia, S.M.; Chávez-Blanco, A.D.; Estrada-Parra, S.; Flores-Mejía, R.; Chacón-Salinas, R. Brucella abortus induces mast cell activation through TLR-2 and TLR-4. Microb. Pathog. 2023, 176, 106005. [Google Scholar] [CrossRef] [PubMed]
- Helal, M.G.; Megahed, N.A.; Abd Elhameed, A.G. Saxagliptin mitigates airway inflammation in a mouse model of acute asthma via modulation of NF-κB and TLR4. Life Sci. 2019, 239, 117017. [Google Scholar] [CrossRef] [PubMed]
- Fatani, S.H. Biomarkers of oxidative stress in acute and chronic bronchial asthma. J. Asthma 2014, 51, 578–584. [Google Scholar] [CrossRef]
- Zou, S.; Hong, J.; Liu, D.; Lai, G.; Ye, J.; Song, Y. Enhanced effect of catalpol on specific immune therapy in treatment of asthmatic mice. Am. J. Transl. Res. 2019, 11, 2463. [Google Scholar]
- Mahalanobish, S.; Saha, S.; Dutta, S.; Sil, P.C. Matrix metalloproteinase: An upcoming therapeutic approach for idiopathic pulmonary fibrosis. Pharmacol. Res. 2020, 152, 104591. [Google Scholar] [CrossRef]
- Chen, H.; Chen, Q.; Jiang, C.-m.; Shi, G.-Y.; Sui, B.-w.; Zhang, W.; Yang, L.-z.; Li, Z.-y.; Liu, L.; Su, Y.-m. Triptolide suppresses paraquat induced idiopathic pulmonary fibrosis by inhibiting TGFB1-dependent epithelial mesenchymal transition. Toxicol. Lett. 2018, 284, 1–9. [Google Scholar] [CrossRef]
- Sutherland, T.E.; Dyer, D.P.; Allen, J.E. The extracellular matrix and the immune system: A mutually dependent relationship. Science 2023, 379, eabp8964. [Google Scholar] [CrossRef]
- Lee, H.L.; Kim, J.M.; Go, M.J.; Kim, T.Y.; Joo, S.G.; Kim, J.H.; Lee, H.S.; Kim, H.-J.; Heo, H.J. Protective effect of Lonicera japonica on PM2.5-induced pulmonary damage in BALB/c mice via the TGF-β and NF-κB pathway. Antioxidants 2023, 12, 968. [Google Scholar] [CrossRef]
- Shang, Q.; Zhu, L.; Shang, W.; Zeng, J.; Qi, Y. Dioscin exhibits protective effects on in vivo and in vitro asthma models via suppressing TGF-β1/Smad2/3 and AKT pathways. J. Biochem. Mol. Toxicol. 2022, 36, e23084. [Google Scholar] [CrossRef] [PubMed]
- Drakopanagiotakis, F.; Xifteri, A.; Polychronopoulos, V.; Bouros, D. Apoptosis in lung injury and fibrosis. Eur. Respir. J. 2008, 32, 1631–1638. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.-H.; Kim, J.-E.; Shim, J.-H.; Yoon, G.; Bang, M.-A.; Bae, C.-S.; Lee, K.-J.; Park, D.-H.; Cho, S.-S. HPLC analysis, optimization of extraction conditions and biological evaluation of Corylopsis coreana Uyeki Flos. Molecules 2016, 21, 94. [Google Scholar] [CrossRef]
- Lee, J.-A.; Lee, M.-Y.; Seo, C.-S.; Jung, D.Y.; Lee, N.-H.; Kim, J.-H.; Ha, H.; Shin, H.K. Anti-asthmatic effects of an Amomum compactum extract on an ovalbumin (OVA)-induced murine asthma model. Biosci. Biotechnol. Biochem. 2010, 74, 1814–1818. [Google Scholar] [CrossRef]
- Kim, D.I.; Song, M.-K.; Lee, K. Comparison of asthma phenotypes in OVA-induced mice challenged via inhaled and intranasal routes. BMC Pulm. Med. 2019, 19, 241. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Oleamide (9-octadecenamide) | ||
|---|---|---|---|
| Linearity range (μg/mL) | 5–100 | ||
| Regression equation | y = 0.6112x + 0.4277 | ||
| Correlation coefficient (R2) | 0.9998 | ||
| Intra-day precision (%) 1 | 0.14 | ||
| Inter-day precision (%) 1 | 0.84 | ||
| LOD (μg/mL) 2 | 0.37 ± 0.00 | ||
| LOQ (μg/mL) 2 | 1.12 ± 0.00 | ||
| Recovery rate (%) 3 | Concentration (μg/mL) | ||
| 5 | 10 | 20 | |
| 96.64 ± 1.10 | 99.62 ± 0.91 | 100.72 ± 1.10 | |
| Groups | Total Leukocytes | Leukocyte Classification Unit: 105/mL | |||
|---|---|---|---|---|---|
| Eosinophils | Lymphocytes | Neutrophils | Monocytes | ||
| Control | 2.70 ± 0.41 | N.D. | N.D. | N.D. | 2.67 ± 0.38 |
| OVA | 7.64 ± 0.43 *** | 3.31 ± 0.88 *** | 1.15 ± 0.19 *** | 0.86 ± 0.31 *** | 2.30 ± 0.43 |
| WCF50 | 5.26 ± 0.56 ## | 1.62 ± 0.23 ## | 0.68 ± 0.19 ## | 0.68 ± 0.22 | 2.29 ± 0.25 |
| WCF100 | 5.16 ± 1.94 # | 1.96 ± 0.84 # | 0.64 ± 0.20 ## | 0.56 ± 0.22 | 2.13 ± 1.00 |
| WCF200 | 5.30 ± 0.82 # | 1.76 ± 0.25 ## | 0.63 ± 0.12 ### | 0.42 ± 0.09 # | 2.50 ± 0.46 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.L.; Ju, Y.H.; Kim, I.Y.; Choi, H.J.; Heo, Y.M.; Na, H.R.; Heo, H.J. Codium fragile Extract Ameliorates Respiratory Function by Controlling Allergic Inflammation in Ovalbumin-Induced Bronchial Disorders in Mice. Mar. Drugs 2025, 23, 221. https://doi.org/10.3390/md23050221
Lee HL, Ju YH, Kim IY, Choi HJ, Heo YM, Na HR, Heo HJ. Codium fragile Extract Ameliorates Respiratory Function by Controlling Allergic Inflammation in Ovalbumin-Induced Bronchial Disorders in Mice. Marine Drugs. 2025; 23(5):221. https://doi.org/10.3390/md23050221
Chicago/Turabian StyleLee, Hyo Lim, Yeong Hyeon Ju, In Young Kim, Hye Ji Choi, Yu Mi Heo, Hwa Rang Na, and Ho Jin Heo. 2025. "Codium fragile Extract Ameliorates Respiratory Function by Controlling Allergic Inflammation in Ovalbumin-Induced Bronchial Disorders in Mice" Marine Drugs 23, no. 5: 221. https://doi.org/10.3390/md23050221
APA StyleLee, H. L., Ju, Y. H., Kim, I. Y., Choi, H. J., Heo, Y. M., Na, H. R., & Heo, H. J. (2025). Codium fragile Extract Ameliorates Respiratory Function by Controlling Allergic Inflammation in Ovalbumin-Induced Bronchial Disorders in Mice. Marine Drugs, 23(5), 221. https://doi.org/10.3390/md23050221