
Ten Years of Research on Fucoidan and Cancer: Focus on Its Antiangiogenic and Antimetastatic Effects



Abstract
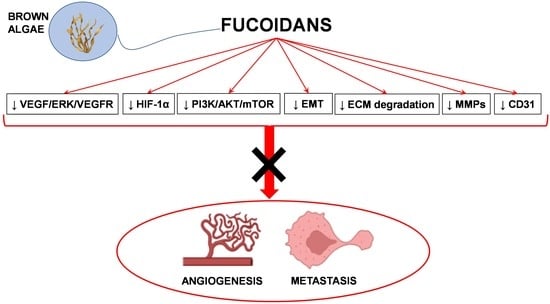
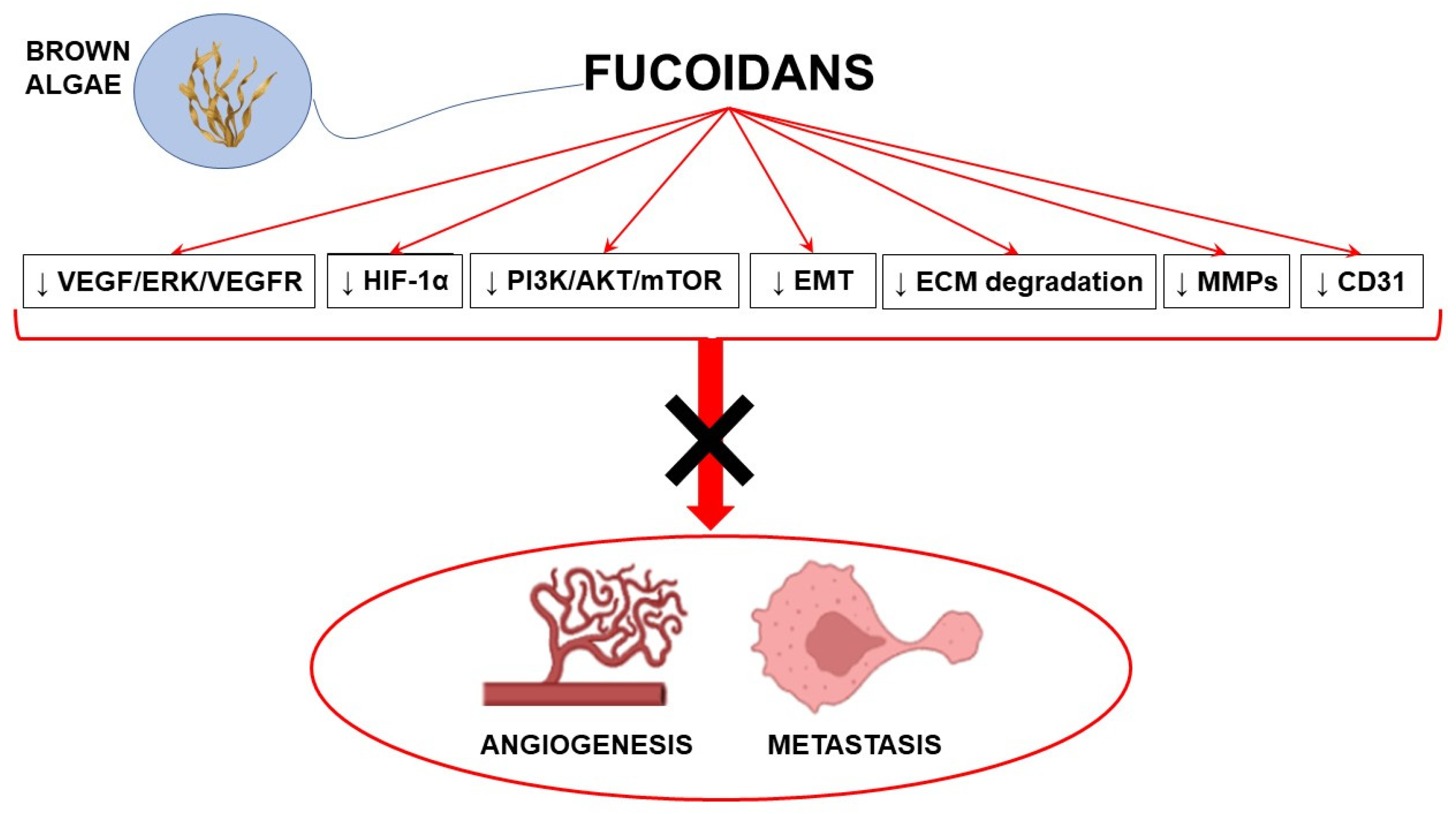
1. Introduction
2. Antiangiogenic and Antimigratory Effects of Fucoidans
2.1. Antiangiogenic and Antimigratory Activity of Fucoidans In Vitro
2.1.1. Cancer Cell Lines
2.1.2. Endothelial Cells
2.2. Antiangiogenic Effects of Fucoidans in Chick Embryo Chorioallantoic Membrane (CAM) Model
2.3. Antiangiogenic and Antimetastatic Effects of Fucoidans in Murine In Vivo Models
2.4. Antiangiogenic and Antimetastatic Effects of Fucoidans in the Zebrafish Model
3. Clinical Studies on Fucoidans
4. Pharmacokinetics of Fucoidans
5. Challenges and Future Perspectives
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mattiuzzi, C.; Lippi, G. Current Cancer Epidemiology. J. Epidemiol. Glob. Health 2019, 9, 217–222. [Google Scholar] [CrossRef]
- Zhao, Y.; Adjei, A.A. Targeting Angiogenesis in Cancer Therapy: Moving Beyond Vascular Endothelial Growth Factor. Oncologist 2015, 20, 660–673. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Jayson, G.C.; Kerbel, R.; Ellis, L.M.; Harris, A.L. Antiangiogenic Therapy in Oncology: Current Status and Future Directions. Lancet 2016, 388, 518–529. [Google Scholar] [CrossRef]
- Catanzaro, E.; Greco, G.; Potenza, L.; Calcabrini, C.; Fimognari, C. Natural Products to Fight Cancer: A Focus on Juglans Regia. Toxins 2018, 10, 469. [Google Scholar] [CrossRef]
- Lenzi, M.; Cocchi, V.; Novaković, A.; Karaman, M.; Sakač, M.; Mandić, A.; Pojić, M.; Barbalace, M.C.; Angeloni, C.; Hrelia, P.; et al. Meripilus Giganteus Ethanolic Extract Exhibits Pro-Apoptotic and Anti-Proliferative Effects in Leukemic Cell Lines. BMC Complement. Altern. Med. 2018, 18, 300. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, M.K.; Warrier, S.; Kumar, A.P.; Sethi, G.; Arfuso, F. Potential Role of Natural Compounds as Anti-Angiogenic Agents in Cancer. Curr. Vasc. Pharmacol. 2017, 15, 503–519. [Google Scholar] [CrossRef]
- Turrini, E.; Maffei, F.; Fimognari, C. Effect of the Marine Polyketide Plocabulin on Tumor Progression. Mar. Drugs 2022, 21, 38. [Google Scholar] [CrossRef] [PubMed]
- Ai, Y.; Zhao, Z.; Wang, H.; Zhang, X.; Qin, W.; Guo, Y.; Zhao, M.; Tang, J.; Ma, X.; Zeng, J. Pull the Plug: Anti-Angiogenesis Potential of Natural Products in Gastrointestinal Cancer Therapy. Phytother. Res. PTR 2022, 36, 3371–3393. [Google Scholar] [CrossRef]
- Rashidi, B.; Malekzadeh, M.; Goodarzi, M.; Masoudifar, A.; Mirzaei, H. Green Tea and Its Anti-Angiogenesis Effects. Biomed. Pharmacother. Biomed. Pharmacother. 2017, 89, 949–956. [Google Scholar] [CrossRef]
- Li, R.; Song, X.; Guo, Y.; Song, P.; Duan, D.; Chen, Z.-S. Natural Products: A Promising Therapeutics for Targeting Tumor Angiogenesis. Front. Oncol. 2021, 11, 772915. [Google Scholar] [CrossRef] [PubMed]
- Yao, W.; Qiu, H.-M.; Cheong, K.-L.; Zhong, S. Advances in Anti-Cancer Effects and Underlying Mechanisms of Marine Algae Polysaccharides. Int. J. Biol. Macromol. 2022, 221, 472–485. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Sun, J.; Su, X.; Yu, Q.; Yu, Q.; Zhang, P. A Review about the Development of Fucoidan in Antitumor Activity: Progress and Challenges. Carbohydr. Polym. 2016, 154, 96–111. [Google Scholar] [CrossRef]
- Vo, T.-S.; Kim, S.-K. Fucoidans as a Natural Bioactive Ingredient for Functional Foods. J. Funct. Foods 2013, 5, 16–27. [Google Scholar] [CrossRef]
- Yang, J.-Y.; Lim, S.Y. Fucoidans and Bowel Health. Mar. Drugs 2021, 19, 436. [Google Scholar] [CrossRef]
- Atashrazm, F.; Lowenthal, R.M.; Woods, G.M.; Holloway, A.F.; Dickinson, J.L. Fucoidan and Cancer: A Multifunctional Molecule with Anti-Tumor Potential. Mar. Drugs 2015, 13, 2327–2346. [Google Scholar] [CrossRef]
- Wang, Y.; Xing, M.; Cao, Q.; Ji, A.; Liang, H.; Song, S. Biological Activities of Fucoidan and the Factors Mediating Its Therapeutic Effects: A Review of Recent Studies. Mar. Drugs 2019, 17, 183. [Google Scholar] [CrossRef]
- Dörschmann, P.; Seeba, C.; Thalenhorst, T.; Roider, J.; Klettner, A. Anti-Inflammatory Properties of Antiangiogenic Fucoidan in Retinal Pigment Epithelium Cells. Heliyon 2023, 9, e15202. [Google Scholar] [CrossRef]
- Khosravifar, M.; Sajadimajd, S.; Bahrami, G. Anti-Diabetic Effects of Macronutrients via Modulation of Angiogenesis: A Comprehensive Review on Carbohydrates and Proteins. Curr. Mol. Med. 2023, 23, 250–265. [Google Scholar] [CrossRef]
- Lu, Y.; Zhu, X.; Hu, C.; Li, P.; Zhao, M.; Lu, J.; Xia, G. A Fucoidan-Gelatin Wound Dressing Accelerates Wound Healing by Enhancing Antibacterial and Anti-Inflammatory Activities. Int. J. Biol. Macromol. 2022, 223, 36–48. [Google Scholar] [CrossRef]
- Bae, H.; Lee, J.-Y.; Yang, C.; Song, G.; Lim, W. Fucoidan Derived from Fucus Vesiculosus Inhibits the Development of Human Ovarian Cancer via the Disturbance of Calcium Homeostasis, Endoplasmic Reticulum Stress, and Angiogenesis. Mar. Drugs 2020, 18, 45. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.-Y.; Li, L.-Z.; Xue, K.-C.; Hu, D.-D.; Gao, Y.-J. Antitumor Activity of Fucoidan in Anaplastic Thyroid Cancer via Apoptosis and Anti-Angiogenesis. Mol. Med. Rep. 2017, 15, 2620–2624. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.; Granja, S.; Neves, N.M.; Reis, R.L.; Baltazar, F.; Silva, T.H.; Martins, A. Fucoidan from Fucus Vesiculosus Inhibits New Blood Vessel Formation and Breast Tumor Growth in Vivo. Carbohydr. Polym. 2019, 223, 115034. [Google Scholar] [CrossRef]
- Wang, F.; Schmidt, H.; Pavleska, D.; Wermann, T.; Seekamp, A.; Fuchs, S. Crude Fucoidan Extracts Impair Angiogenesis in Models Relevant for Bone Regeneration and Osteosarcoma via Reduction of VEGF and SDF-1. Mar. Drugs 2017, 15, 186. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Luo, G.; Xiao, Q.; Chen, L.; Luo, X.; Lv, J.; Chen, L. Fucoidan Inhibits Angiogenesis Induced by Multiple Myeloma Cells. Oncol. Rep. 2016, 36, 1963–1972. [Google Scholar] [CrossRef] [PubMed]
- Yun, C.W.; Yun, S.; Lee, J.H.; Han, Y.-S.; Yoon, Y.M.; An, D.; Lee, S.H. Silencing Prion Protein in HT29 Human Colorectal Cancer Cells Enhances Anticancer Response to Fucoidan. Anticancer Res. 2016, 36, 4449–4458. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.-S.; Lee, J.H.; Lee, S.H. Antitumor Effects of Fucoidan on Human Colon Cancer Cells via Activation of Akt Signaling. Biomol. Ther. 2015, 23, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.-H.; Chiu, Y.-H.; Chan, Y.-L.; Chiu, Y.-H.; Wang, H.; Huang, K.-C.; Li, T.-L.; Hsu, K.-H.; Wu, C.-J. Prophylactic Administration of Fucoidan Represses Cancer Metastasis by Inhibiting Vascular Endothelial Growth Factor (VEGF) and Matrix Metalloproteinases (MMPs) in Lewis Tumor-Bearing Mice. Mar. Drugs 2015, 13, 1882–1900. [Google Scholar] [CrossRef]
- Xue, M.; Ge, Y.; Zhang, J.; Wang, Q.; Hou, L.; Liu, Y.; Sun, L.; Li, Q. Anticancer Properties and Mechanisms of Fucoidan on Mouse Breast Cancer in Vitro and in Vivo. PLoS ONE 2012, 7, e43483. [Google Scholar] [CrossRef]
- Li, W.; Xue, D.; Xue, M.; Zhao, J.; Liang, H.; Liu, Y.; Sun, T. Fucoidan Inhibits Epithelial-to-Mesenchymal Transition via Regulation of the HIF-1α Pathway in Mammary Cancer Cells under Hypoxia. Oncol. Lett. 2019, 18, 330–338. [Google Scholar] [CrossRef]
- Rui, X.; Pan, H.-F.; Shao, S.-L.; Xu, X.-M. Anti-Tumor and Anti-Angiogenic Effects of Fucoidan on Prostate Cancer: Possible JAK-STAT3 Pathway. BMC Complement. Altern. Med. 2017, 17, 378. [Google Scholar] [CrossRef] [PubMed]
- Hsu, W.-J.; Lin, M.-H.; Kuo, T.-C.; Chou, C.-M.; Mi, F.-L.; Cheng, C.-H.; Lin, C.-W. Fucoidan from Laminaria Japonica Exerts Antitumor Effects on Angiogenesis and Micrometastasis in Triple-Negative Breast Cancer Cells. Int. J. Biol. Macromol. 2020, 149, 600–608. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.-D.; Yao, C.-J.; Chow, J.-M.; Chang, C.-L.; Hwang, P.-A.; Chuang, S.-E.; Whang-Peng, J.; Lai, G.-M. Fucoidan Elevates MicroRNA-29b to Regulate DNMT3B-MTSS1 Axis and Inhibit EMT in Human Hepatocellular Carcinoma Cells. Mar. Drugs 2015, 13, 6099–6116. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.-C.; Hsu, W.-L.; Hwang, P.-A.; Chou, T.-C. Low Molecular Weight Fucoidan Inhibits Tumor Angiogenesis through Downregulation of HIF-1/VEGF Signaling under Hypoxia. Mar. Drugs 2015, 13, 4436–4451. [Google Scholar] [CrossRef]
- Pan, T.-J.; Li, L.-X.; Zhang, J.-W.; Yang, Z.-S.; Shi, D.-M.; Yang, Y.-K.; Wu, W.-Z. Antimetastatic Effect of Fucoidan-Sargassum against Liver Cancer Cell Invadopodia Formation via Targeting Integrin AVβ3 and Mediating AVβ3/Src/E2F1 Signaling. J. Cancer 2019, 10, 4777–4792. [Google Scholar] [CrossRef]
- Chen, H.; Cong, Q.; Du, Z.; Liao, W.; Zhang, L.; Yao, Y.; Ding, K. Sulfated Fucoidan FP08S2 Inhibits Lung Cancer Cell Growth in Vivo by Disrupting Angiogenesis via Targeting VEGFR2/VEGF and Blocking VEGFR2/Erk/VEGF Signaling. Cancer Lett. 2016, 382, 44–52. [Google Scholar] [CrossRef]
- Cong, Q.; Chen, H.; Liao, W.; Xiao, F.; Wang, P.; Qin, Y.; Dong, Q.; Ding, K. Structural Characterization and Effect on Anti-Angiogenic Activity of a Fucoidan from Sargassum Fusiforme. Carbohydr. Polym. 2016, 136, 899–907. [Google Scholar] [CrossRef]
- Delma, C.R.; Somasundaram, S.T.; Srinivasan, G.P.; Khursheed, M.; Bashyam, M.D.; Aravindan, N. Fucoidan from Turbinaria Conoides: A Multifaceted “deliverable” to Combat Pancreatic Cancer Progression. Int. J. Biol. Macromol. 2015, 74, 447–457. [Google Scholar] [CrossRef]
- Teng, H.; Yang, Y.; Wei, H.; Liu, Z.; Liu, Z.; Ma, Y.; Gao, Z.; Hou, L.; Zou, X. Fucoidan Suppresses Hypoxia-Induced Lymphangiogenesis and Lymphatic Metastasis in Mouse Hepatocarcinoma. Mar. Drugs 2015, 13, 3514–3530. [Google Scholar] [CrossRef]
- Yang, Y.; Gao, Z.; Ma, Y.; Teng, H.; Liu, Z.; Wei, H.; Lu, Y.; Cheng, X.; Hou, L.; Zou, X. Fucoidan Inhibits Lymphangiogenesis by Downregulating the Expression of VEGFR3 and PROX1 in Human Lymphatic Endothelial Cells. Oncotarget 2016, 7, 38025–38035. [Google Scholar] [CrossRef]
- Chen, X.; Sun, L.; Wei, X.; Lu, H.; Tan, Y.; Sun, Z.; Jiang, J. Antitumor Effect and Molecular Mechanism of Fucoidan in NSCLC. BMC Complement. Med. Ther. 2021, 21, 25. [Google Scholar] [CrossRef] [PubMed]
- Hou, C.-Y.; Ma, C.-Y.; Lin, Y.-J.; Huang, C.-L.; Wang, H.-D.; Yuh, C.-H. WNK1-OSR1 Signaling Regulates Angiogenesis-Mediated Metastasis towards Developing a Combinatorial Anti-Cancer Strategy. Int. J. Mol. Sci. 2022, 23, 12100. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.-W.; Jung, W.; Choi, C.; Kim, S.-Y.; Son, A.; Kim, H.; Lee, N.; Park, H.C. Fucoidan-Manganese Dioxide Nanoparticles Potentiate Radiation Therapy by Co-Targeting Tumor Hypoxia and Angiogenesis. Mar. Drugs 2018, 16, 510. [Google Scholar] [CrossRef] [PubMed]
- Staton, C.A.; Stribbling, S.M.; Tazzyman, S.; Hughes, R.; Brown, N.J.; Lewis, C.E. Current Methods for Assaying Angiogenesis in Vitro and in Vivo. Int. J. Exp. Pathol. 2004, 85, 233–248. [Google Scholar] [CrossRef] [PubMed]
- Agani, F.; Jiang, B.-H. Oxygen-Independent Regulation of HIF-1: Novel Involvement of PI3K/AKT/MTOR Pathway in Cancer. Curr. Cancer Drug Targets 2013, 13, 245–251. [Google Scholar] [CrossRef]
- Martin, T.A.; Ye, L.; Sanders, A.J.; Lane, J.; Jiang, W.G. Cancer Invasion and Metastasis: Molecular and Cellular Perspective; Landes Bioscience: Austin, TX, USA, 2013. [Google Scholar]
- Zhang, Y.; Weinberg, R.A. Epithelial-to-Mesenchymal Transition in Cancer: Complexity and Opportunities. Front. Med. 2018, 12, 361–373. [Google Scholar] [CrossRef]
- Xu, X.; Tao, Y.; Shan, L.; Chen, R.; Jiang, H.; Qian, Z.; Cai, F.; Ma, L.; Yu, Y. The Role of MicroRNAs in Hepatocellular Carcinoma. J. Cancer 2018, 9, 3557–3569. [Google Scholar] [CrossRef]
- Garzon, R.; Liu, S.; Fabbri, M.; Liu, Z.; Heaphy, C.E.A.; Callegari, E.; Schwind, S.; Pang, J.; Yu, J.; Muthusamy, N.; et al. MicroRNA-29b Induces Global DNA Hypomethylation and Tumor Suppressor Gene Reexpression in Acute Myeloid Leukemia by Targeting Directly DNMT3A and 3B and Indirectly DNMT1. Blood 2009, 113, 6411–6418. [Google Scholar] [CrossRef]
- Hsu, H.-Y.; Lin, T.-Y.; Hwang, P.-A.; Tseng, L.-M.; Chen, R.-H.; Tsao, S.-M.; Hsu, J. Fucoidan Induces Changes in the Epithelial to Mesenchymal Transition and Decreases Metastasis by Enhancing Ubiquitin-Dependent TGFβ Receptor Degradation in Breast Cancer. Carcinogenesis 2013, 34, 874–884. [Google Scholar] [CrossRef]
- Augoff, K.; Hryniewicz-Jankowska, A.; Tabola, R. Invadopodia: Clearing the Way for Cancer Cell Invasion. Ann. Transl. Med. 2020, 8, 902. [Google Scholar] [CrossRef]
- Ribatti, D. The Chick Embryo Chorioallantoic Membrane as an Experimental Model to Study in Vivo Angiogenesis in Glioblastoma Multiforme. Brain Res. Bull. 2022, 182, 26–29. [Google Scholar] [CrossRef] [PubMed]
- Yousaf, S.; Ahmad, M.; Wu, S.; Zia, M.A.; Ahmed, I.; Iqbal, H.M.N.; Liu, Q.; Rehman, S.U. Cellular Prion Protein Role in Cancer Biology: Is It A Potential Therapeutic Target? Biomedicines 2022, 10, 2833. [Google Scholar] [CrossRef] [PubMed]
- Fonsatti, E.; Sigalotti, L.; Arslan, P.; Altomonte, M.; Maio, M. Emerging Role of Endoglin (CD105) as a Marker of Angiogenesis with Clinical Potential in Human Malignancies. Curr. Cancer Drug Targets 2003, 3, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Canesin, G.; Krzyzanowska, A.; Hellsten, R.; Bjartell, A. Cytokines and Janus Kinase/Signal Transducer and Activator of Transcription Signaling in Prostate Cancer: Overview and Therapeutic Opportunities. Curr. Opin. Endocr. Metab. Res. 2020, 10, 36–42. [Google Scholar] [CrossRef]
- Zhu, C.; Cao, R.; Zhang, S.-X.; Man, Y.-N.; Wu, X.-Z. Fucoidan Inhibits the Growth of Hepatocellular Carcinoma Independent of Angiogenesis. Evid.-Based Complement. Altern. Med. ECAM 2013, 2013, 692549. [Google Scholar] [CrossRef] [PubMed]
- Jayatilleke, K.M.; Hulett, M.D. Heparanase and the Hallmarks of Cancer. J. Transl. Med. 2020, 18, 453. [Google Scholar] [CrossRef]
- Abdollah, M.R.A.; Ali, A.A.; Elgohary, H.H.; Elmazar, M.M. Antiangiogenic Drugs in Combination with Seaweed Fucoidan: A Mechanistic in Vitro and in Vivo Study Exploring the VEGF Receptor and Its Downstream Signaling Molecules in Hepatic Cancer. Front. Pharmacol. 2023, 14, 1108992. [Google Scholar] [CrossRef]
- Anisimova, N.Y.; Ustyuzhanina, N.E.; Bilan, M.I.; Morozevich, G.E.; Usov, A.I.; Nifantiev, N.E.; Kiselevskiy, M.V. Anti-Angiogenic Properties of Sulfated Polysaccharides Fucoidans and Their Analogs. Russ. Chem. Bull. 2022, 71, 2505–2514. [Google Scholar] [CrossRef]
- Ustyuzhanina, N.E.; Bilan, M.I.; Ushakova, N.A.; Usov, A.I.; Kiselevskiy, M.V.; Nifantiev, N.E. Fucoidans: Pro- or Antiangiogenic Agents? Glycobiology 2014, 24, 1265–1274. [Google Scholar] [CrossRef]
- Manivasagan, P.; Bharathiraja, S.; Bui, N.Q.; Jang, B.; Oh, Y.-O.; Lim, I.G.; Oh, J. Doxorubicin-Loaded Fucoidan Capped Gold Nanoparticles for Drug Delivery and Photoacoustic Imaging. Int. J. Biol. Macromol. 2016, 91, 578–588. [Google Scholar] [CrossRef]
- Fernandes, A.S.; Oliveira, C.; Reis, R.L.; Martins, A.; Silva, T.H. Marine-Inspired Drugs and Biomaterials in the Perspective of Pancreatic Cancer Therapies. Mar. Drugs 2022, 20, 689. [Google Scholar] [CrossRef] [PubMed]
- Tobia, C.; De Sena, G.; Presta, M. Zebrafish Embryo, a Tool to Study Tumor Angiogenesis. Int. J. Dev. Biol. 2011, 55, 505–509. [Google Scholar] [CrossRef]
- Tulotta, C.; He, S.; van der Ent, W.; Chen, L.; Groenewoud, A.; Spaink, H.P.; Snaar-Jagalska, B.E. Imaging Cancer Angiogenesis and Metastasis in a Zebrafish Embryo Model. Adv. Exp. Med. Biol. 2016, 916, 239–263. [Google Scholar] [CrossRef] [PubMed]
- Tsai, H.-L.; Tai, C.-J.; Huang, C.-W.; Chang, F.-R.; Wang, J.-Y. Efficacy of Low-Molecular-Weight Fucoidan as a Supplemental Therapy in Metastatic Colorectal Cancer Patients: A Double-Blind Randomized Controlled Trial. Mar. Drugs 2017, 15, 122. [Google Scholar] [CrossRef] [PubMed]
- Coussens, L.M.; Zitvoge, L.; Palucka, A.K. Neutralizing Tumor-Promoting Chronic Inflammation: A Magic Bullet? Science 2013, 339, 286–291. [Google Scholar] [CrossRef]
- Crusz, S.M.; Balkwill, F.R. Inflammation and Cancer: Advances and New Agents. Nat. Rev. Clin. Oncol. 2015, 12, 584–596. [Google Scholar] [CrossRef]
- Takahashi, H.; Kawaguchi, M.; Kitamura, K.; Narumiya, S.; Kawamura, M.; Tengan, I.; Nishimoto, S.; Hanamure, Y.; Majima, Y.; Tsubura, S.; et al. An Exploratory Study on the Anti-Inflammatory Effects of Fucoidan in Relation to Quality of Life in Advanced Cancer Patients. Integr. Cancer Ther. 2018, 17, 282–291. [Google Scholar] [CrossRef]
- Romani, A.M.P. Cisplatin in Cancer Treatment. Biochem. Pharmacol. 2022, 206, 115323. [Google Scholar] [CrossRef]
- Ikeguchi, M.; Saito, H.; Miki, Y.; Kimura, T. Effect of Fucoidan Dietary Supplement on the Chemotherapy Treatment of Patients with Unresectable Advanced Gastric Cancer. J. Cancer Ther. 2015, 6, 1020. [Google Scholar] [CrossRef]
- Tocaciu, S.; Oliver, L.J.; Lowenthal, R.M.; Peterson, G.M.; Patel, R.; Shastri, M.; McGuinness, G.; Olesen, I.; Fitton, J.H. The Effect of Undaria Pinnatifida Fucoidan on the Pharmacokinetics of Letrozole and Tamoxifen in Patients With Breast Cancer. Integr. Cancer Ther. 2018, 17, 99–105. [Google Scholar] [CrossRef]
- Shikov, A.N.; Flisyuk, E.V.; Obluchinskaya, E.D.; Pozharitskaya, O.N. Pharmacokinetics of Marine-Derived Drugs. Mar. Drugs 2020, 18, 557. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Sun, D.; Zhao, X.; Jin, W.; Wang, J.; Zhang, Q. Microanalysis and Preliminary Pharmacokinetic Studies of a Sulfated Polysaccharide from Laminaria Japonica. Chin. J. Oceanol. Limnol. 2016, 34, 177–185. [Google Scholar] [CrossRef]
- Zhao, X.; Guo, F.; Hu, J.; Zhang, L.; Xue, C.; Zhang, Z.; Li, B. Antithrombotic Activity of Oral Administered Low Molecular Weight Fucoidan from Laminaria Japonica. Thromb. Res. 2016, 144, 46–52. [Google Scholar] [CrossRef]
- Pozharitskaya, O.N.; Shikov, A.N.; Faustova, N.M.; Obluchinskaya, E.D.; Kosman, V.M.; Vuorela, H.; Makarov, V.G. Pharmacokinetic and Tissue Distribution of Fucoidan from Fucus Vesiculosus after Oral Administration to Rats. Mar. Drugs 2018, 16, 132. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Zhang, E.; Hu, B.; Liang, H.; Song, S.; Ji, A. Study on Absorption Mechanism and Tissue Distribution of Fucoidan. Molecules 2020, 25, 1087. [Google Scholar] [CrossRef]
- Nagamine, T.; Nakazato, K.; Tomioka, S.; Iha, M.; Nakajima, K. Intestinal Absorption of Fucoidan Extracted from the Brown Seaweed, Cladosiphon Okamuranus. Mar. Drugs 2014, 13, 48–64. [Google Scholar] [CrossRef]
- Tokita, Y.; Hirayama, M.; Nakajima, K.; Tamaki, K.; Iha, M.; Nagamine, T. Detection of Fucoidan in Urine after Oral Intake of Traditional Japanese Seaweed, Okinawa Mozuku (Cladosiphon Okamuranus Tokida). J. Nutr. Sci. Vitaminol. 2017, 63, 419–421. [Google Scholar] [CrossRef]
- Kadena, K.; Tomori, M.; Iha, M.; Nagamine, T. Absorption Study of Mozuku Fucoidan in Japanese Volunteers. Mar. Drugs 2018, 16, 254. [Google Scholar] [CrossRef]
- Lin, Y.; Qi, X.; Liu, H.; Xue, K.; Xu, S.; Tian, Z. The Anti-Cancer Effects of Fucoidan: A Review of Both in Vivo and in Vitro Investigations. Cancer Cell Int. 2020, 20, 154. [Google Scholar] [CrossRef]
- Jin, J.-O.; Yadav, D.; Madhwani, K.; Puranik, N.; Chavda, V.; Song, M. Seaweeds in the Oncology Arena: Anti-Cancer Potential of Fucoidan as a Drug-A Review. Molecules 2022, 27, 6032. [Google Scholar] [CrossRef]
- Citkowska, A.; Szekalska, M.; Winnicka, K. Possibilities of Fucoidan Utilization in the Development of Pharmaceutical Dosage Forms. Mar. Drugs 2019, 17, 458. [Google Scholar] [CrossRef] [PubMed]
- Luthuli, S.; Wu, S.; Cheng, Y.; Zheng, X.; Wu, M.; Tong, H. Therapeutic Effects of Fucoidan: A Review on Recent Studies. Mar. Drugs 2019, 17, 487. [Google Scholar] [CrossRef] [PubMed]
- Gideon, T.P.; Rengasamy, R. Toxicological Evaluation of Fucoidan from Cladosiphon Okamuranus. J. Med. Food 2008, 11, 638–642. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Zhang, Q.; Song, J. Toxicological Evaluation of Fucoidan Extracted from Laminaria Japonica in Wistar Rats. Food Chem. Toxicol. 2005, 43, 421–426. [Google Scholar] [CrossRef]
- Chung, H.-J.; Jeun, J.; Houng, S.-J.; Jun, H.-J.; Kweon, D.-K.; Lee, S.-J. Toxicological Evaluation of Fucoidan from Undaria Pinnatifidain Vitro and in Vivo. Phytother. Res. PTR 2010, 24, 1078–1083. [Google Scholar] [CrossRef]
- Myers, S.P.; Mulder, A.M.; Baker, D.G.; Robinson, S.R.; Rolfe, M.I.; Brooks, L.; Fitton, J.H. Effects of Fucoidan from Fucus Vesiculosus in Reducing Symptoms of Osteoarthritis: A Randomized Placebo-Controlled Trial. Biol. Targets Ther. 2016, 10, 81–88. [Google Scholar] [CrossRef]
- Imbs, T.I.; Zvyagintseva, T.N.; Ermakova, S.P. Is the Transformation of Fucoidans in Human Body Possible? Int. J. Biol. Macromol. 2020, 142, 778–781. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Fucoidan | Cancer Model | Experimental Model | Antiangiogenic and Antimetastatic Mechanisms | References |
|---|---|---|---|---|
| Fucoidan (from Fucus vesiculosus) (Sigma Aldrich, St. Louis, MO, USA) | Human ovarian cancer | Vitro: ES-2 and OV-90 cell lines (300 µg/mL for 24/48 h) Vivo: Embryos of transgenic zebrafish Tg(fli1:EGFP) (300 µg/mL for 48 h) or zebrafish xenograft model injected with fucoidan pre-treated ES-2 and OV-90 cells | ↓ mRNA expression of VEGFs (VEGFA-VEGFD), FLT4, KDR ↓ VEGFA, VEGFC, FLT1, FLT4, KDR, KDRL | [21] |
| Fucoidan (from Fucus vesiculosus) | Human anaplastic thyroid cancer | Vitro: FTC133 cell line (10 µM) | ↓ HIF-1α and VEGF in hypoxic-like conditions ↓ Tube formation and the migration of FTC133 cells | [22] |
| Crude fucoidan (from Fucus vesiculosus) (Sigma Aldrich) | Human breast cancer | Vitro: MDA-MB-231 cell line, HPMEC-ST1.6R cells (500 µg/mL crude fucoidan for 4, 8, 12 or 16 h) Vivo: CAM without tumor (3 or 7 days after 0.5 mg/mL fucoidan injection) or CAM with tumor MDA-MB-231 cells onplantation (4 or 8 days after 0.5 mg/mL fucoidan injection) | ↓ PDGF ↓ Migration and formation of new vessel ↓ Blood vessel maturation Disruption of pre-formed tubular-like structures ↓ Blood vessel stability (as demonstrated by the reduction in lectin expression) | [23] |
| Crude fucoidan (from Fucus vesiculosus) (Sigma Aldrich) | Human osteosarcoma | Vitro: OECs in co-culture with MSCs or MG63 (100–500 μg/mL) | ↓ VEGF and SDF-1/CXCR4, collagen type 1 and angiopoietin-1 ↓ angiopoietin-2 | [24] |
| Fucoidan (from Fucus vesiculosus) (Sigma Aldrich) | Human multiple myeloma | Vitro: RPMI-8226 and U266 cell lines (25–200 μg/mL for 72 h) HUVECs (incubated with conditioned medium from fucoidan-pretreated RPMI-8226 or U266 cells for 6 h) Vivo: Multiple myeloma cells xenograft NOD/SCID female mice (intraperitoneal injection with 10 or 50 mg/kg fucoidan every two days for 3 weeks) | ↓ VEGF and HIF-1α both in hypoxic and normoxic conditions ↓ Tube structures and HUVECs migration ↓ p-AKT and p-ERK1/2 ↓ Angiogenesis and CD34+ vessels ↓ Microvessels density | [25] |
| Fucoidan (from Fucus vesiculosus) (Sigma Aldrich) | Human colon cancer | Vitro: Prion silenced HT29 cell line (200 μg/mL fucoidan for 48 h) Vivo: HT29 cancer cells xenograft Balb/c mice (5 mg/kg fucoidan intraperitoneally injected every 2 days for a total of nine administrations) | ↓ Angiogenesis; ↓ Migration ↓ Angiogenesis, VEGF and CD31 | [26] |
| Fucoidan (from Fucus vesiculosus) (Sigma Aldrich) | Human colon cancer | Vivo: HT29 cancer cells xenograft male nude mice (5 or 10 mg/kg fucoidan: intraperitoneally injected every 2 days for 30 days) | ↓ Angiogenesis ↓ VEGF and CD31 | [27] |
| Fucoidan (from Fucus vesiculosus) (Sigma Aldrich) | Murine Lewis lung carcinoma | Vitro: Murine Lewis lung adenocarcinoma cell line (50–400 µg/mL for 24 h) Vivo: Lewis lung carcinoma xenograft male C57BL/6 mice (1 or 3 mg/day by intragastric gavage 7 days prior to tumor inoculation) | ↓ Migration ↓ MMPs ↓ Angiogenesis and metastasis ↓ NF-κB ↓ VEGF ↓ MMPs | [28] |
| Crude Fucoidan (from Fucus vesiculosus) (Sigma Aldrich) | Murine breast cancer | Vitro: 4T1 cancer cells (50–200 µg/mL for 48 h) Vivo: 4T1 cancer cells xenograft BALB/c mice (5 or 10 mg/kg b.w. fucoidan intraperitoneally injected every 2 days for 20 days) | ↓ VEGF in vitro and in vivo ↓ Tube formation in vivo ↓ Angiogenesis in vivo ↓ Lung metastases | [29] |
| Fucoidan (source not specified) (Sigma Aldrich) | Human breast cancer | Vitro: MDA-MB-231 cell line (6.25–25 μg/mL for 48 h) | ↓ Invasion and migration of MDA-MB-231 cells ↓ HIF-1α and HIF-1 target genes (TWIST, Snail, CAIX and GLUT-1) in hypoxic conditions ↓ N-cadherin and vimentin and ↑ E-cadherin and ZO-1 ↓ EMT | [30] |
| Fucoidan (source not specified) (Sigma Aldrich) | Human prostate cancer | Vitro: DU-145 cell line (100–1000 μg/mL) Vivo: DU-145 cancer cells xenograft athymic nude mice (20 mg/kg b.w. of fucoidan for 28 days by oral gavage) | ↓ Angiogenesis and cell migration ↓ Hemoglobin content ↓ mRNA expression of CD31 and CD105 in tumor tissue ↓ JAK and STAT3 pathway ↓ Tube formation | [31] |
| Fucoidan (from Laminaria japonica) | Human triple-negative breast cancer | Vitro: MDA-MB-231 and HCC1806 cell line (125–2000 µg/mL fucoidan for 24/48 h) HUVECs (100–500 µg/mL for 18 h or incubated with conditioned medium from fucoidan-pretreated MDA-MB-231 and HCC1806 cells for 24 h) Vivo: Embryos of transgenic zebrafish Tg(fli1:EGFP) (0.1–2 mg/mL fucoidan for 48 h) or zebrafish xenograft model injected with fucoidan pre-treated GFP-expressing MDA-MB-231 | ↓ VEGFA, IGF-I, bFGF, MMP-2, and MMP-9 ↓ Migration and invasion in HUVEC and cancer cells ↓ Angiogenesis and metastatic capability in vivo | [32] |
| Fucoidan (from Sargassum hemiphyllum) (Hi-Q Marine Biotech International Ltd., Taipei, China) | Human hepatocellular carcinoma | Vitro: SK-Hep1 and HepG2 cell lines (200 µg/mL for 24 or 48 h) | ↓ TGF-signaling Regulation of miR-29b-DNMT3B-MTSS1 axis ↓ Invasion and metastasis ↓ EMT (↓ N-cadherin; ↑ E-cadherin) ↓ ECM degradation (↓ MMPs; ↑ TIMP) | [33] |
| Low molecular weight flucoidan (LMWF) 760 Da (from Sargassum hemiphyllum) (Hi-Q Marine Biotech International Ltd.) | Human bladder cancer cells | Vitro: T24 cell line (25–100 µg/mL) HUVECs (25–100 µg/mL) Vivo: T24 cancer cells xenograft BALB/c nude mice (80, 160 or 300 mg/kg b.w. per day LMWF orally administered for 30 days) | ↓ VEGF and HIF-1α ↓ PI3K/AKT/mTOR/p70S6K/4EBP-1 ↓ Migration and invasion of T24 cells in hypoxic conditions ↓ Tube formation in HUVECs in hypoxic conditions ↓ Tumor angiogenesis in vivo | [34] |
| Fucoidan (from Sargassum fusiforme) | Human hepatocellular carcinoma | Vitro: SMMC-7721, Huh7 and HCCLM3 cell lines (500–30,000 µg/mL for 24/48 h) Vivo: HCCMLM3 xenograft male BALB/c nude mice (1 g/kg b.w. orally administered fucoidan for 21 days) | ↓ Migration and invasion ↓ Invadopodia-related proteins (Src, Cortactin, N-WASP, ARP3, CDC42, MMP2, MT1-MMP) ↓ Integrin αVβ3 ↓ Tumor growth and lung metastatic foci in vivo | [35] |
| Fucoidan FP08S2 (from Sargassum fusiforme) | Human lung cancer | Vitro: HMEC-1 cells (4.21, 8.42, 16.84 μM for 8, 12 or 24 h) [36] (25–800 µg/mL for 72 h) [37] Vivo: CAM (2 days after 50–150 μg/egg fucoidan injection) A549 cancer cells xenograft nude mice (0.5 or 10 mg/kg b.w. via tail vein injection every day for 27 days) | Block of VEGFR2/Erk/VEGF signaling pathway in HMEC-1 cells ↓ Migration and invasion of HMEC-1 cells ↓ Tube formation and VEGF and HIF-1α ↓ Tumor growth and microvessel formation in vivo | [36,37] |
| Fucoidan (from Turbinaria conoides): | Human pancreatic cancer | Vitro: MiaPaCa-2 and Panc-1 cell line (25–100 µg/mL for 48 h) HAEC cells (12.5–100 µg/mL for 3 h) Vivo: CAM (4 days after 250–1000 µg/mL fucoidan injection) | ↓ Migration in MiaPaCa-1 cells ↓ Tube formation ↓ Tubule junctions and neovascularization | [38] |
| Fucoidan (from Undaria pinnatifida sporophylls) (Dalian Aquaculture Group Co., Ltd., Dalian, China) | Murine hepatocarcinoma | Vitro: Hca-F cell line (100–400 µg/mL for 24 h) Murine LEC (50–200 μg/mL for 48 h in a hypoxic environment) [39] Human LEC (100–400 μg/mL for 24, 48 or 72 h) [40] Vivo: Hca-F cancer cells xenograft male 615 mice (15 mg/kg and 30 mg/kg b.w. abdominally injected once a day for 3 weeks) [39] or (30, 60, 120 mg/kg intragastrically administered for 14 days) [40] | In hypoxic conditions: ↓ HIF-1α/VEGFC and HGF ↓ PI3K/Akt/mTOR pathway in murine LEC ↓ NF-κB/PI3K/Akt pathway in human LEC ↓ MMP-2, 9; ↑ TIMP ↓ Lymphangiogenesis; ↓ PROX1 and VEGF3 ↓ Metastasis and lymphangiogenesis ↓ HIF-1α and VEGF-C ↓ VEGF3 and lymphatic vessel density | [39,40] |
| Fucoidan (source not specified) | Human lung cancer | Vitro: A549 and H1650 NSCLC cell lines (10,000 or 16,000 µg/mL for 48 h) Vivo: NSCLC cancer cells xenograft model (25 mg/kg b.w. via oral gavage every day for 14 days) | ↓ Angiogenesis via m-TOR pathway ↓ Migration and invasion via EMT ↓ VEGFA, N-cadherin; ↑ E-cadherin | [41] |
| Oligo-fucoidan (source not specified) | Human hepatocellular carcinoma | Vivo: Transgenic human hepatocellular carcinoma zebrafish model (fucoidan 6.6 mg/kg b.w. oral gavage in combination with WNK463, Regonafenib or Rafoxanide twice a week for one month) | ↓ WNK1–OSR1–PPP2R1A axis ↓ Migration | [42] |
| Fucoidan-coated manganese dioxide nanoparticles (Fuco-MnO2-NPs) | Human pancreatic cancer | Vitro: AsPC-1 and BxPC-3 cell lines (0.5–20 µg/mL for 48 h) Vivo: BxPC3 cancer cells xenograft mice (NPs 200 ng/50 µL and fucoidan 100 ng/50 µL by intratumoral injection weekly up to 30 days) | Suppression HIF-1α in hypoxic conditions ↓ VEGFR2 and CD31 | [43] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Turrini, E.; Maffei, F.; Fimognari, C. Ten Years of Research on Fucoidan and Cancer: Focus on Its Antiangiogenic and Antimetastatic Effects. Mar. Drugs 2023, 21, 307. https://doi.org/10.3390/md21050307
Turrini E, Maffei F, Fimognari C. Ten Years of Research on Fucoidan and Cancer: Focus on Its Antiangiogenic and Antimetastatic Effects. Marine Drugs. 2023; 21(5):307. https://doi.org/10.3390/md21050307
Chicago/Turabian StyleTurrini, Eleonora, Francesca Maffei, and Carmela Fimognari. 2023. "Ten Years of Research on Fucoidan and Cancer: Focus on Its Antiangiogenic and Antimetastatic Effects" Marine Drugs 21, no. 5: 307. https://doi.org/10.3390/md21050307
APA StyleTurrini, E., Maffei, F., & Fimognari, C. (2023). Ten Years of Research on Fucoidan and Cancer: Focus on Its Antiangiogenic and Antimetastatic Effects. Marine Drugs, 21(5), 307. https://doi.org/10.3390/md21050307