

Polysaccharide Fraction Isolated from Saccharina japonica Exhibits Anti-Cancer Effects Through Immunostimulating Activities


 ,
,
Abstract
1. Introduction
2. Results and Discussion
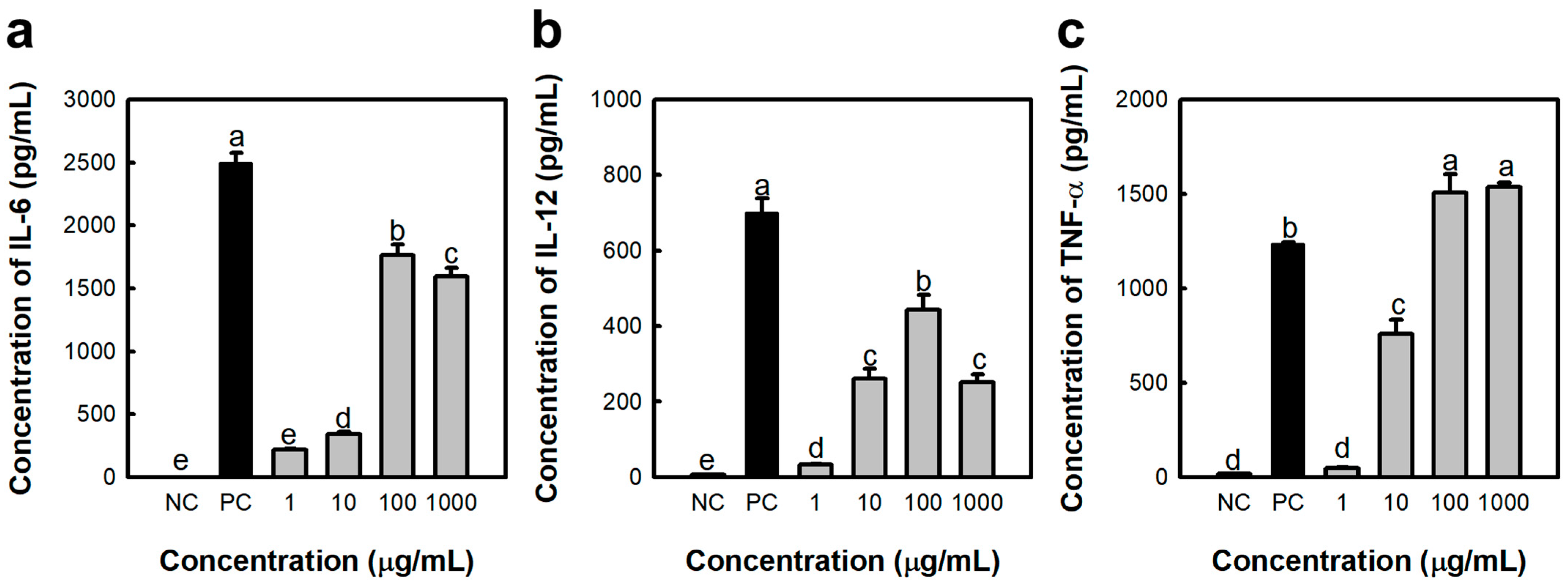
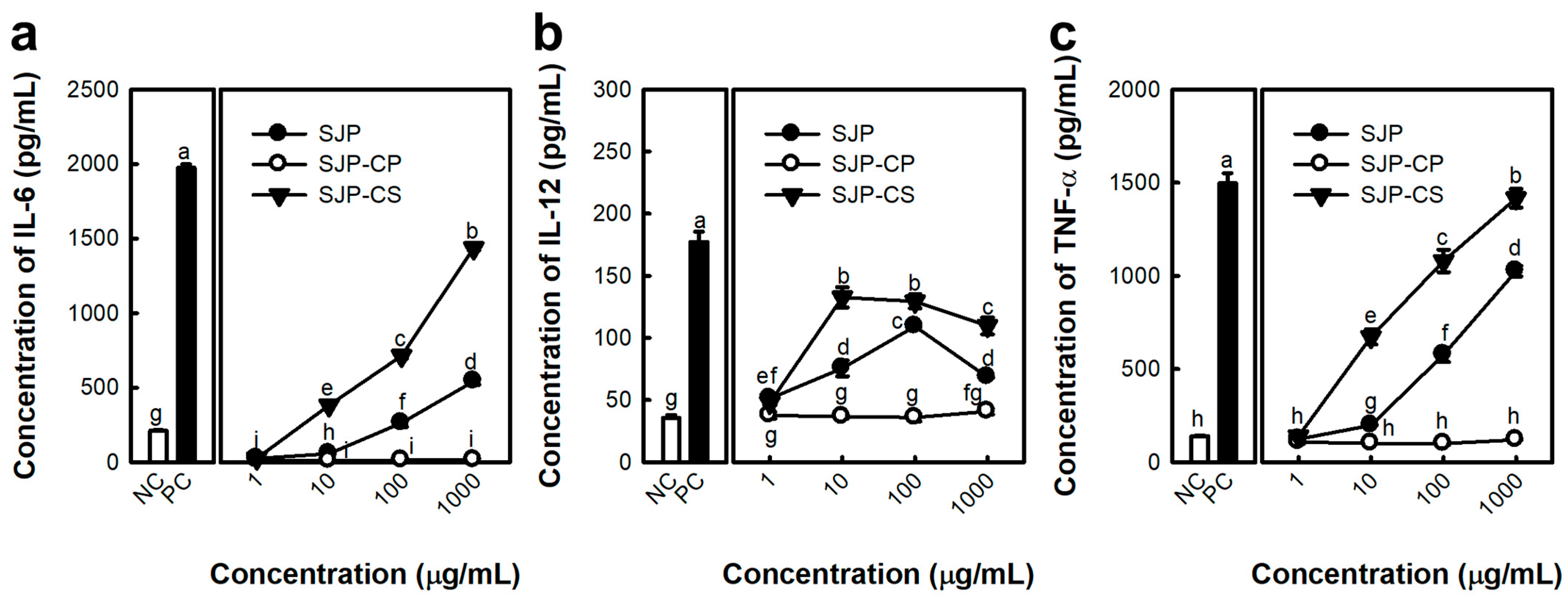
2.1. Activated Macrophage by Polysaccharide Fraction Isolated from S. japonica
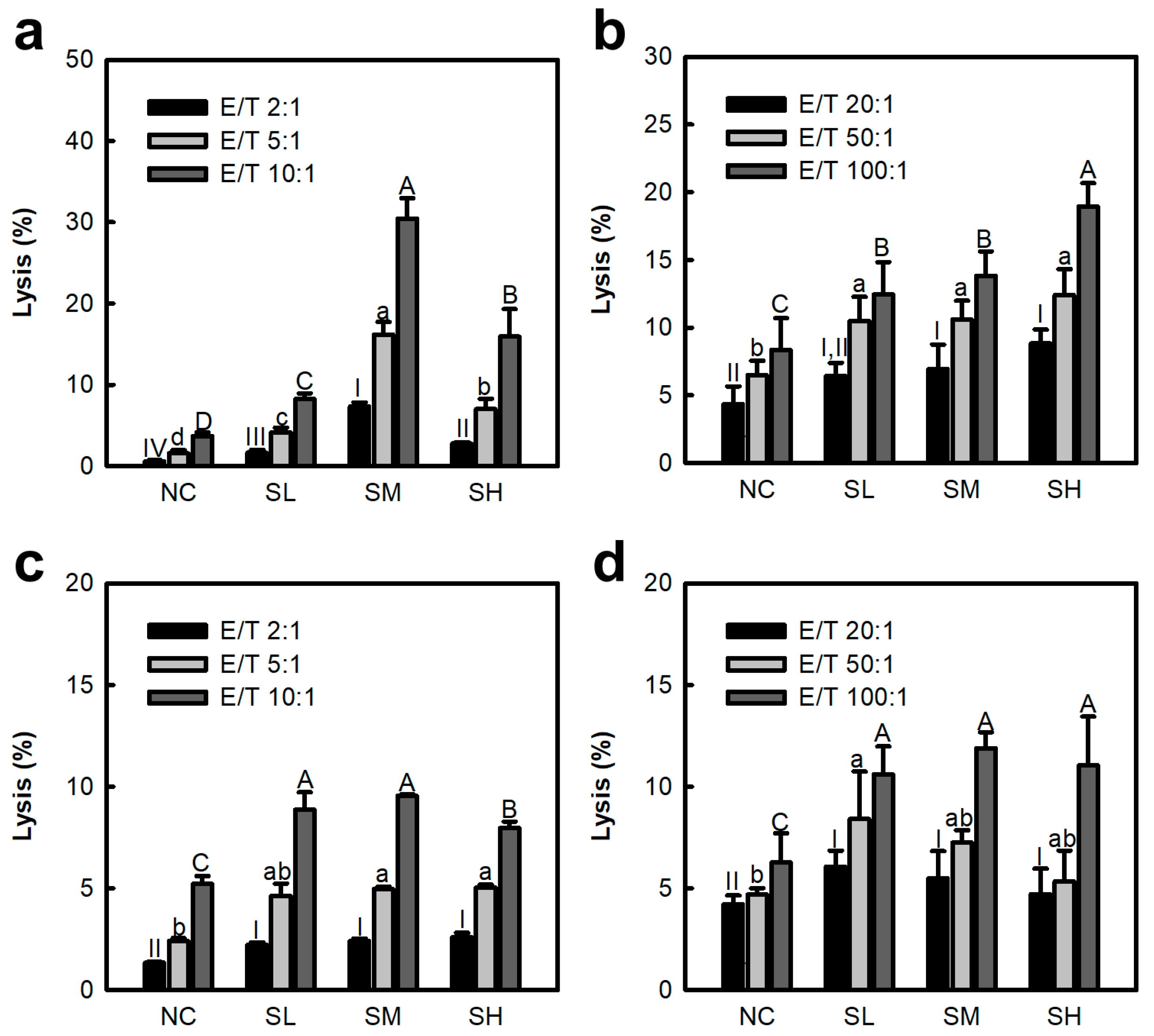
2.2. Effects of Polysaccharide from S. japonica for NK Cells and CTLs
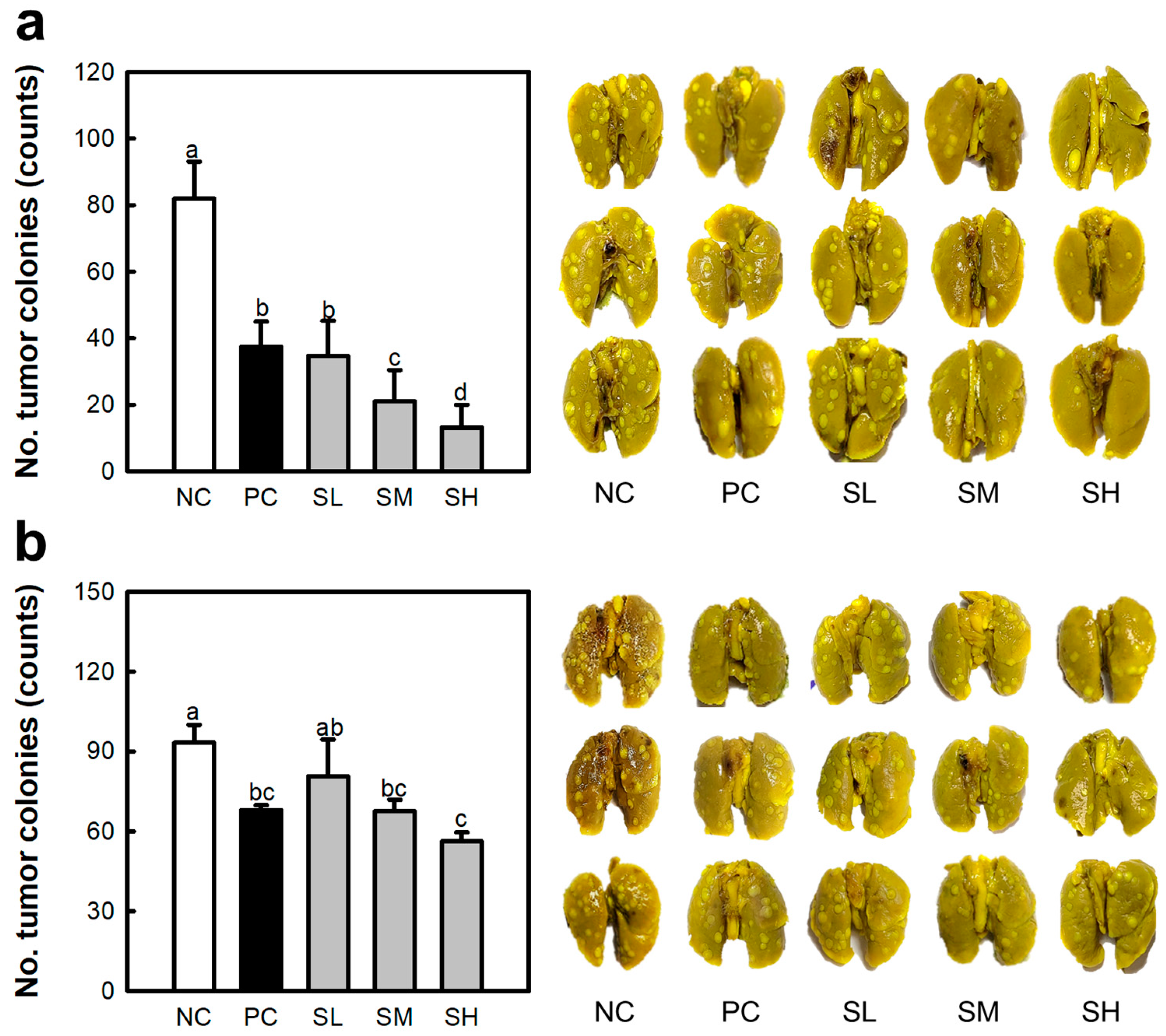
2.3. Inhibitory Efficacy of SJP Against Lung Cancer
2.4. Anti-Cancer Effects of SJP in NK Cell Function Depleted BALB/c Mice
2.5. Determination of Active Region of S. japonica
3. Materials and Methods
3.1. Materials and Animals
3.2. Cytokine Release from Peritoneal Macrophages of Mice
3.3. NK-Cell-Related Cytolytic Effect
3.4. Cytotoxic-T-Lymphocyte-Mediated Cytolytic Effect on Colon26-M3.1
3.5. Lung-Cancer-Bearing Mice Model
3.6. NK-Cell-Activity-Impaired Mice Model
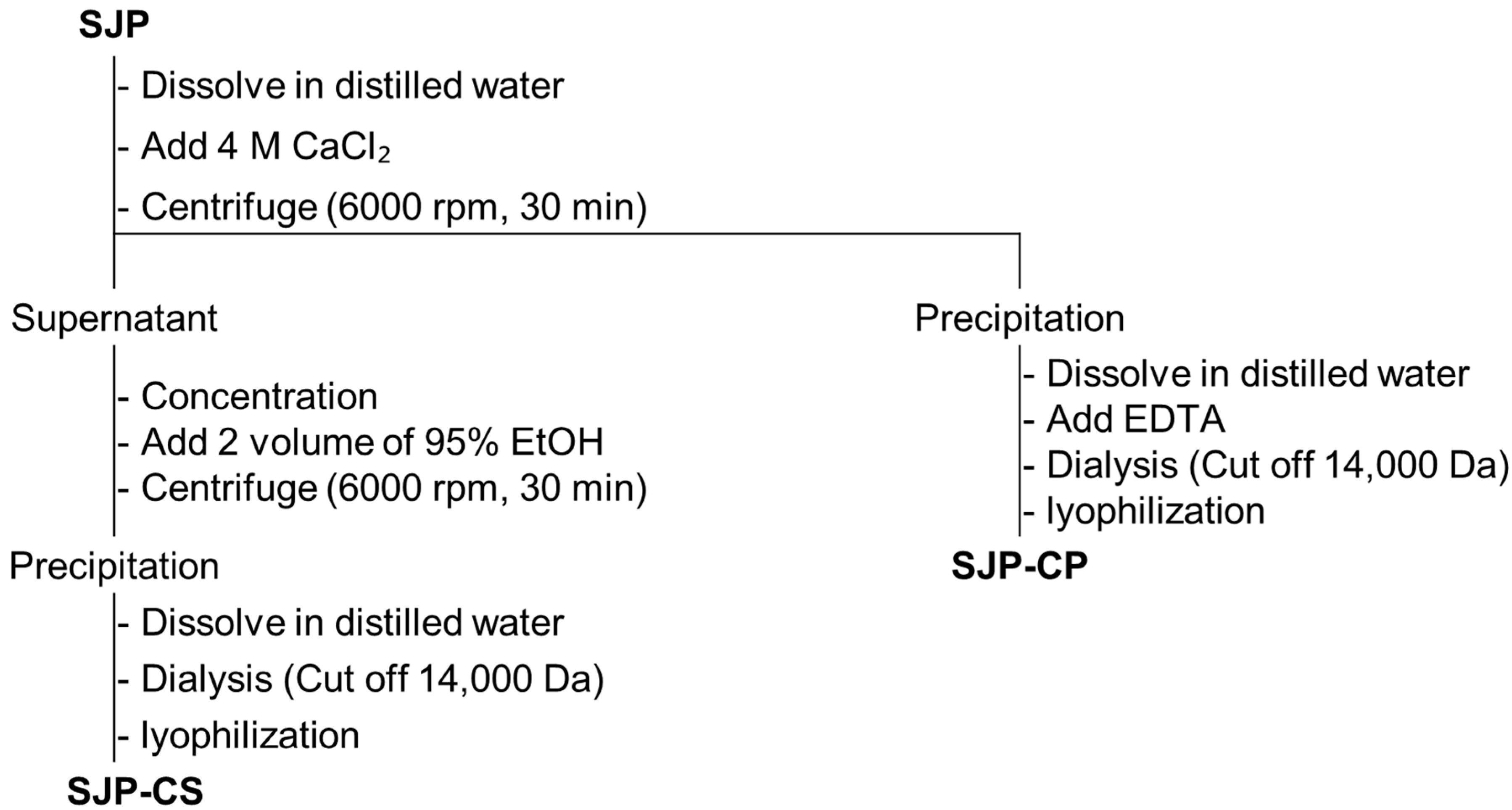
3.7. Fractionation of Polysaccharide
3.8. Component Sugar
3.9. Molecular Weight
3.10. Statistical Test
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, J.; He, Z.; Liang, Y.; Peng, T.; Hu, Z. Insights into algal polysaccharides: A review of their structure, depolymerases, and metabolic pathways. J. Agric. Food Chem. 2022, 70, 1749–1765. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.N.; Ishita, I.J.; Jin, S.E.; Choi, R.J.; Lee, C.M.; Kim, Y.S.; Jung, H.A.; Choi, J.S. Anti-inflammatory activity of edible brown alga Saccharina japonica and its constituents pheophorbide a and pheophytin a in LPS-stimulated RAW 264.7 macrophage cells. Food Chem. Toxicol. 2013, 55, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zheng, Y.; Zhang, Y.; Yang, Y.; Wang, P.; Imre, B.; Wong, A.C.; Hsieh, Y.S.; Wang, D. Brown algae carbohydrates: Structures, pharmaceutical properties, and research challenges. Mar. Drugs. 2021, 19, 620. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Lu, F.; Wei, X.; Zhao, R. Fucoidan: Structure and bioactivity. Molecules 2008, 13, 1671–1695. [Google Scholar] [CrossRef] [PubMed]
- Luthuli, S.; Wu, S.; Cheng, Y.; Zheng, X.; Wu, M.; Tong, H. Therapeutic effects of fucoidan: A review on recent studies. Mar. Drugs 2019, 17, 487. [Google Scholar] [CrossRef]
- Chen, R.; Xu, J.; Wu, W.; Wen, Y.; Lu, S.; El-Seedi, H.R.; Zhao, C. Structure–immunomodulatory activity relationships of dietary polysaccharides. Curr. Res. Food Sci. 2022, 5, 1330–1341. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, Y.; Li, W.; Zhang, B.; Yin, J.; Liuqi, S.; Wang, J.; Peng, B.; Wang, S. Fucoidan ameliorated dextran sulfate sodium-induced ulcerative colitis by modulating gut microbiota and bile acid metabolism. J. Agric. Food Chem. 2022, 70, 14864–14876. [Google Scholar] [CrossRef]
- Son, S.-U.; Suh, H.J.; Shin, K.-S. Characterization of a novel sulfated-rhamnoglucuronan isolated from Korean seaweed Ulva pertusa and its efficacy for treatment of inflammatory bowel disease in mice. Carbohydr. Polym. 2024, 342, 122373. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Yan, C.; Fu, Y.; Ai, C.; Bi, J.; Lin, W.; Song, S. Orally administrated fucoidan and its low-molecular-weight derivatives are absorbed differentially to alleviate coagulation and thrombosis. Int. J. Biol. Macromol. 2024, 255, 128092. [Google Scholar] [CrossRef] [PubMed]
- Oliyaei, N.; Moosavi-Nasab, M.; Mazloomi, S.M. Therapeutic activity of fucoidan and carrageenan as marine algal polysaccharides against viruses. 3 Biotech 2022, 12, 154. [Google Scholar] [CrossRef] [PubMed]
- Yao, W.; Qiu, H.-M.; Cheong, K.-L.; Zhong, S. Advances in anti-cancer effects and underlying mechanisms of marine algae polysaccharides. Int. J. Biol. Macromol. 2022, 221, 472–485. [Google Scholar] [CrossRef] [PubMed]
- Su, F.; Song, Q.; Zhang, C.; Xu, X.; Li, M.; Yao, D.; Wu, L.; Qu, X.; Guan, H.; Yu, G. A β-1, 3/1, 6-glucan from Durvillaea Antarctica inhibits tumor progression in vivo as an immune stimulator. Carbohydr. Polym. 2019, 222, 114993. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Nie, W.; Yu, G.; Li, Y.; Hu, Y.; Lu, J.; Jin, L. Antitumor and immunomodulatory activity of polysaccharides from Sargassum fusiforme. Food Chem. Toxicol. 2012, 50, 695–700. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Xiang, Y.; Sheng, R.; Tomás, H.; Rodrigues, J.; Gu, Z.; Zhang, H.; Gong, Q.; Luo, K. Polysaccharide-based nanomedicines for cancer immunotherapy: A review. Bioact. Mater. 2021, 6, 3358–3382. [Google Scholar] [CrossRef] [PubMed]
- Son, S.-U.; Kim, H.W.; Park, M.S.; Shin, K.-S. Effects of intravenous administration of polysaccharide purified from fermented barley on tumor metastasis inhibition via immunostimulating activities. Food Biosci. 2022, 49, 101833. [Google Scholar] [CrossRef]
- Chen, S.; Saeed, A.F.; Liu, Q.; Jiang, Q.; Xu, H.; Xiao, G.G.; Rao, L.; Duo, Y. Macrophages in immunoregulation and therapeutics. Signal Transduct. Target Ther. 2023, 8, 207. [Google Scholar] [CrossRef] [PubMed]
- Shimasaki, N.; Jain, A.; Campana, D. NK cells for cancer immunotherapy. Nat. Rev. Drug Discov. 2020, 19, 200–218. [Google Scholar] [CrossRef] [PubMed]
- Fielding, C.A.; McLoughlin, R.M.; McLeod, L.; Colmont, C.S.; Najdovska, M.; Grail, D.; Ernst, M.; Jones, S.A.; Topley, N.; Jenkins, B.J. IL-6 regulates neutrophil trafficking during acute inflammation via STAT3. J. Immun. 2008, 181, 2189–2195. [Google Scholar] [CrossRef]
- Shin, M.H.; Kim, J.; Lim, S.A.; Kim, J.; Kim, S.-J.; Lee, K.-M. NK cell-based immunotherapies in cancer. Immune Netw. 2020, 20, e14. [Google Scholar] [CrossRef]
- Lu, H.; Yang, Y.; Gad, E.; Wenner, C.A.; Chang, A.; Larson, E.R.; Dang, Y.; Martzen, M.; Standish, L.J.; Disis, M.L. Polysaccharide krestin is a novel TLR2 agonist that mediates inhibition of tumor growth via stimulation of CD8 T cells and NK cells. Clin. Cancer Res. 2011, 17, 67–76. [Google Scholar] [CrossRef]
- Choi, E.H.; Son, S.U.; Shin, K.S. Tumor inhibitory effect via immunostimulating activities of a rhamnogalacturonan-I-rich polysaccharide isolated from turmeric (Curcuma longa L.). J. Food Biochem. 2022, 46, e14362. [Google Scholar] [CrossRef]
- Xie, J.-H.; Jin, M.-L.; Morris, G.A.; Zha, X.-Q.; Chen, H.-Q.; Yi, Y.; Li, J.-E.; Wang, Z.-J.; Gao, J.; Nie, S.-P. Advances on bioactive polysaccharides from medicinal plants. Crit. Rev. Food Sci. Nutr. 2016, 56, S60–S84. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.M.; Andres, M.L.; Gridley, D.S. NK cell depletion results in accelerated tumor growth and attenuates the antitumor effect of total body irradiation. Int. J. Oncol. 2003, 23, 1585–1592. [Google Scholar] [CrossRef]
- Nishikado, H.; Mukai, K.; Kawano, Y.; Minegishi, Y.; Karasuyama, H. NK cell-depleting anti-asialo GM1 antibody exhibits a lethal off-target effect on basophils in vivo. J. Immun. 2011, 186, 5766–5771. [Google Scholar] [CrossRef]
- Smyth, M.J.; Crowe, N.Y.; Godfrey, D.I. NK cells and NKT cells collaborate in host protection from methylcholanthrene-induced fibrosarcoma. Int. Immunol. 2001, 13, 459–463. [Google Scholar] [CrossRef] [PubMed]
- Sideras, K.; Braat, H.; Kwekkeboom, J.; Van Eijck, C.; Peppelenbosch, M.; Sleijfer, S.; Bruno, M. Role of the immune system in pancreatic cancer progression and immune modulating treatment strategies. Cancer Treat. Rev. 2014, 40, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Dougan, M.; Dranoff, G. Immune therapy for cancer. Annu. Rev. Immunol. 2009, 27, 83–117. [Google Scholar] [CrossRef]
- Usov, A.I.; Bilan, M.I. Fucoidans—Sulfated polysaccharides of brown algae. Russ. Chem. Rev. 2009, 78, 785. [Google Scholar] [CrossRef]
- Salki, I.; Saito, S.; Fujita, C.; Ishida, H.; Iida, J.; Murata, J.; Hasegawa, A.; Azuma, I. Induction of tumoricidal macrophages and production of cytokines by synthetic muramyl dipeptide analogues. Vaccine 1988, 6, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Son, S.-U.; Kim, T.E.; Park, J.-H.; Suh, H.J.; Shin, K.-S. Immunostimulating effects of ulvan type polysaccharide isolated from Korean Ulva pertusa in cyclophosphamide-induced immunosuppressed BALB/c mice. Int. J. Biol. Macromol. 2024, 275, 133518. [Google Scholar] [CrossRef] [PubMed]
- Tsukagoshi, S.; Hashimoto, Y.; Fujii, G.; Kobayashi, H.; Nomoto, K.; Orita, K. Krestin (Psk). Cancer Treat. Rev. 1984, 11, 131–155. [Google Scholar] [CrossRef]
- Shin, H.Y.; Kim, Y.S.; Shin, K.-S.; Han, S.H.; Suh, H.J.; Yu, K.-W. Effect of rhamnogalacturonan-I-rich polysaccharides isolated from crabapple hydrolysates on IL-1β-induced inflammation in intestinal epithelial cells. Int. J. Biol. Macromol. 2024, 277, 134240. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component Sugar (Mole %) | SJP | SJP-CS * | SJP-CP ** |
|---|---|---|---|
| Rhamnose | - | 1.0 ± 0.0 | - |
| Fucose | 9.4 ± 0.3 | 51.7 ± 0.2 | 3.4 ± 0.5 |
| Arabinose | - | - | - |
| Xylose | 0.7 ± 0.0 | 2.1 ± 0.1 | - |
| Mannose | 2.2 ± 0.1 | 8.9 ± 0.1 | 1.7 ± 0.2 |
| Galactose | 4.8 ± 0.1 | 18.8 ± 0.0 | - |
| Glucose | - | - | - |
| Glucuronic acid | 4.4 ± 0.2 | 10.0 ± 0.3 | - |
| Galacturonic acid | - | - | - |
| Guluronic acid | 18.8 ± 1.6 | 1.5 ± 0.2 | 21.9 ± 0.9 |
| Mannuronic acid | 59.7 ± 2.0 | 5.9 ± 0.2 | 73.0 ± 1.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, M.S.; Son, S.-U.; Kim, T.E.; Shim, S.H.; Jang, B.-K.; Park, S.; Shin, K.-S. Polysaccharide Fraction Isolated from Saccharina japonica Exhibits Anti-Cancer Effects Through Immunostimulating Activities. Mar. Drugs 2025, 23, 38. https://doi.org/10.3390/md23010038
Park MS, Son S-U, Kim TE, Shim SH, Jang B-K, Park S, Shin K-S. Polysaccharide Fraction Isolated from Saccharina japonica Exhibits Anti-Cancer Effects Through Immunostimulating Activities. Marine Drugs. 2025; 23(1):38. https://doi.org/10.3390/md23010038
Chicago/Turabian StylePark, Min Seung, Seung-U Son, Tae Eun Kim, Se Hyun Shim, Bong-Keun Jang, Sunyoung Park, and Kwang-Soon Shin. 2025. "Polysaccharide Fraction Isolated from Saccharina japonica Exhibits Anti-Cancer Effects Through Immunostimulating Activities" Marine Drugs 23, no. 1: 38. https://doi.org/10.3390/md23010038
APA StylePark, M. S., Son, S.-U., Kim, T. E., Shim, S. H., Jang, B.-K., Park, S., & Shin, K.-S. (2025). Polysaccharide Fraction Isolated from Saccharina japonica Exhibits Anti-Cancer Effects Through Immunostimulating Activities. Marine Drugs, 23(1), 38. https://doi.org/10.3390/md23010038