
Searching for Novel Sources of Hydrogen Sulfide Donors: Chemical Profiling of Polycarpa aurata Extract and Evaluation of the Anti-Inflammatory Effects

 ,
,  ,
,  ,
,  ,
,  ,
,  ,
, 
 , and
, and
Abstract
:
1. Introduction
2. Results
2.1. Extraction and Fractionation of P. aurata
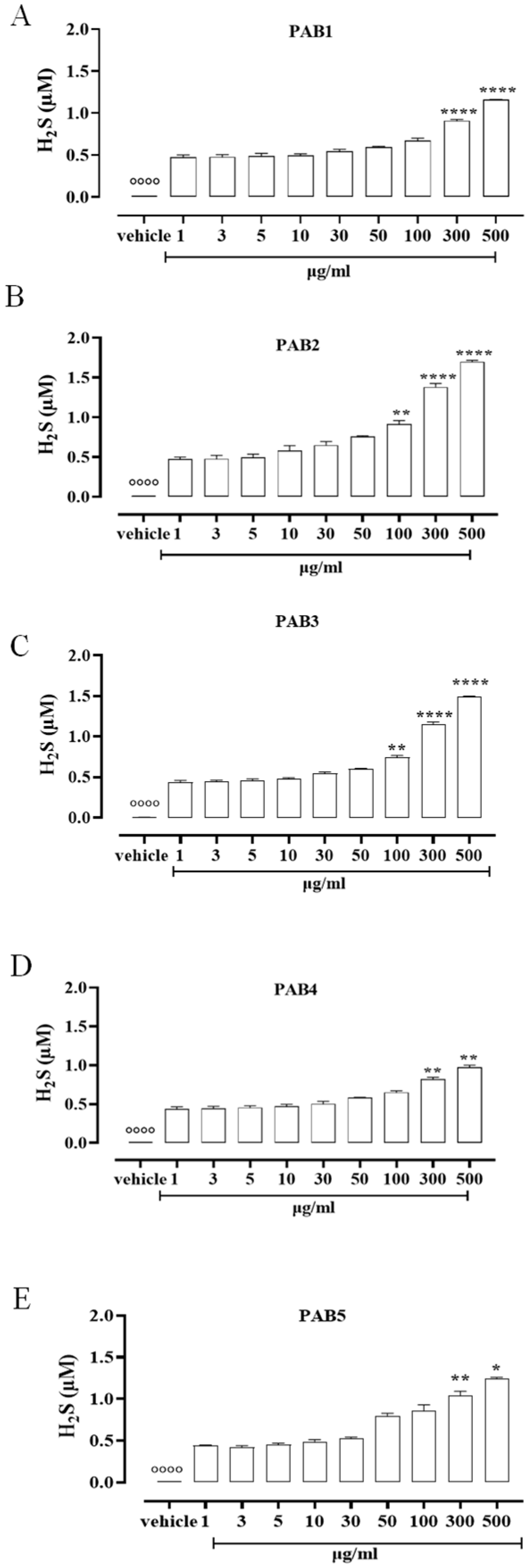
2.2. PAB1-PAB5 Release H2S
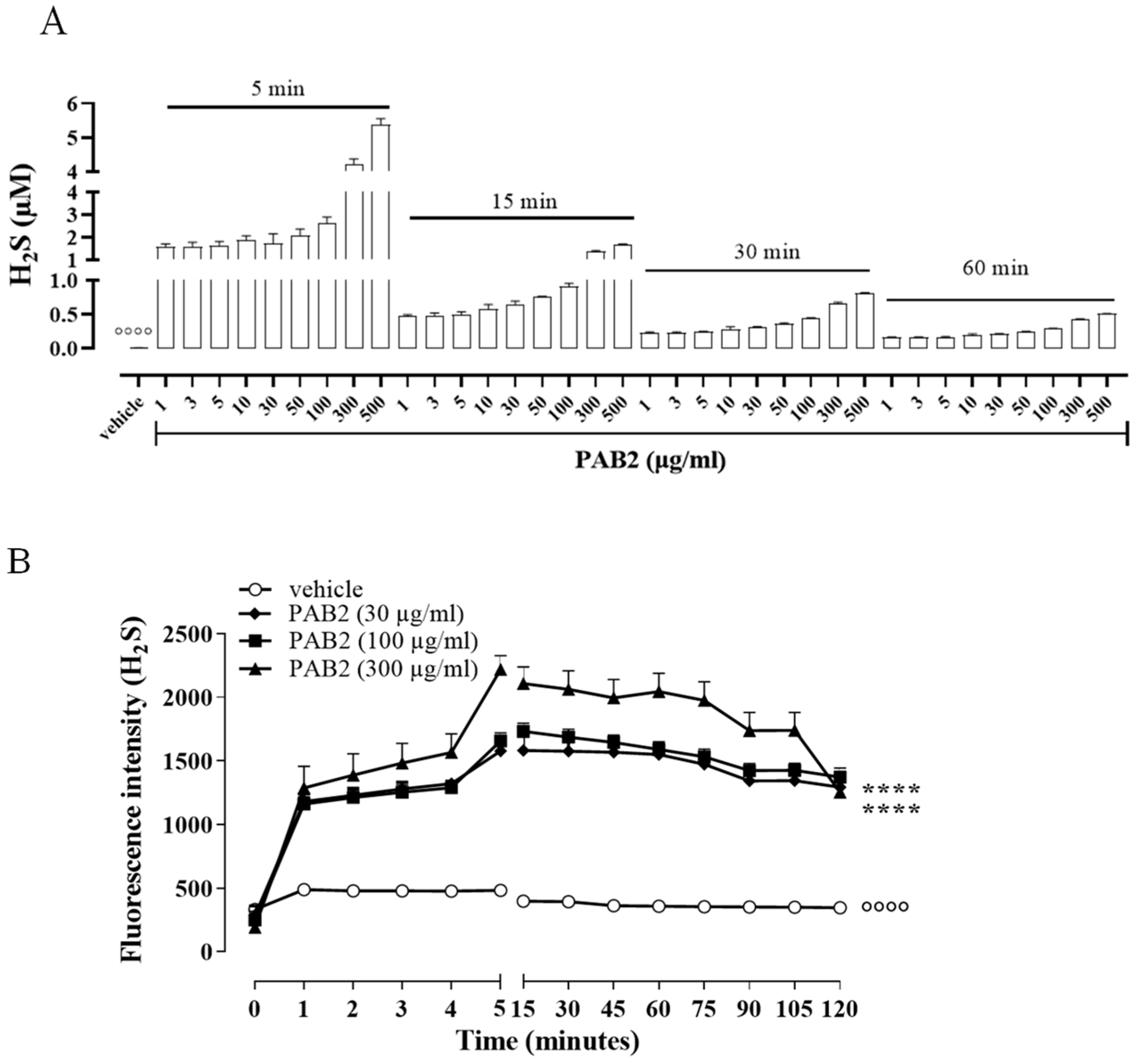
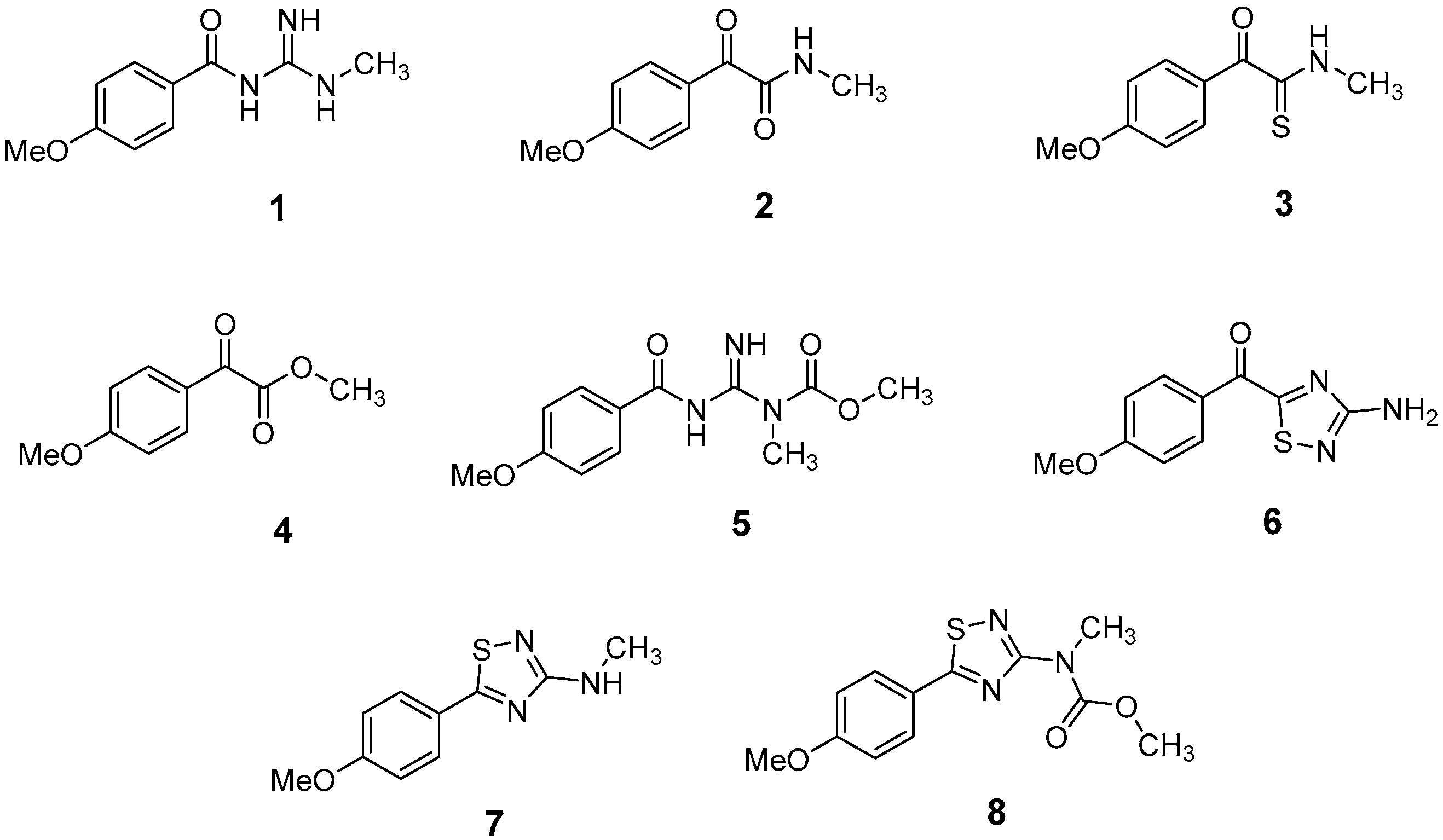
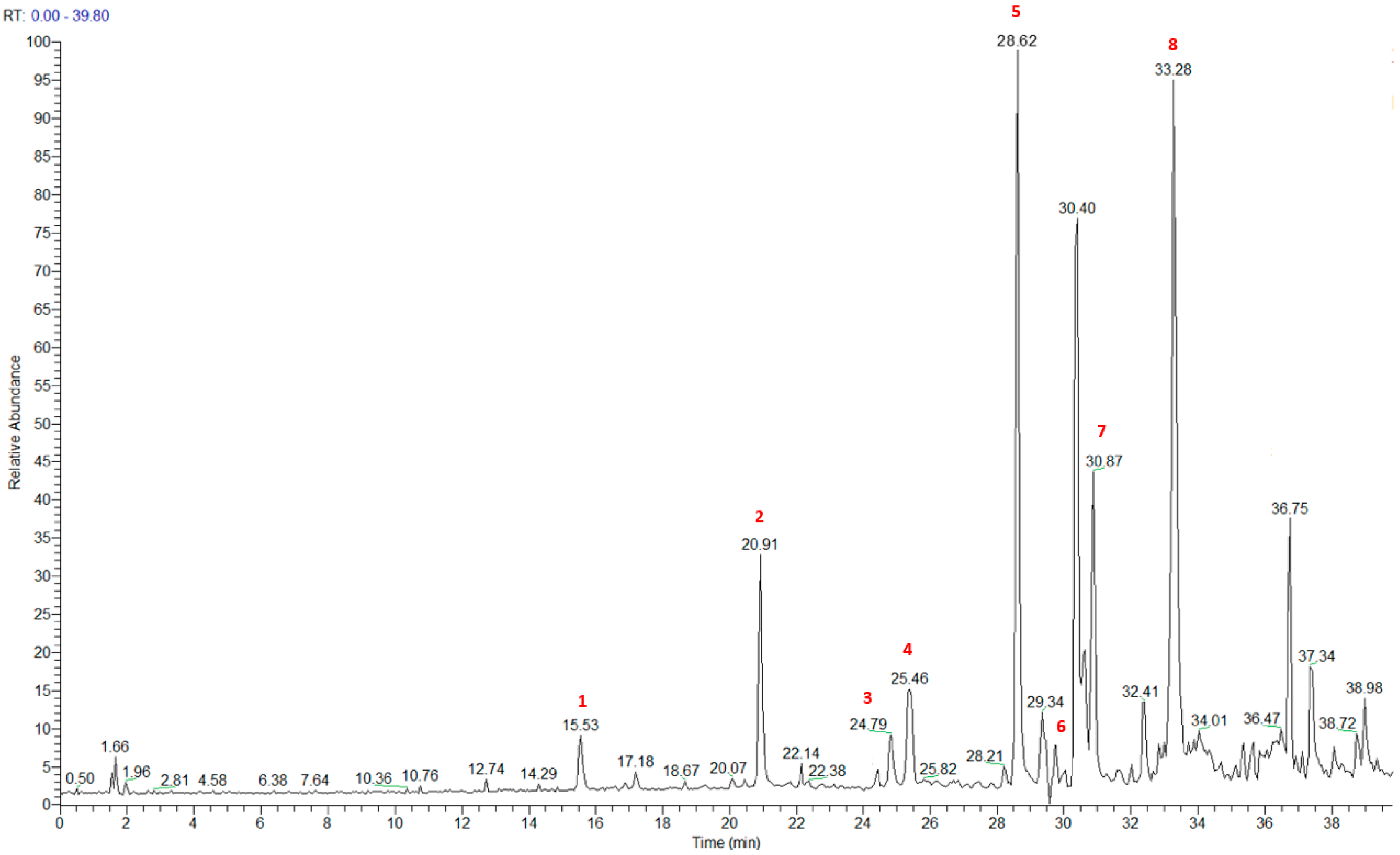
2.3. LC-HRMS and NMR Analyses of PAB2
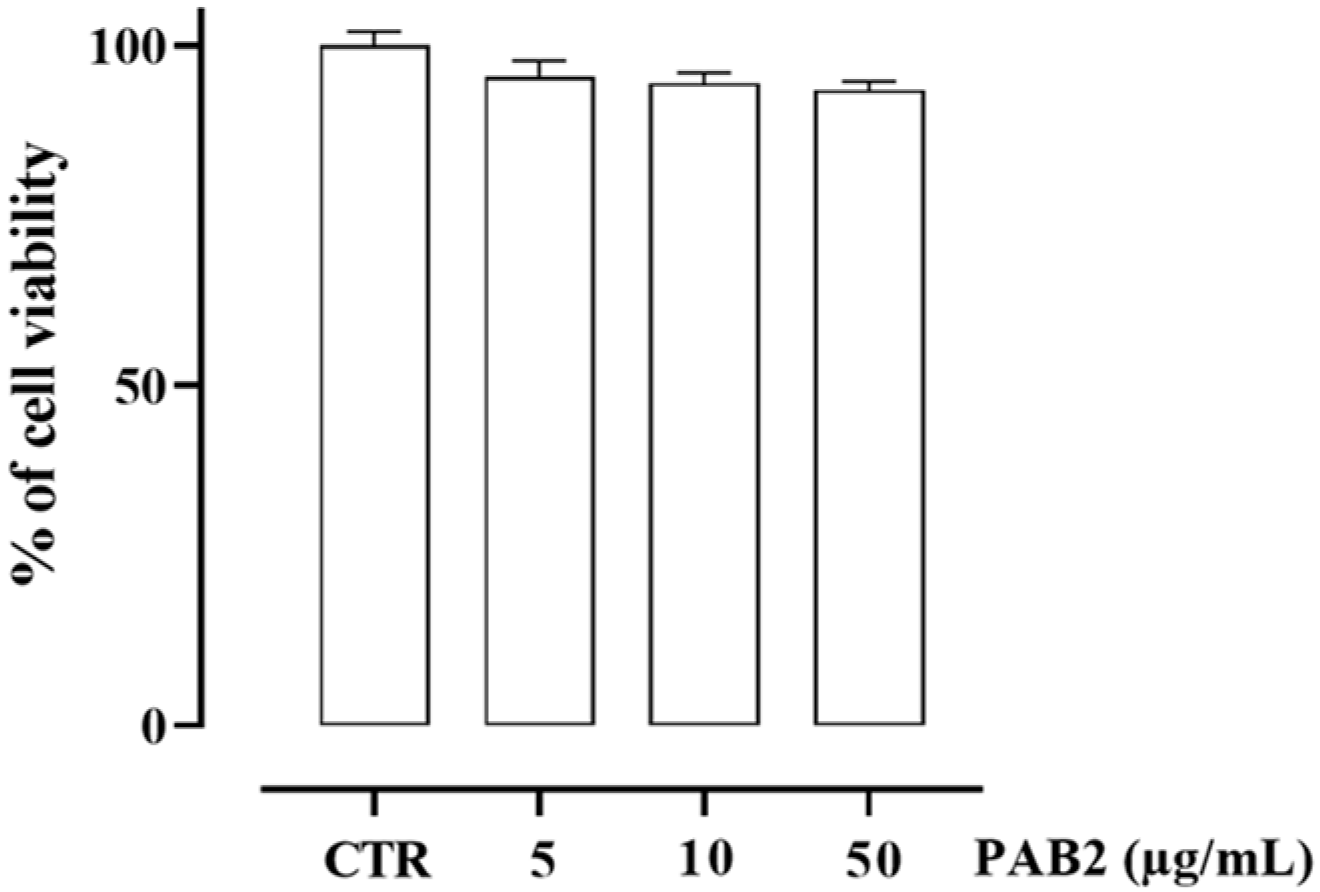
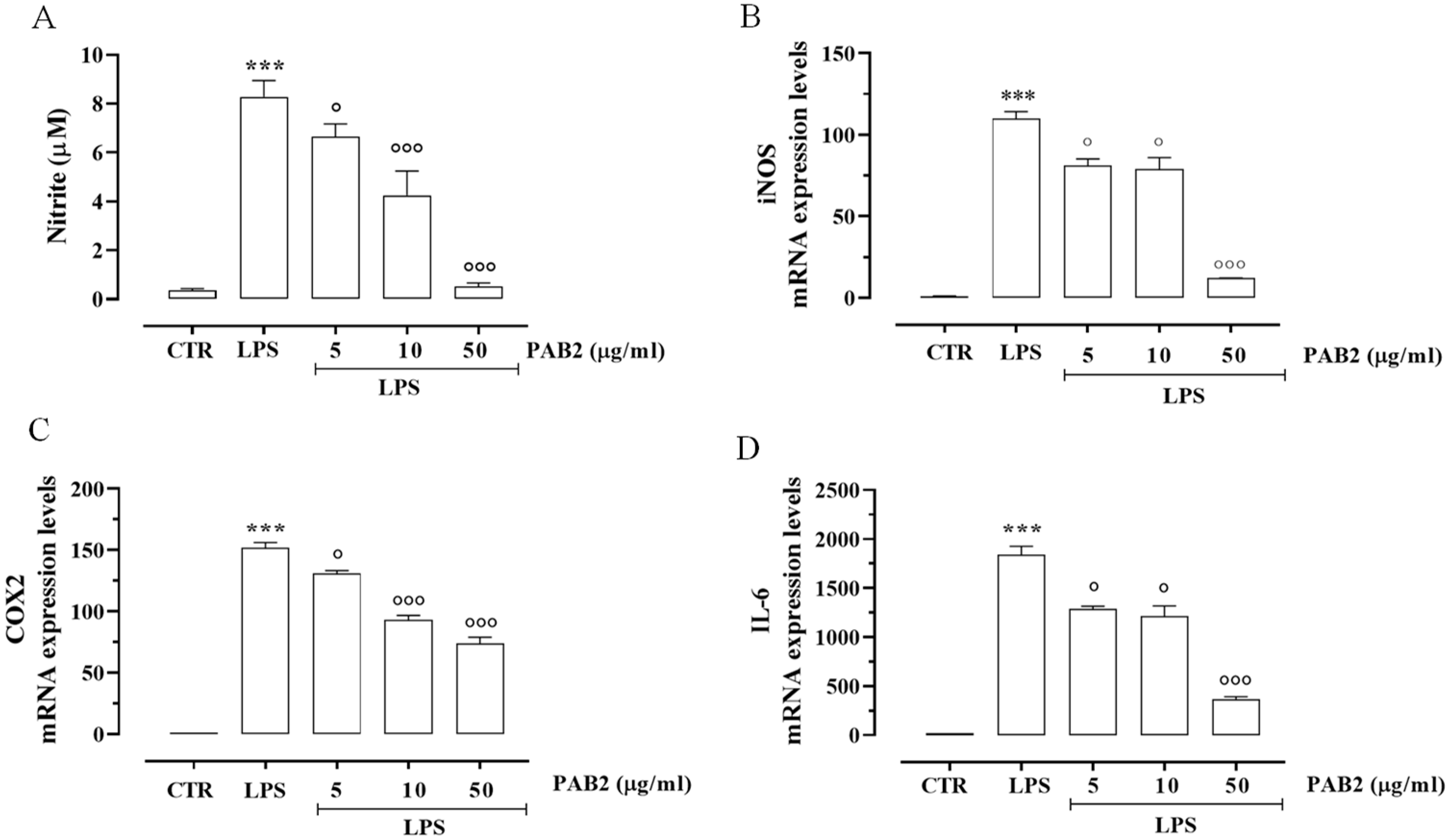
2.4. Effect of PAB2 on LPS-Stimulated J774 Cells
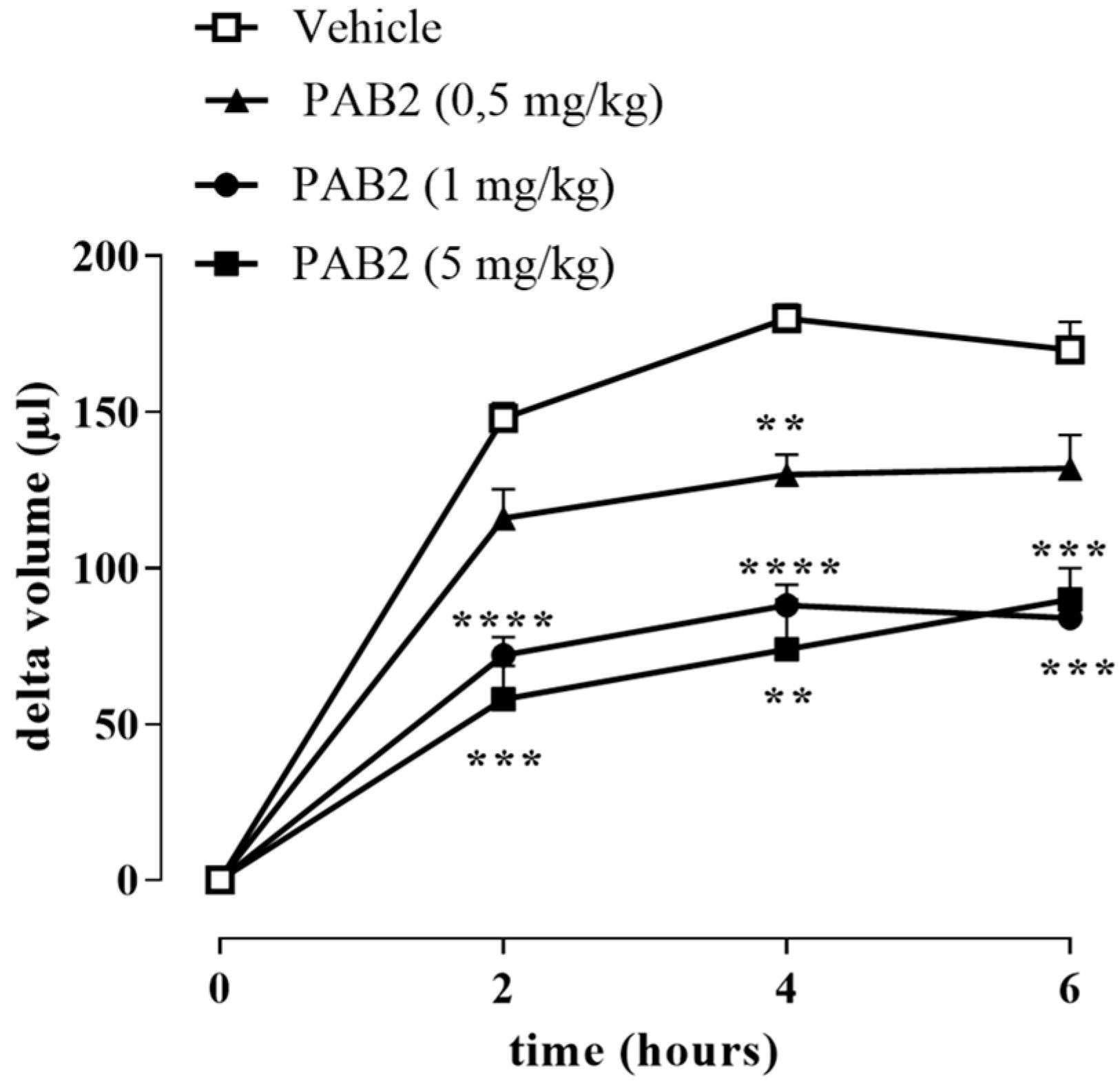
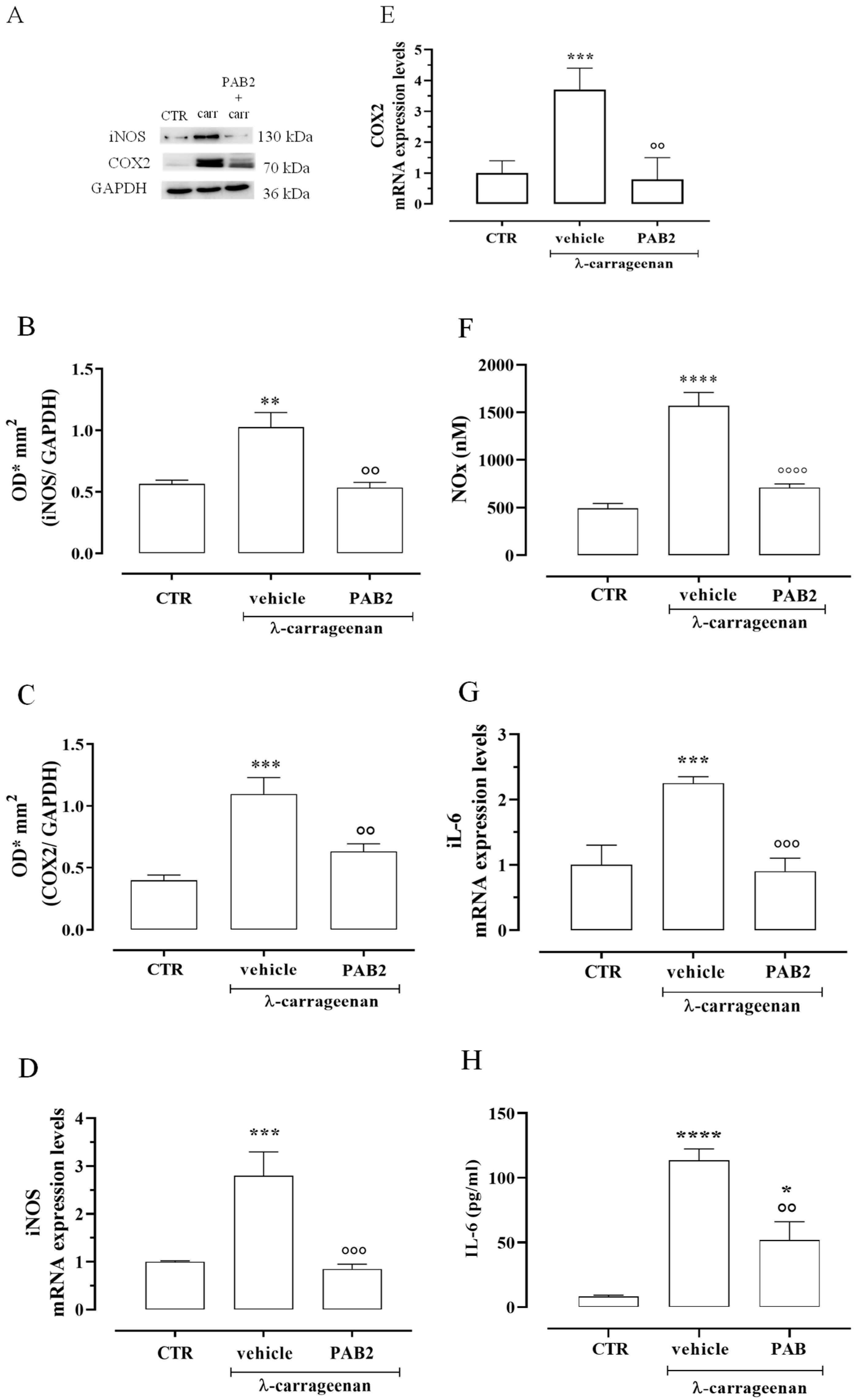
2.5. PAB2 Induces Anti-Inflammatory Effect In Vivo
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Collection, Extraction, and Fractionation of the Ascidian P. aurata
4.3. LC-HRMS/MS Profiling of P. aurata
4.4. Hydrogen Sulfide Measurement in a Cell-Free Assay
4.5. Cell Culture
4.6. Hydrogen Sulfide Measurement in J774 Cells
4.7. Measurement of NO Production in J774 Cells
4.8. RNA Purification and Quantitative Real-Time PCR
4.9. Animals
4.10. Measurement of Mouse Paw Edema
4.11. Western Blotting Analysis
4.12. NOx Measurement in the Mouse Paw
4.13. IL-6 Measurement in the Mouse Paw
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Szabo, C. A Timeline of Hydrogen Sulfide (H2S) Research: From Environmental Toxin to Biological Mediator. Biochem. Pharmacol. 2018, 149, 5–19. [Google Scholar] [CrossRef]
- Cirino, G.; Szabo, C.; Papapetropoulos, A. Physiological Roles of Hydrogen Sulfide in Mammalian Cells, Tissues, and Organs. Physiol. Rev. 2023, 103, 31–276. [Google Scholar] [CrossRef] [PubMed]
- Szabo, C. Hydrogen Sulfide, an Enhancer of Vascular Nitric Oxide Signaling: Mechanisms and Implications. Am. J. Physiol. Cell Physiol. 2017, 312, C3–C15. [Google Scholar] [CrossRef] [PubMed]
- Mitidieri, E.; Gurgone, D.; Caiazzo, E.; Tramontano, T.; Cicala, C.; Sorrentino, R.; d’Emmanuele di Villa Bianca, R. L-cysteine/Cystathionine-β-synthase-induced Relaxation in Mouse Aorta Involves a L-serine/Sphingosine-1-phosphate/NO Pathway. Br. J. Pharmacol. 2020, 177, 734–744. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.Y.; Ping, C.Y.; Mok, Y.-Y.; Ling, L.; Whiteman, M.; Bhatia, M.; Moore, P.K. Regulation of Vascular Nitric Oxide in Vitro and in Vivo; a New Role for Endogenous Hydrogen Sulphide? Br. J. Pharmacol. 2006, 149, 625–634. [Google Scholar] [CrossRef] [PubMed]
- Yong, Q.-C.; Hu, L.-F.; Wang, S.; Huang, D.; Bian, J.-S. Hydrogen Sulfide Interacts with Nitric Oxide in the Heart: Possible Involvement of Nitroxyl. Cardiovasc. Res. 2010, 88, 482–491. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Padovani, D.; Leslie, R.A.; Chiku, T.; Banerjee, R. Relative Contributions of Cystathionine β-Synthase and γ-Cystathionase to H2S Biogenesis via Alternative Trans-Sulfuration Reactions. J. Biol. Chem. 2009, 284, 22457–22466. [Google Scholar] [CrossRef] [PubMed]
- Kabil, O.; Vitvitsky, V.; Xie, P.; Banerjee, R. The Quantitative Significance of the Transsulfuration Enzymes for H2S Production in Murine Tissues. Antioxid. Redox Signal 2011, 15, 363–372. [Google Scholar] [CrossRef]
- Kimura, H. Production and Physiological Effects of Hydrogen Sulfide. Antioxid. Redox Signal 2014, 20, 783–793. [Google Scholar] [CrossRef]
- Shibuya, N.; Mikami, Y.; Kimura, Y.; Nagahara, N.; Kimura, H. Vascular Endothelium Expresses 3-Mercaptopyruvate Sulfurtransferase and Produces Hydrogen Sulfide. J. Biochem. 2009, 146, 623–626. [Google Scholar] [CrossRef]
- Lo Faro, M.L.; Fox, B.; Whatmore, J.L.; Winyard, P.G.; Whiteman, M. Hydrogen Sulfide and Nitric Oxide Interactions in Inflammation. Nitric Oxide 2014, 41, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Rodkin, S.; Nwosu, C.; Sannikov, A.; Tyurin, A.; Chulkov, V.S.; Raevskaya, M.; Ermakov, A.; Kirichenko, E.; Gasanov, M. The Role of Gasotransmitter-Dependent Signaling Mechanisms in Apoptotic Cell Death in Cardiovascular, Rheumatic, Kidney, and Neurodegenerative Diseases and Mental Disorders. Int. J. Mol. Sci. 2023, 24, 6014. [Google Scholar] [CrossRef] [PubMed]
- Giuffrè, A.; Vicente, J.B. Hydrogen Sulfide Biochemistry and Interplay with Other Gaseous Mediators in Mammalian Physiology. Oxid. Med. Cell Longev. 2018, 2018, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Cirino, G.; Vellecco, V.; Bucci, M. Nitric Oxide and Hydrogen Sulfide: The Gasotransmitter Paradigm of the Vascular System. Br. J. Pharmacol. 2017, 174, 4021–4031. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, M.; Gaddam, R.R. Hydrogen Sulfide in Inflammation: A Novel Mediator and Therapeutic Target. Antioxid. Redox Signal 2021, 34, 1368–1377. [Google Scholar] [CrossRef] [PubMed]
- Wallace, J.L.; Ferraz, J.G.P.; Muscara, M.N. Hydrogen Sulfide: An Endogenous Mediator of Resolution of Inflammation and Injury. Antioxid. Redox Signal 2012, 17, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Whiteman, M.; Winyard, P.G. Hydrogen Sulfide and Inflammation: The Good, the Bad, the Ugly and the Promising. Expert. Rev. Clin. Pharmacol. 2011, 4, 13–32. [Google Scholar] [CrossRef]
- Andruski, B.; McCafferty, D.-M.; Ignacy, T.; Millen, B.; McDougall, J.J. Leukocyte Trafficking and Pain Behavioral Responses to a Hydrogen Sulfide Donor in Acute Monoarthritis. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 295, R814–R820. [Google Scholar] [CrossRef]
- Dufton, N.; Natividad, J.; Verdu, E.F.; Wallace, J.L. Hydrogen Sulfide and Resolution of Acute Inflammation: A Comparative Study Utilizing a Novel Fluorescent Probe. Sci. Rep. 2012, 2, 499. [Google Scholar] [CrossRef]
- d’Emmanuele di Villa Bianca, R.; Coletta, C.; Mitidieri, E.; De Dominicis, G.; Rossi, A.; Sautebin, L.; Cirino, G.; Bucci, M.; Sorrentino, R. Hydrogen Sulphide Induces Mouse Paw Oedema through Activation of Phospholipase A2. Br. J. Pharmacol. 2010, 161, 1835–1842. [Google Scholar] [CrossRef]
- Zhang, H.; Hegde, A.; Ng, S.W.; Adhikari, S.; Moochhala, S.M.; Bhatia, M. Hydrogen Sulfide Up-Regulates Substance P in Polymicrobial Sepsis-Associated Lung Injury. J. Immunol. 2007, 179, 4153–4160. [Google Scholar] [CrossRef] [PubMed]
- Vellecco, V.; Esposito, E.; Indolfi, C.; Saviano, A.; Panza, E.; Bucci, M.; Brancaleone, V.; Cirino, G.; d’Emmanuele di Villa Bianca, R.; Sorrentino, R.; et al. Biphasic Inflammatory Response Induced by Intra-Plantar Injection of L-Cysteine: Role of CBS-Derived H2S and S1P/NO Signaling. Biomed. Pharmacother. 2023, 167, 115536. [Google Scholar] [CrossRef] [PubMed]
- Whiteman, M.; Le Trionnaire, S.; Chopra, M.; Fox, B.; Whatmore, J. Emerging Role of Hydrogen Sulfide in Health and Disease: Critical Appraisal of Biomarkers and Pharmacological Tools. Clin. Sci. 2011, 121, 459–488. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Yan, R.; Zhou, X.; Ji, F.; Zhang, B. Hydrogen Sulfide Improves Colonic Barrier Integrity in DSS-Induced Inflammation in Caco-2 Cells and Mice. Int. Immunopharmacol. 2016, 39, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Qian, H.; Zheng, N.; Lu, Q.; Han, Y. GYY4137 Ameliorates Sepsis-Induced Cardiomyopathy via NLRP3 Pathway. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2022, 1868, 166497. [Google Scholar] [CrossRef] [PubMed]
- Whiteman, M.; Li, L.; Rose, P.; Tan, C.-H.; Parkinson, D.B.; Moore, P.K. The Effect of Hydrogen Sulfide Donors on Lipopolysaccharide-Induced Formation of Inflammatory Mediators in Macrophages. Antioxid. Redox Signal 2010, 12, 1147–1154. [Google Scholar] [CrossRef] [PubMed]
- Gerő, D.; Torregrossa, R.; Perry, A.; Waters, A.; Le-Trionnaire, S.; Whatmore, J.L.; Wood, M.; Whiteman, M. The Novel Mitochondria-Targeted Hydrogen Sulfide (H 2 S) Donors AP123 and AP39 Protect against Hyperglycemic Injury in Microvascular Endothelial Cells in Vitro. Pharmacol. Res. 2016, 113, 186–198. [Google Scholar] [CrossRef]
- Levinn, C.M.; Cerda, M.M.; Pluth, M.D. Activatable Small-Molecule Hydrogen Sulfide Donors. Antioxid. Redox Signal 2020, 32, 96–109. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.N.; Zheng, Y.Q.; Wang, B.H. Prodrugs of Hydrogen Sulfide and Related Sulfur Species: Recent Development. Chin. J. Nat. Med. 2020, 18, 296–307. [Google Scholar] [CrossRef]
- Magierowski, M.; Magierowska, K.; Surmiak, M.; Hubalewska-Mazgaj, M.; Kwiecien, S.; Wallace, J.L.; Brzozowski, T. The Effect of Hydrogen Sulfide-Releasing Naproxen (ATB-346) versus Naproxen on Formation of Stress-Induced Gastric Lesions, the Regulation of Systemic Inflammation, Hypoxia and Alterations in Gastric Microcirculation. J. Physiol. Pharmacol. 2017, 68, 749–756. [Google Scholar]
- Dief, A.E.; Mostafa, D.K.; Sharara, G.M.; Zeitoun, T.H. Hydrogen Sulfide Releasing Naproxen Offers Better Anti-Inflammatory and Chondroprotective Effect Relative to Naproxen in a Rat Model of Zymosan Induced Arthritis. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 1537–1546. [Google Scholar]
- Benavides, G.A.; Squadrito, G.L.; Mills, R.W.; Patel, H.D.; Isbell, T.S.; Patel, R.P.; Darley-Usmar, V.M.; Doeller, J.E.; Kraus, D.W. Hydrogen Sulfide Mediates the Vasoactivity of Garlic. Proc. Natl. Acad. Sci. USA 2007, 104, 17977–17982. [Google Scholar] [CrossRef]
- Martelli, A.; Citi, V.; Testai, L.; Brogi, S.; Calderone, V. Organic Isothiocyanates as Hydrogen Sulfide Donors. Antioxid. Redox Signal 2020, 32, 110–144. [Google Scholar] [CrossRef] [PubMed]
- Micheli, L.; Mitidieri, E.; Turnaturi, C.; Vanacore, D.; Ciampi, C.; Lucarini, E.; Cirino, G.; Ghelardini, C.; Sorrentino, R.; Di Cesare Mannelli, L.; et al. Beneficial Effect of H2S-Releasing Molecules in an In Vitro Model of Sarcopenia: Relevance of Glucoraphanin. Int. J. Mol. Sci. 2022, 23, 5955. [Google Scholar] [CrossRef] [PubMed]
- Bello, I.; Smimmo, M.; d’Emmanuele di Villa Bianca, R.; Bucci, M.; Cirino, G.; Panza, E.; Brancaleone, V. Erucin, an H2S-Releasing Isothiocyanate, Exerts Anticancer Effects in Human Triple-Negative Breast Cancer Cells Triggering Autophagy-Dependent Apoptotic Cell Death. Int. J. Mol. Sci. 2023, 24, 6764. [Google Scholar] [CrossRef] [PubMed]
- Martelli, A.; Piragine, E.; Citi, V.; Testai, L.; Pagnotta, E.; Ugolini, L.; Lazzeri, L.; Di Cesare Mannelli, L.; Manzo, O.L.; Bucci, M.; et al. Erucin Exhibits Vasorelaxing Effects and Antihypertensive Activity by H2S-releasing Properties. Br. J. Pharmacol. 2020, 177, 824–835. [Google Scholar] [CrossRef] [PubMed]
- Martelli, A.; Testai, L.; Citi, V.; Marino, A.; Bellagambi, F.G.; Ghimenti, S.; Breschi, M.C.; Calderone, V. Pharmacological Characterization of the Vascular Effects of Aryl Isothiocyanates: Is Hydrogen Sulfide the Real Player? Vasc. Pharmacol. 2014, 60, 32–41. [Google Scholar] [CrossRef]
- Pluth, M.; Bailey, T.; Hammers, M.; Hartle, M.; Henthorn, H.; Steiger, A. Natural Products Containing Hydrogen Sulfide Releasing Moieties. Synlett 2015, 26, 2633–2643. [Google Scholar] [CrossRef]
- Caesar, L.K.; Cech, N.B. Synergy and Antagonism in Natural Product Extracts: When 1 + 1 Does Not Equal 2. Nat. Prod. Rep. 2019, 36, 869–888. [Google Scholar] [CrossRef]
- Genovese, M.; Imperatore, C.; Casertano, M.; Aiello, A.; Balestri, F.; Piazza, L.; Menna, M.; del Corso, A.; Paoli, P. Dual Targeting of PTP1B and Aldose Reductase with Marine Drug Phosphoeleganin: A Promising Strategy for Treatment of Type 2 Diabetes. Mar. Drugs 2021, 19, 535. [Google Scholar] [CrossRef]
- Casertano, M.; Genovese, M.; Piazza, L.; Balestri, F.; Del Corso, A.; Vito, A.; Paoli, P.; Santi, A.; Imperatore, C.; Menna, M. Identifying Human PTP1B Enzyme Inhibitors from Marine Natural Products: Perspectives for Developing of Novel Insulin-Mimetic Drugs. Pharmaceuticals 2022, 15, 325. [Google Scholar] [CrossRef]
- Casertano, M.; Genovese, M.; Santi, A.; Pranzini, E.; Balestri, F.; Piazza, L.; Del Corso, A.; Avunduk, S.; Imperatore, C.; Menna, M.; et al. Evidence of Insulin-Sensitizing and Mimetic Activity of the Sesquiterpene Quinone Avarone, a Protein Tyrosine Phosphatase 1B and Aldose Reductase Dual Targeting Agent from the Marine Sponge Dysidea Avara. Pharmaceutics 2023, 15, 528. [Google Scholar] [CrossRef] [PubMed]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine Natural Products. Nat. Prod. Rep. 2023, 40, 275–325. [Google Scholar] [CrossRef] [PubMed]
- Haque, N.; Parveen, S.; Tang, T.; Wei, J.; Huang, Z. Marine Natural Products in Clinical Use. Mar. Drugs 2022, 20, 528. [Google Scholar] [CrossRef]
- Imperatore, C.; Gimmelli, R.; Persico, M.; Casertano, M.; Guidi, A.; Saccoccia, F.; Ruberti, G.; Luciano, P.; Aiello, A.; Parapini, S.; et al. Investigating the Antiparasitic Potential of the Marine Sesquiterpene Avarone, Its Reduced Form Avarol, and the Novel Semisynthetic Thiazinoquinone Analogue Thiazoavarone. Mar. Drugs 2020, 18, 112. [Google Scholar] [CrossRef] [PubMed]
- Luciano, P.; Imperatore, C.; Senese, M.; Aiello, A.; Casertano, M.; Guo, Y.-W.; Menna, M. Assignment of the Absolute Configuration of Phosphoeleganin via Synthesis of Model Compounds. J. Nat. Prod. 2017, 80, 2118–2123. [Google Scholar] [CrossRef] [PubMed]
- Casertano, M.; Vito, A.; Aiello, A.; Imperatore, C.; Menna, M. Natural Bioactive Compounds from Marine Invertebrates That Modulate Key Targets Implicated in the Onset of Type 2 Diabetes Mellitus (T2DM) and Its Complications. Pharmaceutics 2023, 15, 2321. [Google Scholar] [CrossRef]
- Casertano, M.; Imperatore, C.; Luciano, P.; Aiello, A.; Putra, M.Y.; Gimmelli, R.; Ruberti, G.; Menna, M. Chemical Investigation of the Indonesian Tunicate Polycarpa Aurata and Evaluation of the Effects Against Schistosoma Mansoni of the Novel Alkaloids Polyaurines A and B. Mar. Drugs 2019, 17, 278. [Google Scholar] [CrossRef]
- Pham, C.-D.; Weber, H.; Hartmann, R.; Wray, V.; Lin, W.; Lai, D.; Proksch, P. New Cytotoxic 1,2,4-Thiadiazole Alkaloids from the Ascidian Polycarpa aurata. Org. Lett. 2013, 15, 2230–2233. [Google Scholar] [CrossRef]
- Abas, S.A.; Hossain, M.B.; van der Helm, D.; Schmitz, F.J.; Laney, M.; Cabuslay, R.; Schatzman, R.C. Alkaloids from the Tunicate Polycarpa aurata from Chuuk Atoll. J. Org. Chem. 1996, 61, 2709–2712. [Google Scholar] [CrossRef]
- Wessels, M.; König, G.M.; Wright, A.D. New 4-Methoxybenzoyl Derivatives from the Ascidian Polycarpa aurata. J. Nat. Prod. 2001, 64, 1556–1558. [Google Scholar] [CrossRef]
- Anstis, D.G.; Lindsay, A.C.; Söhnel, T.; Sperry, J. Synthesis of the 1,2,4-Thiadiazole Alkaloid Polyaurine B. J. Nat. Prod. 2020, 83, 1721–1724. [Google Scholar] [CrossRef]
- Nagy, G.; Clark, J.M.; Buzás, E.I.; Gorman, C.L.; Cope, A.P. Nitric Oxide, Chronic Inflammation and Autoimmunity. Immunol. Lett. 2007, 111, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Martelli, A.; Testai, L.; Citi, V.; Marino, A.; Pugliesi, I.; Barresi, E.; Nesi, G.; Rapposelli, S.; Taliani, S.; Da Settimo, F.; et al. Arylthioamides as H2S Donors: L-Cysteine-Activated Releasing Properties and Vascular Effects in Vitro and in Vivo. ACS Med. Chem. Lett. 2013, 4, 904–908. [Google Scholar] [CrossRef] [PubMed]
- Wallace, J.L.; Caliendo, G.; Santagada, V.; Cirino, G. Markedly Reduced Toxicity of a Hydrogen Sulphide-Releasing Derivative of Naproxen (ATB-346). Br. J. Pharmacol. 2010, 159, 1236–1246. [Google Scholar] [CrossRef] [PubMed]
- Hughes, M.N.; Centelles, M.N.; Moore, K.P. Making and Working with Hydrogen Sulfide. Free Radic. Biol. Med. 2009, 47, 1346–1353. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Whiteman, M.; Guan, Y.Y.; Neo, K.L.; Cheng, Y.; Lee, S.W.; Zhao, Y.; Baskar, R.; Tan, C.-H.; Moore, P.K. Characterization of a Novel, Water-Soluble Hydrogen Sulfide–Releasing Molecule (GYY4137). Circulation 2008, 117, 2351–2360. [Google Scholar] [CrossRef] [PubMed]
- Magli, E.; Perissutti, E.; Santagada, V.; Caliendo, G.; Corvino, A.; Esposito, G.; Esposito, G.; Fiorino, F.; Migliaccio, M.; Scognamiglio, A.; et al. H2S Donors and Their Use in Medicinal Chemistry. Biomolecules 2021, 11, 1899. [Google Scholar] [CrossRef] [PubMed]
- Baskar, R.; Bian, J. Hydrogen Sulfide Gas Has Cell Growth Regulatory Role. Eur. J. Pharmacol. 2011, 656, 5–9. [Google Scholar] [CrossRef]
- Rios, E.C.S.; Szczesny, B.; Soriano, F.G.; Olah, G.; Szabo, C. Hydrogen Sulfide Attenuates Cytokine Production through the Modulation of Chromatin Remodeling. Int. J. Mol. Med. 2015, 35, 1741–1746. [Google Scholar] [CrossRef]
- Hirayama, D.; Iida, T.; Nakase, H. The Phagocytic Function of Macrophage-Enforcing Innate Immunity and Tissue Homeostasis. Int. J. Mol. Sci. 2017, 19, 92. [Google Scholar] [CrossRef]
- Wang, Z.; Guan, Y.; Yang, R.; Li, J.; Wang, J.; Jia, A.-Q. Anti-Inflammatory Activity of 3-Cinnamoyltribuloside and Its Metabolomic Analysis in LPS-Activated RAW 264.7 Cells. BMC Complement. Med. Ther. 2020, 20, 329. [Google Scholar] [CrossRef]
- Sharma, J.N.; Al-Omran, A.; Parvathy, S.S. Role of Nitric Oxide in Inflammatory Diseases. Inflammopharmacology 2007, 15, 252–259. [Google Scholar] [CrossRef]
- Hämäläinen, M.; Lilja, R.; Kankaanranta, H.; Moilanen, E. Inhibition of INOS Expression and NO Production by Anti-Inflammatory Steroids. Pulm. Pharmacol. Ther. 2008, 21, 331–339. [Google Scholar] [CrossRef]
- Scheller, J.; Chalaris, A.; Schmidt-Arras, D.; Rose-John, S. The Pro- and Anti-Inflammatory Properties of the Cytokine Interleukin-6. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2011, 1813, 878–888. [Google Scholar] [CrossRef]
- Imperatore, C.; Varriale, A.; Rivieccio, E.; Pennacchio, A.; Staiano, M.; D’Auria, S.; Casertano, M.; Altucci, C.; Valadan, M.; Singh, M.; et al. Spectroscopic Properties of Two 5′-(4-Dimethylamino)Azobenzene Conjugated G-Quadruplex Forming Oligonucleotides. Int. J. Mol. Sci. 2020, 21, 7103. [Google Scholar] [CrossRef] [PubMed]
- Imperatore, C.; Valadan, M.; Tartaglione, L.; Persico, M.; Ramunno, A.; Menna, M.; Casertano, M.; Dell’Aversano, C.; Singh, M.; d’Aulisio Garigliota, M.L.; et al. Exploring the Photodynamic Properties of Two Antiproliferative Benzodiazopyrrole Derivatives. Int. J. Mol. Sci. 2020, 21, 1246. [Google Scholar] [CrossRef] [PubMed]
- Mitidieri, E.; Tramontano, T.; Gurgone, D.; Citi, V.; Calderone, V.; Brancaleone, V.; Katsouda, A.; Nagahara, N.; Papapetropoulos, A.; Cirino, G.; et al. Mercaptopyruvate Acts as Endogenous Vasodilator Independently of 3-Mercaptopyruvate Sulfurtransferase Activity. Nitric Oxide 2018, 75, 53–59. [Google Scholar] [CrossRef]
- Benigno, D.; Virgilio, A.; Bello, I.; La Manna, S.; Vellecco, V.; Bucci, M.; Marasco, D.; Panza, E.; Esposito, V.; Galeone, A. Properties and Potential Antiproliferative Activity of Thrombin-Binding Aptamer (TBA) Derivatives with One or Two Additional G-Tetrads. Int. J. Mol. Sci. 2022, 23, 14921. [Google Scholar] [CrossRef] [PubMed]
- Putra, M.Y.; Ianaro, A.; Panza, E.; Bavestrello, G.; Cerrano, C.; Fattorusso, E.; Taglialatela-Scafati, O. Sinularioside, a Triacetylated Glycolipid from the Indonesian Soft Coral Sinularia Sp., Is an Inhibitor of NO Release. Bioorganic Med. Chem. Lett. 2012, 22, 2723–2725. [Google Scholar] [CrossRef]
- Chatterjee, P.K.; di Villa Bianca, R.D.; Sivarajah, A.; McDonald, M.C.; Cuzzocrea, S.; Thiemermann, C. Pyrrolidine Dithiocarbamate Reduces Renal Dysfunction and Injury Caused by Ischemia/Reperfusion of the Rat Kidney. Eur. J. Pharmacol. 2003, 482, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Kilkenny, C.; Browne, W.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Animal Research: Reporting in Vivo Experiments: The ARRIVE Guidelines. Br. J. Pharmacol. 2010, 160, 1577–1579. [Google Scholar] [CrossRef] [PubMed]
- Lilley, E.; Stanford, S.C.; Kendall, D.E.; Alexander, S.P.H.; Cirino, G.; Docherty, J.R.; George, C.H.; Insel, P.A.; Izzo, A.A.; Ji, Y.; et al. ARRIVE 2.0 and the British Journal of Pharmacology: Updated Guidance for 2020. Br. J. Pharmacol. 2020, 177, 3611–3616. [Google Scholar] [CrossRef]
- Brancaleone, V.; Vellecco, V.; Matassa, D.S.; d’Emmanuele di Villa Bianca, R.; Sorrentino, R.; Ianaro, A.; Bucci, M.; Esposito, F.; Cirino, G. Crucial Role of Androgen Receptor in Vascular H2S Biosynthesis Induced by Testosterone. Br. J. Pharmacol. 2015, 172, 1505–1515. [Google Scholar] [CrossRef] [PubMed]
- Vellecco, V.; Mitidieri, E.; Gargiulo, A.; Brancaleone, V.; Matassa, D.; Klein, T.; Esposito, F.; Cirino, G.; Bucci, M. Vascular Effects of Linagliptin in Non-Obese Diabetic Mice Are Glucose-Independent and Involve Positive Modulation of the Endothelial Nitric Oxide Synthase (ENOS)/Caveolin-1 (CAV-1) Pathway. Diabetes Obes. Metab. 2016, 18, 1236–1243. [Google Scholar] [CrossRef]
- Montanaro, R.; Vellecco, V.; Torregrossa, R.; Casillo, G.M.; Manzo, O.L.; Mitidieri, E.; Bucci, M.; Castaldo, S.; Sorrentino, R.; Whiteman, M.; et al. Hydrogen Sulfide Donor AP123 Restores Endothelial Nitric Oxide-Dependent Vascular Function in Hyperglycemia via a CREB-Dependent Pathway. Redox Biol. 2023, 62, 102657. [Google Scholar] [CrossRef]
- Ebrahimi, B.; Baroutian, S.; Li, J.; Zhang, B.; Ying, T.; Lu, J. Combination of Marine Bioactive Compounds and Extracts for the Prevention and Treatment of Chronic Diseases. Front. Nutr. 2023, 9, 1047026. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exp. MS (m/z) | Δ ppm, RDB | Formula | RT | Putative Compound 1 |
|---|---|---|---|---|
| 208.1078 | −1.217, 5.5 | C10H14N3O2+ | 15.53 | 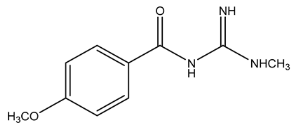 |
| Fragment ions | ||||
| Exp. MS (m/z) | Δ ppm, RDB | Formula | Structural hypothesis | |
| 191.0816 | 0.502, 6.5 | C10H11N2O2+ | 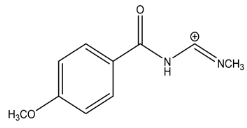 | |
| 152.0705 | −0.691, 4.5 | C8H10NO2+ | 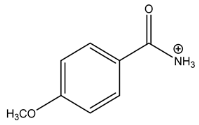 | |
| 135.0439 | −1.155, 5.5 | C8H7O2+ | 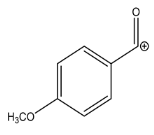 | |
| Exp. MS (m/z) | Δ ppm, RDB | Formula | RT | Putative Compound 2 |
| 194.0809 | −1.390, 5.5 | C10H12NO3+ | 20.91 | 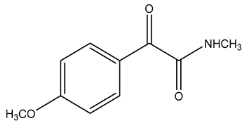 |
| Fragment ions | ||||
| Exp. MS (m/z) | Δ ppm, RDB | Formula | Structural hypothesis | |
| 135.0439 | −1.155, 5.5 | C8H7O2+ | 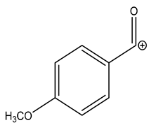 | |
| Exp. MS (m/z) | Δ ppm, RDB | Formula | RT | Putative Compound 3 |
| 210.0584 | 0.353, 5.5 | C10H12NSO2+ | 24.79 | 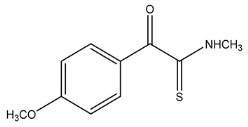 |
| Fragment ions | ||||
| Exp. MS (m/z) | Δ ppm, RDB | Formula | Structural hypothesis | |
| 135.0439 | −0.451, 5.5 | C8H7O2+ | 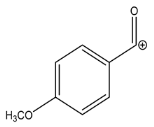 | |
| 109.0648 | 0.078, 3.5 | C7H9O+ | 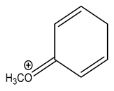 | |
| Exp. MS (m/z) | Δ ppm, RDB | Formula | RT | Putative Compound 4 |
| 195.0650 | −1.155, 5.5 | C10H11O4+ | 25.46 | 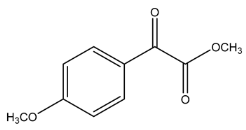 |
| Fragment ions | ||||
| Exp. MS (m/z) | Δ ppm, RDB | Formula | Structural hypothesis | |
| 135.0440 | −0.711, 5.5 | C8H7O2+ | 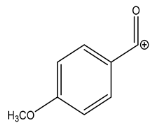 | |
| 121.0283 | −1.289, 5.5 | C7H5O2+ | 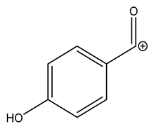 | |
| Exp. MS (m/z) | Δ ppm, RDB | Formula | RT | Putative Compound 5 |
| 266.1134 | −0.498, 6.5 | C12H16N3O4+ | 28.62 | 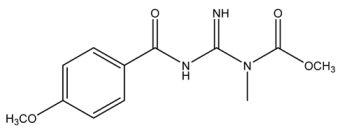 |
| Fragment ions | ||||
| Exp. MS (m/z) | Δ ppm, RDB | Formula | Structural hypothesis | |
| 234.0869 | −1.614, 7.5 | C11H12N3O3+ | 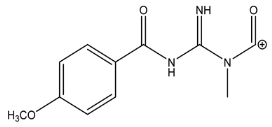 | |
| 135.0437 | −2.710, 5.5 | C8H7O2+ | 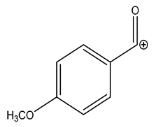 | |
| Exp. MS (m/z) | Δ ppm, RDB | Formula | RT | Putative Compound 6 |
| 236.0487 | −0.567, 7.5 | C10H10N3SO2+ | 29.47 | 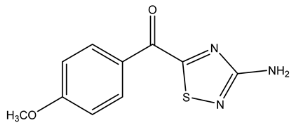 |
| Fragment ions | ||||
| Exp. MS (m/z) | Δ ppm, RDB | Formula | Structural hypothesis | |
| 135.0439 | −1.377, 5.5 | C8H7O2+ | 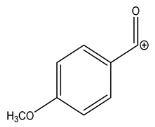 | |
| Exp. MS (m/z) | Δ ppm, RDB | Formula | RT | Putative Compound 7 |
| 222.0695 | −0.447, 6.5 | C10H13N3SO+ | 30.87 | 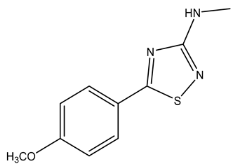 |
| Exp. MS (m/z) | Δ ppm, RDB | Formula | RT | Putative Compound 8 |
| 280.0750 | −0.102, 7.5 | C12H14N3SO3+ | 33.28 | 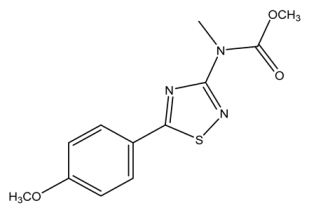 |
| Fragment ions | ||||
| Exp. MS (m/z) | Δ ppm, RDB | Formula | Structural hypothesis | |
| 248.04918 | 1.436, 8.5 | C11H10N3SO2+ | 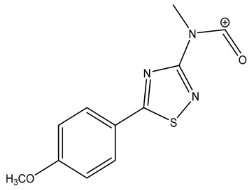 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Casertano, M.; Esposito, E.; Bello, I.; Indolfi, C.; Putra, M.Y.; Di Cesare Mannelli, L.; Ghelardini, C.; Menna, M.; Sorrentino, R.; Cirino, G.; et al. Searching for Novel Sources of Hydrogen Sulfide Donors: Chemical Profiling of Polycarpa aurata Extract and Evaluation of the Anti-Inflammatory Effects. Mar. Drugs 2023, 21, 641. https://doi.org/10.3390/md21120641
Casertano M, Esposito E, Bello I, Indolfi C, Putra MY, Di Cesare Mannelli L, Ghelardini C, Menna M, Sorrentino R, Cirino G, et al. Searching for Novel Sources of Hydrogen Sulfide Donors: Chemical Profiling of Polycarpa aurata Extract and Evaluation of the Anti-Inflammatory Effects. Marine Drugs. 2023; 21(12):641. https://doi.org/10.3390/md21120641
Chicago/Turabian StyleCasertano, Marcello, Erika Esposito, Ivana Bello, Chiara Indolfi, Masteria Yunovilsa Putra, Lorenzo Di Cesare Mannelli, Carla Ghelardini, Marialuisa Menna, Raffaella Sorrentino, Giuseppe Cirino, and et al. 2023. "Searching for Novel Sources of Hydrogen Sulfide Donors: Chemical Profiling of Polycarpa aurata Extract and Evaluation of the Anti-Inflammatory Effects" Marine Drugs 21, no. 12: 641. https://doi.org/10.3390/md21120641
APA StyleCasertano, M., Esposito, E., Bello, I., Indolfi, C., Putra, M. Y., Di Cesare Mannelli, L., Ghelardini, C., Menna, M., Sorrentino, R., Cirino, G., d’Emmanuele di Villa Bianca, R., Imperatore, C., Panza, E., & Mitidieri, E. (2023). Searching for Novel Sources of Hydrogen Sulfide Donors: Chemical Profiling of Polycarpa aurata Extract and Evaluation of the Anti-Inflammatory Effects. Marine Drugs, 21(12), 641. https://doi.org/10.3390/md21120641