


Stress-Induced Production of Bioactive Oxylipins in Marine Microalgae

 ,
,  , , and
, , and
Abstract

1. Introduction
2. Results
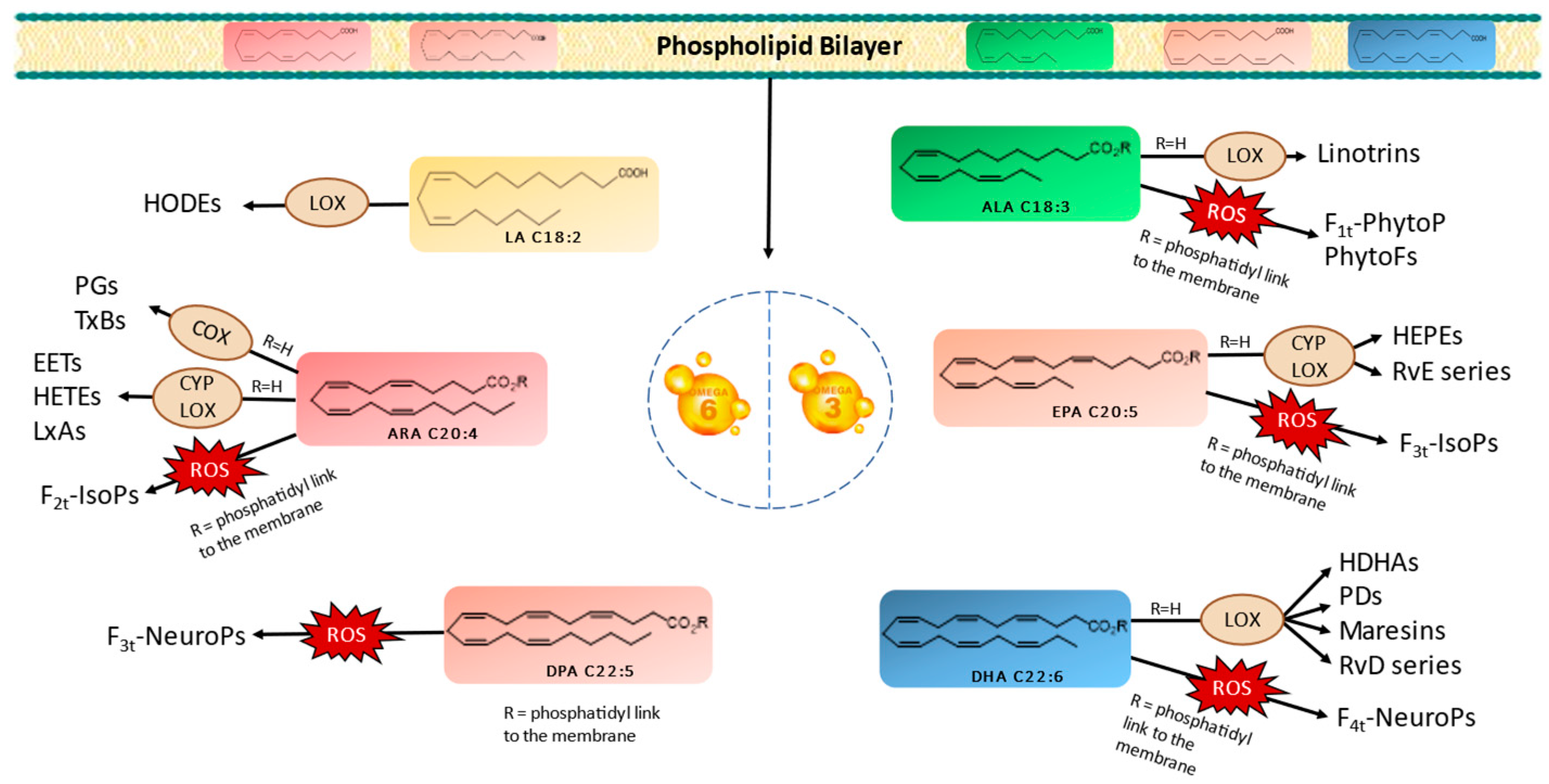
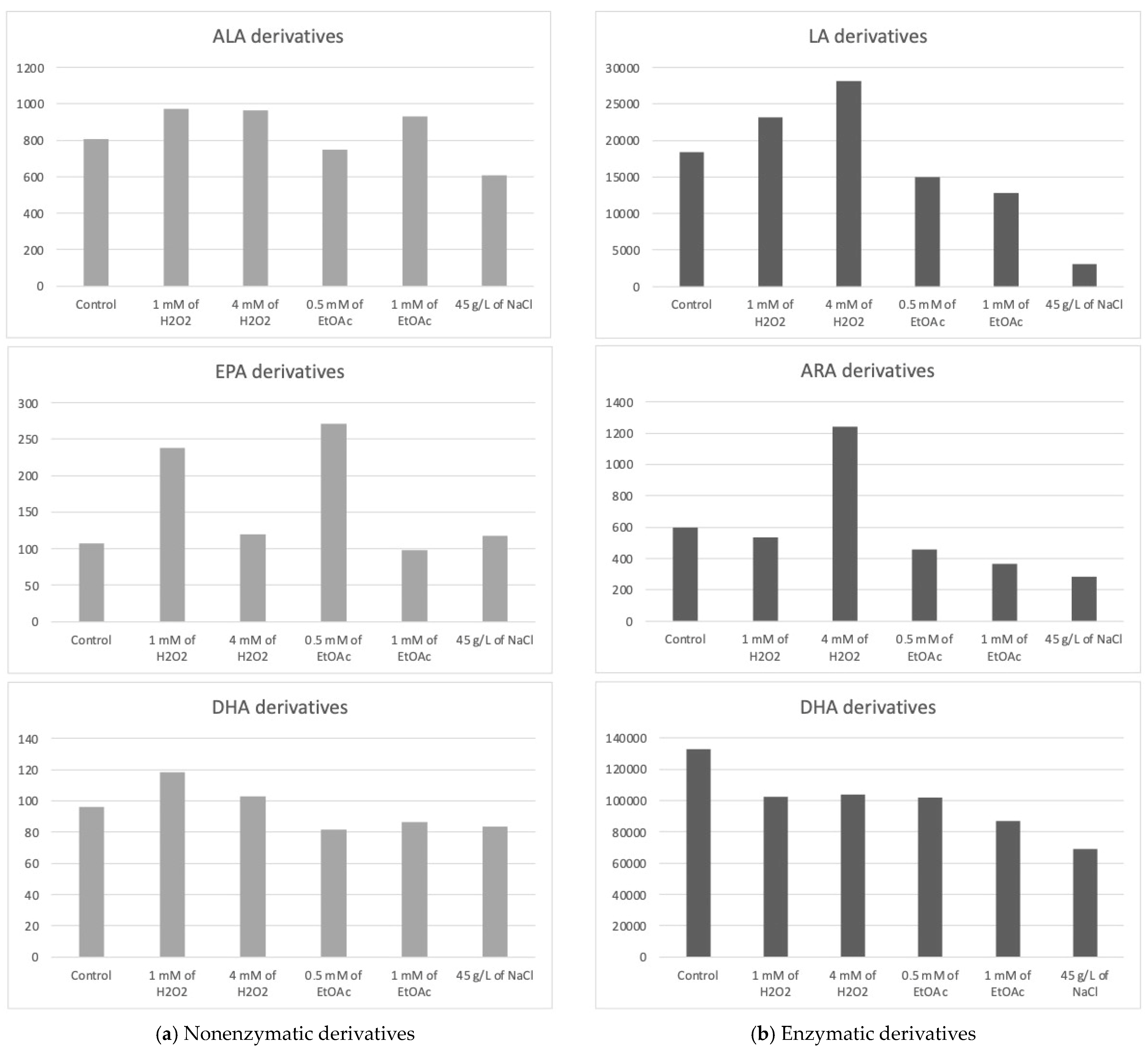
2.1. Profiles of Enzymatic and Nonenzymatic Oxygenated Metabolites in Stressed Mi124
2.1.1. Profiles of Nonenzymatic Oxygenated Metabolites
Derivatives from ARA (C20:4n−6) and DPA (C22:5n−6)
Derivatives from ALA (C18:3n−3)
Derivatives from EPA (C20:5n−3)
Derivatives from DHA (C22:6n−3)
2.1.2. Profiles of Enzymatic Oxygenated Metabolites
Derivatives of LA (C18:2n−6)
Derivatives of ARA (C20:4n−6)
Derivatives from ALA, EPA, and DHA
- (i)
- The addition of H2O2 to the culture medium seemed to induce a slight increase in the concentration of the 9(R),16(RS)-linotrin while EtOAc or NaCl would have the opposite effect;
- (ii)
- The addition of H2O2 (4 mM) would also slightly increase the concentration of 18-HEPE, a derivative of EPA;
- (iii)
- None of them appear to be produced in greater quantities when abiotic stresses are applied, since the highest concentrations for each of the metabolites appeared to be obtained with the control culture medium.
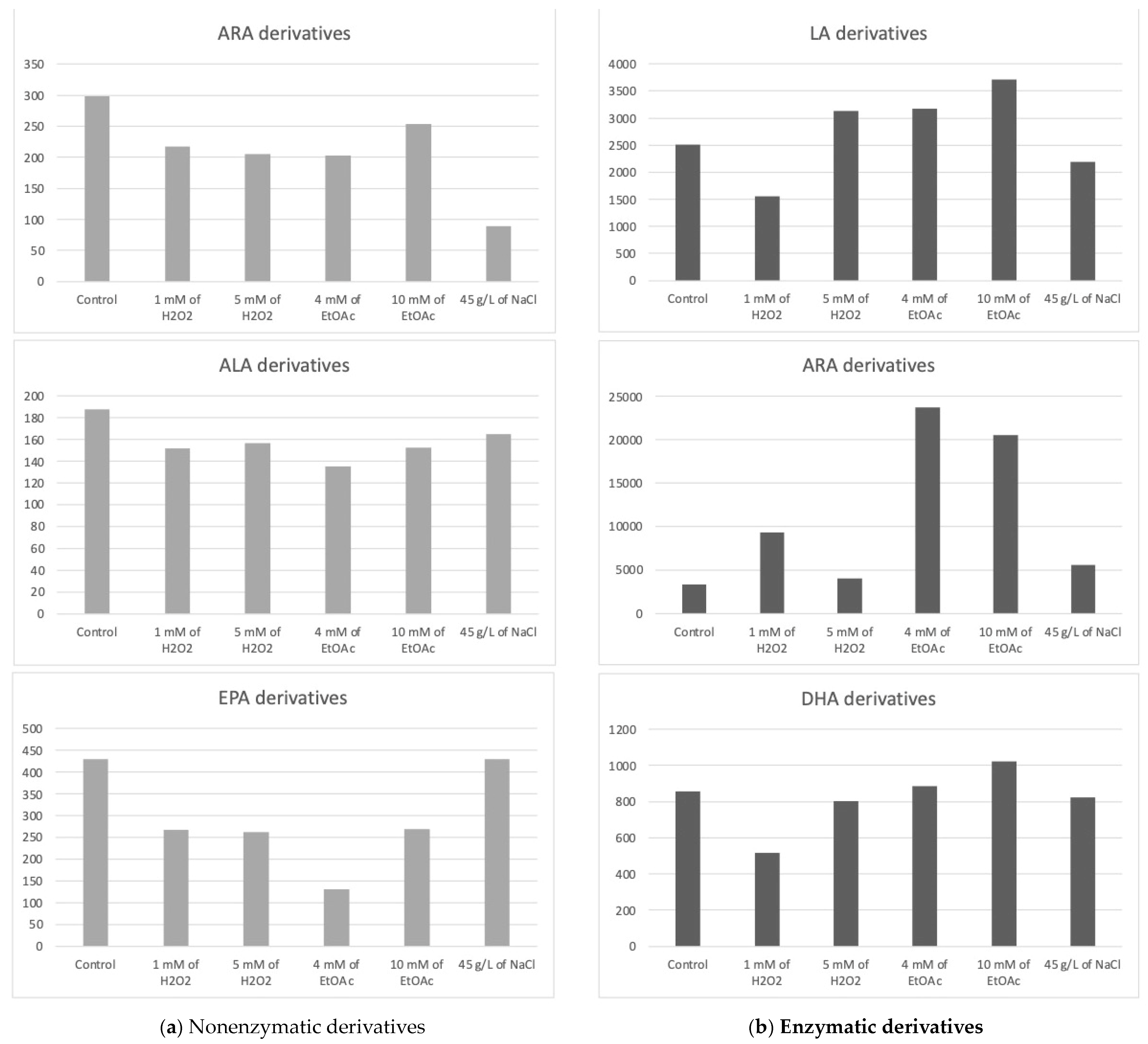
2.2. Profiles of Enzymatic and Nonenzymatic Oxygenated Metabolites in Stressed Mi136
2.2.1. Profiles of Nonenzymatic Oxygenated Metabolites
Derivatives from ARA
Derivatives from ALA
Derivatives from EPA
2.2.2. Profiles of the Enzymatic Oxygenated Metabolites
Derivatives from LA
Derivatives from ARA
Derivatives from EPA
Derivatives from DHA
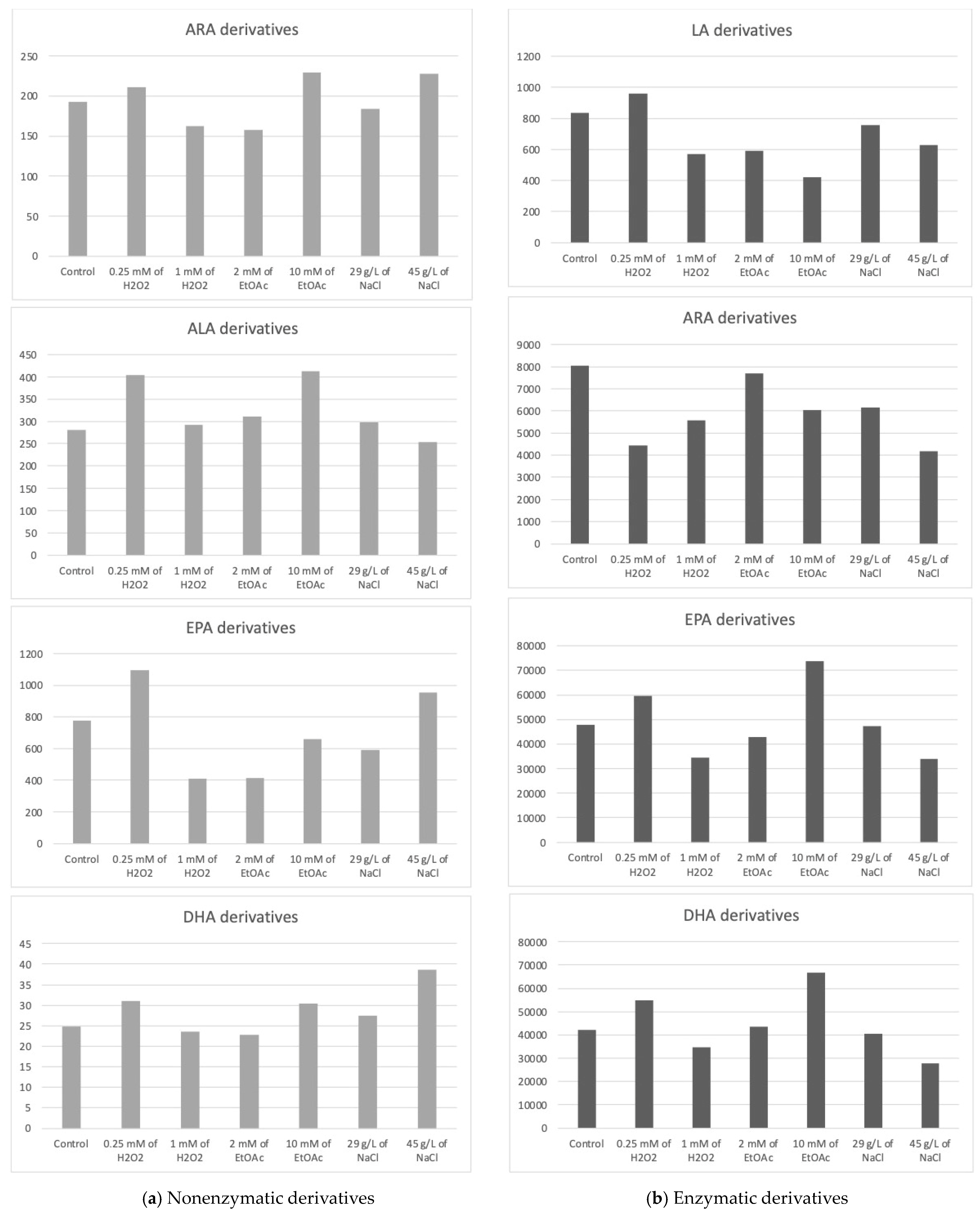
2.3. Profiles of Enzymatic and Nonenzymatic Oxygenated Metabolites in Stressed Mi168
2.3.1. Nonenzymatic Oxygenated Metabolite Profiles
Derivatives from ARA
Derivatives from DPA
Derivatives from ALA
Derivatives from EPA
Derivatives from DHA
2.3.2. Enzymatic Oxygenated Metabolites Profiles
Derivatives from LA
Derivatives from ARA
Derivatives from EPA
Derivatives from DHA
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Microalgal Species and Nonstressful Conditions
4.3. Viability Cell Tests
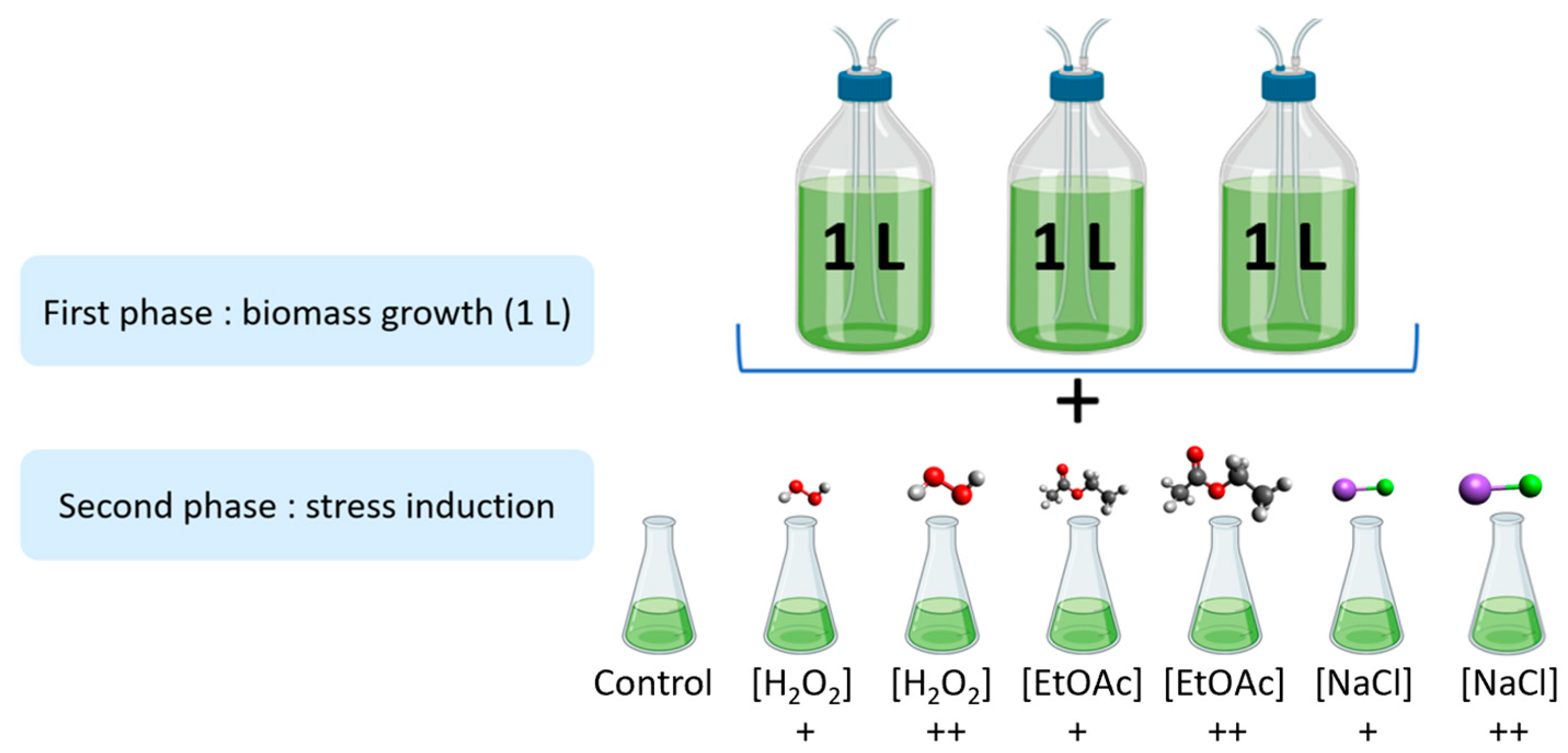
4.4. Stress Induction
4.5. Nonenzymatic Oxylipins Extraction and Analysis
4.6. Enzymatic Oxylipins Extraction and Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Worden, A.Z.; Follows, M.J.; Giovannoni, S.J.; Wilken, S.; Zimmerman, A.E.; Keeling, P.J. Rethinking the Marine Carbon Cycle: Factoring in the Multifarious Lifestyles of Microbes. Science 2015, 347, 1257594. [Google Scholar] [CrossRef]
- Jebali, A.; Sanchez, M.R.; Hanschen, E.R.; Starkenburg, S.R.; Corcoran, A.A. Trait Drift in Microalgae and Applications for Strain Improvement. Biotechnol. Adv. 2022, 60, 108034. [Google Scholar] [CrossRef]
- Dolganyuk, V.; Belova, D.; Babich, O.; Prosekov, A.; Ivanova, S.; Katserov, D.; Patyukov, N.; Sukhikh, S. Microalgae: A Promising Source of Valuable Bioproducts. Biomolecules 2020, 10, 1153. [Google Scholar] [CrossRef]
- Novoveská, L.; Nielsen, S.L.; Eroldoğan, O.T.; Haznedaroglu, B.Z.; Rinkevich, B.; Fazi, S.; Robbens, J.; Vasquez, M.; Einarsson, H. Overview and Challenges of Large-Scale Cultivation of Photosynthetic Microalgae and Cyanobacteria. Mar. Drugs 2023, 21, 445. [Google Scholar] [CrossRef] [PubMed]
- Abdur Razzak, S.; Bahar, K.; Islam, K.M.O.; Haniffa, A.K.; Faruque, M.O.; Hossain, S.M.Z.; Hossain, M.M. Microalgae Cultivation in Photobioreactors: Sustainable Solutions for a Greener Future. Green. Chem. Eng. 2023. [Google Scholar] [CrossRef]
- Linares-Maurizi, A.; Reversat, G.; Awad, R.; Bultel-Poncé, V.; Oger, C.; Galano, J.-M.; Balas, L.; Durbec, A.; Bertrand-Michel, J.; Durand, T.; et al. Bioactive Oxylipins Profile in Marine Microalgae. Mar. Drugs 2023, 21, 136. [Google Scholar] [CrossRef] [PubMed]
- Jónasdóttir, S.H. Fatty Acid Profiles and Production in Marine Phytoplankton. Mar. Drugs 2019, 17, 151. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Li, T.; Du, S.; Chen, H.; Wang, Q. Microalgal Polyunsaturated Fatty Acids: Hotspots and Production Techniques. Front. Bioeng. Biotechnol. 2023, 11, 1146881. [Google Scholar] [CrossRef]
- Ramesh Kumar, B.; Deviram, G.; Mathimani, T.; Duc, P.A.; Pugazhendhi, A. Microalgae as Rich Source of Polyunsaturated Fatty Acids. Biocatal. Agric. Biotechnol. 2019, 17, 583–588. [Google Scholar] [CrossRef]
- Brown, H.A.; Marnett, L.J. Introduction to Lipid Biochemistry, Metabolism, and Signaling. Chem. Rev. 2011, 111, 5817–5820. [Google Scholar] [CrossRef]
- Ahmed, O.S.; Galano, J.-M.; Pavlickova, T.; Revol-Cavalier, J.; Vigor, C.; Lee, J.C.-Y.; Oger, C.; Durand, T. Moving Forward with Isoprostanes, Neuroprostanes and Phytoprostanes: Where Are We Now? Essays Biochem. 2020, 64, 463–484. [Google Scholar] [CrossRef] [PubMed]
- Polyunsaturated Fatty Acids and Fatty Acid-Derived Lipid Mediators: Recent Advances in the Understanding of Their Biosynthesis, Structures, and Functions—ScienceDirect. Available online: https://www.sciencedirect.com/science/article/pii/S0163782722000200?via%3Dihub (accessed on 17 April 2024).
- Vigor, C.; Balas, L.; Guy, A.; Bultel-Poncé, V.; Reversat, G.; Galano, J.-M.; Durand, T.; Oger, C. Isoprostanoids, Isofuranoids and Isoketals—From Synthesis to Lipidomics. Eur. J. Org. Chem. 2022, 2022, e202101523. [Google Scholar] [CrossRef]
- Misheva, M.; Johnson, J.; McCullagh, J. Role of Oxylipins in the Inflammatory-Related Diseases NAFLD, Obesity, and Type 2 Diabetes. Metabolites 2022, 12, 1238. [Google Scholar] [CrossRef] [PubMed]
- Pohnert, G. Phospholipase A2 Activity Triggers the Wound-Activated Chemical Defense in the Diatom Thalassiosira Rotula. Plant Physiol. 2002, 129, 103–111. [Google Scholar] [CrossRef]
- Hajeyah, A.A.; Griffiths, W.J.; Wang, Y.; Finch, A.J.; O’Donnell, V.B. The Biosynthesis of Enzymatically Oxidized Lipids. Front. Endocrinol. 2020, 11, 591819. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, H.-P.; Han, B.; Yu, X. Coupling of Abiotic Stresses and Phytohormones for the Production of Lipids and High-Value by-Products by Microalgae: A Review. Bioresour. Technol. 2019, 274, 549–556. [Google Scholar] [CrossRef]
- Stirk, W.A.; van Staden, J. Potential of Phytohormones as a Strategy to Improve Microalgae Productivity for Biotechnological Applications. Biotechnol. Adv. 2020, 44, 107612. [Google Scholar] [CrossRef]
- Yang, Z.-Y.; Huang, K.-X.; Zhang, Y.-R.; Yang, L.; Zhou, J.-L.; Yang, Q.; Gao, F. Efficient Microalgal Lipid Production Driven by Salt Stress and Phytohormones Synergistically. Bioresour. Technol. 2023, 367, 128270. [Google Scholar] [CrossRef]
- Paliwal, C.; Mitra, M.; Bhayani, K.; Bharadwaj, S.V.V.; Ghosh, T.; Dubey, S.; Mishra, S. Abiotic Stresses as Tools for Metabolites in Microalgae. Bioresour. Technol. 2017, 244, 1216–1226. [Google Scholar] [CrossRef]
- Chen, B.; Wan, C.; Mehmood, M.A.; Chang, J.-S.; Bai, F.; Zhao, X. Manipulating Environmental Stresses and Stress Tolerance of Microalgae for Enhanced Production of Lipids and Value-Added Products–A Review. Bioresour. Technol. 2017, 244, 1198–1206. [Google Scholar] [CrossRef]
- Liu, W.; Au, D.W.T.; Anderson, D.M.; Lam, P.K.S.; Wu, R.S.S. Effects of Nutrients, Salinity, pH and Light:Dark Cycle on the Production of Reactive Oxygen Species in the Alga Chattonella Marina. J. Exp. Mar. Biol. Ecol. 2007, 346, 76–86. [Google Scholar] [CrossRef]
- Wang, X.; Fosse, H.K.; Li, K.; Chauton, M.S.; Vadstein, O.; Reitan, K.I. Influence of Nitrogen Limitation on Lipid Accumulation and EPA and DHA Content in Four Marine Microalgae for Possible Use in Aquafeed. Front. Mar. Sci. 2019, 6, 95. [Google Scholar] [CrossRef]
- Giarikos, D.G.; Brown, J.; Razeghifard, R.; Vo, D.; Castillo, A.; Nagabandi, N.; Gaffney, J.; Zelden, M.; Antakshinova, A.; Rodriguez, S.; et al. Effects of Nitrogen Depletion on the Biosorption Capacities of Neochloris Minuta and Neochloris Alveolaris for Five Heavy Metals. Appl. Water Sci. 2021, 11, 39. [Google Scholar] [CrossRef]
- Yaakob, M.A.; Mohamed, R.M.S.R.; Al-Gheethi, A.; Aswathnarayana Gokare, R.; Ambati, R.R. Influence of Nitrogen and Phosphorus on Microalgal Growth, Biomass, Lipid, and Fatty Acid Production: An Overview. Cells 2021, 10, 393. [Google Scholar] [CrossRef]
- Su, Y. Revisiting Carbon, Nitrogen, and Phosphorus Metabolisms in Microalgae for Wastewater Treatment. Sci. Total Environ. 2021, 762, 144590. [Google Scholar] [CrossRef]
- YuanKun, L. Algal Nutrition. In Handbook of Microalgal Culture: Biotechnology and Applied Phycology; Wiley: Hoboken, NJ, USA, 2004; pp. 116–124. [Google Scholar]
- Shetty, P.; Gitau, M.M.; Maróti, G. Salinity Stress Responses and Adaptation Mechanisms in Eukaryotic Green Microalgae. Cells 2019, 8, 1657. [Google Scholar] [CrossRef] [PubMed]
- Qiao, T.; Zhao, Y.; Zhong, D.; Yu, X. Hydrogen Peroxide and Salinity Stress Act Synergistically to Enhance Lipids Production in Microalga by Regulating Reactive Oxygen Species and Calcium. Algal Res. 2021, 53, 102017. [Google Scholar] [CrossRef]
- Bazzani, E.; Lauritano, C.; Mangoni, O.; Bolinesi, F.; Saggiomo, M. Chlamydomonas Responses to Salinity Stress and Possible Biotechnological Exploitation. J. Mar. Sci. Eng. 2021, 9, 1242. [Google Scholar] [CrossRef]
- Tammam, A.A.; Fakhry, E.M.; El-Sheekh, M. Effect of Salt Stress on Antioxidant System and the Metabolism of the Reactive Oxygen Species in Dunaliella Salina and Dunaliella Tertiolecta. Afr. J. Biotechnol. 2011, 10, 3795–3808. [Google Scholar] [CrossRef]
- Lupette, J.; Jaussaud, A.; Vigor, C.; Oger, C.; Galano, J.-M.; Réversat, G.; Vercauteren, J.; Jouhet, J.; Durand, T.; Maréchal, E. Non-Enzymatic Synthesis of Bioactive Isoprostanoids in the Diatom Phaeodactylum Following Oxidative Stress. Plant Physiol. 2018, 178, 1344–1357. [Google Scholar] [CrossRef]
- Pinto, E.; Sigaud-kutner, T.C.S.; Leitão, M.A.S.; Okamoto, O.K.; Morse, D.; Colepicolo, P. Heavy Metal–Induced Oxidative Stress in Algae1. J. Phycol. 2003, 39, 1008–1018. [Google Scholar] [CrossRef]
- Collins, J.R.; Edwards, B.R.; Fredricks, H.F.; Van Mooy, B.A.S. LOBSTAHS: An Adduct-Based Lipidomics Strategy for Discovery and Identification of Oxidative Stress Biomarkers. Anal. Chem. 2016, 88, 7154–7162. [Google Scholar] [CrossRef]
- Anand, V.; Kashyap, M.; Sharma, M.P.; Bala, K. Impact of Hydrogen Peroxide on Microalgae Cultivated in Varying Salt-Nitrate-Phosphate Conditions. J. Environ. Chem. Eng. 2021, 9, 105814. [Google Scholar] [CrossRef]
- Roussel, A.; Mériot, V.; Jauffrais, T.; Berteaux-Lecellier, V.; Lebouvier, N. OMICS Approaches to Assess Dinoflagellate Responses to Chemical Stressors. Biology 2023, 12, 1234. [Google Scholar] [CrossRef]
- Vigor, C.; Oger, C.; Reversat, G.; Rocher, A.; Zhou, B.; Linares-Maurizi, A.; Guy, A.; Bultel-Poncé, V.; Galano, J.-M.; Vercauteren, J.; et al. Isoprostanoid Profiling of Marine Microalgae. Biomolecules 2020, 10, 1073. [Google Scholar] [CrossRef] [PubMed]
- Cunha, A.E.; Sátiro, J.R.; Escobar, B.P.; Simões, R.M. Chlorella Vulgaris Growth, Pigment and Lipid Accumulation: Effect of Progressive Light and Hydrogen Peroxide Exposure. J. Chem. Technol. Biotechnol. 2023, 98, 442–450. [Google Scholar] [CrossRef]
- Xie, H.; Zhao, X.; Zhou, G.; He, X.; Lan, H.; Jiang, Z. Investigating the Performance of CoxOy/Activated Carbon Catalysts for Ethyl Acetate Catalytic Combustion. Appl. Surf. Sci. 2015, 326, 119–123. [Google Scholar] [CrossRef]
- Zhang, C.; Hasunuma, T.; Shiung Lam, S.; Kondo, A.; Ho, S.-H. Salinity-Induced Microalgal-Based Mariculture Wastewater Treatment Combined with Biodiesel Production. Bioresour. Technol. 2021, 340, 125638. [Google Scholar] [CrossRef]
- Kumar Patel, A.; Tseng, Y.-S.; Rani Singhania, R.; Chen, C.-W.; Chang, J.-S.; Di Dong, C. Novel Application of Microalgae Platform for Biodesalination Process: A Review. Bioresour. Technol. 2021, 337, 125343. [Google Scholar] [CrossRef]
- Asai, K.; Hirabayashi, T.; Houjou, T.; Uozumi, N.; Taguchi, R.; Shimizu, T. Human Group IVC Phospholipase A2(cPLA2γ): ROLES IN THE MEMBRANE REMODELING AND ACTIVATION INDUCED BY OXIDATIVE STRESS *. J. Biol. Chem. 2003, 278, 8809–8814. [Google Scholar] [CrossRef]
- Guihéneuf, F.; Stengel, D.B. LC-PUFA-Enriched Oil Production by Microalgae: Accumulation of Lipid and Triacylglycerols Containing n-3 LC-PUFA Is Triggered by Nitrogen Limitation and Inorganic Carbon Availability in the Marine Haptophyte Pavlova Lutheri. Mar. Drugs 2013, 11, 4246–4266. [Google Scholar] [CrossRef] [PubMed]
- Hikihara, R.; Yamasaki, Y.; Shikata, T.; Nakayama, N.; Sakamoto, S.; Kato, S.; Hatate, H.; Tanaka, R. Analysis of Phytosterol, Fatty Acid, and Carotenoid Composition of 19 Microalgae and 6 Bivalve Species. J. Aquat. Food Prod. Technol. 2020, 29, 461–479. [Google Scholar] [CrossRef]
- Cañavate, J.P. Advancing Assessment of Marine Phytoplankton Community Structure and Nutritional Value from Fatty Acid Profiles of Cultured Microalgae. Rev. Aquac. 2019, 11, 527–549. [Google Scholar] [CrossRef]
- Delbrut, A.; Albina, P.; Lapierre, T.; Pradelles, R.; Dubreucq, E. Fucoxanthin and Polyunsaturated Fatty Acids Co-Extraction by a Green Process. Molecules 2018, 23, 874. [Google Scholar] [CrossRef] [PubMed]
- Premaratne, M.; Liyanaarachchi, V.; Nimarshana, P.; Ariyadasa, T.; Malik, A.; Attalage, R. Co-Production of Fucoxanthin, Docosahexaenoic Acid (DHA) and Bioethanol from the Marine Microalga Tisochrysis Lutea. Biochem. Eng. J. 2021, 176, 108160. [Google Scholar] [CrossRef]
- Steinrücken, P.; Prestegard, S.K.; de Vree, J.H.; Storesund, J.E.; Pree, B.; Mjøs, S.A.; Erga, S.R. Comparing EPA Production and Fatty Acid Profiles of Three Phaeodactylum Tricornutum Strains under Western Norwegian Climate Conditions. Algal Res. 2018, 30, 11–22. [Google Scholar] [CrossRef]
- Stiefvatter, L.; Lehnert, K.; Frick, K.; Montoya-Arroyo, A.; Frank, J.; Vetter, W.; Schmid-Staiger, U.; Bischoff, S.C. Oral Bioavailability of Omega-3 Fatty Acids and Carotenoids from the Microalgae Phaeodactylum Tricornutum in Healthy Young Adults. Mar. Drugs 2021, 19, 700. [Google Scholar] [CrossRef]
- He, Q.; Yang, H.; Wu, L.; Hu, C. Effect of Light Intensity on Physiological Changes, Carbon Allocation and Neutral Lipid Accumulation in Oleaginous Microalgae. Bioresour. Technol. 2015, 191, 219–228. [Google Scholar] [CrossRef]
- Zhang, Y.-M.; Chen, H.; He, C.-L.; Wang, Q. Nitrogen Starvation Induced Oxidative Stress in an Oil-Producing Green Alga Chlorella Sorokiniana C3. PLoS ONE 2013, 8, e69225. [Google Scholar] [CrossRef]
- Yilancioglu, K.; Cokol, M.; Pastirmaci, I.; Erman, B.; Cetiner, S. Oxidative Stress Is a Mediator for Increased Lipid Accumulation in a Newly Isolated Dunaliella Salina Strain. PLoS ONE 2014, 9, e91957. [Google Scholar] [CrossRef]
- Renaud, S.M.; Thinh, L.-V.; Lambrinidis, G.; Parry, D.L. Effect of Temperature on Growth, Chemical Composition and Fatty Acid Composition of Tropical Australian Microalgae Grown in Batch Cultures. Aquaculture 2002, 211, 195–214. [Google Scholar] [CrossRef]
- Church, J.; Hwang, J.-H.; Kim, K.-T.; McLean, R.; Oh, Y.-K.; Nam, B.; Joo, J.C.; Lee, W.H. Effect of Salt Type and Concentration on the Growth and Lipid Content of Chlorella Vulgaris in Synthetic Saline Wastewater for Biofuel Production. Bioresour. Technol. 2017, 243, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Arvin, E.; Pedersen, L.-F. Hydrogen Peroxide Decomposition Kinetics in Aquaculture Water. Aquac. Eng. 2015, 64, 1–7. [Google Scholar] [CrossRef]
- Wang, T.; Ge, H.; Liu, T.; Tian, X.; Wang, Z.; Guo, M.; Chu, J.; Zhuang, Y. Salt Stress Induced Lipid Accumulation in Heterotrophic Culture Cells of Chlorella Protothecoides: Mechanisms Based on the Multi-Level Analysis of Oxidative Response, Key Enzyme Activity and Biochemical Alteration. J. Biotechnol. 2016, 228, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Nieman, D.C.; Meaney, M.P.; John, C.S.; Knagge, K.J.; Chen, H. 9- and 13-Hydroxy-Octadecadienoic Acids (9+13 HODE) Are Inversely Related to Granulocyte Colony Stimulating Factor and IL-6 in Runners after 2h Running. Brain Behav. Immun. 2016, 56, 246–252. [Google Scholar] [CrossRef] [PubMed]
- White, P.J.; Arita, M.; Taguchi, R.; Kang, J.X.; Marette, A. Transgenic Restoration of Long-Chain n-3 Fatty Acids in Insulin Target Tissues Improves Resolution Capacity and Alleviates Obesity-Linked Inflammation and Insulin Resistance in High-Fat–Fed Mice. Diabetes 2010, 59, 3066–3073. [Google Scholar] [CrossRef] [PubMed]
- Hagihara, M.; Yamashita, M.; Ariyoshi, T.; Eguchi, S.; Minemura, A.; Miura, D.; Higashi, S.; Oka, K.; Nonogaki, T.; Mori, T.; et al. Clostridium Butyricum-Induced ω-3 Fatty Acid 18-HEPE Elicits Anti-Influenza Virus Pneumonia Effects through Interferon-λ Upregulation. Cell Rep. 2022, 41, 111755. [Google Scholar] [CrossRef]
- Collado-González, J.; Durand, T.; Ferreres, F.; Medina, S.; Torrecillas, A.; Gil-Izquierdo, Á. Phytoprostanes. Lipid Technology 2015, 27, 127–130. [Google Scholar] [CrossRef]
- Guillard, R.R.; Ryther, J.H. Studies of Marine Planktonic Diatoms. I. Cyclotella Nana Hustedt, and Detonula Confervacea (Cleve) Gran. Can. J. Microbiol. 1962, 8, 229–239. [Google Scholar] [CrossRef]
- Le Faouder, P.; Baillif, V.; Spreadbury, I.; Motta, J.-P.; Rousset, P.; Chêne, G.; Guigné, C.; Tercé, F.; Vanner, S.; Vergnolle, N.; et al. LC–MS/MS Method for Rapid and Concomitant Quantification of pro-Inflammatory and pro-Resolving Polyunsaturated Fatty Acid Metabolites. J. Chromatogr. B 2013, 932, 123–133. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mi124 | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Oxylipins | Control | 1 mM of H2O2 | 4 mM of H2O2 | 0.5 mM of EtOAc | 1 mΜ of EtOAc | 45 g/L of NaCl | ||||||||||||
| Concentration (pg/mg) | ± | SD | Concentration (pg/mg) | ± | SD | Concentration (pg/mg) | ± | SD | Concentration (pg/mg) | ± | SD | Concentration (pg/mg) | ± | SD | Concentration (pg/mg) | ± | SD | |
| 9-HODE | 4.58 × 103 | ± | 3.12 × 102 | 9.14 × 103 | ± | 5.95 × 102 | 9.33 × 103 | ± | 1.87 × 103 | 3.87 × 103 | ± | 6.53 × 102 | 3.15 × 103 | ± | 8.08 × 102 | 3.09 × 103 | ± | 3.53 × 102 |
| 13-HODΕ | 1.38 × 104 | ± | 3.00 × 103 | 1.40 × 104 | ± | 1.76 × 103 | 1.88 × 104 | ± | 6.97 × 103 | 1.11 × 104 | ± | 5.13 × 103 | 9.71 × 103 | ± | 5.71 × 103 | 7.50 × 103 | ± | 2.08 × 103 |
| 5oxoETE | 3.12 × 10 | ± | 9.24 | 3.04 × 10 | ± | 2.60 × 10 | 9.28 × 10 | ± | 2.05 × 10 | 1.97 × 10 | ± | 1.47 × 10 | 1.78 × 10 | ± | 1.50 × 10 | 2.07 × 10 | ± | 8.41 |
| 5-HΕΤΕ | 3.95 × 10 | ± | 2.02 | 5.55 × 10 | ± | 2.54 | 1.52 × 102 | ± | 4.34 × 10 | 3.18 × 10 | ± | 1.27 × 10 | 2.11 × 10 | ± | 1.10 × 10 | 2.40 × 10 | ± | 8.37 |
| 8-HΕΤΕ | 2.19 × 10 | ± | 8.33 | 2.72 × 10 | ± | 2.10 | 6.46 × 10 | ± | 1.62 × 10 | 1.79 × 10 | ± | 9.25 | 1.43 × 10 | ± | 1.10 × 10 | 1.22 × 10 | ± | 2.55 |
| 12-HΕΤΕ | 3.18 × 10 | ± | 1.19 × 10 | 4.83 × 10 | ± | 1.14 × 10 | 1.21 × 102 | ± | 2.24 × 10 | 3.13 × 10 | ± | 2.35 × 10 | 1.88 × 10 | ± | 1.55 × 10 | 1.12 × 10 | ± | 8.60 |
| 15-HΕΤΕ | 4.49 × 102 | ± | 1.23 × 102 | 3.57 × 102 | ± | 1.01 × 102 | 7.76 × 102 | ± | 3.42 × 102 | 3.31 × 102 | ± | 2.03 × 102 | 2.79 × 102 | ± | 1.96 × 102 | 2.10 × 102 | ± | 8.19 × 10 |
| 14.15-ΕΕΤ | 2.51 × 10 | ± | 1.79 × 10 | 1.71 × 10 | ± | 1.23 × 10 | 3.66 × 10 | ± | 1.55 × 10 | 2.73 × 10 | ± | 2.10 × 10 | 1.57 × 10 | ± | 2.12 × 10 | 4.85 | ± | 6.85 |
| 9(R).16(R.S)-Linotrines | 4.30 × 102 | ± | 4.96 × 10 | 4.04 × 102 | ± | 9.40 × 10 | 4.24 × 102 | ± | 1.42 × 102 | 2.89 × 102 | ± | 2.19 × 10 | 1.91 × 102 | ± | 6.39 × 10 | 2.7 × 102 | ± | 6.59 × 10 |
| 18-HEPE | 8.55 × 102 | ± | 2.34 × 102 | 7.53 × 102 | ± | 8.60 | 1.01 × 103 | ± | 2.36 × 102 | 6.81 × 102 | ± | 2.53 × 102 | 5.87 × 102 | ± | 4.02 × 102 | 5.43 × 102 | ± | 2.08 × 102 |
| ΡDx | 1.04 × 102 | ± | 5.17 × 10 | 3.83 × 10 | ± | 1.03 × 10 | 4.85 × 10 | ± | 2.48 × 10 | 8.87 × 10 | ± | 5.57 × 10 | 8.42 × 10 | ± | 9.81 × 10 | 3.15 × 10 | ± | 1.94 × 10 |
| 14-HDoHE | 1.07 × 104 | ± | 1.23 × 103 | 1.15 × 104 | ± | 1.39 × 103 | 1.33 × 104 | ± | 4.03 × 103 | 8.65 × 103 | ± | 3.06 × 103 | 6.27 × 103 | ± | 3.08 × 103 | 6.20 × 103 | ± | 2.22 × 103 |
| 17-HDoHE | 1.22 × 105 | ± | 3.81 × 104 | 9.10 × 104 | ± | 2.11 × 104 | 9.06 × 104 | ± | 3.05 × 104 | 9.31 × 104 | ± | 5.57 × 104 | 8.07 × 104 | ± | 5.98 × 104 | 6.28 × 104 | ± | 3.68 × 104 |
| 9-L1t-PhytoP | 65.31 | ± | 11.53 | 90.89 | ± | 7.24 | 89.25 | ± | 1.89 | 50.80 | ± | 6.89 | 66.48 | ± | 12.64 | 59.95 | ± | 8.57 |
| 9-F1t-PhytoP | 32.88 | ± | 2.31 | 64.26 | ± | 17.70 | 54.69 | ± | 4.54 | 28.53 | ± | 5.43 | 30.30 | ± | 1.57 | 49.25 | ± | 12.30 |
| 16-B1t-PhytoP | 64.90 | ± | 9.96 | 94.93 | ± | 8.60 | 92.14 | ± | 2.15 | 52.04 | ± | 6.88 | 66.24 | ± | 11.28 | 70.40 | ± | 11.31 |
| 16-epi-16-F1t-PhytoP | 136.68 | ± | 35.56 | 154.10 | ± | 29.85 | 166.69 | ± | 43.22 | 162.60 | ± | 12.12 | 158.19 | ± | 68.11 | 97.43 | ± | 17.70 |
| 16-F1t-PhytoP | 191.01 | ± | 51.00 | 202.69 | ± | 32.71 | 234.50 | ± | 66.10 | 232.24 | ± | 15.52 | 228.76 | ± | 102.82 | 132.98 | ± | 23.32 |
| ent-9A-12-epi-ST-Δ10-13-PhytoF | 44.25 | ± | 13.08 | 62.05 | ± | 3.04 | 55.00 | ± | 4.73 | 33.29 | ± | 7.76 | 54.11 | ± | 14.81 | 40.92 | ± | 4.58 |
| ent-9B-12-epi-ST-Δ10-13-PhytoF | 14.75 | ± | 4.45 | 20.95 | ± | 2.77 | 16.44 | ± | 1.54 | 11.42 | ± | 2.67 | 16.54 | ± | 4.35 | 19.95 | ± | 3.44 |
| ent-16A-9-epi-ST-Δ14-10-PhytoF | 95.51 | ± | 31.37 | 105.70 | ± | 11.86 | 94.80 | ± | 18.53 | 66.76 | ± | 14.37 | 122.56 | ± | 47.00 | 49.93 | ± | 6.80 |
| ent-16B-9-epi-ST-Δ14-10-PhytoF | 122.81 | ± | 42.73 | 131.16 | ± | 15.43 | 115.29 | ± | 22.14 | 83.16 | ± | 9.69 | 146.41 | ± | 51.37 | 58.83 | ± | 10.08 |
| ent-16A-13-epi-ST-Δ14-9-PhytoF | 15.75 | ± | 1.21 | 21.30 | ± | 2.10 | 20.83 | ± | 0.90 | 13.70 | ± | 3.21 | 14.77 | ± | 2.42 | 16.24 | ± | 1.21 |
| ent-16B-13-epi-ST-Δ14-9-PhytoF | 23.87 | ± | 7.07 | 26.67 | ± | 3.92 | 24.34 | ± | 4.66 | 16.14 | ± | 4.32 | 28.32 | ± | 11.17 | 12.93 | ± | 1.48 |
| 5(R)-5-F3t-lsoP | 40.49 | ± | 28.74 | 110.61 | ± | 92.55 | 31.46 | ± | 22.71 | 116.24 | ± | 103.04 | 27.35 | ± | 19.60 | 57.62 | ± | 19.22 |
| 8(R)-8-F3t-lsoP | 10.31 | ± | 3.35 | 16.28 | ± | 0.67 | 12.64 | ± | 2.30 | 10.68 | ± | 2.36 | 16.43 | ± | 10.83 | 6.45 | ± | 1.89 |
| 18(R)-8-F3t-IsoP | 56.50 | ± | 9.99 | 111.57 | ± | 83.51 | 75.67 | ± | 16.67 | 144.45 | ± | 122.63 | 54.16 | ± | 38.59 | 54.00 | ± | 5.26 |
| 4(RS)-4-F4t-NeuroP | 15.47 | ± | 1.78 | 37.48 | ± | 13.54 | 25.77 | ± | 0.72 | 14.51 | ± | 2.30 | 15.61 | ± | 3.68 | 20.32 | ± | 3.23 |
| 10(R)-10-F4t-NeuroP | 27.06 | ± | 18.53 | 12.61 | ± | 3.63 | 15.96 | ± | 3.46 | 18.54 | ± | 4.06 | 20.37 | ± | 9.16 | 14.45 | ± | 0.90 |
| 10(S)-10-F4t-NeuroP | 23.49 | ± | 6.60 | 16.37 | ± | 6.44 | 19.77 | ± | 2.76 | 20.81 | ± | 5.48 | 18.09 | ± | 6.77 | 11.81 | ± | 1.67 |
| 13A(RS)-13-F4t-NeuroP | 16.55 | ± | 3.76 | 28.61 | ± | 6.14 | 24.18 | ± | 2.19 | 16.01 | ± | 4.49 | 20.80 | ± | 2.72 | 19.71 | ± | 2.50 |
| 13B(RS)-13-F4t-NeuroP | 13.57 | ± | 4.57 | 23.01 | ± | 6.00 | 17.27 | ± | 2.32 | 11.80 | ± | 3.52 | 11.60 | ± | 1.90 | 17.22 | ± | 5.37 |
| Mi136 | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Oxylipins | Control | 1 mM of H2O2 | 5 mM of H2O2 | 4 mM of EtOAc | 10 mΜ of EtOAc | 45 g/L of NaCl | ||||||||||||
| Concentration (pg/mg) | ± | SD | Concentration (pg/mg) | ± | SD | Concentration (pg/mg) | ± | SD | Concentration (pg/mg) | ± | SD | Concentration (pg/mg) | ± | SD | Concentration (pg/mg) | ± | SD | |
| 9-HODE | 9.26 × 102 | ± | 1.54 × 102 | 5.44 × 102 | ± | 1.24 × 102 | 1.17 × 103 | ± | 1.46 × 102 | 1.16 × 103 | ± | 2.54 × 10 | 1.31 × 103 | ± | 2.06 × 102 | 8.48 × 102 | ± | 1.42 × 102 |
| 13-HODΕ | 1.59 × 103 | ± | 2.89 × 102 | 1.01 × 103 | ± | 1.91 × 102 | 1.96 × 103 | ± | 2.40 × 102 | 2.02 × 103 | ± | 8.21 × 10 | 2.40 × 103 | ± | 2.86 × 102 | 1.35 × 103 | ± | 2.66 × 102 |
| 5-HΕΤΕ | 2.49 × 102 | ± | 2.71 × 10 | 1.95 × 102 | ± | 7.63 | 3.11 × 102 | ± | 8.77 | 2.53 × 102 | ± | 8.86 | 2.20 × 102 | ± | 4.75 × 10 | 1.96 × 102 | ± | 4.23 × 10 |
| 8-HΕΤΕ | 1.60 × 102 | ± | 1.23 × 10 | 1.17 × 102 | ± | 1.25 × 10 | 1.93 × 102 | ± | 1.11 × 10 | 1.79 × 102 | ± | 1.45 × 10 | 1.63 × 102 | ± | 3.34 × 10 | 1.31 × 102 | ± | 1.49 × 10 |
| 12-HΕΤΕ | 2.82 × 102 | ± | 7.18 | 1.82 × 102 | ± | 2.46 × 10 | 2.84 × 102 | ± | 1.47 × 10 | 2.93 × 102 | ± | 2.03 × 10 | 2.77 × 102 | ± | 4.63 × 10 | 1.75 × 102 | ± | 2.93 × 10 |
| 15-HΕΤΕ | 3.49 × 102 | ± | 3.70 × 10 | 2.85 × 102 | ± | 2.79 × 10 | 3.59 × 102 | ± | 1.42 × 10 | 3.83 × 102 | ± | 1.57 | 3.58 × 102 | ± | 4.86 × 10 | 1.98 × 102 | ± | 1.26 × 10 |
| 5oxoETE | 1.19 × 102 | ± | 1.98 × 10 | 9.26 × 10 | ± | 5.77 | 8.74 × 10 | ± | 1.53 × 10 | 1.05 × 102 | ± | 1.07 × 10 | 1.07 × 102 | ± | 1.85 × 10 | 8.19 × 10 | ± | 2.23 × 10 |
| 8.9-EET | 1.35 × 102 | ± | 2.82 × 10 | 1.23 × 102 | ± | 2.32 × 10 | 1.38 × 102 | ± | 2.14 × 10 | 1.47 × 102 | ± | 1.85 | 1.42 × 102 | ± | 1.65 × 10 | 8.80 × 10 | ± | 3.45 × 10 |
| 11.12-EET | 1.50 × 102 | ± | 2.27 × 10 | 1.23 × 102 | ± | 2.48 × 10 | 1.30 × 102 | ± | 2.79 × 10 | 1.46 × 102 | ± | 1.74 × 10 | 1.13 × 102 | ± | 7.94 | 7.00 × 10 | ± | 1.94 × 10 |
| 14.15-ΕΕΤ | 1.43 × 102 | ± | 3.70 × 10 | 1.24 × 102 | ± | 3.57 × 10 | 1.10 × 102 | ± | 3.85 × 10 | 1.59 × 102 | ± | 9.89 | 1.16 × 102 | ± | 2.00 × 10 | 6.08 × 10 | ± | 2.25 × 10 |
| PGE2 | 1.80 × 102 | ± | 2.70 × 102 | 3.64 × 102 | ± | 4.82 × 102 | 9.01 × 10 | ± | 8.28 × 10 | 1.04 × 103 | ± | 1.05 × 103 | 6.87 × 102 | ± | 8.42 × 102 | 3.42 × 102 | ± | 2.19 × 102 |
| PGE3 | 1.51 × 103 | ± | 1.54 × 103 | 7.43 × 103 | ± | 4.02 × 103 | 2.23 × 103 | ± | 2.07 × 103 | 2.04 × 104 | ± | 8.99 × 103 | 1.81 × 104 | ± | 1.64 × 104 | 4.22 × 103 | ± | 5.71 × 103 |
| 15-d-PGJ2 | 3.59 × 10 | ± | 6.16 × 10 | 2.97 × 102 | ± | 1.69 × 103 | 6.66 × 10 | ± | 1.50 × 10 | 5.81 × 102 | ± | 1.08 × 103 | 2.19 × 102 | ± | 3.44 × 10 | 4.21 × 10 | ± | 1.12 × 102 |
| 18-HEPE | 4.73 × 104 | ± | 1.05 × 105 | 2.36 × 105 | ± | 1.27 × 106 | 6.69 × 104 | ± | 1.78 × 104 | 6.82 × 105 | ± | 1.09 × 106 | 2.62 × 105 | ± | 2.47 × 104 | 1.26 × 105 | ± | 1.17 × 105 |
| 14-HDoHE | 3.73 × 102 | ± | 7.66 × 10 | 2.25 × 102 | ± | 5.08 × 10 | 3.61 × 102 | ± | 4.39 × 10 | 4.20 × 102 | ± | 2.67 × 10 | 4.66 × 102 | ± | 8.73 × 10 | 3.85 × 102 | ± | 5.08 × 10 |
| 17-HDoHE | 4.83 × 102 | ± | 9.38 × 10 | 2.90 × 102 | ± | 8.94 × 10 | 4.40 × 102 | ± | 4.88 × 10 | 4.64 × 102 | ± | 5.74 × 10 | 5.56 × 102 | ± | 1.13 × 102 | 4.38 × 102 | ± | 7.32 × 10 |
| 5(RS)-5-F2t-lsoP | 167.17 | ± | 89.06 | 115.41 | ± | 40.19 | 109.59 | ± | 10.60 | 97.28 | ± | 20.92 | 116.45 | ± | 49.26 | 55.15 | ± | 8.06 |
| 15-A2-IsoP | 130.51 | ± | 46.56 | 101.89 | ± | 21.26 | 94.74 | ± | 7.68 | 105.64 | ± | 19.85 | 138.11 | ± | 55.59 | 34.07 | ± | 7.63 |
| 9-L1t-PhytoP | 6.55 | ± | 4.64 | 3.50 | ± | 0.94 | 3.05 | ± | 1.10 | 2.98 | ± | 0.73 | 4.64 | ± | 1.12 | 9.00 | ± | 10.73 |
| 9-F1t-PhytoP | 8.97 | ± | 12.69 | 3.19 | ± | 4.51 | 2.23 | ± | 3.15 | 0.00 | ± | 0.00 | 0.00 | ± | 0.00 | 4.69 | ± | 6.63 |
| 16-B1t-PhytoP | 8.43 | ± | 7.14 | 4.35 | ± | 1.43 | 3.57 | ± | 1.35 | 3.54 | ± | 0.66 | 4.70 | ± | 1.23 | 11.24 | ± | 13.06 |
| 16-epi-16-F1t-PhytoP | 57.05 | ± | 3.84 | 49.25 | ± | 8.16 | 56.88 | ± | 9.46 | 45.42 | ± | 11.67 | 50.85 | ± | 3.25 | 52.50 | ± | 6.67 |
| 16-F1t-PhytoP | 68.99 | ± | 5.64 | 62.36 | ± | 11.22 | 71.36 | ± | 12.47 | 62.42 | ± | 11.31 | 64.17 | ± | 6.92 | 68.13 | ± | 6.81 |
| ent-9A-12-epi-ST-Δ10-13-PhytoF | 5.75 | ± | 2.69 | 5.80 | ± | 2.06 | 3.64 | ± | 0.35 | 3.96 | ± | 0.82 | 4.60 | ± | 1.13 | 4.07 | ± | 2.64 |
| ent-16A-9-epi-ST-Δ14-10-PhytoF | 6.95 | ± | 0.75 | 5.83 | ± | 1.47 | 6.60 | ± | 0.59 | 6.81 | ± | 1.53 | 8.42 | ± | 1.76 | 5.48 | ± | 3.50 |
| ent-16B-9-epi-ST-Δ14-10-PhytoF | 10.05 | ± | 1.16 | 8.70 | ± | 1.88 | 9.34 | ± | 1.25 | 8.46 | ± | 1.81 | 11.17 | ± | 2.40 | 6.88 | ± | 6.56 |
| ent-16A-13-epi-ST-Δ14-9-PhytoF | 9.09 | ± | 11.95 | 4.95 | ± | 5.86 | 1.03 | ± | 0.27 | 0.68 | ± | 0.11 | 0.83 | ± | 0.16 | 1.50 | ± | 1.15 |
| ent-16B-13-epi-ST-Δ14-9-PhytoF | 5.49 | ± | 4.58 | 4.55 | ± | 4.42 | 1.73 | ± | 0.48 | 2.04 | ± | 0.60 | 3.16 | ± | 1.35 | 1.41 | ± | 1.02 |
| 5(R)-5-F3t-lsoP | 165.29 | ± | 27.74 | 112.96 | ± | 7.80 | 139.18 | ± | 16.00 | 112.42 | ± | 11.47 | 142.33 | ± | 26.24 | 242.00 | ± | 54.01 |
| 5(S)-5-F3t-lsoP | 76.83 | ± | 23.15 | 43.78 | ± | 3.55 | 53.92 | ± | 10.80 | 45.54 | ± | 10.36 | 50.77 | ± | 11.22 | 88.03 | ± | 24.68 |
| 8(R)-8-F3t-lsoP | 16.41 | ± | 6.10 | 21.36 | ± | 3.24 | 15.82 | ± | 2.62 | 20.11 | ± | 2.20 | 14.45 | ± | 2.47 | 17.88 | ± | 2.20 |
| 18(R)-8-F3t-IsoP | 163.16 | ± | 172.45 | 77.53 | ± | 55.02 | 42.79 | ± | 8.53 | 39.20 | ± | 10.63 | 44.79 | ± | 17.65 | 52.50 | ± | 11.36 |
| 18(S)-8-F3t-IsoP | 7.84 | ± | 6.18 | 10.72 | ± | 1.73 | 10.93 | ± | 2.26 | 13.68 | ± | 0.69 | 16.72 | ± | 2.68 | 29.24 | ± | 8.78 |
| Mi168 | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Oxylipins | Control | 0.25 mM of H2O2 | 1 mM of H2O2 | 2 mM of EtOAc | 10 mΜ of EtOAc | 29 g/L of NaCl | 45 g/L of NaCl | ||||||||||||||
| Concentration (pg/mg) | ± | SD | Concentration (pg/mg) | ± | SD | Concentration (pg/mg) | ± | SD | Concentration (pg/mg) | ± | SD | Concentration (pg/mg) | ± | SD | Concentration (pg/mg) | ± | SD | Concentration (pg/mg) | ± | SD | |
| 9-HODE | 3.70 × 102 | ± | 3.27 × 10 | 4.39 × 102 | ± | 1.72 × 102 | 2.49 × 102 | ± | 6.53 × 10 | 2.62 × 102 | ± | 4.07 × 10 | 4.22 × 102 | ± | 6.45 × 10 | 3.47 × 102 | ± | 7.87 × 10 | 2.70 × 102 | ± | 3.28 |
| 13-HODΕ | 4.65 × 102 | ± | 4.61 × 10 | 5.21 × 102 | ± | 1.91 × 102 | 3.22 × 102 | ± | 9.20 × 10 | 3.31 × 102 | ± | 6.70 × 10 | 5.20 × 102 | ± | 7.22 × 10 | 4.12 × 102 | ± | 7.05 × 10 | 3.58 × 102 | ± | 2.02 × 10 |
| 5-HΕΤΕ | 3.66 × 103 | ± | 6.23 × 10 | 4.28 × 103 | ± | 1.70 × 103 | 2.57 × 103 | ± | 5.18 × 102 | 3.76 × 103 | ± | 3.71 × 102 | 5.94 × 103 | ± | 2.41 × 103 | 2.75 × 103 | ± | 5.15 × 102 | 1.80 × 103 | ± | 1.67 × 102 |
| 8-HΕΤΕ | 2.23 × 103 | ± | 1.10 × 102 | 2.73 × 103 | ± | 1.02 × 103 | 1.57 × 103 | ± | 4.92 × 102 | 2.19 × 103 | ± | 1.44 × 102 | 3.26 × 103 | ± | 1.34 × 103 | 1.67 × 103 | ± | 2.91 × 102 | 8.63 × 102 | ± | 8.31 × 10 |
| 12-HΕΤΕ | 3.07 × 103 | ± | 2.18 × 102 | 3.67 × 103 | ± | 1.38 × 103 | 2.18 × 103 | ± | 7.48 × 102 | 2.80 × 103 | ± | 1.95 × 102 | 4.42 × 103 | ± | 1.90 × 103 | 2.27 × 103 | ± | 3.95 × 102 | 1.29 × 103 | ± | 7.59 × 10 |
| 15-HΕΤΕ | 4.22 × 103 | ± | 2.09 × 102 | 5.06 × 103 | ± | 1.91 × 103 | 2.95 × 103 | ± | 1.10 × 103 | 3.88 × 103 | ± | 5.55 × 102 | 6.04 × 103 | ± | 1.88 × 103 | 3.33 × 103 | ± | 5.33 × 102 | 2.48 × 103 | ± | 2.33 × 102 |
| 5oxoETE | 9.41 × 102 | ± | 6.88 × 10 | 1.19 × 103 | ± | 4.76 × 102 | 6.73 × 102 | ± | 1.82 × 102 | 9.59 × 102 | ± | 7.00 × 10 | 1.64 × 103 | ± | 6.79 × 102 | 7.73 × 102 | ± | 1.20 × 102 | 5.77 × 102 | ± | 7.91 |
| 8.9-EET | 5.79 × 102 | ± | 3.04 × 10 | 6.05 × 102 | ± | 2.40 × 102 | 3.92 × 102 | ± | 2.96 × 10 | 7.82 × 102 | ± | 2.11 × 102 | 1.22 × 103 | ± | 5.95 × 102 | 4.40 × 102 | ± | 5.47 × 10 | 3.12 × 102 | ± | 2.64 × 10 |
| 11.12-EET | 4.37 × 102 | ± | 3.33 × 10 | 4.52 × 102 | ± | 1.36 × 102 | 3.29 × 102 | ± | 4.50 × 10 | 6.02 × 102 | ± | 1.75 × 102 | 9.73 × 102 | ± | 5.31 × 102 | 3.52 × 102 | ± | 5.29 × 10 | 2.47 × 102 | ± | 2.58 × 10 |
| 14.15-ΕΕΤ | 4.61 × 102 | ± | 5.58 × 10 | 4.98 × 1022 | ± | 1.75 × 102 | 3.24 × 102 | ± | 4.31 × 10 | 6.45 × 102 | ± | 1.52 × 102 | 9.77 × 102 | ± | 5.55 × 102 | 3.57 × 102 | ± | 5.98 × 10 | 2.11 × 102 | ± | 1.29 × 10 |
| TXB2 | 3.86 × 10 | ± | 1.14 × 10 | 4.47 × 10 | ± | 3.08 × 10 | 2.65 × 10 | ± | 6.16 | 2.92 × 10 | ± | 8.55 | 7.86 × 10 | ± | 2.82 × 10 | 2.88 × 10 | ± | 4.45 | 1.31 × 10 | ± | 4.00 × 10−1 |
| PGD2 | 1.87 × 10 | ± | 6.70 | 3.26 × 10 | ± | 1.46 × 10 | 2.16 × 10 | ± | 6.85 | 2.59 × 10 | ± | 1.47 | 7.95 × 10 | ± | 5.34 × 10 | 1.84 × 10 | ± | 4.27 | 2.08 × 10 | ± | 4.49 |
| PGE2 | 1.57 × 102 | ± | 1.07 × 102 | 8.41 × 10 | ± | 6.39 × 10 | 1.05 × 102 | ± | 9.99 × 10 | 1.25 × 102 | ± | 7.83 × 10 | 1.17 × 102 | ± | 8.36 × 10 | 1.78 × 102 | ± | 1.24 × 102 | 4.81 × 10 | ± | 2.48 × 10 |
| PGE3 | 6.59 × 102 | ± | 1.84 × 102 | 8.48 × 102 | ± | 6.07 × 102 | 3.80 × 102 | ± | 5.86 × 10 | 5.75 × 102 | ± | 1.24 × 102 | 1.15 × 103 | ± | 5.73 × 102 | 3.50 × 102 | ± | 5.02 × 10 | 2.10 × 102 | ± | 7.89 |
| 15-d-PGJ2 | 1.27 × 10 | ± | 3.49 × 10−1 | 1.51 × 10 | ± | 1.53 × 10 | 9.64 | ± | 1.82 | 1.93 × 10 | ± | 7.63 | 4.16 × 10 | ± | 2.05 × 10 | 1.13 × 10 | ± | 2.83 | 3.28 | ± | 1.36 |
| 18-HEPE | 4.71 × 104 | ± | 3.23 × 103 | 5.87 × 104 | ± | 2.64 × 104 | 3.41 × 104 | ± | 8.83 × 103 | 4.24 × 104 | ± | 4.61 × 103 | 7.26 × 104 | ± | 2.00 × 104 | 4.70 × 104 | ± | 7.55 × 103 | 3.37 × 104 | ± | 3.39 × 103 |
| RvD5 | 1.15 × 10 | ± | 2.58 | 1.32 × 10 | ± | 5.78 | 1.30 × 10 | ± | 5.23 | 8.23 | ± | 1.19 | 1.37 × 10 | ± | 1.23 × 10 | 1.04 × 10 | ± | 5.50 | 4.23 | ± | 7.07 × 10−2 |
| PDX | 1.24 × 10 | ± | 2.80 | 1.75 × 10 | ± | 6.97 | 1.26 × 10 | ± | 5.36 | 1.30 × 10 | ± | 2.49 | 1.81 × 10 | ± | 9.13 | 1.46 × 10 | ± | 2.60 | 1.05 × 10 | ± | 3.18 |
| 14-HDoHE | 1.02 × 104 | ± | 1.15 × 103 | 1.39 × 104 | ± | 5.14 × 103 | 8.90 × 103 | ± | 2.15 × 103 | 1.13 × 104 | ± | 1.19 × 103 | 1.60 × 104 | ± | 6.11 × 103 | 9.80 × 103 | ± | 2.28 × 103 | 5.52 × 103 | ± | 8.59 × 102 |
| 17-HDoHE | 3.21 × 104 | ± | 4.23 × 103 | 4.09 × 104 | ± | 1.37 × 104 | 2.59 × 104 | ± | 1.09 × 104 | 3.23 × 104 | ± | 4.76 × 103 | 5.07 × 104 | ± | 1.20 × 104 | 3.08 × 104 | ± | 5.16 × 103 | 2.24 × 104 | ± | 3.87 × 103 |
| 5(RS)-5-F2c-lsoP | 111.13 | ± | 3.01 | 115.61 | ± | 4.08 | 90.74 | ± | 20.99 | 90.61 | ± | 2.74 | 129.47 | ± | 15.19 | 100.71 | ± | 33.99 | 133.33 | ± | 21.74 |
| 5(RS)-5-F2t-lsoP | 14.10 | ± | 0.83 | 16.19 | ± | 0.94 | 12.01 | ± | 1.28 | 13.60 | ± | 2.74 | 17.55 | ± | 5.87 | 13.26 | ± | 2.56 | 21.60 | ± | 3.92 |
| 15-A2-IsoP | 57.46 | ± | 13.60 | 65.81 | ± | 14.22 | 49.11 | ± | 16.03 | 43.12 | ± | 0.59 | 68.26 | ± | 11.57 | 58.43 | ± | 18.03 | 56.91 | ± | 13.79 |
| 15-epi-15-F2t-IsoP | 6.70 | ± | 1.37 | 9.11 | ± | 1.65 | 6.63 | ± | 0.74 | 6.74 | ± | 1.88 | 9.29 | ± | 1.08 | 7.97 | ± | 1.82 | 10.89 | ± | 1.69 |
| 15(RS)-15-F2t-lsoP | 3.33 | ± | 0.26 | 4.42 | ± | 1.79 | 3.64 | ± | 0.42 | 3.38 | ± | 0.56 | 5.07 | ± | 1.87 | 3.32 | ± | 0.41 | 5.13 | ± | 0.55 |
| 4(RS)-14-F3t-NeuroP | 18.59 | ± | 2.37 | 23.44 | ± | 4.62 | 19.18 | ± | 3.00 | 17.33 | ± | 1.10 | 22.13 | ± | 5.16 | 19.76 | ± | 4.11 | 33.50 | ± | 4.55 |
| 9-L1t-PhytoP | 3.09 | ± | 0.74 | 6.22 | ± | 3.77 | 1.22 | ± | 0.25 | 4.29 | ± | 2.57 | 2.95 | ± | 0.87 | 2.24 | ± | 0.56 | 3.56 | ± | 0.99 |
| 16-B1t-PhytoP | 3.71 | ± | 0.55 | 6.98 | ± | 4.56 | 1.34 | ± | 0.33 | 4.51 | ± | 2.98 | 3.82 | ± | 1.04 | 2.99 | ± | 0.20 | 4.69 | ± | 1.08 |
| 16-epi-16-F1t-PhytoP | 71.71 | ± | 11.99 | 113.39 | ± | 55.66 | 83.34 | ± | 39.74 | 84.27 | ± | 17.68 | 103.26 | ± | 27.17 | 82.56 | ± | 26.79 | 77.59 | ± | 13.51 |
| 16-F1t-PhytoP | 196.36 | ± | 23.86 | 267.43 | ± | 75.80 | 203.33 | ± | 81.34 | 213.81 | ± | 28.03 | 297.12 | ± | 77.60 | 205.93 | ± | 60.06 | 161.55 | ± | 41.06 |
| ent-16A-9-epi-ST-Δ14-10-PhytoF | 1.60 | ± | 0.37 | 3.17 | ± | 1.55 | 1.02 | ± | 0.18 | 1.52 | ± | 1.08 | 2.09 | ± | 0.39 | 1.49 | ± | 0.18 | 2.00 | ± | 0.05 |
| ent-16B-9-epi-ST-Δ14-10-PhytoF | 2.15 | ± | 0.62 | 4.60 | ± | 2.79 | 1.38 | ± | 0.28 | 2.47 | ± | 0.68 | 2.20 | ± | 0.39 | 1.70 | ± | 0.47 | 3.19 | ± | 0.76 |
| ent-16A-13-epi-ST-Δ14-9-PhytoF | 1.77 | ± | 1.16 | 2.24 | ± | 2.14 | 0.67 | ± | 0.15 | 0.57 | ± | 0.41 | 1.22 | ± | 0.56 | 0.98 | ± | 0.11 | 1.02 | ± | 0.73 |
| 5(R)-5-F3t-lsoP | 247.40 | ± | 45.11 | 283.17 | ± | 43.02 | 209.98 | ± | 31.07 | 204.07 | ± | 37.03 | 319.24 | ± | 64.80 | 279.09 | ± | 64.13 | 495.36 | ± | 9.54 |
| 5(S)-5-F3t-lsoP | 87.74 | ± | 12.50 | 96.90 | ± | 21.94 | 74.47 | ± | 6.85 | 76.78 | ± | 18.69 | 109.24 | ± | 31.62 | 99.86 | ± | 26.60 | 171.49 | ± | 5.36 |
| 8(R)-8-F3t-lsoP | 13.05 | ± | 3.25 | 18.33 | ± | 5.29 | 12.24 | ± | 2.03 | 13.59 | ± | 1.22 | 25.82 | ± | 6.01 | 15.82 | ± | 2.63 | 25.63 | ± | 4.18 |
| 8(S)-8-F3t-lsoP | 12.07 | ± | 3.39 | 11.00 | ± | 3.69 | 8.44 | ± | 1.99 | 8.65 | ± | 2.56 | 12.66 | ± | 2.99 | 10.87 | ± | 2.76 | 11.96 | ± | 2.89 |
| 18(R)-8-F3t-IsoP | 392.01 | ± | 375.37 | 659.01 | ± | 738.83 | 83.54 | ± | 19.69 | 89.87 | ± | 42.52 | 167.32 | ± | 43.50 | 159.82 | ± | 34.50 | 199.22 | ± | 27.56 |
| 18(S)-8-F3t-IsoP | 23.03 | ± | 5.23 | 27.85 | ± | 1.37 | 20.46 | ± | 3.16 | 20.45 | ± | 10.01 | 27.20 | ± | 6.80 | 28.07 | ± | 4.57 | 51.69 | ± | 5.57 |
| 4(RS)-4-F4t-NeuroP | 8.23 | ± | 0.49 | 10.72 | ± | 2.18 | 8.12 | ± | 1.46 | 7.84 | ± | 0.24 | 10.36 | ± | 2.27 | 8.44 | ± | 2.00 | 13.65 | ± | 1.38 |
| 10(R)-10-F4t-NeuroP | 3.98 | ± | 0.46 | 4.39 | ± | 0.88 | 3.05 | ± | 0.82 | 4.26 | ± | 0.97 | 5.40 | ± | 2.59 | 4.03 | ± | 0.90 | 6.16 | ± | 0.47 |
| 10(S)-10-F4t-NeuroP | 3.17 | ± | 0.59 | 4.22 | ± | 2.26 | 3.52 | ± | 0.84 | 2.87 | ± | 0.29 | 4.57 | ± | 1.39 | 4.71 | ± | 0.86 | 5.06 | ± | 0.32 |
| 13A(RS)-13-F4t-NeuroP | 5.22 | ± | 0.95 | 5.81 | ± | 1.54 | 4.53 | ± | 0.66 | 5.19 | ± | 0.88 | 6.34 | ± | 1.87 | 6.36 | ± | 1.75 | 8.39 | ± | 0.91 |
| 13B(RS)-13-F4t-NeuroP | 4.28 | ± | 0.61 | 5.92 | ± | 0.34 | 4.39 | ± | 0.58 | 2.68 | ± | 1.99 | 3.82 | ± | 0.36 | 3.95 | ± | 0.63 | 5.46 | ± | 1.94 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Linares-Maurizi, A.; Awad, R.; Durbec, A.; Reversat, G.; Gros, V.; Galano, J.-M.; Bertrand-Michel, J.; Durand, T.; Pradelles, R.; Oger, C.; et al. Stress-Induced Production of Bioactive Oxylipins in Marine Microalgae. Mar. Drugs 2024, 22, 406. https://doi.org/10.3390/md22090406
Linares-Maurizi A, Awad R, Durbec A, Reversat G, Gros V, Galano J-M, Bertrand-Michel J, Durand T, Pradelles R, Oger C, et al. Stress-Induced Production of Bioactive Oxylipins in Marine Microalgae. Marine Drugs. 2024; 22(9):406. https://doi.org/10.3390/md22090406
Chicago/Turabian StyleLinares-Maurizi, Amandyne, Rana Awad, Anaelle Durbec, Guillaume Reversat, Valérie Gros, Jean-Marie Galano, Justine Bertrand-Michel, Thierry Durand, Rémi Pradelles, Camille Oger, and et al. 2024. "Stress-Induced Production of Bioactive Oxylipins in Marine Microalgae" Marine Drugs 22, no. 9: 406. https://doi.org/10.3390/md22090406
APA StyleLinares-Maurizi, A., Awad, R., Durbec, A., Reversat, G., Gros, V., Galano, J.-M., Bertrand-Michel, J., Durand, T., Pradelles, R., Oger, C., & Vigor, C. (2024). Stress-Induced Production of Bioactive Oxylipins in Marine Microalgae. Marine Drugs, 22(9), 406. https://doi.org/10.3390/md22090406