
Heterologous Expression and Characterization of a pH-Stable Chitinase from Micromonospora aurantiaca with a Potential Application in Chitin Degradation

 , and
, and
Abstract

1. Introduction
2. Results and Discussion
2.1. Sequence Analysis of MaChi1

2.2. Expression and Purification of MaChi1
2.3. Enzymatic Characterization of MaChi1

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism | Chitinase | MW (kDa) | Optimal Temperature (°C) | Optimal pH | pH Stability | Ref |
|---|---|---|---|---|---|---|
| Streptomyces alfalfae | SaHEX | 60 | 60 | 5.5; relative activity was above 80% at 5.0–6.0 | pH range of 4.5–8.5, incubation for 1 h, residual activity was above 80% | [44] |
| Streptomyces sp. F-3 | SsChi18C | – | 60 | 5.0; relative activity was above 80% at 3.0–7.0 | pH range of 3.0–8.0, incubation for 0.5 h, residual activity was above 80% | [45] |
| Microbulbifer sp. BN3 | Chi1602 | 60 | 60 | 9.0; relative activity was above 80% at 4.0–9.0 | pH range of 5.0–9.0, incubation for 1 h, residual activity was above 80% | [46] |
| Paenibacillus sp. | A1 | 30 | 50 | 4.5; relative activity was above 90% at 4.0–5.0 | pH range of 4.5–5.5, incubation for 1 h, residual activity was above 80% | [47] |
| Thermophilic sp. | Chi304 | 70.95 | 80 | 9.0; relative activity was above 80% at 8.0–10.0 | pH range of 6.0–10.0, incubation for 1 h, residual activity was above 60% | [48] |
| Trichoderma gamsii R1 | ChiTg | 42 | 40 | 5.0; relative activity was above 70% at 4.0–6.0 | pH range of 5.0–8.0, incubation for 0.5 h, residual activity was above 80% | [2] |
| Paenibacillus sp. | Y412MC10 | 52 | 60 | 5.5; relative activity was above 70% at 4.5–6.5 | pH range of 4.5–6.5, incubation for 1 h, residual activity was above 90% | [49] |
| Chitinilyticum sp. C8 | ChiC8–1 | 100 | 50 | 6.0; relative activity was above 70% at 4.0–9.0 | pH range of 5.0–8.0, incubation for 1 h, residual activity was above 90% | [50] |
| Aeromonas media CZW001 | AmChi | 40 | 55 | 8.0; relative activity was above 75% at 6.0–9.0 | pH range of 4.0–9.0, incubation for 2 h, residual activity was above 70% | [51] |
| Trichoderma harzianum GIM 3.442 | Chit46 | 46 | 45 | 6.0; relative activity was above 80% at 6.0–7.0 | pH range of 5.0–9.0, incubation for 1 h, residual activity was above 80% | [52] |
| M. aurantiaca | MaChi1 | 57 | 55 | 5.0; relative activity was above 82% at 4.0–9.0 | pH range of 3.0–10.0, incubation for 1 h, residual activity was above 88.6% | This study |
2.4. Substrate Spectrum and Kinetic Parameters of MaChi1
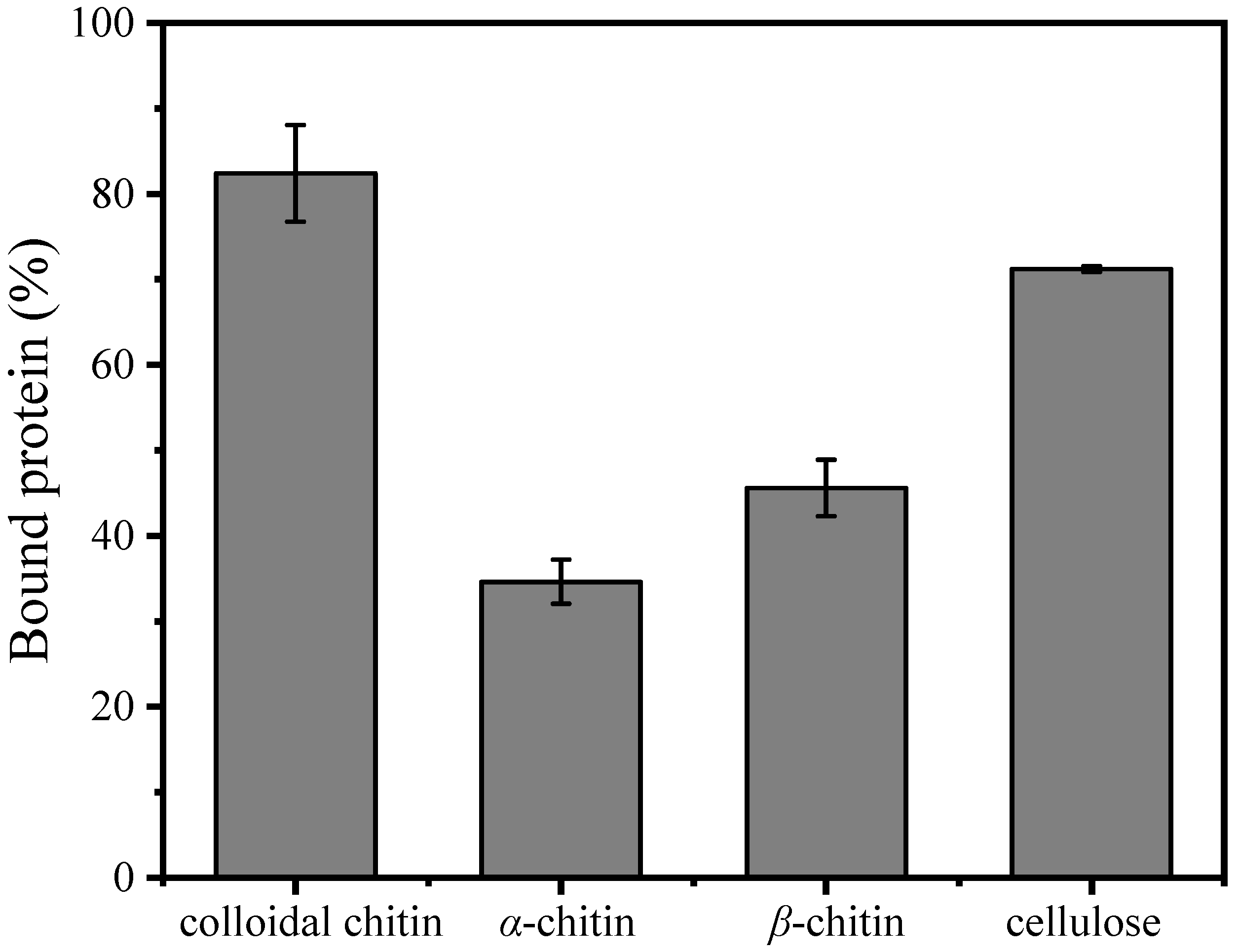
2.5. Affinity of MaChi1 for Polysaccharides
2.6. Hydrolytic Property of MaChi1
2.7. Hydrolysis of Colloidal Chitin
3. Materials and Methods
3.1. Materials
3.2. Strains and Culture Conditions
3.3. Cloning, Expression, and Bioinformatic Analysis of the MaChi1 Gene
3.4. Purification of Recombinant MaChi1
3.5. Chitinase Activity Assay
3.6. Biochemical Characteristics of MaChi1
3.7. Substrate Specificity and Enzyme Kinetics
3.8. Substrate-Binding Capacity of MaChi1
3.9. Degradation Mode of MaChi1
3.10. Enzymatic Hydrolysis of Colloidal Chitin
3.11. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Amiri, H.; Aghbashlo, M.; Sharma, M.; Gaffey, J.; Manning, L.; Moosavi Basri, S.M.; Kennedy, J.F.; Gupta, V.K.; Tabatabaei, M. Chitin and chitosan derived from crustacean waste valorization streams can support food systems and the UN sustainable development goals. Nat. Food. 2022, 3, 822–828. [Google Scholar] [CrossRef]
- Wang, J.; Zhu, M.; Wang, P.; Chen, W. Biochemical properties of a Cold-Active chitinase from marine Trichoderma gamsii R1 and its application to preparation of chitin oligosaccharides. Mar. Drugs 2023, 21, 332. [Google Scholar] [CrossRef] [PubMed]
- Minguet-Lobato, M.; Cervantes, F.V.; Miguez, N.; Plou, F.J.; Fernandez-Lobato, M. Chitinous material bioconversion by three new chitinases from the yeast Mestchnikowia pulcherrima. Microb. Cell. Fact. 2024, 23, 31. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Dang, Y.; Liu, S.; Huang, K.; Qin, Q.; Chen, X.; Zhang, Y.; Wang, Y.; Li, P. Identification and characterization of three chitinases with potential in direct conversion of crystalline chitin into N,N′-diacetylchitobiose. Mar. Drugs 2022, 20, 165. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Yang, L.; Yang, D.; Jiang, M.; Ling, C.; Chen, H.; Ji, F.; Pan, L. Biochemical purification and characterization of a truncated acidic, thermostable chitinase from marine fungus for N-acetylglucosamine production. Front. Bioeng. Biotechnol. 2022, 10, 1013313. [Google Scholar] [CrossRef]
- Zhang, X.; Yuan, J.; Li, F.; Xiang, J. Chitin synthesis and degradation in crustaceans: A genomic view and application. Mar. Drugs 2021, 19, 153. [Google Scholar] [CrossRef]
- Husson, E.; Hadad, C.; Huet, G.; Laclef, S.; Lesur, D.; Lambertyn, V.; Jamali, A.; Gottis, S.; Sarazin, C.; Nguyen Van Nhien, A. The effect of room temperature ionic liquids on the selective biocatalytic hydrolysis of chitin via sequential or simultaneous strategies. Green Chem. 2017, 19, 4122–4131. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, K.; Li, L.; Song, X.; He, Y.; Ding, N.; Li, L.; Wang, S.; Liu, Z. A review of the immune activity of chitooligosaccharides. Food Sci. Technol. 2023, 43, e97822. [Google Scholar] [CrossRef]
- Kumar, M.; Madhuprakash, J.; Balan, V.; Kumar Singh, A.; Vivekanand, V.; Pareek, N. Chemoenzymatic production of chitooligosaccharides employing ionic liquids and Thermomyces lanuginosus chitinase. Bioresour. Technol. 2021, 337, 125399. [Google Scholar] [CrossRef]
- Ji, X.; Zhu, L.; Chang, K.; Zhang, R.; Chen, Y.; Yin, H.; Jin, J.; Zhao, L. Chitooligosaccahrides: Digestion characterization and effect of the degree of polymerization on gut microorganisms to manage the metabolome functional diversity in vitro. Carbohydr. Polym. 2022, 275, 118716. [Google Scholar] [CrossRef]
- Qin, X.; Xin, Y.Z.; Su, X.Y.; Wang, X.L.; Zhang, J.; Tu, T.; Wang, Y.R.; Yao, B.; Huang, H.Q.; Luo, H.Y. Heterologous expression and characterization of thermostable chitinase and β-N-acetylhexosaminidase from Caldicellulosiruptor acetigenus and their synergistic action on the bioconversion of chitin into N-acetyl-D-glucosamine. Int. J. Biol. Macromol. 2021, 192, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Behera, P.K.; Madhuprakash, J. Efficient conversion of crystalline chitin to N-acetylglucosamine and N,N’-diacetylchitobiose by the enzyme cocktail produced by Paenibacillus sp. LS1. Carbohydr. Polym. 2020, 250, 116889. [Google Scholar] [CrossRef]
- Okoro, O.V.; Nie, L.; Gunduz, O.; Ulag, S.; Hamidi, M.; Shavandi, A. Technoeconomic assessment of biopolymer production from crustacean waste with the UK as a case study. Sustainability 2023, 15, 2280. [Google Scholar] [CrossRef]
- Oyeleye, A.; Normi, Y.M. Chitinase: Diversity, limitations, and trends in engineering for suitable applications. Biosci. Rep. 2018, 38, BSR2018032300. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Yu, M.; Wu, Y.; Ran, L.; Liu, W.; Zhang, X. Two highly similar chitinases from marine Vibrio species have different enzymatic properties. Mar. Drugs 2020, 18, 139. [Google Scholar] [CrossRef] [PubMed]
- Lombard, V.; Ramulu, H.G.; Drula, E.; Coutinho, P.M.; Henrissat, B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic. Acids. Res. 2014, 42, D490–D495. [Google Scholar] [CrossRef] [PubMed]
- Taokaew, S.; Kriangkrai, W. Chitinase-assisted bioconversion of chitinous waste for development of value-added chito-oligosaccharides products. Biology 2023, 12, 87. [Google Scholar] [CrossRef] [PubMed]
- Itoh, T.; Hibi, T.; Fujii, Y.; Sugimoto, I.; Fujiwara, A.; Suzuki, F.; Iwasaki, Y.; Kim, J.K.; Taketo, A.; Kimoto, H. Cooperative degradation of chitin by extracellular and cell surface-expressed chitinases from Paenibacillus sp. strain FPU-7. Appl. Environ. Microbiol. 2013, 79, 7482–7490. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.Y.; Li, J.H.; Lv, X.Q.; Du, G.C.; Liu, L. Molecular engineering of chitinase from Bacillus sp. DAU101 for enzymatic production of chitooligosaccharides. Enzyme. Microb. Technol. 2019, 124, 54–62. [Google Scholar] [CrossRef]
- Li, Z.K.; Xia, C.Y.; Wang, Y.X.; Li, X.; Qiao, Y.; Li, C.Y.; Zhou, J.; Zhang, L.; Ye, X.F.; Huang, Y.; et al. Identification of an endo-chitinase from Corallococcus sp. EGB and evaluation of its antifungal properties. Int. J. Biol. Macromol. 2019, 132, 1235–1243. [Google Scholar] [CrossRef]
- Hirsch, A.M.; Valdés, M. Micromonospora: An important microbe for biomedicine and potentially for biocontrol and biofuels. Soil Biol. Biochem. 2010, 42, 536–542. [Google Scholar] [CrossRef]
- Gasmi, M.; Kitouni, M.; Carro, L.; Pujic, P.; Normand, P.; Boubakri, H. Chitinolytic actinobacteria isolated from an Algerian semi-arid soil: Development of an antifungal chitinase-dependent assay and GH18 chitinase gene identification. Ann. Microbiol. 2019, 69, 395–405. [Google Scholar] [CrossRef]
- Teregulova, G.A.; Sineva, O.N.; Markelova, N.N.; Sadikova, V.S.; Uvarov, G.V.; Kovalenko, M.A.; Manucharova, N.A. Evaluation of chitinolytic and antibiotic activity of Streptomyces avidinii INA 01467 and Micromonospora aurantiaca INA 01468. Eurasian Soil Sci. 2023, 56, 611–618. [Google Scholar] [CrossRef]
- Mane, U.V.; Deshmukh, A.M. Chitin degrading potential of three aquatic actinomycetes and its optimization. Afr. J. Biotechnol. 2009, 8, 6617–6620. [Google Scholar]
- Harvinda, Y.; Ustadi, U.; Putra, M.M.P. Production, purification and characterization of chitinase from Micromonospora sp. AR17. Indones. J. Biotechnol. 2023, 28, 46–55. [Google Scholar] [CrossRef]
- Forsberg, Z.; Bissaro, B.; Gullesen, J.; Dalhus, B.; Vaaje-Kolstad, G.; Eijsink, V. Structural determinants of bacterial lytic polysaccharide monooxygenase functionality. J. Biol. Chem. 2018, 293, 1397–1412. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Chiba, D.; Yoshida, S.; Takahashi, M.; Totani, K.; Shida, Y.; Ogasawara, W.; Nakagawa, Y.S. Functional analysis of a novel lytic polysaccharide monooxygenase from Streptomyces griseus on cellulose and chitin. Int. J. Biol. Macromol. 2020, 164, 2085–2091. [Google Scholar] [CrossRef]
- Li, J.; Goddard-Borger, E.D.; Raji, O.; Saxena, H.; Solhi, L.; Mathieu, Y.; Master, E.R.; Wakarchuk, W.W.; Brumer, H. Chitin-active lytic polysaccharide monooxygenases are rare in Cellulomonas species. Appl. Environ. Microbiol. 2022, 88, e00922–e00968. [Google Scholar] [CrossRef]
- Honda, S.; Kunii, T.; Nohara, K.; Wakita, S.; Sugahara, Y.; Kawakita, M.; Oyama, F.; Sakaguchi, M. Characterization of a Bacillus thuringiensis chitinase that binds to cellulose and chitin. AMB Express. 2017, 7, 51. [Google Scholar] [CrossRef]
- Nakamura, T.; Mine, S.; Hagihara, Y.; Ishikawa, K.; Ikegami, T.; Uegaki, K. Tertiary structure and carbohydrate recognition by the chitin-binding domain of a hyperthermophilic chitinase from Pyrococcus furiosus. J. Mol. Biol. 2008, 381, 670–680. [Google Scholar] [CrossRef]
- Xu, G.Y.; Ong, E.; Gilkes, N.R.; Kilburn, D.G.; Muhandiram, D.R.; Harris-Brandts, M.; Carver, J.P.; Kay, L.E.; Harvey, T.S. Solution structure of a cellulose-binding domain from Cellulomonas fimi by nuclear magnetic resonance spectroscopy. Biochemistry 1995, 34, 6993–7009. [Google Scholar] [CrossRef] [PubMed]
- Tunyasuvunakool, K.; Adler, J.; Wu, Z.; Green, T.; Zielinski, M.; Zídek, A.; Bridgland, A.; Cowie, A.; Meyer, C.; Laydon, A.; et al. Highly accurate protein structure prediction for the human proteome. Nature 2021, 596, 590. [Google Scholar] [CrossRef] [PubMed]
- David, A.; Islam, S.; Tankhilevich, E.; Sternberg, M. The Alphafold database of protein structures: A biologist’s guide. J. Mol. Biol. 2022, 434, 167336. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.J.; Shao, S.J.; Li, L.L.; Cheng, Z.; Tian, L.; Gao, P.J.; Wang, L.S. Substrate-binding specificity of chitinase and chitosanase as revealed by active-site architecture analysis. Carbohydr. Res. 2015, 418, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Xie, Y.; Ma, J.; Luo, X.; Nie, P.; Zuo, Z.; Lahrmann, U.; Zhao, Q.; Zheng, Y.; Zhao, Y.; et al. IBS: An illustrator for the presentation and visualization of biological sequences. Bioinformatics 2015, 31, 3359–3361. [Google Scholar] [CrossRef] [PubMed]
- Nawani, N.N.; Kapadnis, B.P.; Das, A.D.; Rao, A.S.; Mahajan, S.K. Purification and characterization of a thermophilic and acidophilic chitinase from Microbispora sp. V2. J. Appl. Microbiol. 2002, 93, 965–975. [Google Scholar] [CrossRef]
- Zhang, W.J.; Liu, Y.H.; Ma, J.W.; Yan, Q.J.; Jiang, Z.Q.; Yang, S.Q. Biochemical characterization of a bifunctional chitinase/lysozyme from Streptomyces sampsonii suitable for N-acetyl chitobiose production. Biotechnol. Lett. 2020, 42, 1489–1499. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Gao, K.; Secundo, F.; Mao, X. Biochemical characterization of two β-N-acetylglucosaminidases from Streptomyces violascens for efficient production of N-acetyl-D-glucosamine. Food Chem. 2021, 364, 130393. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Li, A.; Han, H.; Liu, T.; Yang, Q. A potent chitinase from Bacillus subtilis for the efficient bioconversion of chitin-containing wastes. Int. J. Biol. Macromol. 2018, 116, 863–868. [Google Scholar] [CrossRef]
- Gao, L.; Sun, J.; Secundo, F.; Gao, X.; Mao, X. Cloning, characterization and substrate degradation mode of a novel chitinase from Streptomyces albolongus ATCC 27414. Food Chem. 2018, 261, 329–336. [Google Scholar] [CrossRef]
- Lu, Y.; Wang, N.; He, J.; Li, Y.; Gao, X.; Huang, L.; Yan, X. Expression and characterization of a novel chitinase with antifungal activity from a rare actinomycete, Saccharothrix yanglingensis Hhs.015. Protein. Expr. Purif. 2018, 143, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Lv, C.; Gu, T.; Ma, R.; Yao, W.; Huang, Y.; Gu, J.; Zhao, G. Biochemical characterization of a GH19 chitinase from Streptomyces alfalfae and its applications in crystalline chitin conversion and biocontrol. Int. J. Biol. Macromol. 2021, 167, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Brzezinska, M.S.; Jankiewicz, U.; Kalwasinska, A.; Swiatczak, J.; Zero, K. Characterization of chitinase from Streptomyces luridiscabiei U05 and its antagonist potential against fungal plant pathogens. J. Phytopathol. 2019, 167, 404–412. [Google Scholar] [CrossRef]
- Lv, C.; Gu, T.; Xu, K.; Gu, J.; Li, L.; Liu, X.; Zhang, A.; Gao, S.; Li, W.; Zhao, G. Biochemical characterization of a β-N-acetylhexosaminidase from Streptomyces alfalfae and its application in the production of N-acetyl-D-glucosamine. J. Biosci. Bioeng. 2019, 128, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.M.; Li, Y.J.; Tian, Z.N.; Qian, Y.C.; Zhang, H.Q.; Wang, L.S. A novel thermostable chitinolytic machinery of Streptomyces sp. F-3 consisting of chitinases with different action modes. Biotechnol. Biofuels. 2019, 12, 136. [Google Scholar] [CrossRef] [PubMed]
- Li, R.K.; Hu, Y.J.; He, Y.J.; Ng, T.B.; Zhou, Z.M.; Ye, X.Y. A thermophilic chitinase 1602 from the marine bacterium Microbulbifer sp. BN3 and its high-level expression in Pichia pastoris. Biotechnol. Appl. Biochem. 2021, 68, 1076–1085. [Google Scholar] [CrossRef] [PubMed]
- Du, J.H.; Duan, S.; Miao, J.Y.; Zhai, M.M.; Cao, Y. Purification and characterization of chitinase from Paenibacillus sp. Biotechnol. Appl. Biochem. 2021, 68, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Guan, F.; Xu, G.; Liu, X.; Zhang, Y.; Sun, J.; Yao, B.; Huang, H.; Wu, N.; Tian, J. A novel Thermophilic chitinase directly mined from the marine metagenome using the deep learning tool Preoptem. Bioresour. Bioprocess. 2022, 9, 54. [Google Scholar] [CrossRef]
- Arooj, B.; Mutahir, Z.; Ali, M.; Akhter, M.; Mahmood, M.S.; Hamid, A.; Saleem, M. A thermally stable acidic chitinase from Paenibacillus sp. Y412MC10: Molecular characterization and its structural modeling. Pak. J. Zool. 2023, 55, 2527–2538. [Google Scholar]
- Zhao, Q.; Fan, L.; Deng, C.; Ma, C.; Zhang, C.; Zhao, L. Bioconversion of chitin into chitin oligosaccharides using a novel chitinase with high chitin-binding capacity. Int. J. Biol. Macromol. 2023, 244, 125241. [Google Scholar] [CrossRef]
- Ding, Z.W.; Li, T.; Chen, M.; Fang, Y.W.; Hou, X.Y.; Yang, G.; Lu, J.; Ye, Q.W.; Zhu, R.J.; He, F.X.; et al. Purification and characterization of a chitinase from Aeromonas media CZW001 as a biocatalyst for producing chitinpentaose and chitinhexaose. Biotechnol. Appl. Biochem. 2023, 70, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.J.; Shi, D.; Mao, H.H.; Li, Z.W.; Liang, S.; Ke, Y.; Luo, X.C. Heterologous expression and characterization of an antifungal chitinase (Chit46) from Trichoderma harzianum GIM 3.442 and its application in colloidal chitin conversion. Int. J. Biol. Macromol. 2019, 134, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Wu, X.L.; Guo, X.X.; Tang, J.; Zong, M.H.; Lou, W.Y. Double-chitinase hydrolysis of crab shell chitin pretreated by ionic liquid to generate chito-oligosaccharide. ACS Sustain. Chem. Eng. 2019, 7, 1683–1691. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, S.; Yang, D.; Yang, L.; Wang, Q.; Yu, J.; Li, N.; Pan, L. The discovery, enzymatic characterization and functional analysis of a newly isolated chitinase from marine-derived fungus Aspergillus fumigatus df347. Mar. Drugs 2022, 20, 520. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Guo, Y.; Jin, Y.; Ma, M. Bioconversion of chitin waste using a cold-adapted chitinase to produce chitin oligosaccharides. LWT Food Sci. Technol. 2020, 133, 109863. [Google Scholar] [CrossRef]
- Dai, D.H.; Hu, W.L.; Huang, G.R.; Li, W. Purification and characterization of a novel extracellular chitinase from Thermophilic bacillus sp. Hu1. Afr. J. Biotechnol. 2011, 10, 2476–2485. [Google Scholar]
- Yahiaoui, M.; Laribi-Habchi, H.; Bouacem, K.; Asmani, K.L.; Jaouadi, B. Purification and biochemical characterization of a new organic solvent-tolerant chitinase from Paenibacillus timonensis strain LK-DZ15 isolated from the Djurdjura Mountains in Kabylia, Algeria. Carbohydr. Res. 2019, 483, 107747. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Fu, X.; Yan, Q.; Guo, Y.; Liu, Z.; Jiang, Z. Cloning, expression, purification and application of a novel chitinase from a thermophilic marine bacterium Paenibacillus barengoltzii. Food Chem. 2016, 192, 1041–1048. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.H.; Fu, X.; Yan, X.Y.; Peng, W.F.; Kang, L.X. A broad-specificity chitinase from Penicillium oxalicum k10 exhibits antifungal activity and biodegradation properties of chitin. Mar. Drugs. 2021, 19, 356. [Google Scholar] [CrossRef]
- Zhang, A.; Gao, C.; Wang, J.; Chen, K.; Ouyang, P. An efficient enzymatic production of N-acetyl-D-glucosamine from crude chitin powders. Green Chem. 2016, 18, 2147–2154. [Google Scholar] [CrossRef]
- Fu, X.; Yan, Q.; Wang, J.; Yang, S.; Jiang, Z. Purification and biochemical characterization of novel acidic chitinase from Paenicibacillus barengoltzii. Int. J. Biol. Macromol. 2016, 91, 973–979. [Google Scholar] [CrossRef] [PubMed]
- Vaikuntapu, P.R.; Mallakuntla, M.K.; Das, S.N.; Bhuvanachandra, B.; Ramakrishna, B.; Nadendla, S.R.; Podile, A.R. Applicability of endochitinase of Flavobacterium johnsoniae with transglycosylation activity in generating long-chain chitooligosaccharides. Int. J. Biol. Macromol. 2018, 117, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Joe, S.; Sarojini, S. An efficient method of production of colloidal chitin for enumeration of chitinase producing bacteria. Mapana J. Sci. 2017, 4, 37–45. [Google Scholar] [CrossRef]
- Yang, Y.; Li, J.; Liu, X.; Pan, X.; Hou, J.; Ran, C.; Zhou, Z. Improving extracellular production of Serratia marcescens lytic polysaccharide monooxygenase CBP21 and Aeromonas veronii B565 chitinase Chi92 in Escherichia coli and their synergism. AMB Express 2017, 7, 170. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with alphafold. Nature 2021, 596, 583. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. Autodock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- García-Fraga, B.; Da-Silva, A.F.; López-Seijas, J.; Sieiro, C. A novel family 19 chitinase from the marine-derived Pseudoalteromonas tunicata CCUG 44952T: Heterologous expression, characterization and antifungal activity. Biochem. Eng. J. 2015, 93, 84–93. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Xu, P.; Zong, M.; Lou, W. Purification and characterization of alkaline chitinase from Paenibacillus pasadenensis CS0611. Chin. J. Catal. 2017, 38, 665–672. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, X.; He, Y.; Li, Y. The synergistic action of two chitinases from Vibrio harveyi on chitin degradation. Carbohydr. Polym. 2023, 307, 120640. [Google Scholar] [CrossRef]




| Substrate | Specific Activity (U/mg) |
|---|---|
| α-Chitin | 0.50 ± 0.01 |
| β-Chitin | 1.44 ± 0.02 |
| Colloidal chitin | 1.78 ± 0.03 |
| Shrimp shell powder | 0.69 ± 0.00 |
| (GlcNAc)2 | ND a |
| (GlcNAc)3 | 1.15 ± 0.03 |
| (GlcNAc)4 | 1.28 ± 0.02 |
| (GlcNAc)5 | 1.85 ± 0.01 |
| (GlcNAc)6 | 3.11 ± 0.05 |
| Chitosan (DD 95%) | 0.19 ± 0.01 |
| Chitosan (DD 90%) | 0.12 ± 0.01 |
| Chitosan (DD 85%) | 0.25 ± 0.01 |
| Chitosan (DD 80%) | 0.39 ± 0.01 |
| Chitosan (DD 70%) | 0.69 ± 0.01 |
| Chitosan (DD 55%) | 1.38 ± 0.02 |
| Cellulose | 0.34 ± 0.02 |
| Soluble starch | 0.34 ± 0.00 |
| Substrate | Vmax (μmol·min−1·mg−1) | Km (mg/mL) | Kcat (s−1) | Kcat/Km (mg−1·mL·s−1) |
|---|---|---|---|---|
| Colloidal chitin | 2.33 | 4.60 | 0.43 | 0.09 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, H.-Z.; Wang, D.; Yang, H.-T.; Wu, Y.-L.; Li, Y.-C.; Xia, G.-H.; Zhang, X.-Y. Heterologous Expression and Characterization of a pH-Stable Chitinase from Micromonospora aurantiaca with a Potential Application in Chitin Degradation. Mar. Drugs 2024, 22, 287. https://doi.org/10.3390/md22060287
Guo H-Z, Wang D, Yang H-T, Wu Y-L, Li Y-C, Xia G-H, Zhang X-Y. Heterologous Expression and Characterization of a pH-Stable Chitinase from Micromonospora aurantiaca with a Potential Application in Chitin Degradation. Marine Drugs. 2024; 22(6):287. https://doi.org/10.3390/md22060287
Chicago/Turabian StyleGuo, Han-Zhong, Dou Wang, Hui-Ting Yang, Yu-Le Wu, Yong-Cheng Li, Guang-Hua Xia, and Xue-Ying Zhang. 2024. "Heterologous Expression and Characterization of a pH-Stable Chitinase from Micromonospora aurantiaca with a Potential Application in Chitin Degradation" Marine Drugs 22, no. 6: 287. https://doi.org/10.3390/md22060287
APA StyleGuo, H.-Z., Wang, D., Yang, H.-T., Wu, Y.-L., Li, Y.-C., Xia, G.-H., & Zhang, X.-Y. (2024). Heterologous Expression and Characterization of a pH-Stable Chitinase from Micromonospora aurantiaca with a Potential Application in Chitin Degradation. Marine Drugs, 22(6), 287. https://doi.org/10.3390/md22060287