
Echinochrome Ameliorates Physiological, Immunological, and Histopathological Alterations Induced by Ovalbumin in Asthmatic Mice by Modulating the Keap1/Nrf2 Signaling Pathway


 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. High-Performance Liquid Chromatography (HPLC)
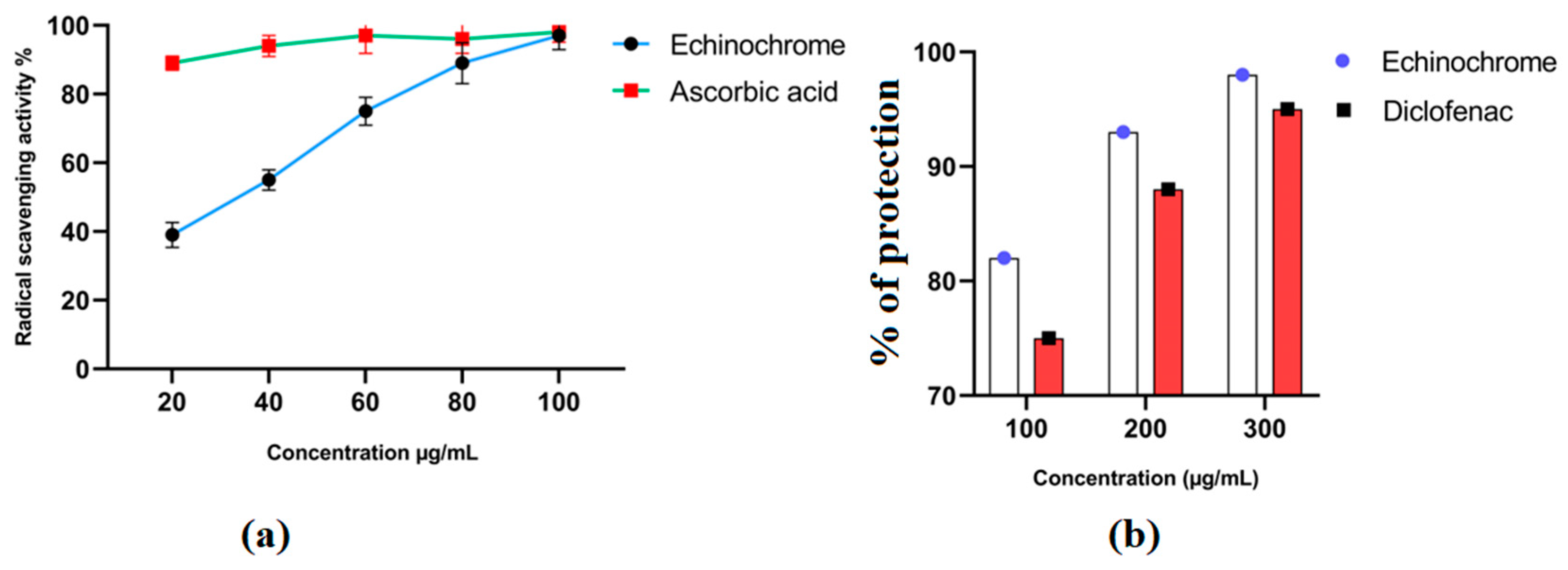
2.2. In Vitro Antioxidant and Anti-Inflammatory Activities
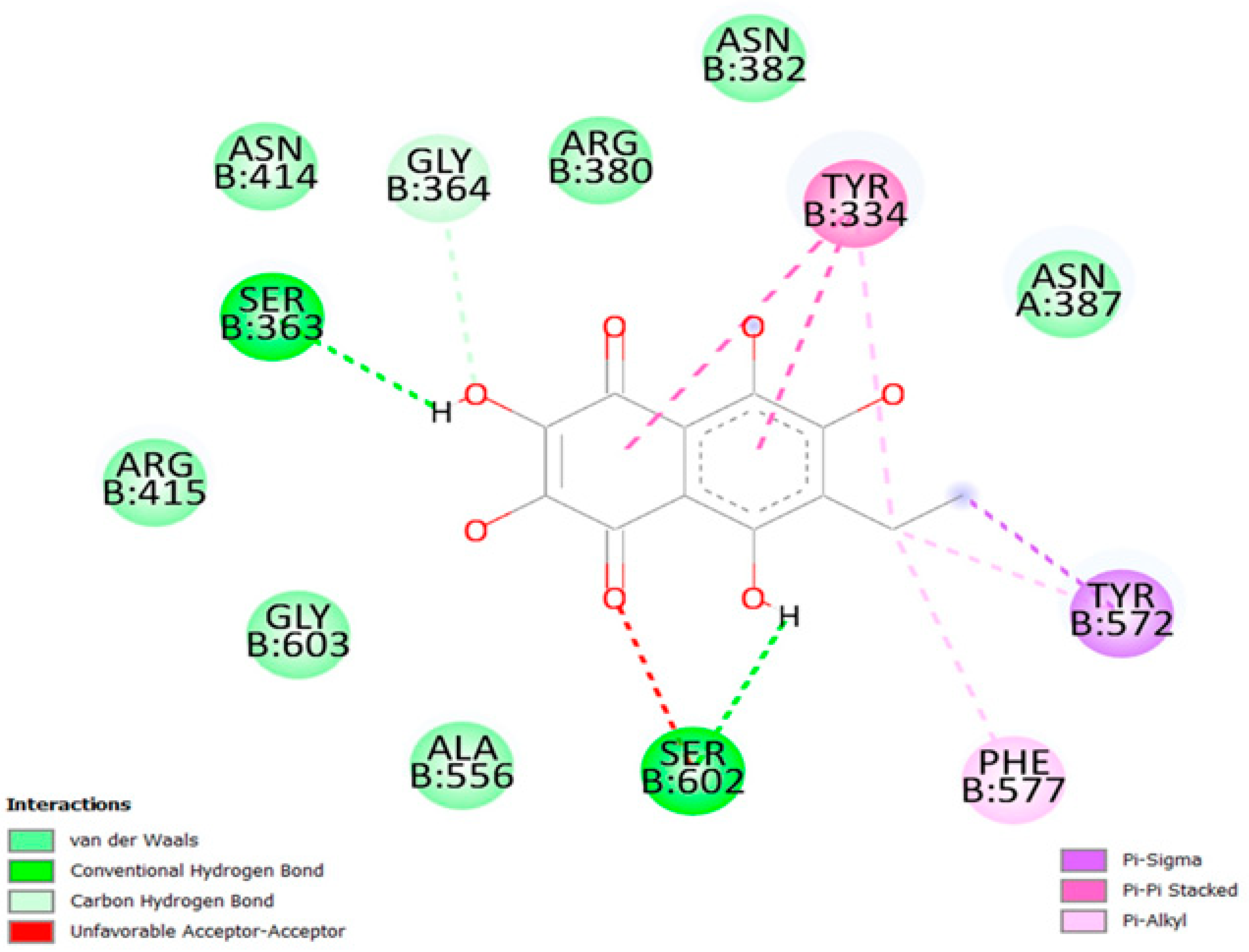
2.3. Molecular Docking Interaction between Ech and Keap1
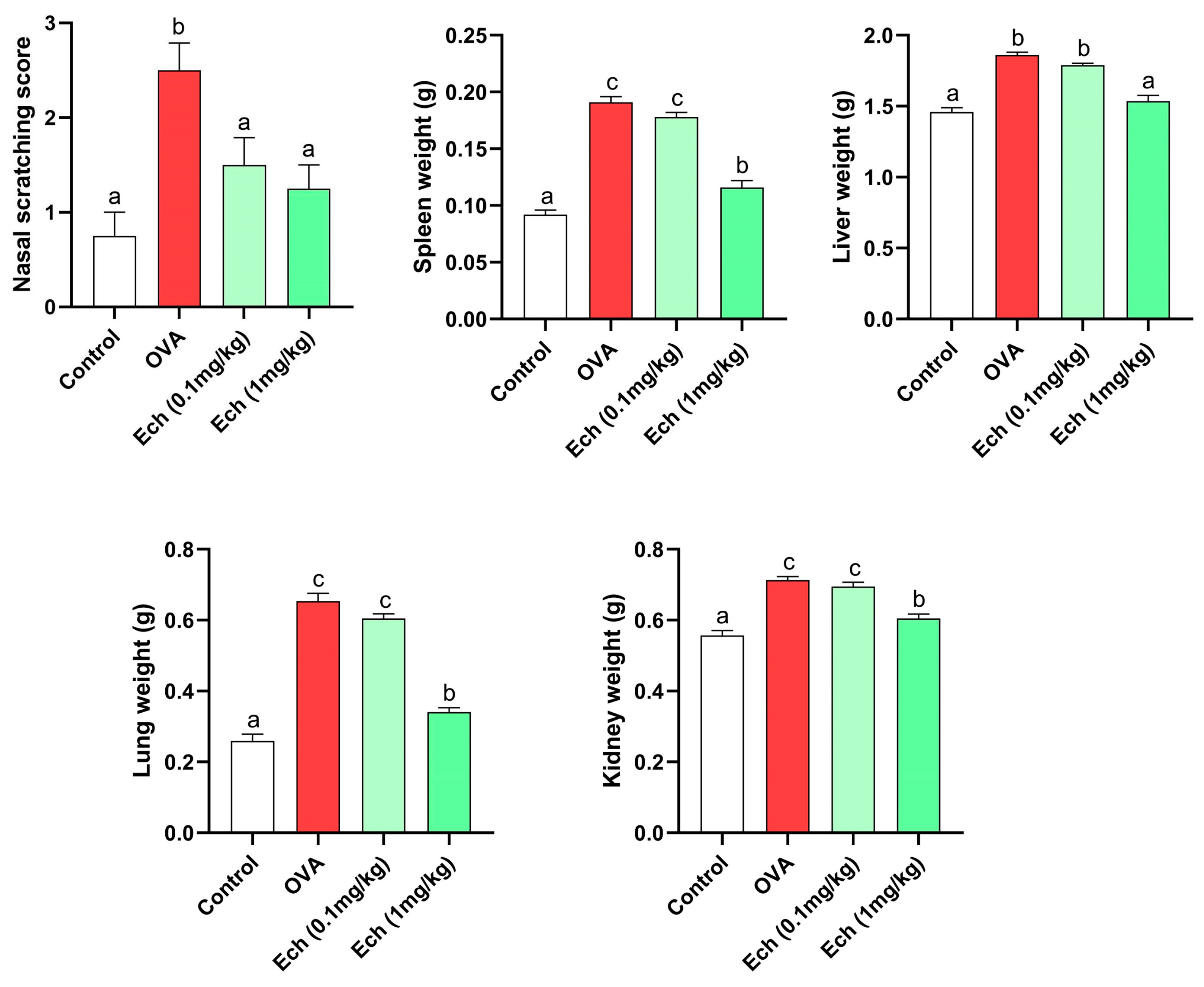
2.4. Effect of Ech on Morphology, Body Weight, and Nasal Scratching
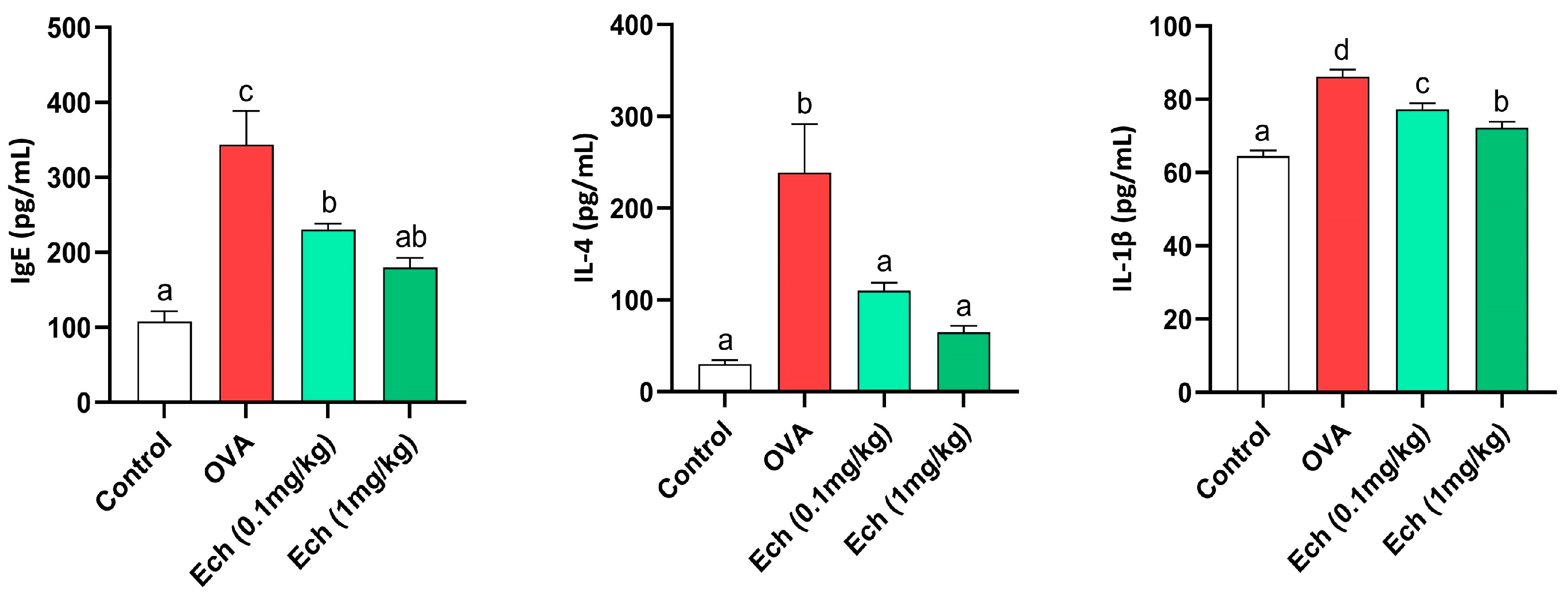
2.5. Ech Reduced the Serum Levels of IgE, IL-4, and IL-1β
2.6. Effect of Ech on Liver and Kidney Function
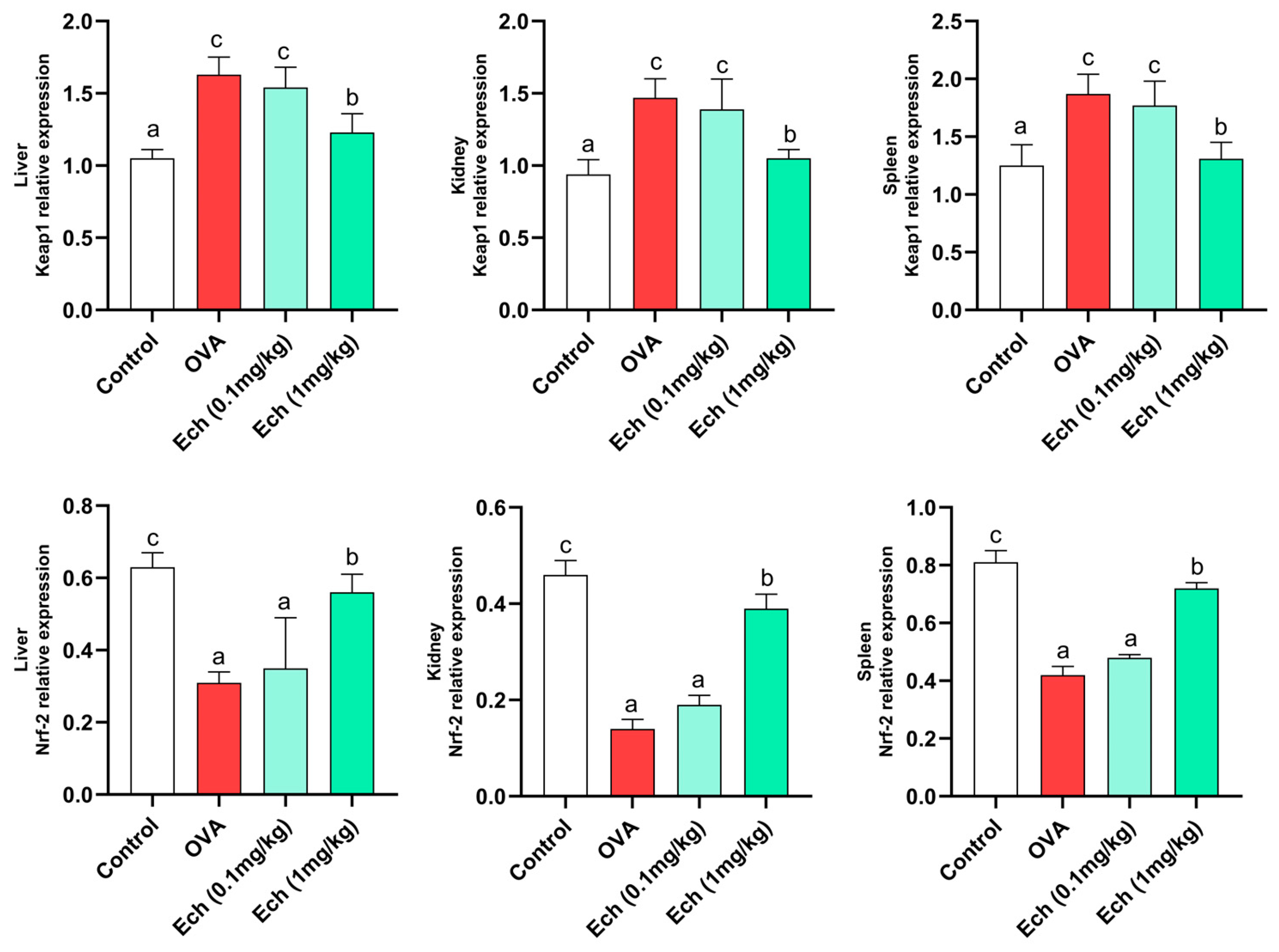
2.7. Effect of Ech on Keap1 and Nrf2 Protein Levels in the Liver, Kidney, and Spleen of Asthmatic Mice
2.8. Effects of Ech on the Expression of Keap1 and Nrf2 Genes in the Liver, Kidney, and Spleen of Asthmatic Mice
2.9. Effect of Ech on Kidney Antioxidants and Oxidative Stress Markers
2.10. Effect of Ech on Liver Antioxidants and Oxidative Stress Markers
2.11. Effect of Ech on Spleen Antioxidants and Oxidative Stress Markers
2.12. Effect of Ech on Lung Histopathology
2.13. Effect of Ech on Liver Histopathology
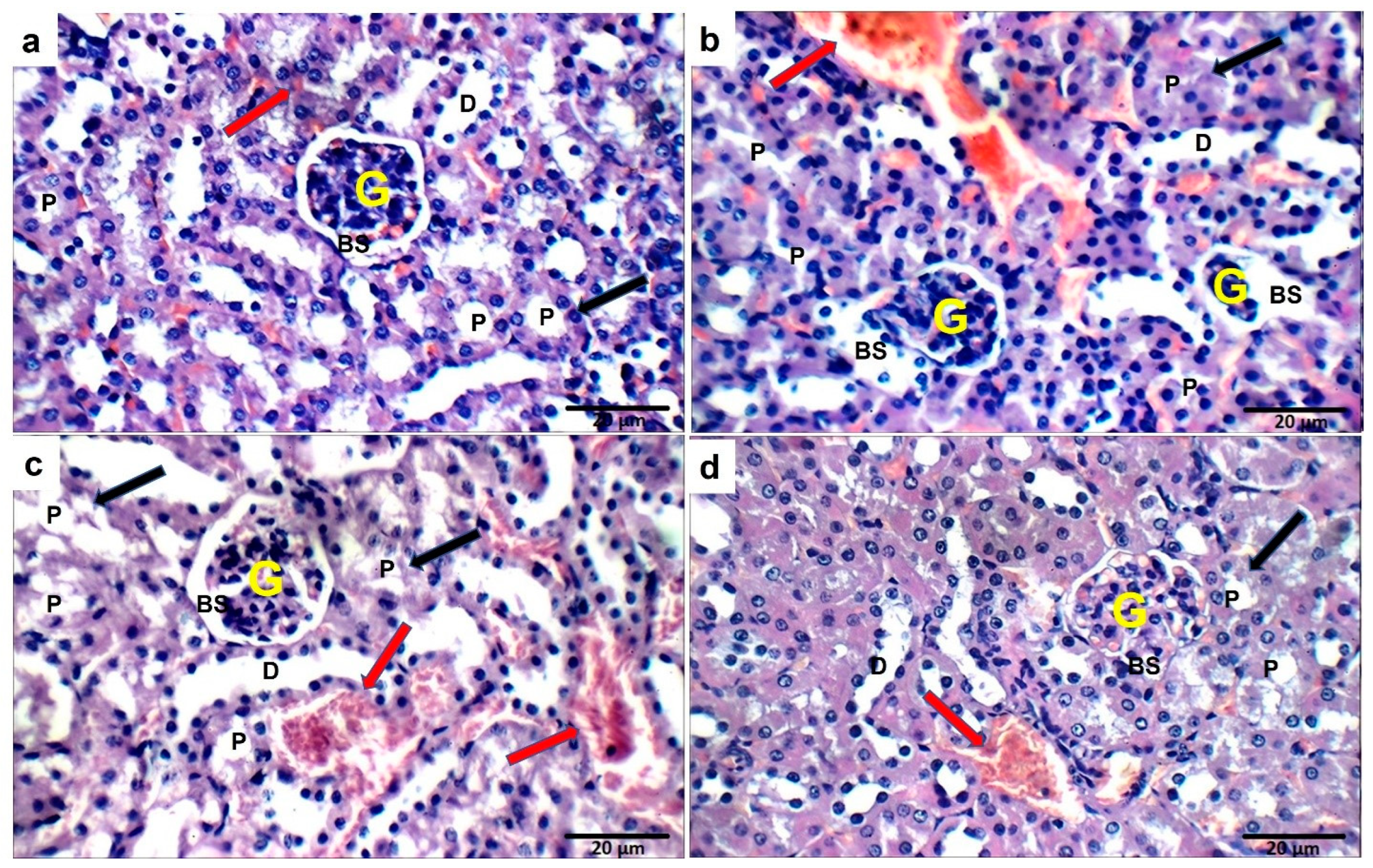
2.14. Effect of Ech on Kidney Histopathology
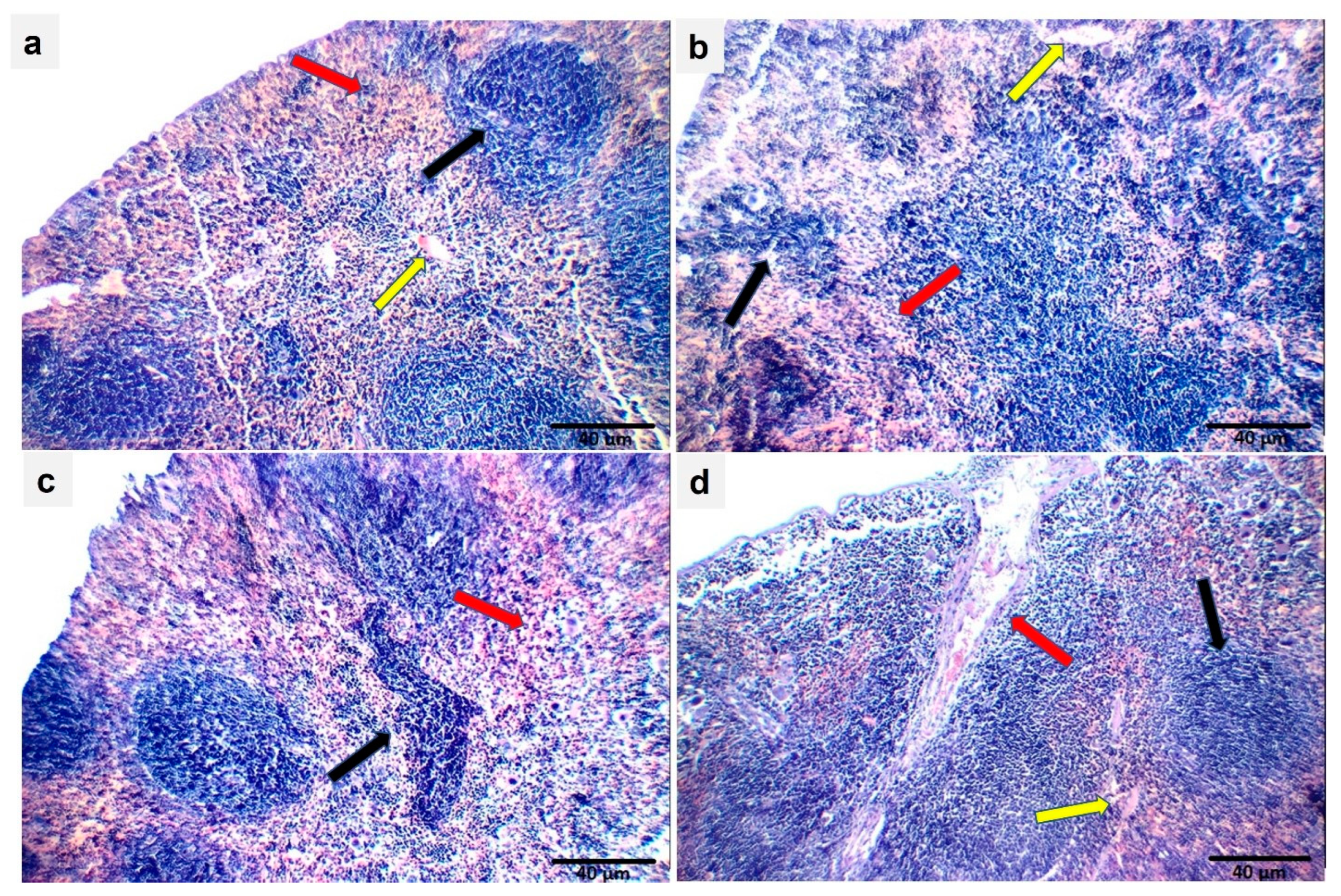
2.15. Effect of Ech on Spleen Histopathology
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Ech Extraction
4.3. HPLC Analysis
4.4. In Vitro Biological Studies
4.4.1. Antioxidant Activity Using DPPH Radical Scavenging Protocol
4.4.2. Anti-Inflammatory Activity Using Heat-Induced Hemolysis Protocol
4.5. Molecular Docking Interaction between Ech and Keap1
4.6. Experimental Animals
4.7. Animal Grouping and Experiment Design
4.8. Evaluation of Body Weight and Nasal Scratching
4.9. Sample Collection
4.10. Measurement of Serum Levels of IgE, IL-4, and IL-1β
4.11. Evaluation of Serum Biochemical Parameters
4.12. Western Blot Assays
4.13. Real-Time Reverse Transcriptase-Polymerase Chain Reaction (RT-PCR)
4.14. Determination of Liver, Kidney, and Spleen Oxidative Stress Biomarkers
4.15. Histopathological Examination of Lung, Kidney, Liver, and Spleen
4.16. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Zhu, Y.; Sun, D.; Liu, H.; Sun, L.; Jie, J.; Luo, J.; Peng, L.; Song, L. Bixin protects mice against bronchial asthma though modulating PI3K/Akt pathway. Int. Immunopharmacol. 2021, 101, 108266. [Google Scholar] [CrossRef]
- Zheng, M.; Guo, X.; Pan, R.; Gao, J.; Zang, B.; Jin, M. Hydroxysafflor Yellow A Alleviates Ovalbumin-Induced Asthma in a Guinea Pig Model by Attenuateing the Expression of Inflammatory Cytokines and Signal Transduction. Front. Pharmacol. 2019, 10, 328. [Google Scholar] [CrossRef] [PubMed]
- Zou, B.; Fu, Y.; Cao, C.; Pan, D.; Wang, W.; Kong, L. Gentiopicroside ameliorates ovalbumin-induced airway inflammation in a mouse model of allergic asthma via regulating SIRT1/NF-κB signaling pathway. Pulm. Pharmacol. Ther. 2021, 68, 102034. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Luo, Z.; Chen, J.; Li, B.; Li, L.; Shen, C. Bavachin inhibits IL-4 expression by downregulating STAT6 phosphorylation and GATA-3 expression and ameliorates asthma inflammation in an animal model. Immunobiology 2022, 227, 152182. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, C.; Luo, J.; Hua, S.; Li, D.; Peng, L.; Liu, H.; Song, L. The protective role of Zingerone in a murine asthma model via activation of the AMPK/Nrf2/HO-1 pathway. Food Funct. 2021, 12, 3120–3131. [Google Scholar] [CrossRef]
- Daenen, K.; Andries, A.; Mekahli, D.; Van Schepdael, A.; Jouret, F.; Bammens, B. Oxidative stress in chronic kidney disease. Pediatr. Nephrol. 2019, 34, 975–991. [Google Scholar] [CrossRef]
- Muriel, P.; Gordillo, K.R. Role of Oxidative Stress in Liver Health and Disease. Oxidative Med. Cell. Longev. 2016, 2016, 9037051. [Google Scholar] [CrossRef]
- Li, A.-N.; Li, S.; Zhang, Y.-J.; Xu, X.-R.; Chen, Y.-M.; Li, H.-B. Resources and Biological Activities of Natural Polyphenols. Nutrients 2014, 6, 6020–6047. [Google Scholar] [CrossRef] [PubMed]
- Ozbek, E. Induction of Oxidative Stress in Kidney. Int. J. Nephrol. 2012, 2012, 465897. [Google Scholar] [CrossRef]
- Mebius, R.E.; Kraal, G. Structure and function of the spleen. Nat. Rev. Immunol. 2005, 5, 606–616. [Google Scholar] [CrossRef]
- Lu, M.-C.; Ji, J.-A.; Jiang, Z.-Y.; You, Q.-D. The Keap1–Nrf2–ARE Pathway As a Potential Preventive and Therapeutic Target: An Update. Med. Res. Rev. 2016, 36, 924–963. [Google Scholar] [CrossRef] [PubMed]
- Hassanein, E.H.M.; Sayed, A.M.; Hussein, O.E.; Mahmoud, A.M. Coumarins as Modulators of the Keap1/Nrf2/ARE Signaling Pathway. Oxidative Med. Cell. Longev. 2020, 2020, 1675957. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Dong, M.; Teng, F.; Cui, J.; Zhu, X.; Wang, W.; Wuniqiemu, T.; Qin, J.; Yi, L.; Wang, S.; et al. TMT-based quantitative proteomics reveals suppression of SLC3A2 and ATP1A3 expression contributes to the inhibitory role of acupuncture on airway inflammation in an OVA-induced mouse asthma model. Biomed. Pharmacother. 2021, 134, 111001. [Google Scholar] [CrossRef]
- Mayer, A.M.S.; Guerrero, A.J.; Rodríguez, A.D.; Taglialatela-Scafati, O.; Nakamura, F.; Fusetani, N. Marine Pharmacology in 2016-2017: Marine Compounds with Antibacterial, Antidiabetic, Antifungal, Anti-Inflammatory, Antiprotozoal, Antituberculosis and Antiviral Activities; Affecting the Immune and Nervous Systems, and Other Miscellaneous Mechanisms of Action. Mar. Drugs 2021, 19, 49. [Google Scholar]
- Anderson, H.A.; Mathieson, J.W.; Thomson, R.H. Distribution of spinochrome pigments in echinoids. Comp. Biochem. Physiol. 1969, 28, 333–345. [Google Scholar] [CrossRef]
- Park, G.-T.; Yoon, J.-W.; Yoo, S.-B.; Song, Y.-C.; Song, P.; Kim, H.-K.; Han, J.; Bae, S.-J.; Ha, K.-T.; Mishchenko, N.P.; et al. Echinochrome A Treatment Alleviates Fibrosis and Inflammation in Bleomycin-Induced Scleroderma. Mar. Drugs 2021, 19, 237. [Google Scholar] [CrossRef]
- Fedoreyev, S.A.; Krylova, N.V.; Mishchenko, N.P.; Vasileva, E.A.; Pislyagin, E.A.; Iunikhina, O.V.; Lavrov, V.F.; Svitich, O.A.; Ebralidze, L.K.; Leonova, G.N. Antiviral and Antioxidant Properties of Echinochrome A. Mar. Drugs 2018, 16, 509. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, A.S.; Sadek, S.A.; Hassanein, S.S.; Soliman, A.M. Hepatoprotective Effect of Echinochrome Pigment in Septic Rats. J. Surg. Res. 2019, 234, 317–324. [Google Scholar] [CrossRef]
- Han, J.; Zhang, S.; Jiang, B.; Wang, J.; Ge, X.; Wu, B.; Zhang, S.; Wang, D. Sesquiterpene lactones from Xanthium sibiricum Patrin alleviate asthma by modulating the Th1/Th2 balance in a murine model. Phytomedicine 2022, 99, 154032. [Google Scholar] [CrossRef]
- Yang, Z.; Li, X.; Fu, R.; Hu, M.; Wei, Y.; Hu, X.; Tan, W.; Tong, X.; Huang, F. Therapeutic Effect of Renifolin F on Airway Allergy in an Ovalbumin-Induced Asthma Mouse Model In Vivo. Molecules 2022, 27, 3789. [Google Scholar] [CrossRef]
- Dai, R.; Niu, M.; Wang, N.; Wang, Y. Syringin alleviates ovalbumin-induced lung inflammation in BALB/c mice asthma model via NF-κB signaling pathway. Environ. Toxicol. 2021, 36, 433–444. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, L.; Chen, B.; Zhuo, Q.; Bao, C.; Lin, L. Propofol inhibits NF-κB activation to ameliorate airway inflammation in ovalbumin (OVA)-induced allergic asthma mice. Int. Immunopharmacol. 2017, 51, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Adelusi, T.I.; Abdul-Hammed, M.; Idris, M.O.; Oyedele, Q.K.; Adedotun, I.O.J.H. Molecular dynamics, quantum mechanics and docking studies of some Keap1 inhibitors–An insight into the atomistic mechanisms of their antioxidant potential. Heliyon 2021, 7, e07317. [Google Scholar] [CrossRef]
- Cuadrado, A.; Rojo, A.I.; Wells, G.; Hayes, J.D.; Cousin, S.P.; Rumsey, W.L.; Attucks, O.C.; Franklin, S.; Levonen, A.-L.; Kensler, T.W.; et al. Therapeutic targeting of the NRF2 and KEAP1 partnership in chronic diseases. Nat. Rev. Drug Discov. 2019, 18, 295–317. [Google Scholar] [CrossRef]
- Crisman, E.; Duarte, P.; Dauden, E.; Cuadrado, A.; Rodríguez-Franco, M.I.; López, M.G.; León, R.J.M.R.R. KEAP1-NRF2 protein–protein interaction inhibitors: Design, pharmacological properties and therapeutic potential. Med. Res. Rev. 2023, 43, 237–287. [Google Scholar] [CrossRef] [PubMed]
- Magesh, S.; Chen, Y.; Hu, L.J.M. Small molecule modulators of K eap1-N rf2-ARE pathway as potential preventive and therapeutic agents. Med. Res. Rev. 2012, 32, 687–726. [Google Scholar] [CrossRef]
- Lee, S.; Hu, L. Nrf2 activation through the inhibition of Keap1-Nrf2 protein-protein interaction. Med. Chem. Res. Int. J. Rapid Commun. Des. Mech. Action Biol. Act. Agents 2020, 29, 846–867. [Google Scholar] [CrossRef]
- Mishchenko, N.P.; Vasileva, E.A.; Gerasimenko, A.V.; Grigorchuk, V.P.; Dmitrenok, P.S.; Fedoreyev, S.A. Isolation and Structure Determination of Echinochrome A Oxidative Degradation Products. Molecules 2020, 25, 4778. [Google Scholar] [CrossRef]
- Cho, K.-S.; Park, M.-K.; Kang, S.-A.; Park, H.-Y.; Hong, S.-L.; Park, H.-K.; Yu, H.-S.; Roh, H.-J. Adipose-Derived Stem Cells Ameliorate Allergic Airway Inflammation by Inducing Regulatory T Cells in a Mouse Model of Asthma. Mediat. Inflamm. 2014, 2014, 436476. [Google Scholar] [CrossRef] [PubMed]
- Yan, G.H.; Choi, Y.H. Salidroside Attenuates Allergic Airway Inflammation Through Negative Regulation of Nuclear Factor-Kappa B and p38 Mitogen–Activated Protein Kinase. J. Pharmacol. Sci. 2014, 126, 126–135. [Google Scholar] [CrossRef]
- Lambrecht, B.N.; Hammad, H. The immunology of asthma. Nat. Immunol. 2015, 16, 45–56. [Google Scholar] [CrossRef]
- Roh, J.-H.; Lee, H.; Yun-Jeong, B.; Park, C.S.; Kim, H.-J.; Yoon, S.-Y. A nationwide survey of the association between nonalcoholic fatty liver disease and the incidence of asthma in Korean adults. PLoS ONE 2022, 17, e0262715. [Google Scholar] [CrossRef] [PubMed]
- Ozer, J.; Ratner, M.; Shaw, M.; Bailey, W.; Schomaker, S. The current state of serum biomarkers of hepatotoxicity. Toxicology 2008, 245, 194–205. [Google Scholar] [CrossRef] [PubMed]
- Khaldi, T.; Chekchaki, N.; Rouibah, Z.; Chouala, K.; Cheniti, H.; Boumendjel, M.; Taibi, F.; Messarah, M.; Boumendjel, A. Preventive effects of oral administration of Nigella sativa oil against smokeless tobacco induced toxicity and oxidative stress in the liver and kidney of allergic asthma induced rats. Toxicol. Environ. Health Sci. 2022, 14, 291–300. [Google Scholar] [CrossRef]
- Iribarren, C.; Sidney, S.; Lydick, E.; Sorel, M.E.; Eisner, M.D. The association between asthma, asthma therapeutic classes and hepatic enzyme elevation among adult HMO members. Compr. Ther. 2001, 27, 133–139. [Google Scholar] [CrossRef]
- Huang, H.-L.; Ho, S.-Y.; Li, C.-H.; Chu, F.-Y.; Ciou, L.-P.; Lee, H.-C.; Chen, W.-L.; Tzeng, N.-S. Bronchial asthma is associated with increased risk of chronic kidney disease. BMC Pulm. Med. 2014, 14, 80. [Google Scholar] [CrossRef]
- Small, I.; Moreira, A.; Couto, M. Practical approach to managing exercise-induced asthma in children and adults. Prim. Care Respir. J. 2013, 22, 126–129. [Google Scholar] [CrossRef] [PubMed]
- Assayag, M.; Goldstein, S.; Samuni, A.; Kaufman, A.; Berkman, N. The nitroxide/antioxidant 3-carbamoyl proxyl attenuates disease severity in murine models of severe asthma. Free Radic. Biol. Med. 2021, 177, 181–188. [Google Scholar] [CrossRef]
- Rawash, M.A.; Mohamed, A.S.; El-Zayat, E.M. The Concurrent Therapeutic Potential of Adipose-derived Mesenchymal Stem Cells on Gentamycin-induced Hepatorenal Toxicity in Rats. Curr. Stem. Cell Res. Ther. 2022, 17, 808–814. [Google Scholar]
- Ajayi, B.O.; Olajide, T.A.; Olayinka, E.T. 6-gingerol attenuates pulmonary inflammation and oxidative stress in mice model of house dust mite-induced asthma. Adv. Redox Res. 2022, 5, 100036. [Google Scholar] [CrossRef]
- Malaquias, M.A.S.; Oyama, L.A.; Jericó, P.C.; Costa, I.; Padilha, G.; Nagashima, S.; Lopes-Pacheco, M.; Rebelatto, C.L.K.; Michelotto, P.V.; Xisto, D.G.; et al. Effects of mesenchymal stromal cells play a role the oxidant/antioxidant balance in a murine model of asthma. Allergol. Immunopathol. 2018, 46, 136–143. [Google Scholar] [CrossRef]
- Tiwari, M.; Dwivedi, U.N.; Kakkar, P. Tinospora cordifolia extract modulates COX-2, iNOS, ICAM-1, pro-inflammatory cytokines and redox status in murine model of asthma. J. Ethnopharmacol. 2014, 153, 326–337. [Google Scholar] [CrossRef] [PubMed]
- Sadek, S.A.; Hassanein, S.S.; Mohamed, A.S.; Soliman, A.M.; Fahmy, S.R. Echinochrome pigment extracted from sea urchin suppress the bacterial activity, inflammation, nociception, and oxidative stress resulted in the inhibition of renal injury in septic rats. J. Food Biochem. 2022, 46, e13729. [Google Scholar] [CrossRef]
- Tang, X.; Nishimura, A.; Ariyoshi, K.; Nishiyama, K.; Kato, Y.; Vasileva, E.A.; Mishchenko, N.P.; Fedoreyev, S.A.; Stonik, V.A.; Kim, H.K.; et al. Echinochrome Prevents Sulfide Catabolism-Associated Chronic Heart Failure after Myocardial Infarction in Mice. Mar. Drugs 2023, 21, 52. [Google Scholar] [CrossRef]
- Giles, G.I.; Jacob, C. Reactive sulfur species: An emerging concept in oxidative stress. Biol. Chem. 2002, 383, 375–388. [Google Scholar] [CrossRef]
- Clark, A.H. Monograph of shallow-water Indo-West Pacific echinoderms. Br. Mus. (Nat. Hist.) Publ. 1971, 690, 234. [Google Scholar]
- Amarowicz, R.; Synowiecki, J.; Shahidi, F. Sephadex LH-20 separation of pigments from shells of red sea urchin (Strongylocentrotus franciscanus). Food Chem. 1994, 51, 227–229. [Google Scholar] [CrossRef]
- Tung, Y.-T.; Wu, J.-H.; Kuo, Y.-H.; Chang, S.-T. Antioxidant activities of natural phenolic compounds from Acacia confusa bark. Bioresour. Technol. 2007, 98, 1120–1123. [Google Scholar] [CrossRef]
- Abdelfattah, M.A.; Mohamed, A.S.; Ibrahim, S.A.; Fahmy, S.R. Allolobophora caliginosa coelomic fluid and extract alleviate glucocorticoid-induced osteoporosis in mice by suppressing oxidative stress and regulating osteoblastic/osteoclastic-related markers. Sci. Rep. 2023, 13, 2090. [Google Scholar] [CrossRef] [PubMed]
- Parameswari, P.; Devika, R.; Vijayaraghavan, P. In vitro anti-inflammatory and antimicrobial potential of leaf extract from Artemisia nilagirica (Clarke) Pamp. Saudi J. Biol. Sci. 2019, 26, 460–463. [Google Scholar] [CrossRef]
- DeLano, W.L. Pymol: An open-source molecular graphics tool. CCP4 Newsl. Protein Crystallogr. 2002, 40, 82–92. [Google Scholar]
- Jnoff, E.; Albrecht, C.; Barker, J.J.; Barker, O.; Beaumont, E.; Bromidge, S.; Brookfield, F.; Brooks, M.; Bubert, C.; Ceska, T.; et al. Binding Mode and Structure–Activity Relationships around Direct Inhibitors of the Nrf2–Keap1 Complex. ChemMedChem 2014, 9, 699–705. [Google Scholar] [CrossRef]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open Babel: An open chemical toolbox. J. Cheminf. 2011, 3, 33. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Adasme, M.F.; Linnemann, K.L.; Bolz, S.N.; Kaiser, F.; Salentin, S.; Haupt, V.J.; Schroeder, M. PLIP 2021: Expanding the scope of the protein–ligand interaction profiler to DNA and RNA. Nucleic Acids Res. 2021, 49, W530–W534. [Google Scholar] [CrossRef] [PubMed]
- Bai, D.; Sun, T.; Lu, F.; Shen, Y.; Zhang, Y.; Zhang, B.; Yu, G.; Li, H.; Hao, J. Eupatilin Suppresses OVA-Induced Asthma by Inhibiting NF-κB and MAPK and Activating Nrf2 Signaling Pathways in Mice. Int. J. Mol. Sci. 2022, 23, 1582. [Google Scholar]
- Yun, H.R.; Ahn, S.W.; Seol, B.; Vasileva, E.A.; Mishchenko, N.P.; Fedoreyev, S.A.; Stonik, V.A.; Han, J.; Ko, K.S.; Rhee, B.D. Echinochrome A treatment alleviates atopic dermatitis-like skin lesions in NC/Nga mice via IL-4 and IL-13 suppression. Mar. Drugs 2021, 19, 622. [Google Scholar] [CrossRef]
- Liu, C.; You, J.; Lu, Y.; Sun, J.; Pan, J.; Li, Y.; Liu, T.; Li, Y.; Wang, A.; Zhang, X. Protective effects on ovalbumin-induced mouse asthma models and qualitative and quantitative analysis of multiple compounds in Gerberae Piloselloidis Herba. J. Sep. Sci. 2022, 45, 990–1005. [Google Scholar] [CrossRef]
- Abdelaziz, M.H.; El-Dakdoky, M.H.; Ahmed, T.A.; Mohamed, A.S. Biological impacts of the green synthesized silver nanoparticles on the pregnant albino rats and their fetuses. Birth Defects Res. 2023, 115, 441–457. [Google Scholar] [CrossRef]
- Abdelaziz, M.H.; Abdelfattah, M.A.; Bahaaeldine, M.A.; Rashed, A.R.; Mohamed, A.S.; Ali, M.F.; Elbatran, M.M.; Saad, D.Y. Earthworm Extract Enhanced Organ Functions in Diabetic Rats by Ameliorating Physiological and Structural Changes. Biointerface Res. Appl. Chem. 2022, 13, 445. [Google Scholar]
- Koura, R.A.A.; Mohamed, H.R.H.; Baiomy, A.A.; Bahaaeldine, M.A.; Mohamed, A.S. The Therapeutic Role of Chitosan-Saponin-Bentonite Nanocomposite on Acute Kidney Injury Induced by Chromium in Male Wistar Rats. Biointerface Res. Appl. Chem. 2023, 13, 595. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein–Ligand Interactions Profile | |||||
|---|---|---|---|---|---|
| Index | Residue | Amino Acid | Distance (Å) | Ligand Atom | Type of Interaction |
| 1 | 334B | TYR | 3.54 (3.08) | 19 O [O3] | Hydrogen bond acceptor |
| 2 | 363B | SER | 2.95 (2.06) | 16 H [O2] | Hydrogen bond acceptor |
| 3 | 382B | ASN | 3.88 (3.36) | 21 O [O3] | Hydrogen bond acceptor |
| 4 | 387A | ASN | 3.08 (2.15) | 19 O [O3] | Hydrogen bond donor |
| 5 | 414B | ASN | 4.04 (3.24) | 16 H [O2] | Hydrogen bond donor |
| 6 | 602B | SER | 2.88 (2.35) | 17 O [O3] | Hydrogen bond acceptor |
| 7 | 334B | TYR | 3.85 | 1 O | Hydrophobic interaction |
| 8 | 572B | TYR | 3.79 | 2 O | Hydrophobic interaction |
| 9 | 577B | PHE | 3.63 | 1 O | Hydrophobic interaction |
| 10 | 334 B | TYR | 3.88 | 5 C, 6 C, 7 C, 8 C, 9 C, 10 C | π-Stacking |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdelmawgood, I.A.; Mahana, N.A.; Badr, A.M.; Mohamed, A.S.; Al Shawoush, A.M.; Atia, T.; Abdelrazak, A.E.; Sakr, H.I. Echinochrome Ameliorates Physiological, Immunological, and Histopathological Alterations Induced by Ovalbumin in Asthmatic Mice by Modulating the Keap1/Nrf2 Signaling Pathway. Mar. Drugs 2023, 21, 455. https://doi.org/10.3390/md21080455
Abdelmawgood IA, Mahana NA, Badr AM, Mohamed AS, Al Shawoush AM, Atia T, Abdelrazak AE, Sakr HI. Echinochrome Ameliorates Physiological, Immunological, and Histopathological Alterations Induced by Ovalbumin in Asthmatic Mice by Modulating the Keap1/Nrf2 Signaling Pathway. Marine Drugs. 2023; 21(8):455. https://doi.org/10.3390/md21080455
Chicago/Turabian StyleAbdelmawgood, Islam Ahmed, Noha Ahmed Mahana, Abeer Mahmoud Badr, Ayman Saber Mohamed, Abdeljalil Mohamed Al Shawoush, Tarek Atia, Amir Elhadi Abdelrazak, and Hader I. Sakr. 2023. "Echinochrome Ameliorates Physiological, Immunological, and Histopathological Alterations Induced by Ovalbumin in Asthmatic Mice by Modulating the Keap1/Nrf2 Signaling Pathway" Marine Drugs 21, no. 8: 455. https://doi.org/10.3390/md21080455
APA StyleAbdelmawgood, I. A., Mahana, N. A., Badr, A. M., Mohamed, A. S., Al Shawoush, A. M., Atia, T., Abdelrazak, A. E., & Sakr, H. I. (2023). Echinochrome Ameliorates Physiological, Immunological, and Histopathological Alterations Induced by Ovalbumin in Asthmatic Mice by Modulating the Keap1/Nrf2 Signaling Pathway. Marine Drugs, 21(8), 455. https://doi.org/10.3390/md21080455