


Identification of Potential Factors for the Promotion of Fucoxanthin Synthesis by Methyl Jasmonic Acid Treatment of Phaeodactylum tricornutum
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effect of MeJA on Cell Growth and Fucoxanthin Content of P. tricornutum
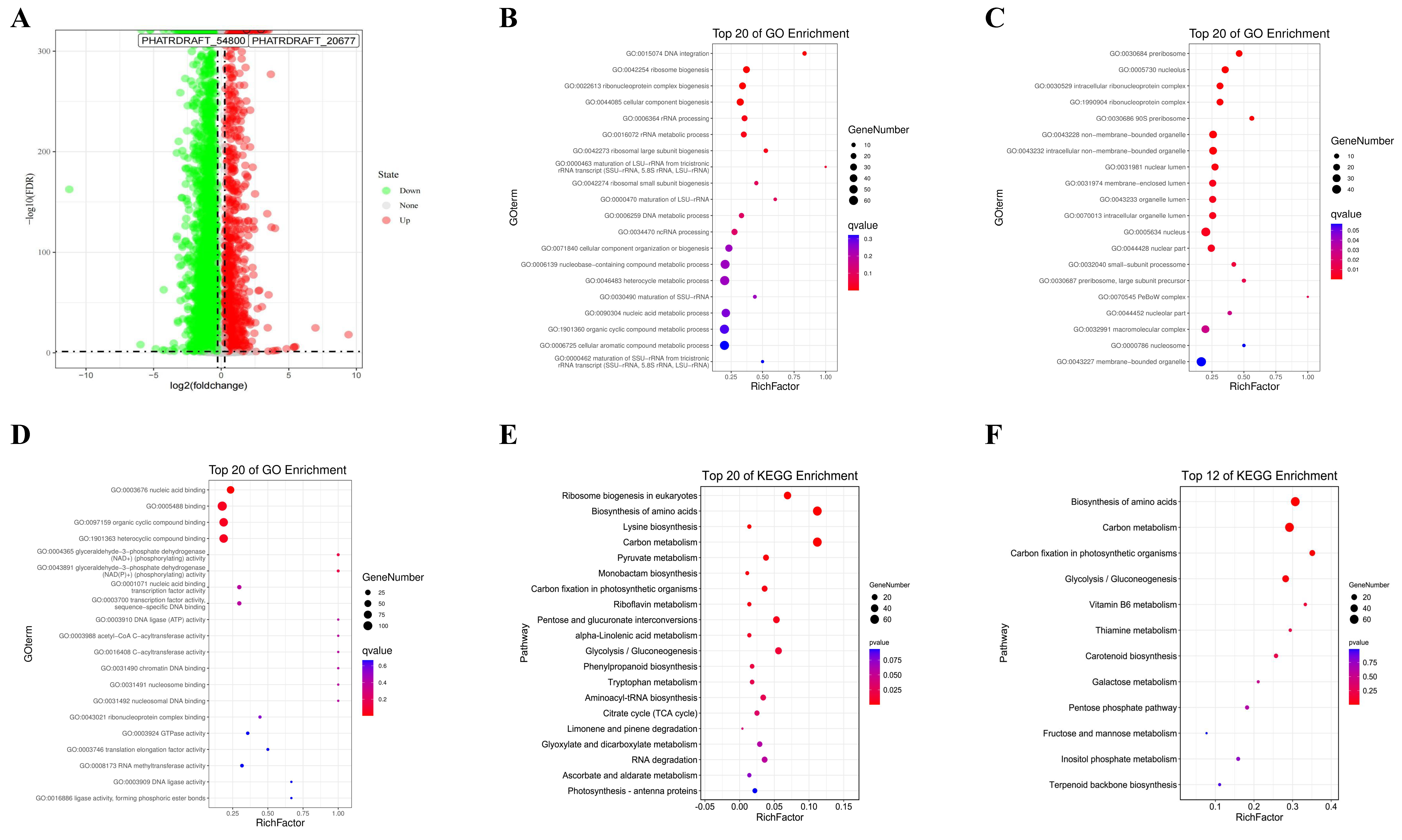
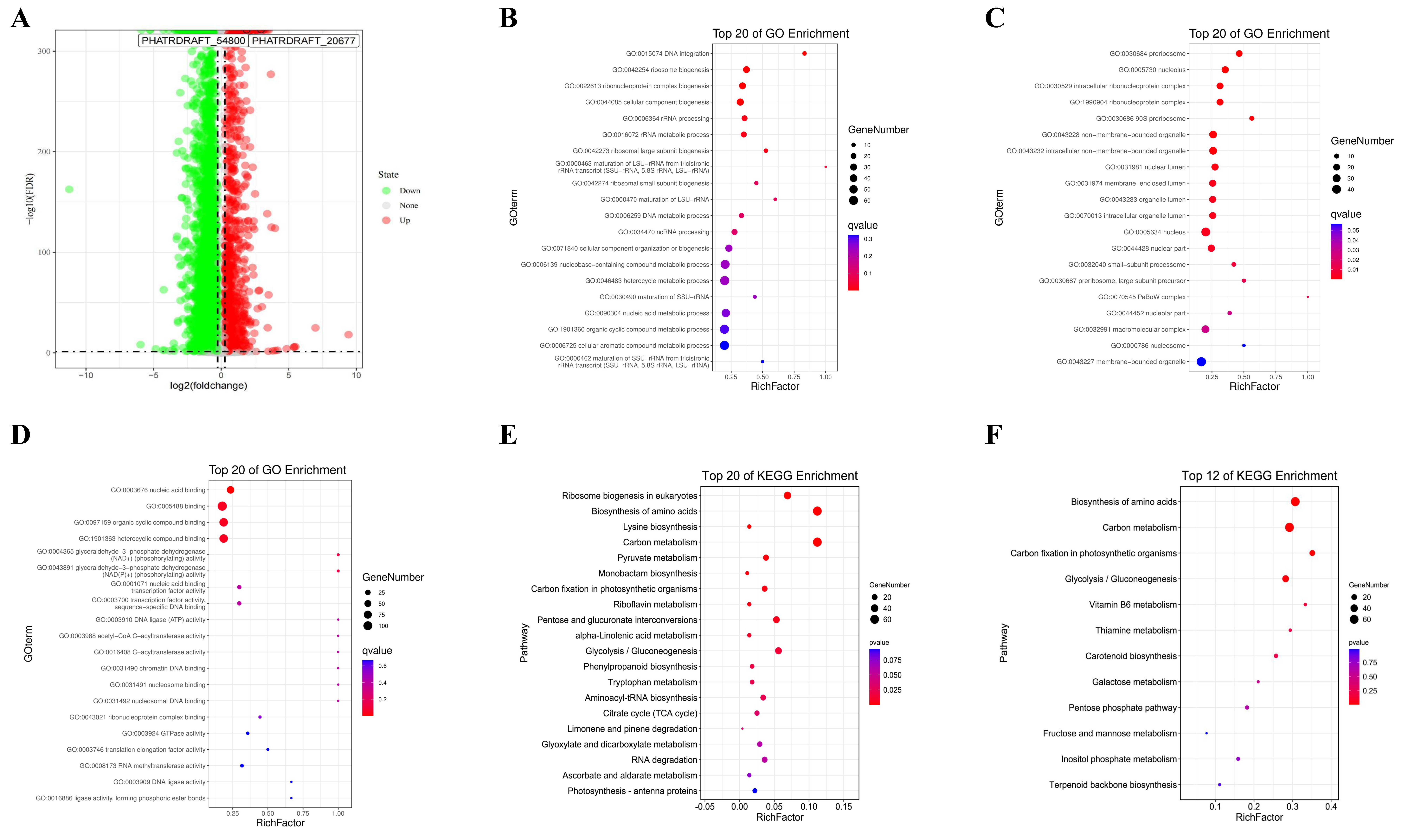
2.2. Differential Gene Expression Analysis
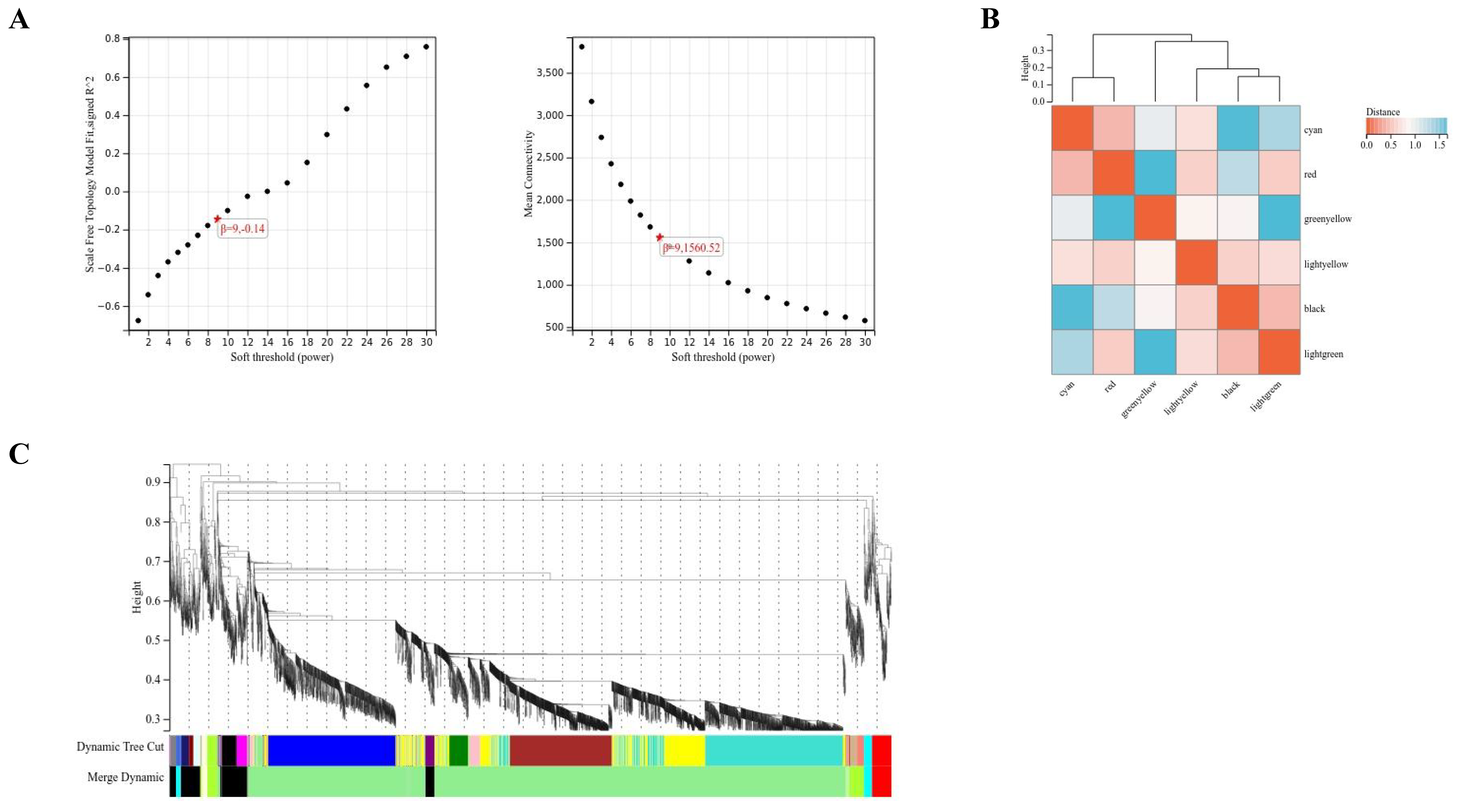
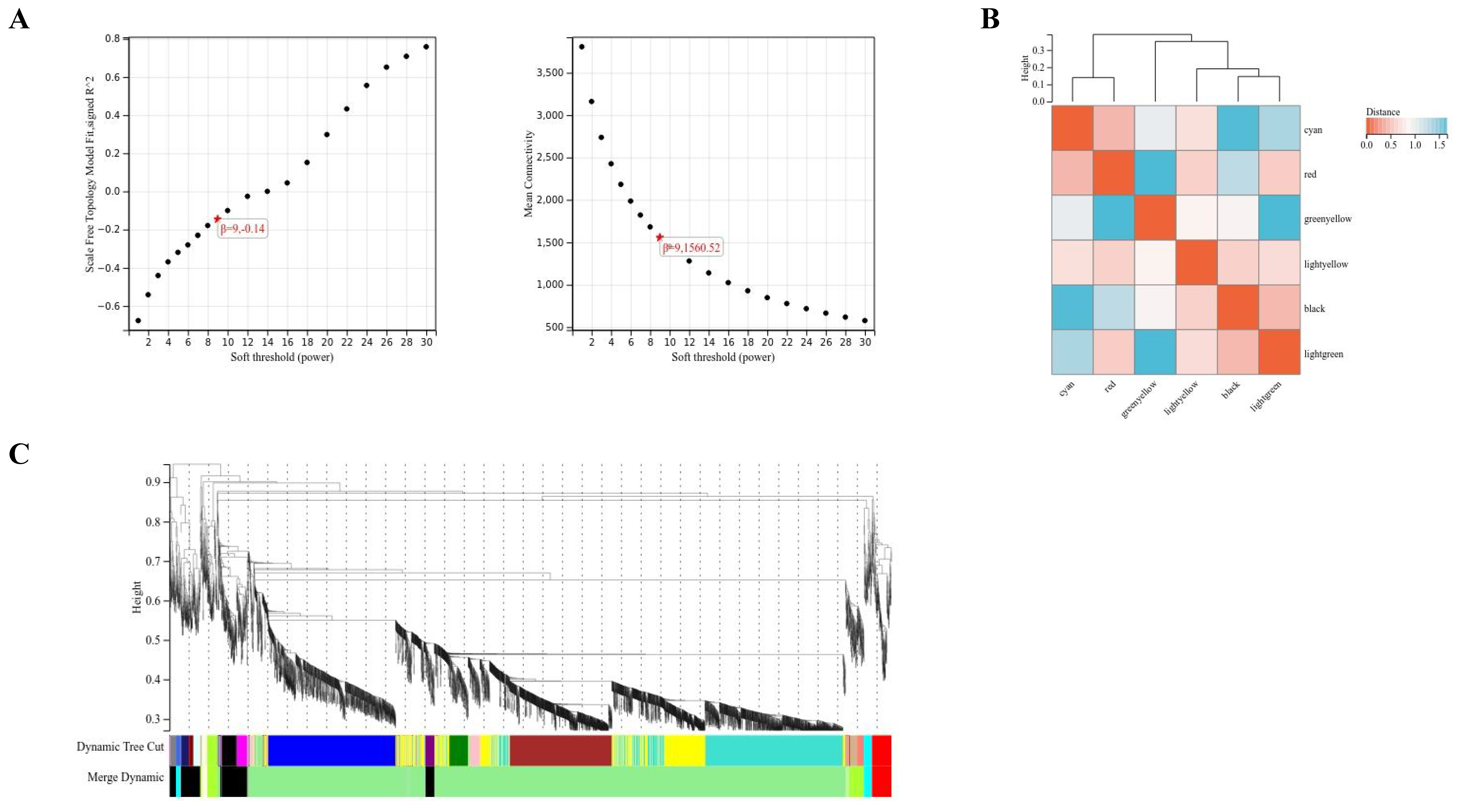
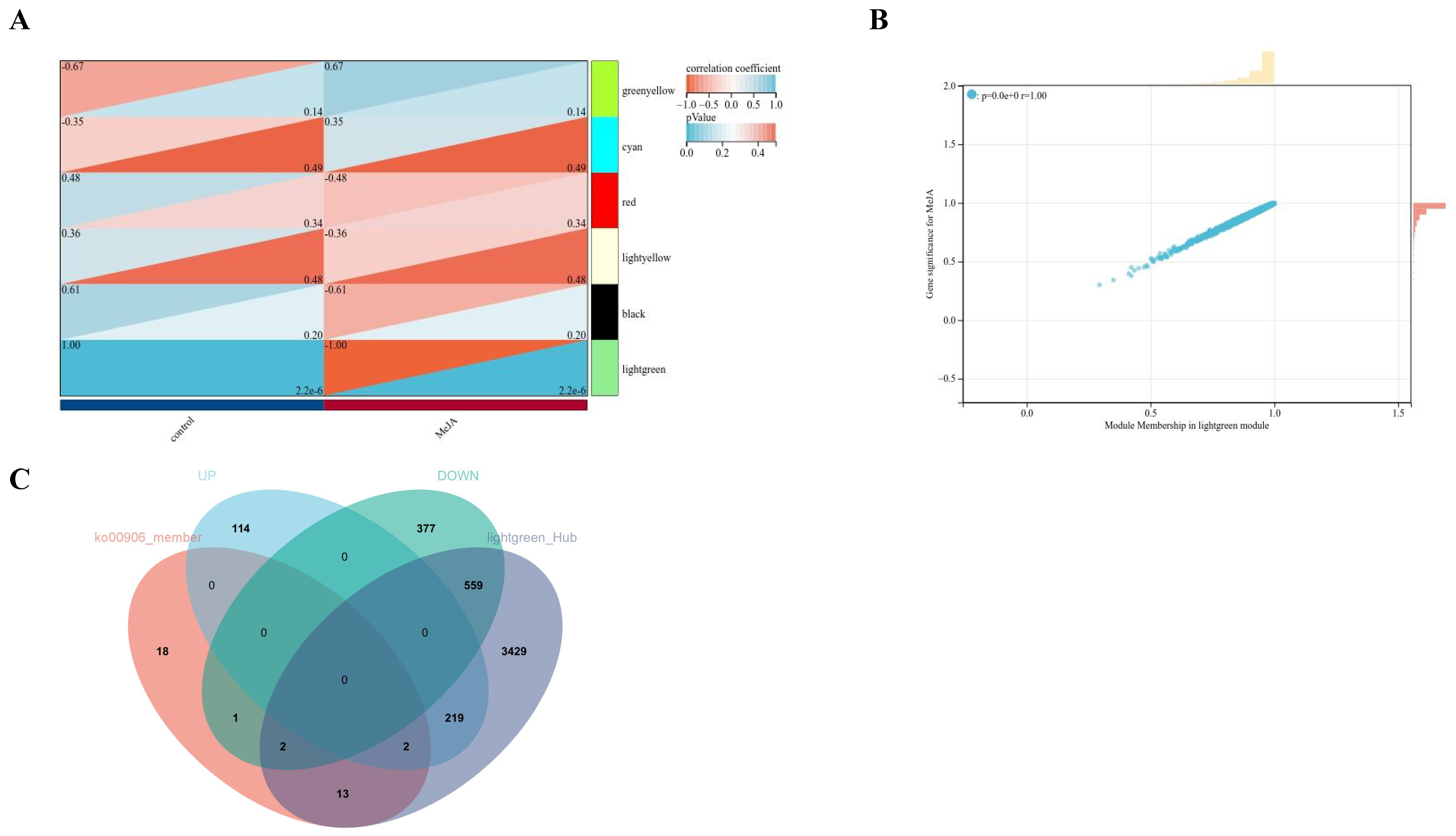
2.3. WGCNA
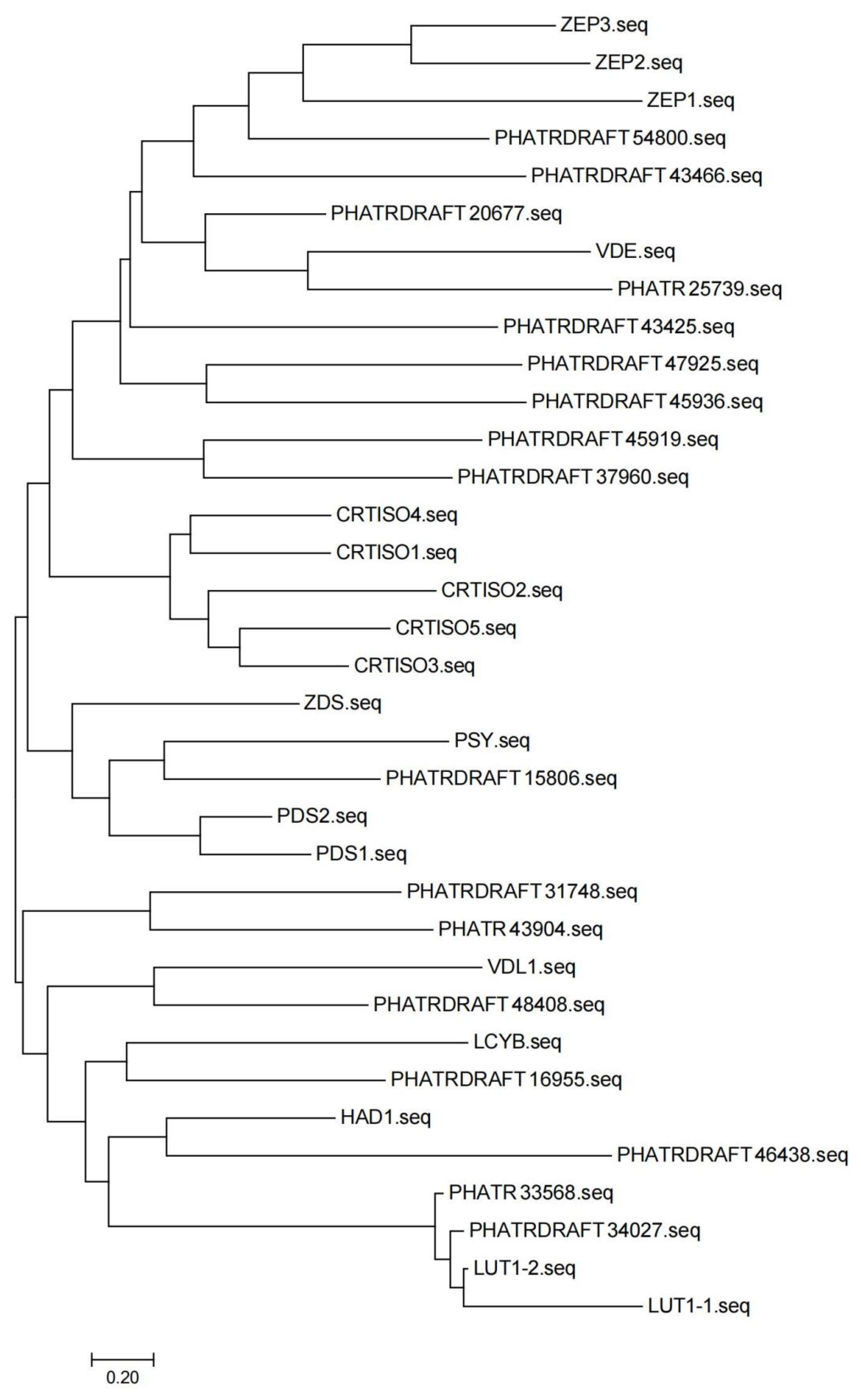
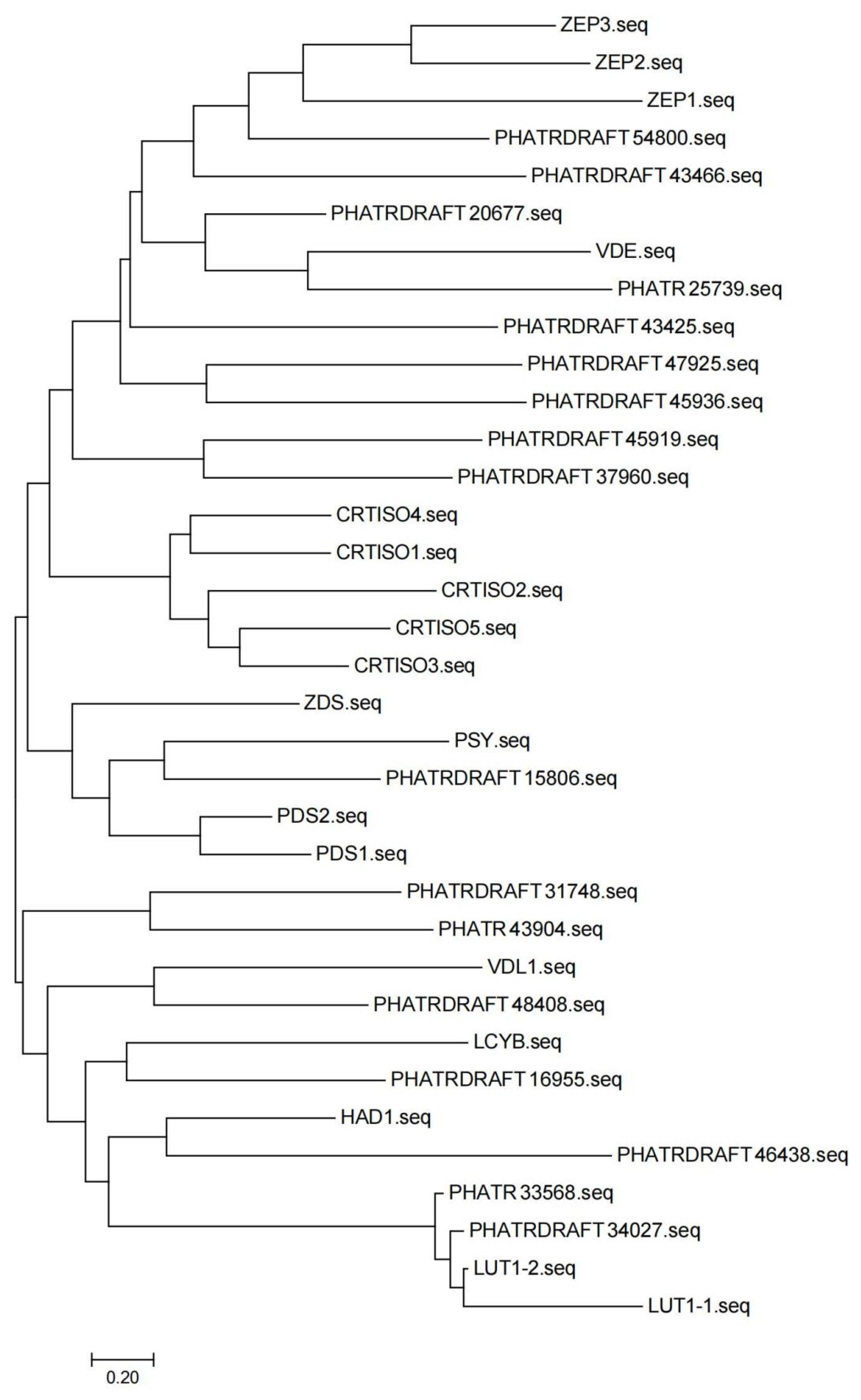
2.4. Analysis of Hub Genes for the Promotion of Fucoxanthin Synthesis in MeJA-Treated P. tricornutum
3. Discussion
4. Materials and Methods
4.1. Cultivation of P. tricornutum
4.2. Determination of Growth Curves of P. tricornutum
4.3. Extraction and Determination of Fucoxanthin
4.4. Extraction of Total RNA from P. tricornutum
4.5. Transcriptome Sequencing
4.6. Differential Genes Expression Analysis
4.7. WGCNA
4.8. Functional Analysis of Key Genes for Fucoxanthin Synthesis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Kwon, D.Y.; Vuong, T.T.; Choi, J.; Lee, T.S.; Um, J.-I.; Koo, S.Y.; Hwang, K.T.; Kim, S.M. Fucoxanthin biosynthesis has a positive correlation with the specific growth rate in the culture of microalga Phaeodactylum tricornutum. J. Appl. Phycol. 2021, 33, 1473–1485. [Google Scholar] [CrossRef]
- McClure, D.D.; Luiz, A.; Gerber, B.; Barton, G.W.; Kavanagh, J.M. An investigation into the effect of culture conditions on fucoxanthin production using the marine microalgae Phaeodactylum tricornutum. Algal. Res. 2018, 29, 41–48. [Google Scholar] [CrossRef]
- Neumann, U.; Derwenskus, F.; Flaiz Flister, V.; Schmid-Staiger, U.; Hirth, T.; Bischoff, S.C. Fucoxanthin, A Carotenoid Derived from Phaeodactylum tricornutum Exerts Antiproliferative and Antioxidant Activities In Vitro. Antioxidants 2019, 8, 183. [Google Scholar] [CrossRef]
- Bauer, C.; Schmitz, C.; Corrêa, R.; Herrera, C.; Ramlov, F.; Oliveira, E.; Pizzato, A.; Varela, L.; Cabral, D.; Yunes, R. In vitro fucoxanthin production by the Phaeodactylum tricornutum diatom. Stud. Nat. Prod. Chem. 2019, 63, 211–242. [Google Scholar]
- Sun, J.; Zhou, C.; Cheng, P.; Zhu, J.; Hou, Y.; Li, Y.; Zhang, J.; Yan, X. A simple and efficient strategy for fucoxanthin extraction from the microalga Phaeodactylum tricornutum. Algal. Res. 2022, 61, 102610. [Google Scholar] [CrossRef]
- Gómez-Loredo, A.; Benavides, J.; Rito-Palomares, M. Growth kinetics and fucoxanthin production of Phaeodactylum tricornutum and Isochrysis galbana cultures at different light and agitation conditions. J. Appl. Phycol. 2016, 28, 849–860. [Google Scholar] [CrossRef]
- Kadono, T.; Kira, N.; Suzuki, K.; Iwata, O.; Ohama, T.; Okada, S.; Nishimura, T.; Akakabe, M.; Tsuda, M.; Adachi, M. Effect of an Introduced Phytoene Synthase Gene Expression on Carotenoid Biosynthesis in the Marine Diatom Phaeodactylum tricornutum. Mar. Drugs 2015, 13, 5334–5357. [Google Scholar] [CrossRef]
- Satomi, Y. Antitumor and Cancer-preventative Function of Fucoxanthin: A Marine Carotenoid. Anticancer Res. 2017, 37, 1557–1562. [Google Scholar] [CrossRef]
- Yu, K.; Gong, Y.; Zhu, S.; Liu, H.; Wang, H. Effects of different exogenous elicitors on lcyb gene transcription and fucoxanthin content in Phaeodactylum tricornutum. J. Agric. Biotechnol. 2017, 25, 2009–2017. [Google Scholar]
- Sirhindi, G.; Mushtaq, R.; Gill, S.S.; Sharma, P.; Abd Allah, E.F.; Ahmad, P. Jasmonic acid and methyl jasmonate modulate growth, photosynthetic activity and expression of photosystem II subunit genes in Brassica oleracea L. Sci. Rep. 2020, 10, 9322. [Google Scholar] [CrossRef]
- Ho, T.T.; Murthy, H.N.; Park, S.Y. Methyl Jasmonate Induced Oxidative Stress and Accumulation of Secondary Metabolites in Plant Cell and Organ Cultures. Int. J. Mol. Sci. 2020, 21, 716. [Google Scholar] [CrossRef]
- Mc Gee, D.; Archer, L.; Parkes, R.; Fleming, G.T.A.; Santos, H.M.; Touzet, N. The role of methyl jasmonate in enhancing biomass yields and bioactive metabolites in Stauroneis sp. (Bacillariophyceae) revealed by proteome and biochemical profiling. J. Proteom. 2021, 249, 104381. [Google Scholar] [CrossRef] [PubMed]
- Nabi, N.; Singh, S.; Saffeullah, P. Responses of in vitro cell cultures to elicitation: Regulatory role of jasmonic acid and methyl jasmonate: A review. Vitr. Cell. Dev. Biol.-Plant 2021, 57, 341–355. [Google Scholar] [CrossRef]
- Singh, A.; Dwivedi, P. Methyl-jasmonate and salicylic acid as potent elicitors for secondary metabolite production in medicinal plants: A review. J. Pharmacogn. Phytochem. 2018, 7, 750–757. [Google Scholar]
- Wang, S.Y.; Shi, X.C.; Liu, F.Q.; Laborda, P. Effects of exogenous methyl jasmonate on quality and preservation of postharvest fruits: A review. Food Chem. 2021, 353, 129482. [Google Scholar] [CrossRef]
- Chen, M.; Guo, H.; Chen, S.; Li, T.; Li, M.; Rashid, A.; Xu, C.; Wang, K. Methyl Jasmonate Promotes Phospholipid Remodeling and Jasmonic Acid Signaling To Alleviate Chilling Injury in Peach Fruit. J. Agric. Food Chem. 2019, 67, 9958–9966. [Google Scholar] [CrossRef] [PubMed]
- Arano-Varela, H.; Cruz-Sosa, F.; Estrada-Zúñiga, M.E.; Fernández, F.J. Effects of phenylalanine and methyl jasmonate on verbascoside production in Buddleja cordata Kunth cell suspension cultures. S. Afr. J. Bot. 2020, 135, 41–49. [Google Scholar] [CrossRef]
- Baek, M.W.; Choi, H.R.; Solomon, T.; Jeong, C.S.; Lee, O.H.; Tilahun, S. Preharvest Methyl Jasmonate Treatment Increased the Antioxidant Activity and Glucosinolate Contents of Hydroponically Grown Pak Choi. Antioxidants 2021, 10, 131. [Google Scholar] [CrossRef]
- Luo, H.; He, W.; Li, D.; Bao, Y.; Riaz, A.; Xiao, Y.; Song, J.; Liu, C. Effect of methyl jasmonate on carotenoids biosynthesis in germinated maize kernels. Food Chem. 2020, 307, 125525. [Google Scholar] [CrossRef]
- Griffiths, G. Jasmonates: Biosynthesis, perception and signal transduction. Essays Biochem. 2020, 64, 501–512. [Google Scholar]
- Guan, Y.; Gong, Y.; Zhu, S. Cloning and expression analysis of carotenoid isomerase gene from Phaeodactylum tricornutum. J. Nucl. Agric. Sci. 2018, 32, 2098–2106. [Google Scholar]
- Zhao, Z.; Chen, J.; Ci, F.; Pang, H.; Cheng, N.; Xing, A. α-Carotene: A valuable carotenoid in biological and medical research. J. Sci. Food Agric. 2022, 102, 5606–5617. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Yuan, Q.; Zheng, H.; Zheng, A.; Zhang, F.; Sun, B. Molecular cloning and sequence analysis of the carotenoid isomerase gene in yellow-flower Chinese kale. AIP Conf. Proc. 2019, 2058, 020024. [Google Scholar]
- Ma, X.; Zheng, B.; Ma, Y.; Xu, W.; Wu, H.; Wang, S. Carotenoid accumulation and expression of carotenoid biosynthesis genes in mango flesh during fruit development and ripening. Sci. Hortic. 2018, 237, 201–206. [Google Scholar] [CrossRef]
- Xi, W.; Zhang, L.; Liu, S.; Zhao, G. The Genes of CYP, ZEP, and CCD1/4 Play an Important Role in Controlling Carotenoid and Aroma Volatile Apocarotenoid Accumulation of Apricot Fruit. Front. Plant Sci. 2020, 11, 607715. [Google Scholar] [CrossRef] [PubMed]
- Hornero-Méndez, D.; Britton, G. Involvement of NADPH in the cyclization reaction of carotenoid biosynthesis. FEBS Lett. 2002, 515, 133–136. [Google Scholar] [CrossRef] [PubMed]
- Corpas, F.J.; González-Gordo, S.; Palma, J.M. Nitric oxide and hydrogen sulfide modulate the NADPH-generating enzymatic system in higher plants. J. Exp. Bot. 2021, 72, 830–847. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Shi, F.; Zhan, W. Overexpression of ZWF1 and POS5 improves carotenoid biosynthesis in recombinant Saccharomyces cerevisiae. Lett. Appl. Microbiol. 2015, 61, 354–360. [Google Scholar] [CrossRef]
- Kim, S.M.; Jung, Y.J.; Kwon, O.N.; Cha, K.H.; Um, B.H.; Chung, D.; Pan, C.H. A potential commercial source of fucoxanthin extracted from the microalga Phaeodactylum tricornutum. Appl Biochem Biotechnol. 2012, 166, 1843–1855. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Chen, Y.; Wang, H.; Huang, Y.; Hu, Y.; Zhao, Y.; Gong, Y. Identification of Potential Factors for the Promotion of Fucoxanthin Synthesis by Methyl Jasmonic Acid Treatment of Phaeodactylum tricornutum. Mar. Drugs 2024, 22, 7. https://doi.org/10.3390/md22010007
Liu H, Chen Y, Wang H, Huang Y, Hu Y, Zhao Y, Gong Y. Identification of Potential Factors for the Promotion of Fucoxanthin Synthesis by Methyl Jasmonic Acid Treatment of Phaeodactylum tricornutum. Marine Drugs. 2024; 22(1):7. https://doi.org/10.3390/md22010007
Chicago/Turabian StyleLiu, Hao, Yawen Chen, Heyu Wang, Yaxuan Huang, Ying Hu, Yuxiang Zhao, and Yifu Gong. 2024. "Identification of Potential Factors for the Promotion of Fucoxanthin Synthesis by Methyl Jasmonic Acid Treatment of Phaeodactylum tricornutum" Marine Drugs 22, no. 1: 7. https://doi.org/10.3390/md22010007
APA StyleLiu, H., Chen, Y., Wang, H., Huang, Y., Hu, Y., Zhao, Y., & Gong, Y. (2024). Identification of Potential Factors for the Promotion of Fucoxanthin Synthesis by Methyl Jasmonic Acid Treatment of Phaeodactylum tricornutum. Marine Drugs, 22(1), 7. https://doi.org/10.3390/md22010007