
Amelioration of Particulate Matter-Induced Oxidative Stress by a Bioactive Hizikia fusiformis Extract: A Functional Biomaterial for Cosmeceutical Applications

 , , , , and
, , , , and
Abstract
1. Introduction
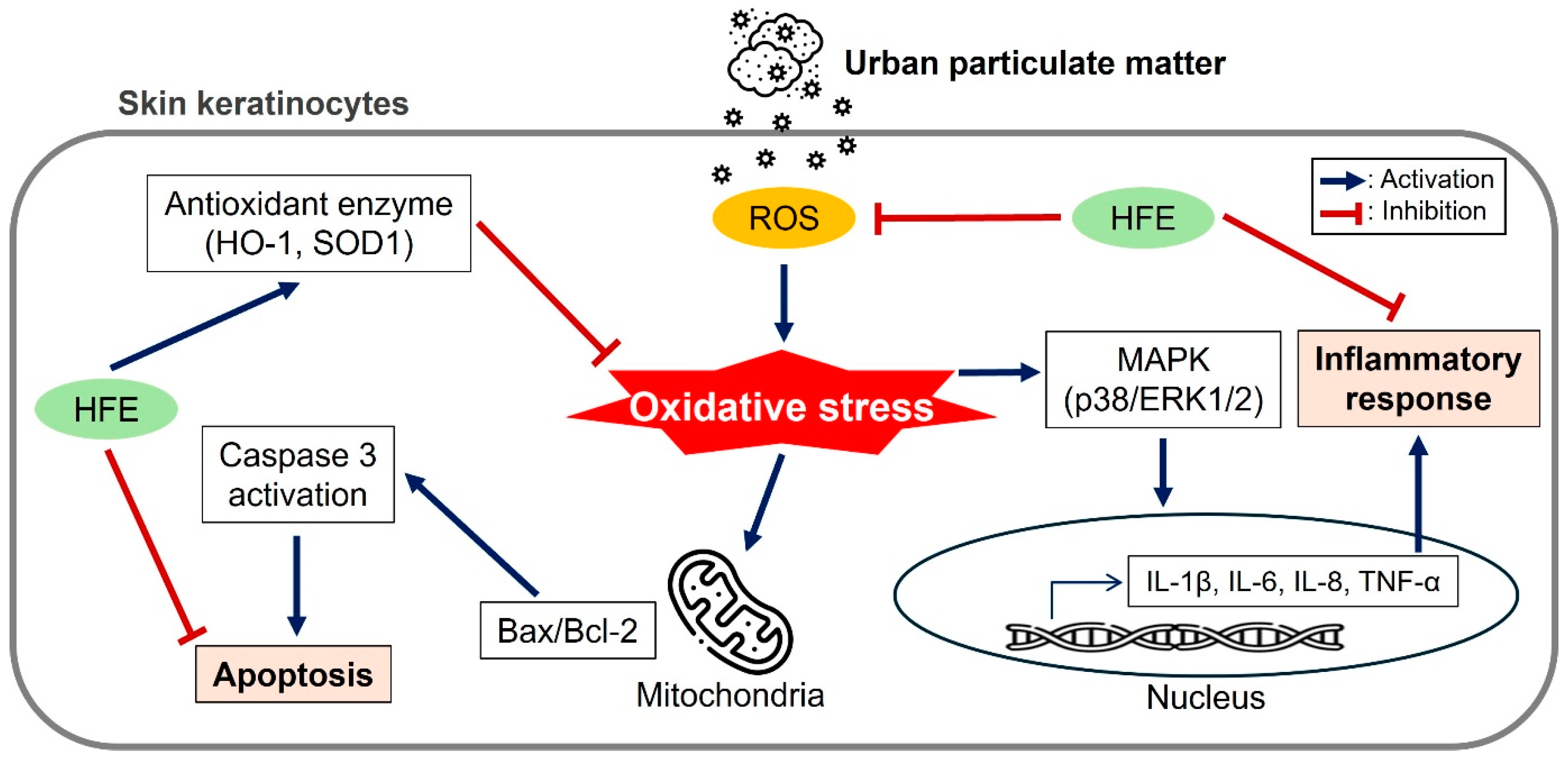
2. Results and Discussion
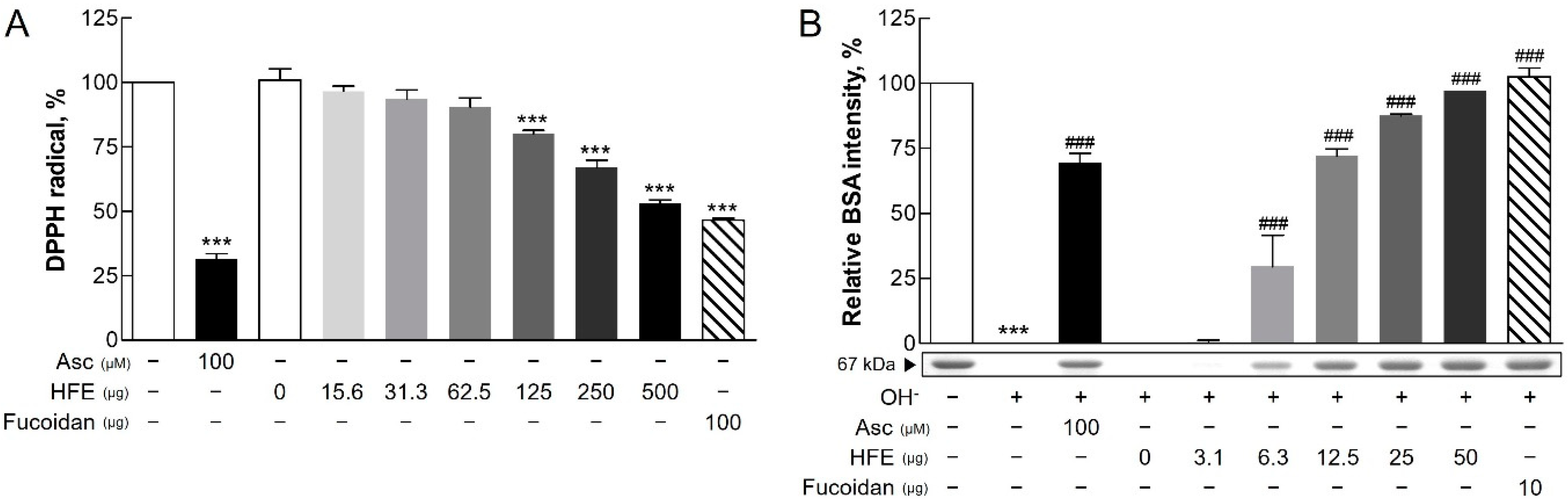
2.1. Antioxidant Activities
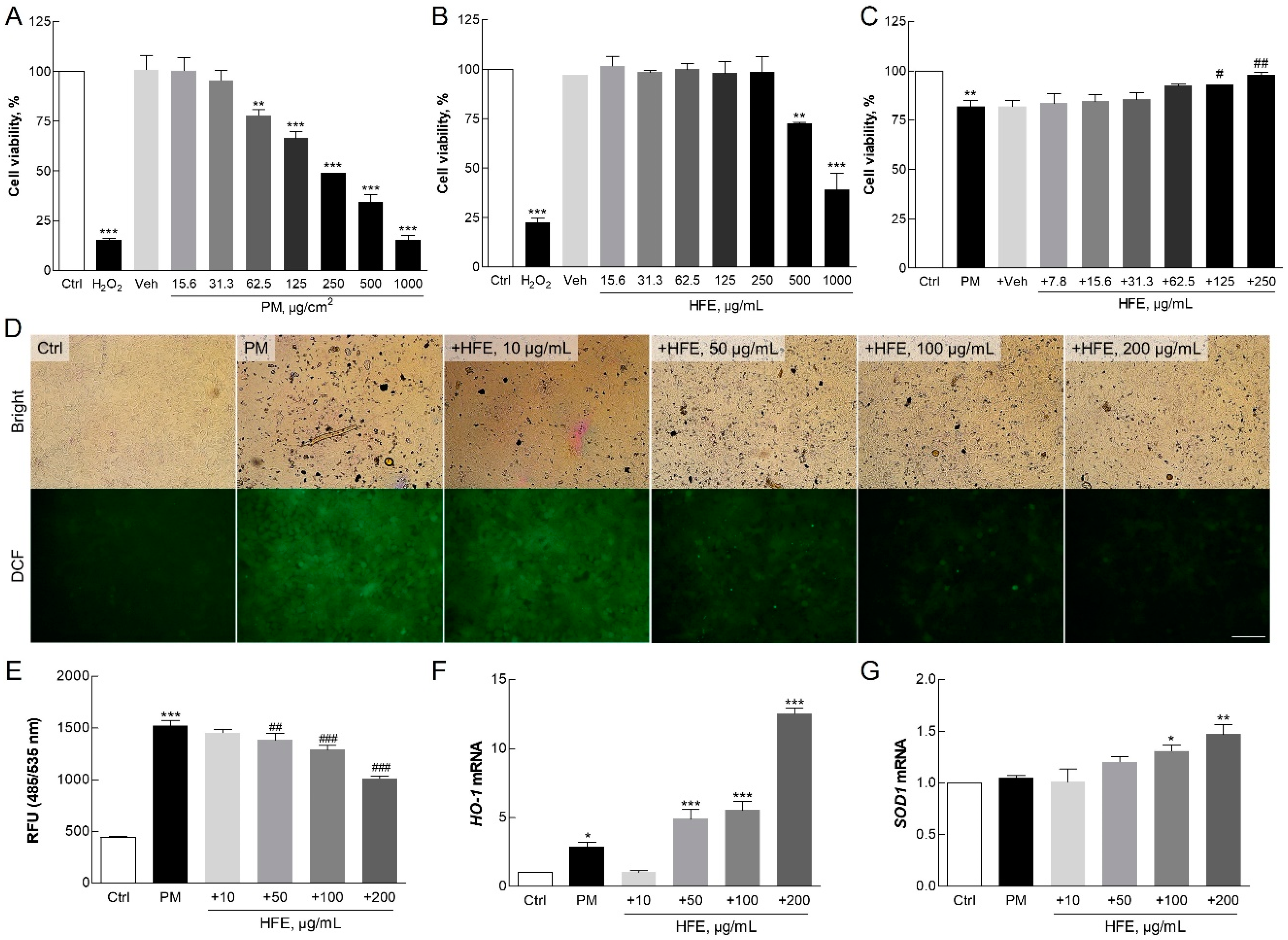
2.2. Effects of HFE on Cell Viability and Oxidative Stress Induced by PM
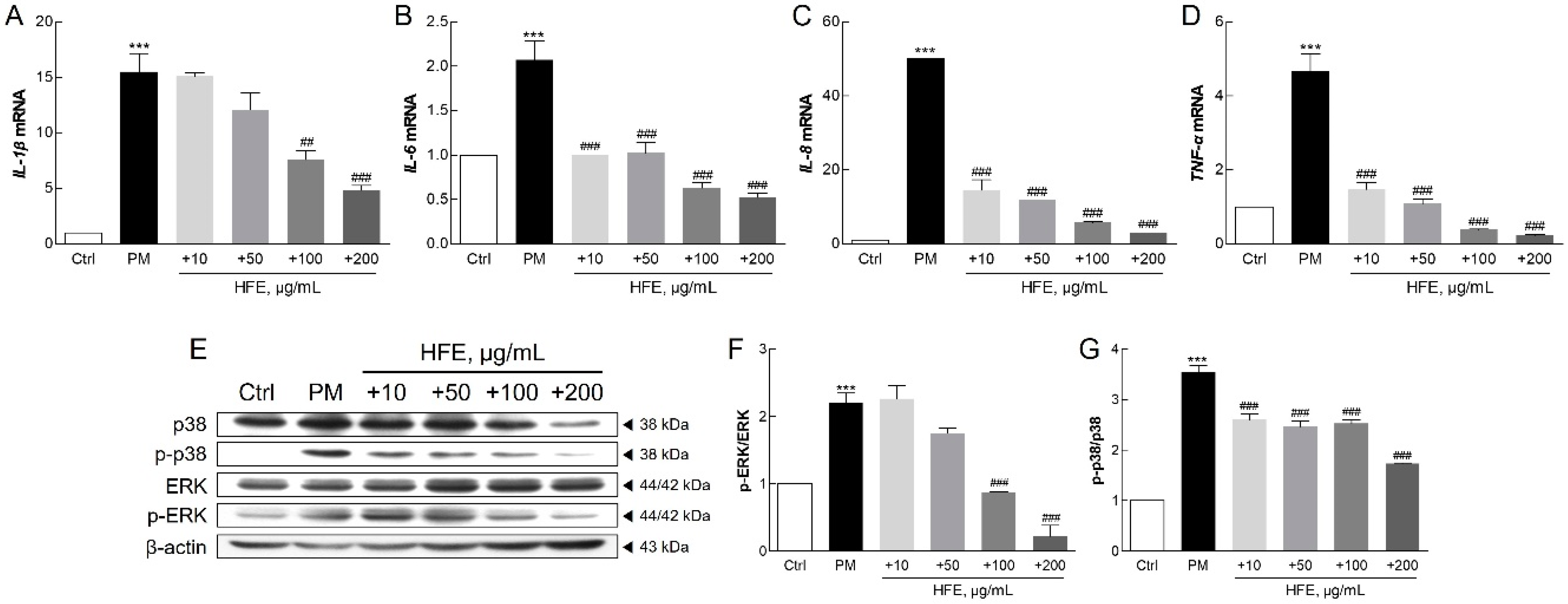
2.3. Effects of HFE on PM-Induced Inflammatory Response via the MAPK Signaling Pathway
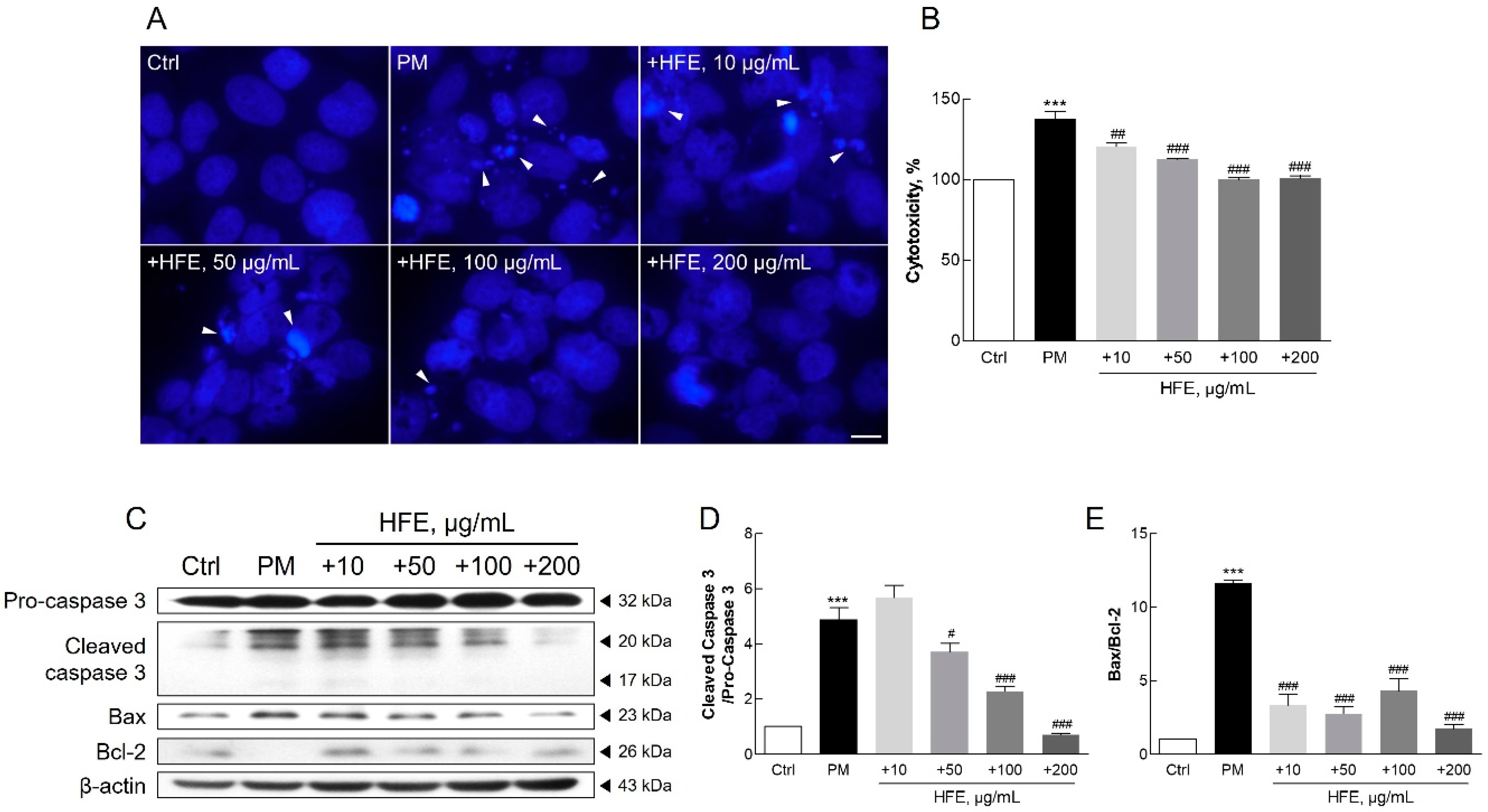
2.4. Protective Effects of HFE Against PM-Induced Apoptosis
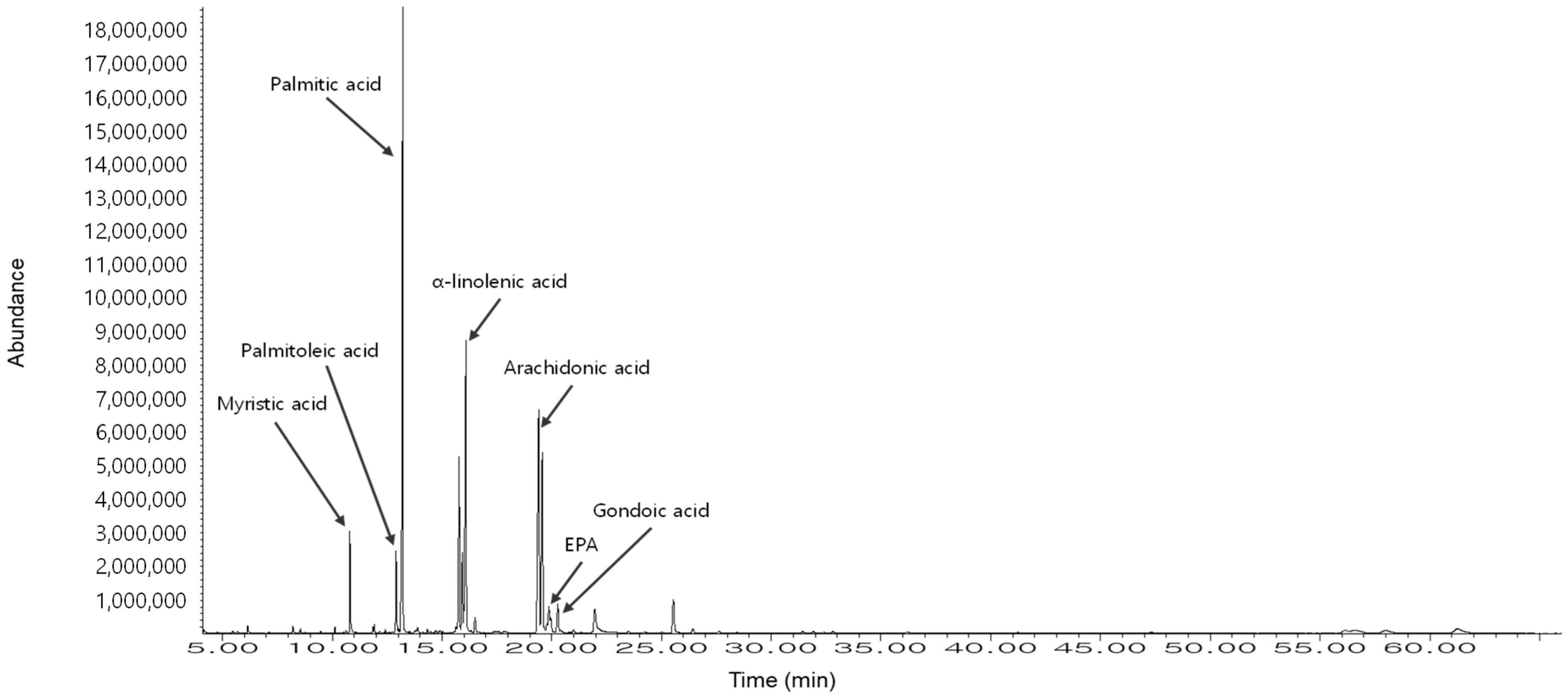
2.5. Fatty Acids and Fucoidan Content in HFE
3. Materials and Methods
3.1. Particulate Matter Preparation
3.2. Sample Preparation and Content Analysis
3.3. Antioxidant Assay
3.4. Cell Culture
3.5. Cell Viability Assay
3.6. Cytotoxicity Assay
3.7. Nuclear Staining
3.8. ROS Assay
3.9. qPCR Assay
3.10. Western Blot Analysis
3.11. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ANOVA | Analysis of variance |
| ARE | Antioxidant response element |
| BSA | Bovine serum albumin |
| DCFH2-DA | 2′,7′-Dichlorodihydrofluorescin diacetate |
| DMEM | Dulbecco’s modified Eagle’s medium |
| DMSO | Dimethyl sulfoxide |
| DPPH | 2,2-Diphenyl-1-picrylhydrazyl |
| ECL | Enhanced chemiluminescence |
| ERK | Extracellular signal-related kinase |
| FBS | Fetal bovine serum |
| HFE | Hizikia fusiformis ethanol extract |
| HO-1 | Heme oxygenase 1 |
| HRP | Horseradish peroxidase |
| IL | Interleukin |
| LDH | Lactate dehydrogenase |
| MAPK | Mitogen-activated protein kinase |
| MMP | Matrix metalloproteinase |
| NF-κB | Nuclear factor κ-light-chain-enhancer of activated B cells |
| NRF2 | Nuclear factor erythroid 2-related factor 2 |
| PBS | Phosphate-buffered saline |
| PM | Particulate matter |
| PVDF | Polyvinylidene difluoride |
| qPCR | Quantitative polymerase chain reaction |
| SD | Standard deviation |
| SDS-PAGE | Sodium dodecyl sulfate–polyacrylamide gel electrophoresis |
| SOD1 | Superoxide dismutase 1 |
| TLR | Toll-like receptor |
| TNF α | Tumor necrosis factor α |
References
- Fernando, I.P.S.; Jayawardena, T.U.; Kim, H.-S.; Lee, W.W.; Vaas, A.P.J.P.; De Silva, H.I.C.; Abayaweera, G.S.; Nanayakkara, C.M.; Abeytunga, D.T.U.; Lee, D.-S.; et al. Beijing urban particulate matter-induced injury and inflammation in human lung epithelial cells and the protective effects of fucosterol from Sargassum binderi (Sonder ex J. Agardh). Environ. Res. 2019, 172, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Fernando, P.D.; Piao, M.J.; Kang, K.A.; Zhen, A.X.; Herath, H.M.; Kang, H.K.; Choi, Y.H.; Hyun, J.W. Hesperidin Protects Human HaCaT Keratinocytes from Particulate Matter 2.5-Induced Apoptosis via the Inhibition of Oxidative Stress and Autophagy. Antioxidants 2022, 11, 1363. [Google Scholar] [CrossRef]
- Hammond, J.; Maher, B.A.; Gonet, T.; Bautista, F.; Allsop, D. Oxidative Stress, Cytotoxic and Inflammatory Effects of Urban Ultrafine Road-Deposited Dust from the UK and Mexico in Human Epithelial Lung (Calu-3) Cells. Antioxidants 2022, 11, 1814. [Google Scholar] [CrossRef]
- Li, Q.; Kang, Z.; Jiang, S.; Zhao, J.; Yan, S.; Xu, F.; Xu, J. Effects of Ambient Fine Particles PM2.5 on Human HaCaT Cells. Int. J. Environ. Res. Public. Health 2017, 14, 72. [Google Scholar] [CrossRef]
- Kim, K.-H.; Kabir, E.; Kabir, S. A review on the human health impact of airborne particulate matter. Environ. Int. 2015, 74, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Chao, L.; Feng, B.; Liang, H.; Zhao, X.; Song, J. Particulate matter and inflammatory skin diseases: From epidemiological and mechanistic studies. Sci. Total Environ. 2023, 905, 167111. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Li, Z.; Su, J. Air pollution and skin diseases: A comprehensive evaluation of the associated mechanism. Ecotoxicol. Environ. Saf. 2024, 278, 116429. [Google Scholar] [CrossRef] [PubMed]
- Diao, P.; He, H.; Tang, J.; Xiong, L.; Li, L. Natural compounds protect the skin from airborne particulate matter by attenuating oxidative stress. Biomed. Pharmacother. 2021, 138, 111534. [Google Scholar] [CrossRef]
- Wang, X.; Huang, C.; Yang, F.; Wang, K.; Cha, S.-H.; Mao, X.; Jeon, Y.-J.; Wang, L. Fucoidan isolated from the edible seaweed Sargassum fusiforme suppresses skin damage stimulated by airborne particulate matter. Algal Res. 2024, 77, 103339. [Google Scholar] [CrossRef]
- Piao, M.J.; Ahn, M.J.; Kang, K.A.; Ryu, Y.S.; Hyun, Y.J.; Shilnikova, K.; Zhen, A.X.; Jeong, J.W.; Choi, Y.H.; Kang, H.K.; et al. Particulate matter 2.5 damages skin cells by inducing oxidative stress, subcellular organelle dysfunction, and apoptosis. Arch. Toxicol. 2018, 92, 2077–2091. [Google Scholar] [CrossRef]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Orhan, I.E.; Banach, M.; Rollinger, J.M.; Barreca, D.; Weckwerth, W.; Bauer, R.; Bayer, E.A.; et al. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef] [PubMed]
- Hahn, D.; Kim, M.J.; Kwon, Y.; Kim, E.; Park, D.H.; Bae, J.-S. Natural products ameliorating the adverse health effects by air particulate matter. Biotechnol. Bioprocess. Eng. 2024, 29, 1–24. [Google Scholar] [CrossRef]
- Meinita, M.D.N.; Harwanto, D.; Sohn, J.H.; Kim, J.S.; Choi, J.S. Hizikia fusiformis: Pharmacological and Nutritional Properties. Foods 2021, 10, 1660. [Google Scholar] [CrossRef]
- Lee, K.H.; Kim, H.J.; Kim, H.B.; Kim, S.T.; Choi, Y.R.; Seo, D.W.; Yu, J.M.; Jang, S.K.; Kim, S.M.; Lee, D.-I.; et al. Hizikia fusiformis fractions successfully improve atopic dermatitis indices in anti-CD3-stimulated splenocytes and 2,4-dinitrochlorobenzene-treated BALB/c mice. J. Pharm. Pharmacol. 2014, 66, 466–476. [Google Scholar] [CrossRef] [PubMed]
- Siriwardhana, N.; Lee, K.W.; Jeon, Y.J.; Kim, S.H.; Haw, J.W. Antioxidant Activity of Hizikia fusiformis on Reactive Oxygen Species Scavenging and Lipid Peroxidation Inhibition. Food Sci. Technol. Int. 2003, 9, 339–346. [Google Scholar] [CrossRef]
- Han, Y.R.; Ali, M.Y.; Woo, M.-H.; Jung, H.A.; Choi, J.S. Anti-Diabetic and Anti-Inflammatory Potential of the Edible Brown Alga izikia Fusiformis. J. Food Biochem. 2015, 39, 417–428. [Google Scholar] [CrossRef]
- Dai, Y.-L.; Jiang, Y.-F.; Lee, H.G.; Jeon, Y.-J.; Kang, M.-C. Characterization and screening of anti-tumor activity of fucoidan from acid-processed hijiki (Hizikia fusiforme). Int. J. Biol. Macromol. 2019, 139, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Choi, M.H.; Jang, E.-s.; Shin, H.-J.; Lee, J.H. Polysaccharide from Hizikia Fusiformis Enhances the Immunomodulatory Activity of Macrophages. J. Rhinol. 2020, 27, 8–15. [Google Scholar] [CrossRef]
- Wang, L.; Lee, W.; Oh, J.Y.; Cui, Y.R.; Ryu, B.; Jeon, Y.J. Protective Effect of Sulfated Polysaccharides from Celluclast-Assisted Extract of Hizikia fusiforme Against Ultraviolet B-Induced Skin Damage by Regulating NF-κB, AP-1, and MAPKs Signaling Pathways In Vitro in Human Dermal Fibroblasts. Mar. Drugs 2018, 16, 239. [Google Scholar] [CrossRef]
- Kim, H.S.; Ahn, J.W.; Ha, N.R.; Damodar, K.; Jang, S.K.; Yoo, Y.M.; Gyoung, Y.S.; Joo, S.S. Antibacterial and Immunosuppressive Effects of a Novel Marine Brown Alga-Derived Ester in Atopic Dermatitis. Mar. Drugs 2024, 22, 354. [Google Scholar] [CrossRef]
- Husni, A.; Izmi, N.; Ayunani, F.Z.; Kartini, A.; Husnayain, N.; Isnansetyo, A. Characteristics and Antioxidant Activity of Fucoidan from Sargassum hystrix: Effect of Extraction Method. Int. J. Food Sci. 2022, 2022, 3689724. [Google Scholar] [CrossRef]
- Davis, T.A.; Volesky, B.; Mucci, A. A review of the biochemistry of heavy metal biosorption by brown algae. Water Res. 2003, 37, 4311–4330. [Google Scholar] [CrossRef] [PubMed]
- Nagahawatta, D.P.; Liyanage, N.M.; Jayawardhana, H.; Lee, H.G.; Jayawardena, T.U.; Jeon, Y.J. Anti-Fine Dust Effect of Fucoidan Extracted from Ecklonia maxima Laves in Macrophages via Inhibiting Inflammatory Signaling Pathways. Mar. Drugs 2022, 20, 413. [Google Scholar] [CrossRef] [PubMed]
- Liyanage, N.M.; Lee, H.G.; Nagahawatta, D.P.; Jayawardhana, H.; Song, K.M.; Choi, Y.S.; Jeon, Y.J.; Kang, M.C. Fucoidan from Sargassum autumnale Inhibits Potential Inflammatory Responses via NF-κB and MAPK Pathway Suppression in Lipopolysaccharide-Induced RAW 264.7 Macrophages. Mar. Drugs 2023, 21, 374. [Google Scholar] [CrossRef]
- Jeong, J.-W.; Hwang, S.J.; Han, M.H.; Lee, D.-S.; Yoo, J.S.; Choi, I.-W.; Cha, H.-J.; Kim, S.; Kim, H.-S.; Kim, G.-Y.; et al. Fucoidan inhibits lipopolysaccharide-induced inflammatory responses in RAW 264.7 macrophages and zebrafish larvae. Mol. Cell. Toxicol. 2017, 13, 405–417. [Google Scholar] [CrossRef]
- Flórez-Fernández, N.; Vaamonde-García, C.; Torres, M.D.; Buján, M.; Muíños, A.; Muiños, A.; Lamas-Vázquez, M.J.; Meijide-Faílde, R.; Blanco, F.J.; Domínguez, H. Relevance of the Extraction Stage on the Anti-Inflammatory Action of Fucoidans. Pharmaceutics 2023, 15, 808. [Google Scholar] [CrossRef]
- Khan, A.Q.; Agha, M.V.; Sheikhan, K.S.A.M.; Younis, S.M.; Tamimi, M.A.; Alam, M.; Ahmad, A.; Uddin, S.; Buddenkotte, J.; Steinhoff, M. Targeting deregulated oxidative stress in skin inflammatory diseases: An update on clinical importance. Biomed. Pharmacother. 2022, 154, 113601. [Google Scholar] [CrossRef]
- Forman, H.J.; Zhang, H. Targeting oxidative stress in disease: Promise and limitations of antioxidant therapy. Nat. Rev. Drug Discov. 2021, 20, 689–709. [Google Scholar] [CrossRef]
- Campbell, N.K.; Fitzgerald, H.K.; Dunne, A. Regulation of inflammation by the antioxidant haem oxygenase 1. Nat. Rev. Immunol. 2021, 21, 411–425. [Google Scholar] [CrossRef]
- Ma, Q. Role of nrf2 in oxidative stress and toxicity. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 401–426. [Google Scholar] [CrossRef]
- Ryu, Y.S.; Kang, K.A.; Piao, M.J.; Ahn, M.J.; Yi, J.M.; Hyun, Y.-M.; Kim, S.H.; Ko, M.K.; Park, C.O.; Hyun, J.W. Particulate matter induces inflammatory cytokine production via activation of NFκB by TLR5-NOX4-ROS signaling in human skin keratinocyte and mouse skin. Redox Biol. 2019, 21, 101080. [Google Scholar] [CrossRef] [PubMed]
- Seok, J.K.; Lee, J.W.; Kim, Y.M.; Boo, Y.C. Punicalagin and (-)-Epigallocatechin-3-Gallate Rescue Cell Viability and Attenuate Inflammatory Responses of Human Epidermal Keratinocytes Exposed to Airborne Particulate Matter PM10. Ski. Pharmacol. Physiol. 2018, 31, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-P.; Schunck, M.; Kallen, K.-J.; Neumann, C.; Trautwein, C.; Rose-John, S.; Proksch, E. The Interleukin-6 Cytokine System Regulates Epidermal Permeability Barrier Homeostasis. J. Investig. Dermatol. 2004, 123, 124–131. [Google Scholar] [CrossRef]
- Johnson, B.Z.; Stevenson, A.W.; Prêle, C.M.; Fear, M.W.; Wood, F.M. The Role of IL-6 in Skin Fibrosis and Cutaneous Wound Healing. Biomedicines 2020, 8, 101. [Google Scholar] [CrossRef]
- Cai, Y.; Xue, F.; Quan, C.; Qu, M.; Liu, N.; Zhang, Y.; Fleming, C.; Hu, X.; Zhang, H.-g.; Weichselbaum, R.; et al. A Critical Role of the IL-1β–IL-1R Signaling Pathway in Skin Inflammation and Psoriasis Pathogenesis. J. Investig. Dermatol. 2019, 139, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Dijkhoff, I.M.; Drasler, B.; Karakocak, B.B.; Petri-Fink, A.; Valacchi, G.; Eeman, M.; Rothen-Rutishauser, B. Impact of airborne particulate matter on skin: A systematic review from epidemiology to in vitro studies. Part. Fibre Toxicol. 2020, 17, 35. [Google Scholar] [CrossRef]
- Kaminska, B. MAPK signalling pathways as molecular targets for anti-inflammatory therapy—From molecular mechanisms to therapeutic benefits. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2005, 1754, 253–262. [Google Scholar] [CrossRef]
- Kim, M.; Kim, J.H.; Jeong, G.J.; Park, K.Y.; Lee, M.-K.; Seo, S.J. Particulate matter induces pro-inflammatory cytokines via phosphorylation of p38 MAPK possibly leading to dermal inflammaging. Exp. Dermatol. 2019, 28, 809–815. [Google Scholar] [CrossRef]
- Ziv, E.; Rotem, C.; Miodovnik, M.; Ravid, A.; Koren, R. Two modes of ERK activation by TNF in keratinocytes: Different cellular outcomes and bi-directional modulation by vitamin D. J. Cell. Biochem. 2008, 104, 606–619. [Google Scholar] [CrossRef]
- Aghaei-Zarch, S.M.; Nia, A.H.S.; Nouri, M.; Mousavinasab, F.; Najafi, S.; Bagheri-Mohammadi, S.; Aghaei-Zarch, F.; Toolabi, A.; Rasoulzadeh, H.; Ghanavi, J.; et al. The impact of particulate matters on apoptosis in various organs: Mechanistic and therapeutic perspectives. Biomed. Pharmacother. 2023, 165, 115054. [Google Scholar] [CrossRef]
- Zhen, A.X.; Piao, M.J.; Hyun, Y.J.; Kang, K.A.; Ryu, Y.S.; Cho, S.J.; Kang, H.K.; Koh, Y.S.; Ahn, M.J.; Kim, T.H.; et al. Purpurogallin Protects Keratinocytes from Damage and Apoptosis Induced by Ultraviolet B Radiation and Particulate Matter 2.5. Biomol. Ther. 2019, 27, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Paik, K.; Na, J.I.; Huh, C.H.; Shin, J.W. Particulate Matter and Its Molecular Effects on Skin: Implications for Various Skin Diseases. Int. J. Mol. Sci. 2024, 25, 9888. [Google Scholar] [CrossRef] [PubMed]
- Fernando, P.; Piao, M.J.; Zhen, A.X.; Ahn, M.J.; Yi, J.M.; Choi, Y.H.; Hyun, J.W. Extract of Cornus officinalis Protects Keratinocytes from Particulate Matter-induced Oxidative Stress. Int. J. Med. Sci. 2020, 17, 63–70. [Google Scholar] [CrossRef]
- Lee, J.; Kwon, J.; Jo, Y.J.; Yoon, S.B.; Hyeon, J.H.; Park, B.J.; You, H.J.; Youn, C.; Kim, Y.; Choi, H.W.; et al. Particulate matter 10 induces oxidative stress and apoptosis in rhesus macaques skin fibroblast. PeerJ 2023, 11, e16589. [Google Scholar] [CrossRef]
- Hahn, T.; Schulz, M.; Stadtmüller, R.; Zayed, A.; Muffler, K.; Lang, S.; Ulber, R. Cationic Dye for the Specific Determination of Sulfated Polysaccharides. Anal. Lett. 2016, 49, 1948–1962. [Google Scholar] [CrossRef]
- Fernando, I.P.S.; Dias, M.; Madusanka, D.M.D.; Han, E.J.; Kim, M.J.; Heo, S.J.; Lee, K.; Cheong, S.H.; Ahn, G. Low molecular weight fucoidan fraction ameliorates inflammation and deterioration of skin barrier in fine-dust stimulated keratinocytes. Int. J. Biol. Macromol. 2021, 168, 620–630. [Google Scholar] [CrossRef]
- Kirindage, K.; Jayasinghe, A.M.K.; Cho, N.; Cho, S.H.; Yoo, H.M.; Fernando, I.P.S.; Ahn, G. Fine-Dust-Induced Skin Inflammation: Low-Molecular-Weight Fucoidan Protects Keratinocytes and Underlying Fibroblasts in an Integrated Culture Model. Mar. Drugs 2022, 21, 12. [Google Scholar] [CrossRef]
- Zhao, M.; Garcia-Vaquero, M.; Przyborska, J.; Sivagnanam, S.P.; Tiwari, B. The development of analytical methods for the purity determination of fucoidan extracted from brown seaweed species. Int. J. Biol. Macromol. 2021, 173, 90–98. [Google Scholar] [CrossRef]
- Ahn, J.W.; Jang, S.K.; Jo, B.R.; Kim, H.S.; Park, J.Y.; Park, H.Y.; Yoo, Y.M.; Joo, S.S. A therapeutic intervention for Alzheimer’s disease using ginsenoside Rg3: Its role in M2 microglial activation and non-amyloidogenesis. J. Physiol. Pharmacol. 2021, 72, 185–193. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Composition | Common Name | Library Name | Relative Area% |
|---|---|---|---|---|
| 1 | 14:0 | Myristic acid | Tetradecanoic acid, methyl ester | 4.42 |
| 2 | 16:1 | Palmitoleic acid | 9-Hexadecenoic acid, methyl ester | 4.11 |
| 3 | 16:0 | Palmitic acid | Hexadecanoic acid, methyl ester | 31.19 |
| 4 | 18:3 | α-Linolenic acid | 9,12,15-Octadecatrienoic acid, methyl ester | 26.68 |
| 5 | 20:4 | Arachidonic acid | 5,8,11,14-Eicasatetraenoic acid, methyl ester | 18.03 |
| 6 | 20:5 | EPA | 5,8,11,14,17-Eicosapentaenoic acid, methyl ester | 12.94 |
| 7 | 20:1 | Gondoic acid | 11-Eicosenoic acid, methyl ester | 2.64 |
| Species | Gene | Primer Sequences | Accession No. | |
|---|---|---|---|---|
| Human | HO-1 | Fwd | 5′-CTT CAC CTT CCC CAA CAT TG | NM_002133.3 |
| Rvs | 5′-CCT CAA AGA GCT GGA TGT TG | |||
| SOD1 | Fwd | 5′-GCA TCA TCA ATT TCG AGC AGA | NM_000454.5 | |
| Rvs | 5′-CAA TAG ACA CAT CGG CCA CAC | |||
| IL-1β | Fwd | 5′-GTA CCT GAG CTC GCC AGT GA | NM_000576.3 | |
| Rvs | 5′-TGA AGC CCT TGC TGT AGT GG | |||
| IL-6 | Fwd | 5′-CCT AGA GTA CCT CCA GAA CA | NM_001318095.2 | |
| Rvs | 5′-AGA TGA GTT GTC ATG TCC TG | |||
| IL-8 | Fwd | 5′-CAA ACC TTT CCA CCC CAA AT | NM_000584 | |
| Rvs | 5′-ACA ACC CTC TGC ACC CAG TT | |||
| TNF-α | Fwd | 5′-TTG TTC CTC AGC CTC TTC TC | NM_000594.4 | |
| Rvs | 5′-AAG ATG ATC TGA CTG CCT GG | |||
| β-Actin | Fwd | 5′-ACC TGA CTG ACT ACC TCA TG | NM_001101.5 | |
| Rvs | 5′-CTC ATT GCC AAT GGT GAT GA | |||
| Epitope | Manufacturer | Cat. No. | Dilution | Host |
|---|---|---|---|---|
| β-Actin (43 kDa) | Santa Cruz Biotechnology | sc-81178 | 1:1000 | Mouse |
| Bax (23 kDa) | sc-70407 | 1:1000 | Mouse | |
| Bcl-2 (26 kDa) | sc-7382 | 1:1000 | Mouse | |
| Caspase-3 (32/20/17 kDa) | sc-56053 | 1:1000 | Mouse | |
| p-p38 (38 kDa) | sc-7973 | 1:1000 | Mouse | |
| p38 (38 kDa) | sc-535 | 1:1000 | Rabbit | |
| p-ERK1/2 (44/42 kDa) | ABclonal | AP0974 | 1:1000 | Rabbit |
| EKR1/2 (44/42 kDa) | A16686 | 1:1000 | Rabbit | |
| Mouse IgG (HRP-linked) | Cell Signaling Technology | #7076 | 1:5000 | Goat |
| Rabbit IgG (HRP-linked) | #7074 | 1:10000 | Horse |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahn, J.W.; Kim, H.S.; Kim, S.H.; Yang, H.S.; Damodar, K.; Yoo, Y.-M.; Hong, J.T.; Joo, S.S. Amelioration of Particulate Matter-Induced Oxidative Stress by a Bioactive Hizikia fusiformis Extract: A Functional Biomaterial for Cosmeceutical Applications. Mar. Drugs 2025, 23, 135. https://doi.org/10.3390/md23030135
Ahn JW, Kim HS, Kim SH, Yang HS, Damodar K, Yoo Y-M, Hong JT, Joo SS. Amelioration of Particulate Matter-Induced Oxidative Stress by a Bioactive Hizikia fusiformis Extract: A Functional Biomaterial for Cosmeceutical Applications. Marine Drugs. 2025; 23(3):135. https://doi.org/10.3390/md23030135
Chicago/Turabian StyleAhn, Jeong Won, Hyun Soo Kim, So Hui Kim, Hye Soo Yang, Kongara Damodar, Yeong-Min Yoo, Jin Tae Hong, and Seong Soo Joo. 2025. "Amelioration of Particulate Matter-Induced Oxidative Stress by a Bioactive Hizikia fusiformis Extract: A Functional Biomaterial for Cosmeceutical Applications" Marine Drugs 23, no. 3: 135. https://doi.org/10.3390/md23030135
APA StyleAhn, J. W., Kim, H. S., Kim, S. H., Yang, H. S., Damodar, K., Yoo, Y.-M., Hong, J. T., & Joo, S. S. (2025). Amelioration of Particulate Matter-Induced Oxidative Stress by a Bioactive Hizikia fusiformis Extract: A Functional Biomaterial for Cosmeceutical Applications. Marine Drugs, 23(3), 135. https://doi.org/10.3390/md23030135