
Arctic Sea Ice Microalga Chlamydomonas latifrons KNF0041: Identification and Statistical Optimization of Medium for Enhanced Biomass and Omega-3/Omega-6

 ,
,  , and
, and
Abstract
:
1. Introduction
2. Results and Discussion
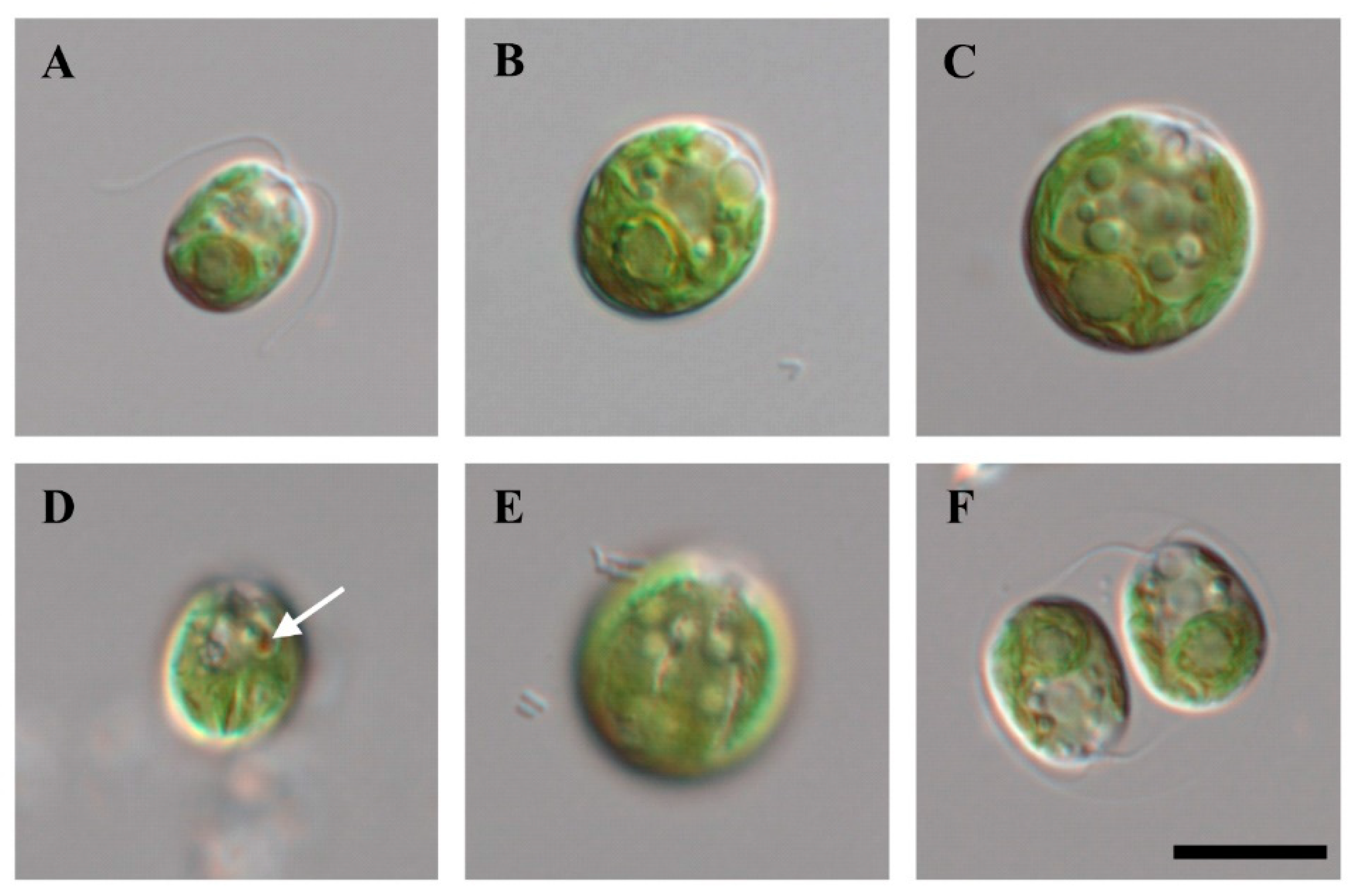
2.1. Morphological Characteristics of the Strain KNF0041
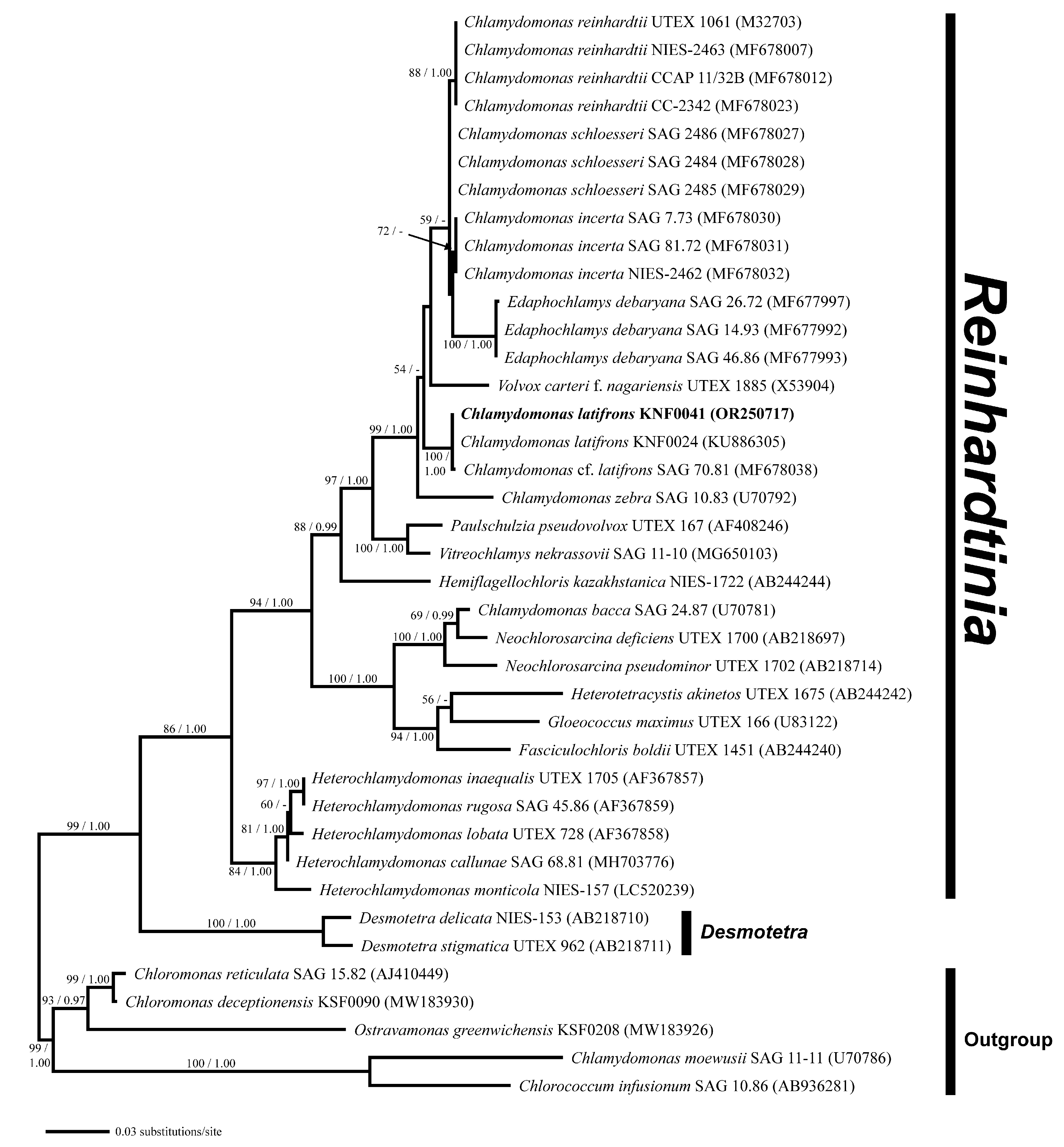
2.2. Molecular Phylogenetic Analyses
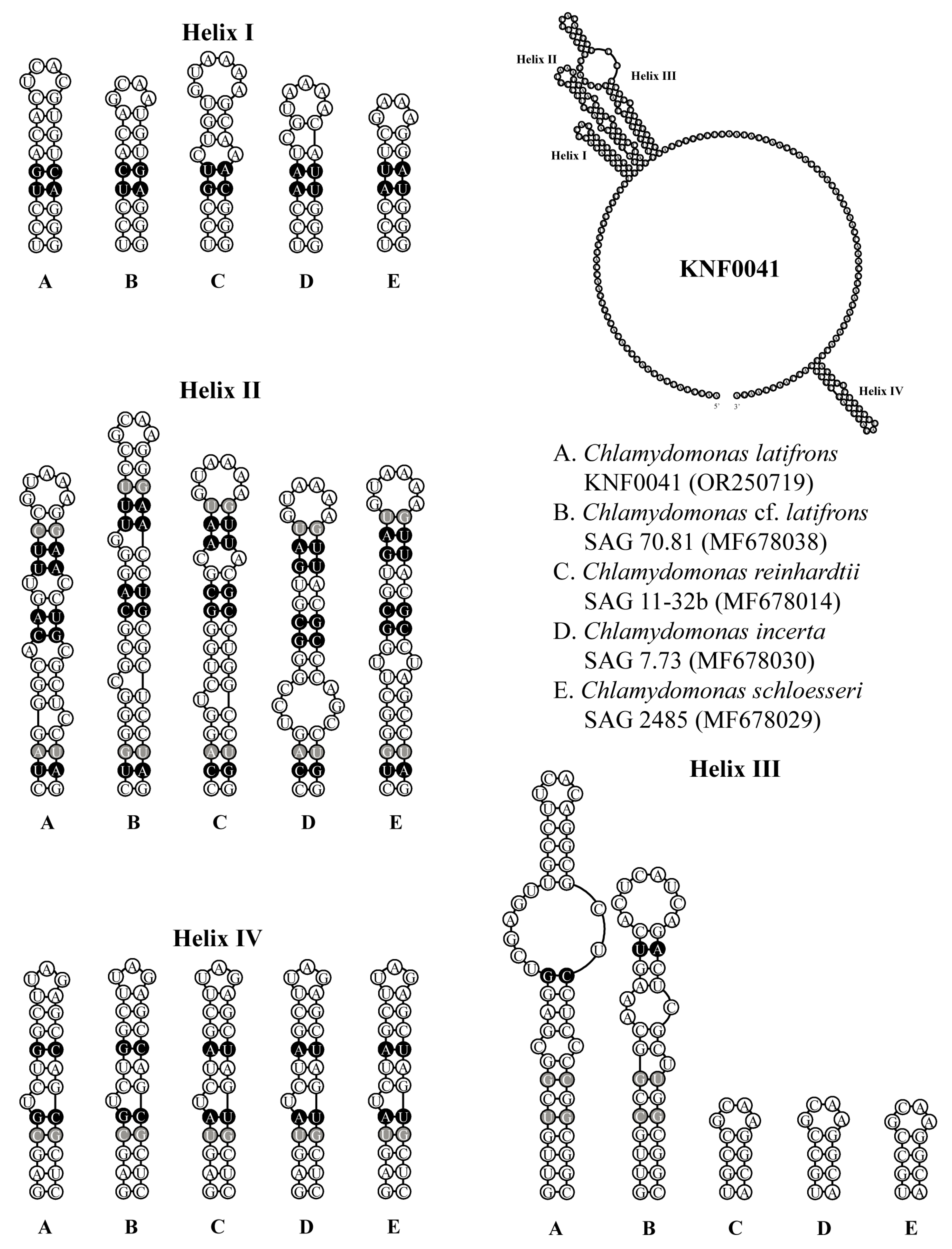
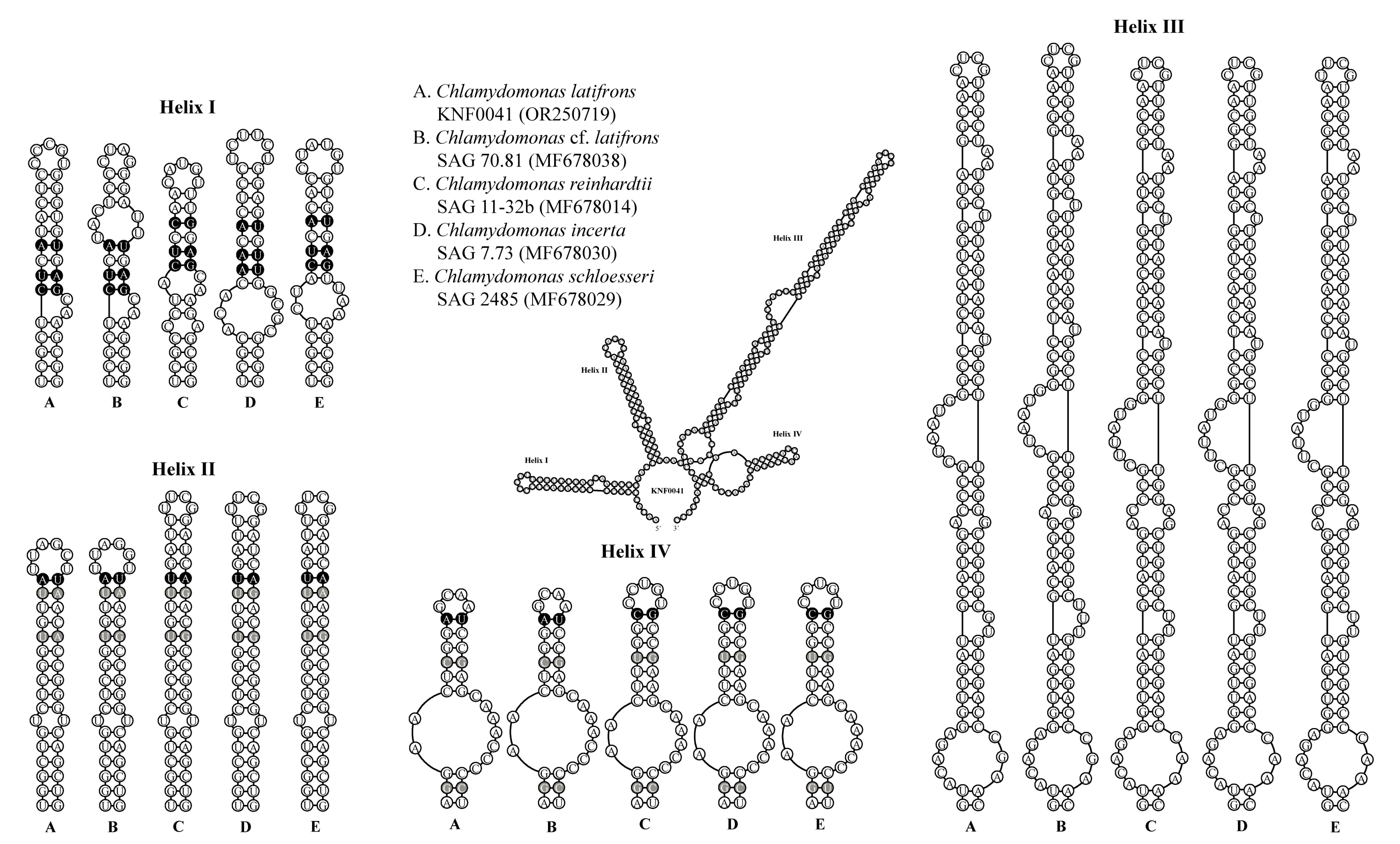
2.3. Secondary Structures of ITS1 and ITS2
2.4. Determination of Growth Temperature, Medium, and Light Conditions
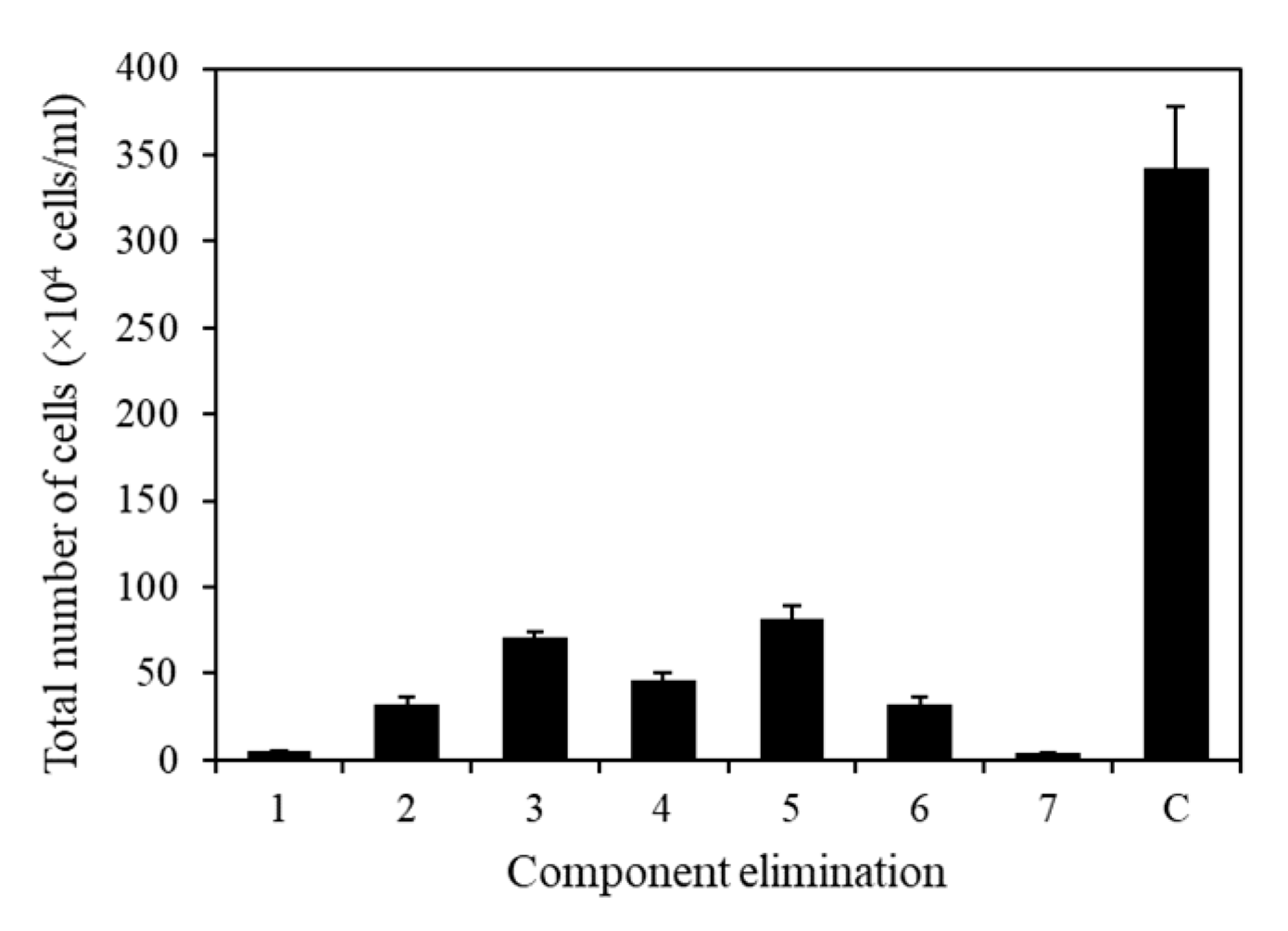
2.5. Component Selection
2.6. Screening of Components Using the Plackett–Burman Design
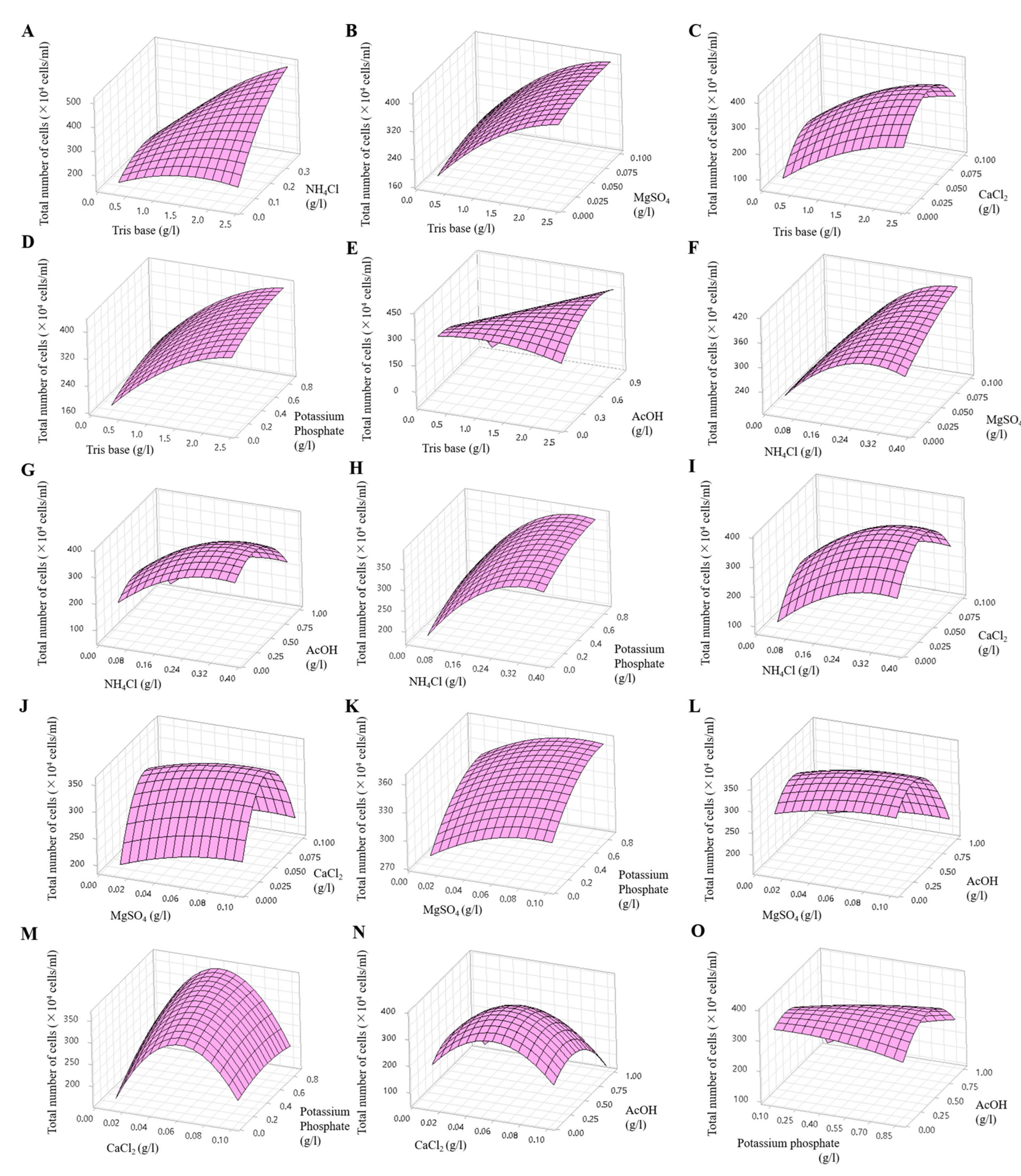
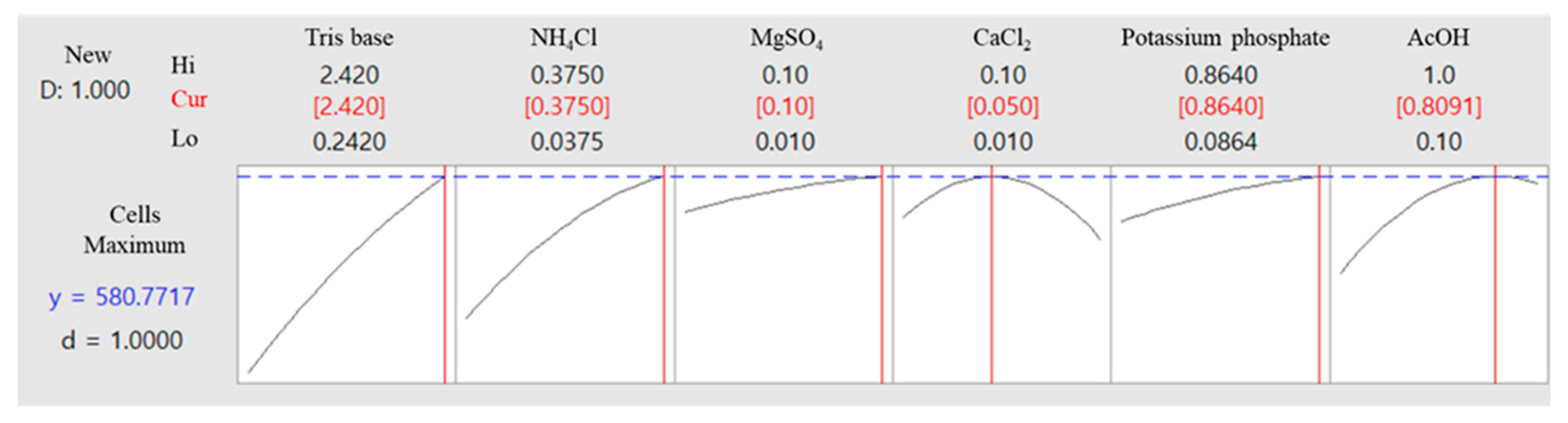
2.7. Optimization of Medium Components for KNF0041 Growth
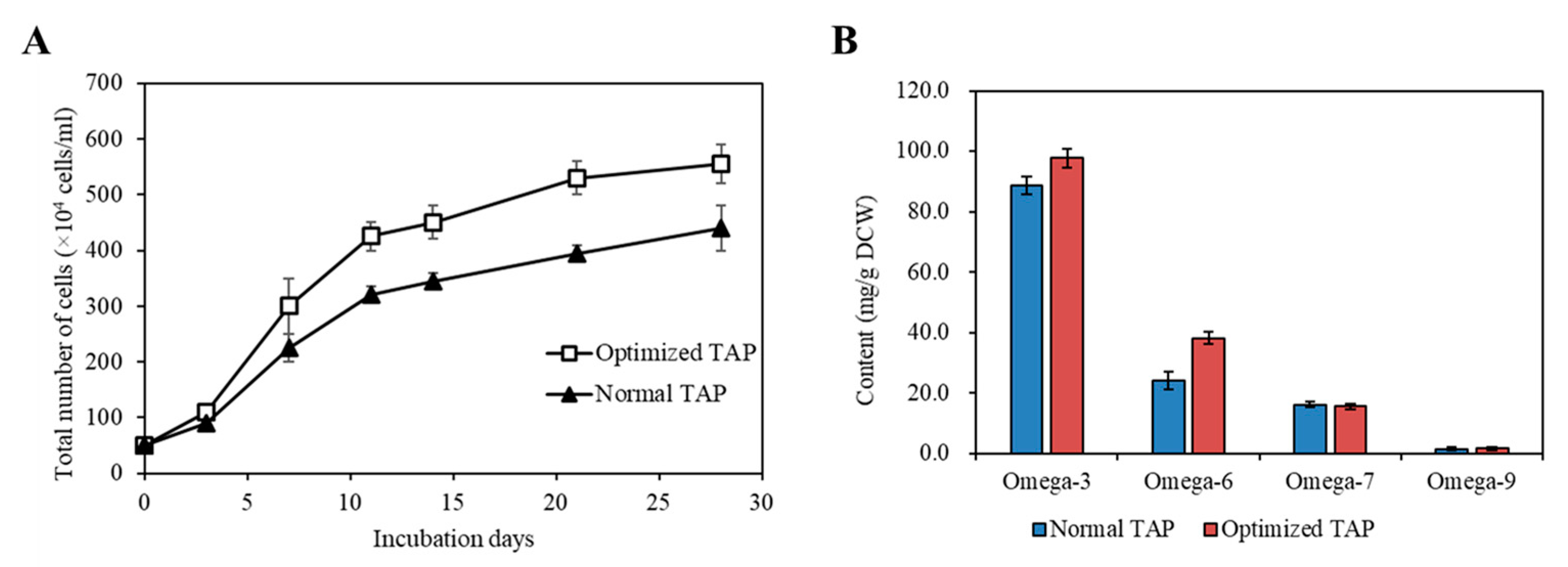
2.8. Comparison of the Strain KNF0041’s Growth and Omega-3/Omega-6 Fatty Acid Production in Normal and Optimized Media
3. Materials and Methods
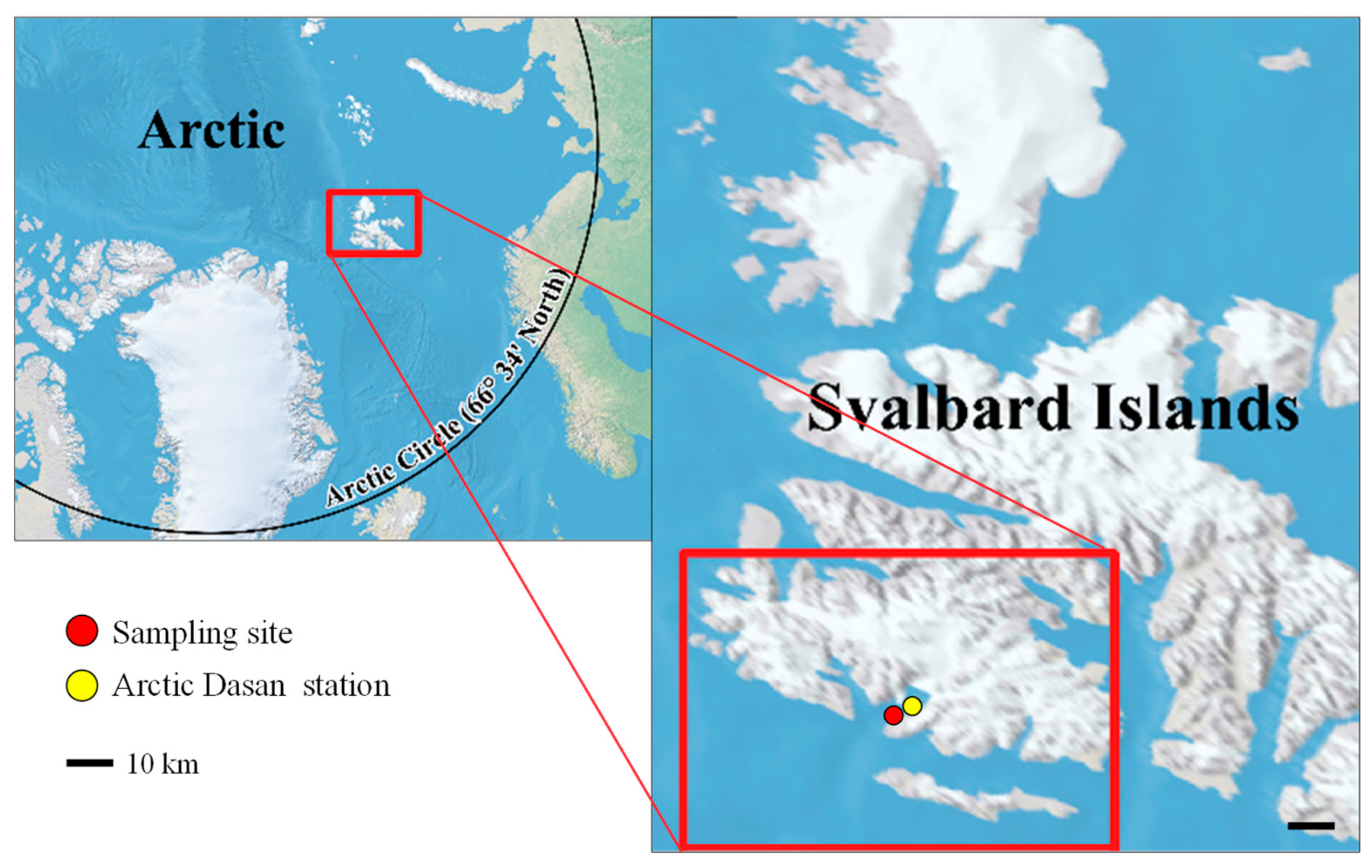
3.1. Isolation and Culture Condition
3.2. Morphological and Molecular Identification and Phylogenetic Analyses
3.3. ITS1 and ITS2 Secondary Structures
3.4. Statistical Optimization
3.5. Fatty Acid Methyl Ester (FAME) Analysis
3.6. Statistical Tests
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Borchert, E.; Jackson, S.A.; O’Gara, F.; Dobson, A.D. Psychrophiles as a source of novel antimicrobials. In Psychrophiles: From Biodiversity to Biotechnology; Margesin, R., Ed.; Springer: Singapore, 2017; pp. 527–540. [Google Scholar]
- Nguyen, D.L.; Hwang, J.; Kim, E.J.; Lee, J.H.; Han, S.J. Production and Characterization of a Recombinant Cold-Active Acetyl Xylan Esterase from Psychrophilic Paenibacillus sp. R4 Strain. Appl. Biochem. Microbiol. 2022, 58, 428–434. [Google Scholar] [CrossRef]
- Kim, E.J.; Kim, J.E.; Hwang, J.; Kim, I.-C.; Lee, S.G.; Kim, S.; Lee, J.H.; Han, S.J. Increased productivity and antifreeze activity of ice-binding protein from Flavobacterium frigoris PS1 produced using Escherichia coli as bioreactor. Appl. Biochem. Microbiol. 2019, 55, 489–494. [Google Scholar] [CrossRef]
- Brouchkov, A.; Melnikov, V.; Kalenova, L.; Fursova, O.; Pogorelko, G.; Potapov, V.; Fursova, N.; Ignatov, S.; Brenner, E.; Bezrukov, V. Permafrost bacteria in biotechnology: Biomedical applications. In Psychrophiles: From Biodiversity to Biotechnology; Margesin, R., Ed.; Springer: Singapore, 2017; pp. 541–554. [Google Scholar]
- Do, H.; Lee, J.H.; Lee, S.G.; Kim, H.J. Crystallization and preliminary X-ray crystallographic analysis of an ice-binding protein (FfIBP) from Flavobacterium frigoris PS1. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2012, 68, 806–809. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.J.; Jung, W.; Lim, S.; Kim, S.; Han, S.J.; Choi, H.-G. Growth and lipid content at low temperature of Arctic alga Chlamydomonas sp. KNM0029C. Bioproc. Biosyst. Eng. 2016, 39, 151–157. [Google Scholar] [CrossRef]
- Ettl, H. Die Gattung Chlamydomonas Ehrenberg (Chlamydomonas und die Nächstverwandten Gattungen II). Beih. Nova Hedwigia 1976, 49, 1–1122. [Google Scholar]
- Pröschold, T.; Marin, B.; Schlösser, U.G.; Melkonian, M. Molecular phylogeny and taxonomic revision of Chlamydomonas (Chlorophyta). I. Emendation of Chlamydomonas Ehrenberg and Chloromonas Gobi, and description of Oogamochlamys gen. nov. and Lobochlamys gen. nov. Protist 2001, 152, 265–300. [Google Scholar] [CrossRef] [PubMed]
- Hoham, R.W.; Bonome, T.A.; Martin, C.W.; Leebens-Mack, J.H. A combined 18S rDNA and rbcL phylogenetic analysis of Chloromonas and Chlamydomonas (Chlorophyceae, Volvocales) emphasizing snow and other cold-temperature habitats. J. Phycol. 2002, 38, 1051–1064. [Google Scholar] [CrossRef]
- Gutiérrez, S.; Svahn, S.L.; Johansson, M.E. Effects of omega-3 fatty acids on immune cells. Int. J. Mol. Sci. 2019, 20, 5028. [Google Scholar] [CrossRef]
- Remize, M.; Brunel, Y.; Silva, J.L.; Berthon, J.-Y.; Filaire, E. Microalgae n-3 PUFAs Production and Use in Food and Feed Industries. Mar. Drugs 2021, 19, 113. [Google Scholar] [CrossRef] [PubMed]
- Suh, S.S.; Hong, J.M.; Kim, E.J.; Jung, S.W.; Kim, S.M.; Kim, J.E.; Kim, I.C.; Kim, S. Anti-inflammation and Anti-Cancer Activity of Ethanol Extract of Antarctic Freshwater Microalga, Micractinium sp. Int. J. Med. Sci. 2018, 15, 929–936. [Google Scholar] [CrossRef]
- Kim, E.J.; Chae, H.; Koo, M.H.; Yu, J.; Kim, H.; Cho, S.M.; Hong, K.W.; Lee, J.Y.; Youn, U.J.; Kim, S. Statistical optimization of phytol and polyunsaturated fatty acid production in the Antarctic microalga Micractinium variabile KSF0031. Algae 2022, 37, 175–183. [Google Scholar] [CrossRef]
- Cvetkovska, M.; Hüner, N.P.; Smith, D.R. Chilling out: The evolution and diversification of psychrophilic algae with a focus on Chlamydomonadales. Polar Biol. 2017, 40, 1169–1184. [Google Scholar] [CrossRef]
- Sforza, E.; Simionato, D.; Giacometti, G.M.; Bertucco, A.; Morosinotto, T. Adjusted light and dark cycles can optimize photosynthetic efficiency in algae growing in photobioreactors. PLoS ONE 2012, 7, e38975. [Google Scholar] [CrossRef]
- Huarachi-Olivera, R.; Mata, M.T.; Valdés, J.; Riquelme, C. Biosorption of Zn (II) from Seawater Solution by the Microalgal Biomass of Tetraselmis marina AC16-MESO. Int. J. Mol. Sci. 2021, 22, 12799. [Google Scholar] [CrossRef]
- Fernandes, T.; Cordeiro, N. Effects of phosphorus-induced changes on the growth, nitrogen uptake, and biochemical composition of Pavlova pinguis and Hemiselmis cf. andersenii. J. Appl. Phycol. 2022, 34, 757–775. [Google Scholar] [CrossRef]
- Gorgich, M.; Passos, M.L.; Mata, T.M.; Martins, A.A.; Saraiva, M.L.M.; Caetano, N.S. Enhancing extraction and purification of phycocyanin from Arthrospira sp. with lower energy consumption. Energy Rep. 2020, 6, 312–318. [Google Scholar] [CrossRef]
- Kim, E.J.; Jung, W.; Lim, S.; Kim, S.; Choi, H.-G.; Han, S.J. Lipid production by arctic microalga Chlamydomonas sp. KNF0008 at low temperatures. Appl. Biochem. Biotechnol. 2019, 188, 326–337. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, D.P.K.; Silva, A.F.; Araujo, O.Q.; Chaloub, R.M. Impact of temperature and light intensity on triacylglycerol accumulation in marine microalgae. Biomass Bioenergy 2015, 72, 280–287. [Google Scholar] [CrossRef]
- Kim, E.J.; Kim, S.; Choi, H.-G.; Han, S.J. Co-production of biodiesel and bioethanol using psychrophilic microalga Chlamydomonas sp. KNM0029C isolated from Arctic sea ice. Biotechnol. Biofuels 2020, 13, 20. [Google Scholar] [CrossRef]
- Zheng, S.; Zou, S.; Feng, T.; Sun, S.; Guo, X.; He, M.; Wang, C.; Chen, H.; Wang, Q. Low temperature combined with high inoculum density improves alpha-linolenic acid production and biochemical characteristics of Chlamydomonas reinhardtii. Bioresour. Technol. 2022, 348, 126746. [Google Scholar] [CrossRef] [PubMed]
- Tsai, H.-P.; Chuang, L.-T.; Chen, C.-N.N. Production of long chain omega-3 fatty acids and carotenoids in tropical areas by a new heat-tolerant microalga Tetraselmis sp. DS3. Food Chem. 2016, 192, 682–690. [Google Scholar] [CrossRef] [PubMed]
- Çalişkan, E.; Haznedaroğlu, B.Z. Characterization of omega-3 and omega-6 fatty acid accumulation in Chlorococcum novae-angliae microalgae grown under various culture conditions. Int. J. Life. Sci. Biotechnol. 2022, 5, 346–369. [Google Scholar] [CrossRef]
- Pröschold, T.; Darienko, T.; Krienitz, L.; Coleman, A.W. Chlamydomonas schloesseri sp. nov. (Chlamydophyceae, Chlorophyta) revealed by morphology, autolysin cross experiments, and multiple gene analyses. Phytotaxa 2018, 362, 21–38. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef] [PubMed]
- Plackett, R.L.; Burman, J.P. The design of optimum multifactorial experiments. Biometrika 1946, 33, 305–325. [Google Scholar] [CrossRef]
- Ekpenyong, M.G.; Antai, S.P.; Asitok, A.D.; Ekpo, B.O. Plackett-Burman design and response surface optimization of medium trace nutrients for glycolipopeptide biosurfactant production. Iran. Biomed. J. 2017, 21, 249. [Google Scholar] [CrossRef] [PubMed]
- Box, G.E.; Behnken, D.W. Some new three level designs for the study of quantitative variables. Technometrics 1960, 2, 455–475. [Google Scholar] [CrossRef]
- Czyrski, A.; Sznura, J. The application of Box-Behnken-Design in the optimization of HPLC separation of fluoroquinolones. Sci. Rep. 2019, 9, 19458. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.J.; Jung, W.; Kim, S.; Choi, H.-G.; Han, S.J. Enhancing Lipid Production in the Arctic Microalga Chlamydomonas sp. KNM0029C by Using Statistical Optimization of Medium Components. KSBB J. 2019, 34, 173–177. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Component | −Value (g/L) | +Value (g/L) | Effect | T-Statistic | p-Value |
|---|---|---|---|---|---|---|
| X1 | Tris base | 0.484 | 4.84 | −30.58 | −5.54 | 0 |
| X2 | NH4Cl | 0.075 | 0.75 | −12.48 | −2.26 | 0.036 |
| X3 | MgSO4 | 0.02 | 0.2 | −65.72 | −11.91 | 0 |
| X4 | CaCl2 | 0.01 | 0.1 | 30.93 | 5.6 | 0 |
| X5 | Potassium phosphate | 0.0864 | 0.864 | 25.37 | 4.6 | 0 |
| X6 | Trace metals | 0.01954 | 0.1954 | −7.83 | −1.42 | 0.173 |
| X7 | AcOH | 0.2 (mL) | 2 (mL) | −67.82 | −12.29 | 0 |
| Variable | Component | −Value (g/L) | 0 Value (g/L) | Value (g/L) |
|---|---|---|---|---|
| X1 | Tris base | 0.242 | 1.331 | 2.42 |
| X2 | NH4Cl | 0.0375 | 0.20625 | 0.375 |
| X3 | MgSO4 | 0.01 | 0.055 | 0.1 |
| X4 | CaCl2 | 0.01 | 0.055 | 0.1 |
| X5 | Potassium phosphate | 0.0864 | 0.4752 | 0.864 |
| X6 | AcOH | 0.1 (mL) | 0.55 (mL) | 1 (mL) |
| Component | Normal TAP (g/L) | Optimized TAP (g/L) |
|---|---|---|
| Tris base | 2.42 | 2.42 |
| NH4Cl | 0.375 | 0.375 |
| MgSO4 | 0.1 | 0.1 |
| CaCl2 | 0.05 | 0.05 |
| Potassium phosphate | 0.432 | 0.864 |
| Trace metals | 0.097 | 0.097 |
| AcOH | 1 (mL) | 0.81 (mL) |
| Fatty Acid | Normal TAP | Optimized TAP | ||
|---|---|---|---|---|
| Content (%) | Yield (mg/g DCW) | Content (%) | Yield (mg/g DCW) | |
| C12:0 | 0.51 | 1.18 | 0.50 | 1.29 |
| C14:0 | 0.52 | 1.18 | 0.51 | 1.29 |
| C16:0 | 10.29 | 23.61 | 10.01 | 25.35 |
| C16:1n-7 | 7.06 | 16.18 | 6.15 | 15.58 |
| C16:2n-6 | 2.27 | 5.20 | 2.95 | 7.46 |
| C16:3n-6 | 2.45 | 5.63 | 2.59 | 6.56 |
| C16:4n-3 | 11.95 | 27.41 | 12.07 | 30.56 |
| C18:0 | 0.78 | 1.78 | 0.83 | 2.11 |
| C18:1n-9 | 0.65 | 1.49 | 0.64 | 1.62 |
| C18:2n-6 | 2.09 | 4.79 | 5.51 | 13.95 |
| C18:3n-6 | 3.71 | 8.52 | 4.04 | 10.22 |
| C18:3n-3 | 16.31 | 37.42 | 16.41 | 41.55 |
| C18:4n-3 | 10.44 | 23.95 | 10.10 | 25.58 |
| phytol | 1.12 | 2.56 | 1.67 | 4.23 |
| SFA | 12.19 | 27.76 | 11.85 | 30.04 |
| MUFA | 7.70 | 17.67 | 6.79 | 17.20 |
| PUFA | 49.23 | 112.91 | 53.66 | 135.89 |
| TFA | 69.12 | 158.35 | 72.31 | 183.13 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chae, H.; Kim, S.; Choi, H.-G.; Kim, J.H.; Han, S.J.; Kim, E.J. Arctic Sea Ice Microalga Chlamydomonas latifrons KNF0041: Identification and Statistical Optimization of Medium for Enhanced Biomass and Omega-3/Omega-6. Mar. Drugs 2023, 21, 454. https://doi.org/10.3390/md21080454
Chae H, Kim S, Choi H-G, Kim JH, Han SJ, Kim EJ. Arctic Sea Ice Microalga Chlamydomonas latifrons KNF0041: Identification and Statistical Optimization of Medium for Enhanced Biomass and Omega-3/Omega-6. Marine Drugs. 2023; 21(8):454. https://doi.org/10.3390/md21080454
Chicago/Turabian StyleChae, Hyunsik, Sanghee Kim, Han-Gu Choi, Ji Hee Kim, Se Jong Han, and Eun Jae Kim. 2023. "Arctic Sea Ice Microalga Chlamydomonas latifrons KNF0041: Identification and Statistical Optimization of Medium for Enhanced Biomass and Omega-3/Omega-6" Marine Drugs 21, no. 8: 454. https://doi.org/10.3390/md21080454
APA StyleChae, H., Kim, S., Choi, H.-G., Kim, J. H., Han, S. J., & Kim, E. J. (2023). Arctic Sea Ice Microalga Chlamydomonas latifrons KNF0041: Identification and Statistical Optimization of Medium for Enhanced Biomass and Omega-3/Omega-6. Marine Drugs, 21(8), 454. https://doi.org/10.3390/md21080454