Extending the Structural Diversity of Labdane Diterpenoids from Marine-Derived Fungus Talaromyces sp. HDN151403 Using Heterologous Expression

 , , , and
, , , and 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Prediction of the Labdane-Related Diterpenes BGC in Talaromyces sp. HDN151403
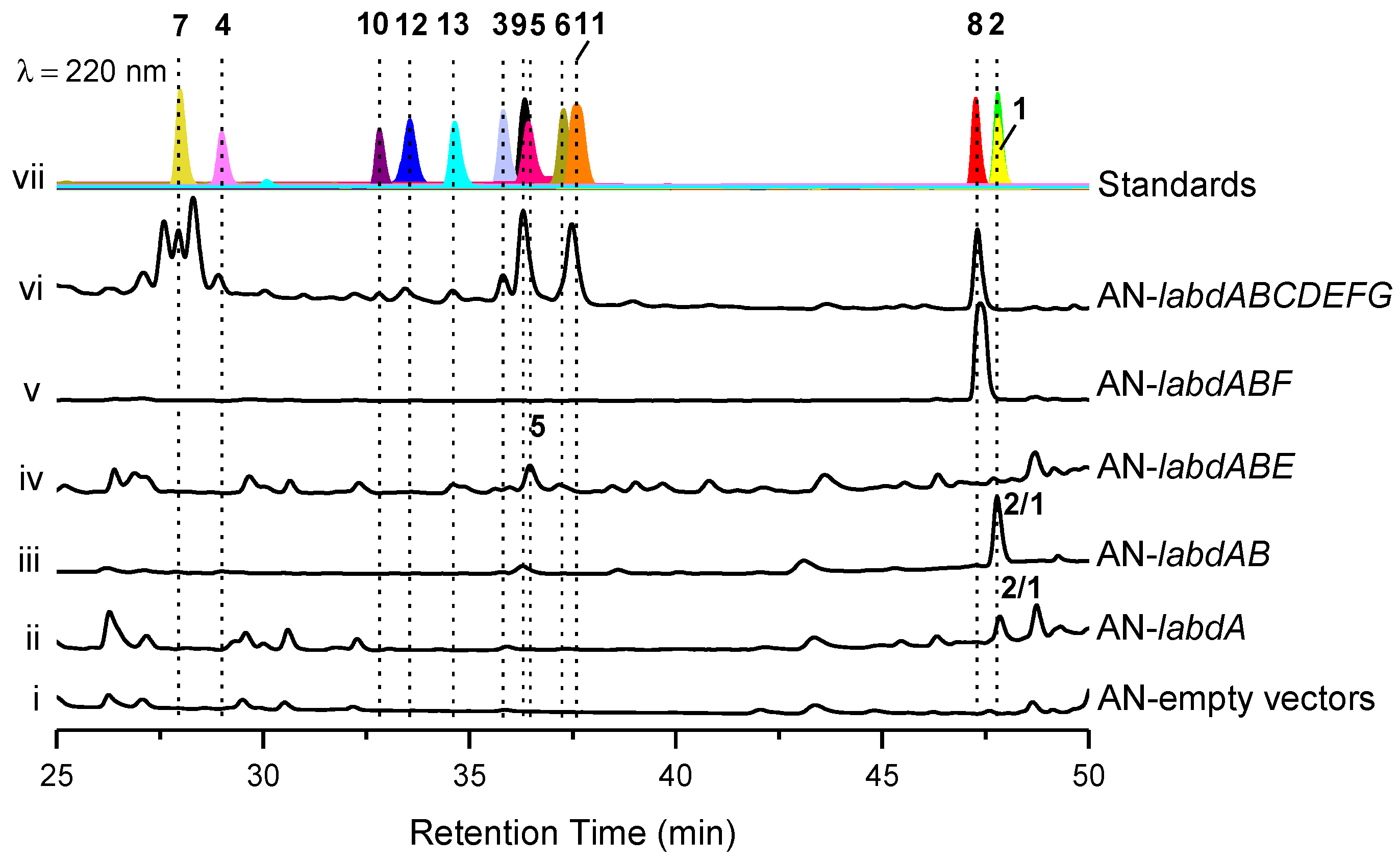
2.2. Heterologous Expression the labd BGC in A. nidulans A1145
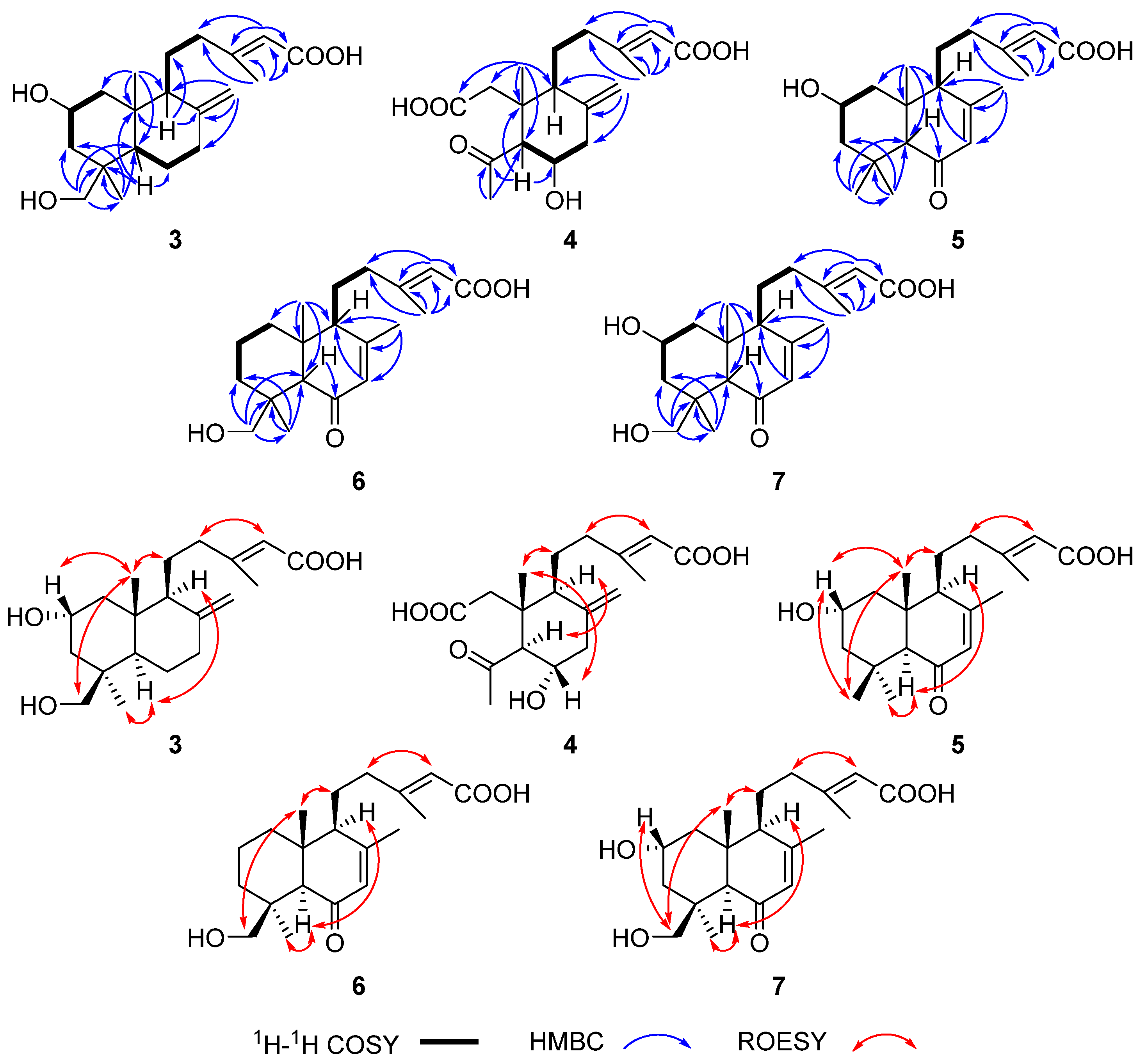
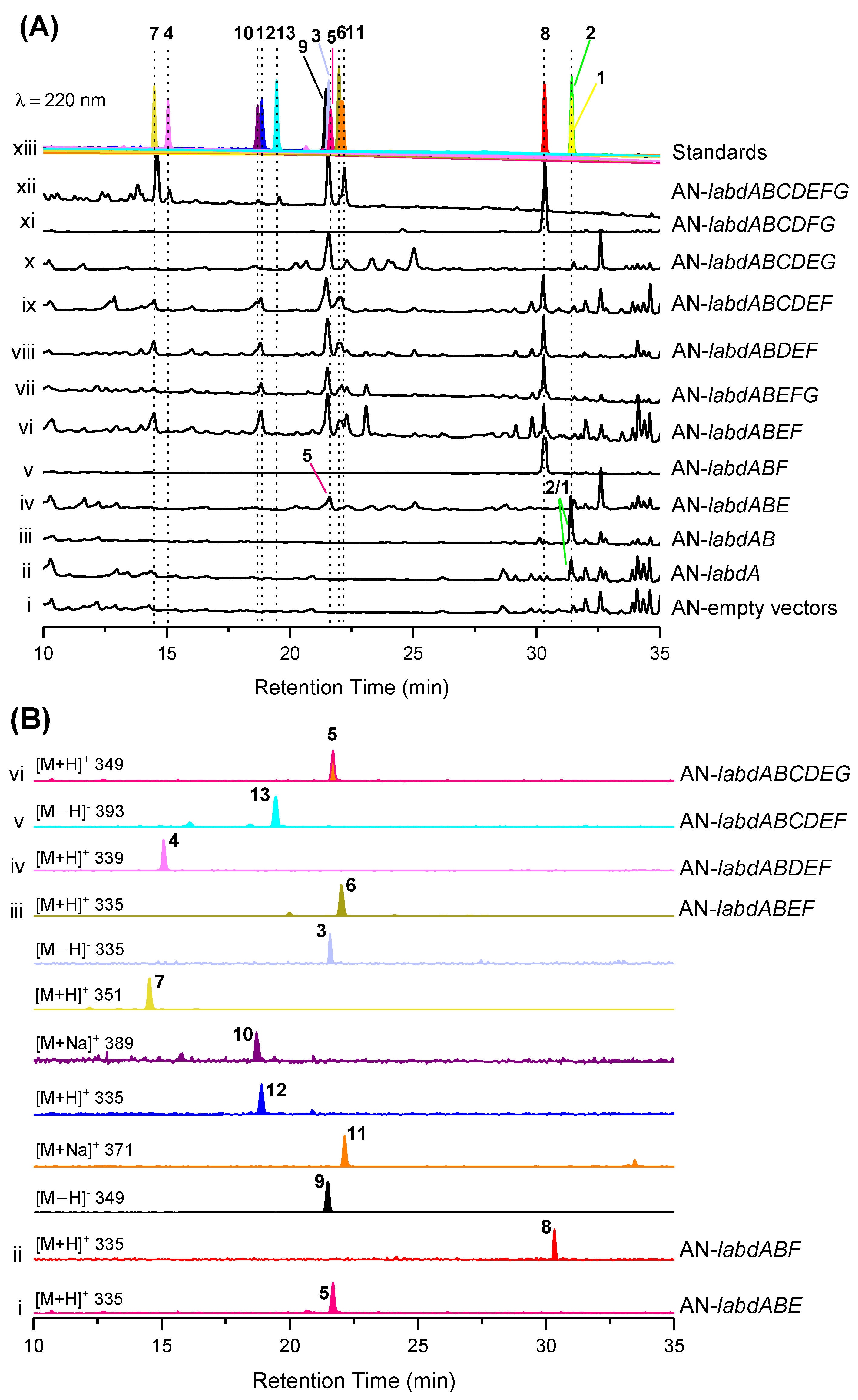
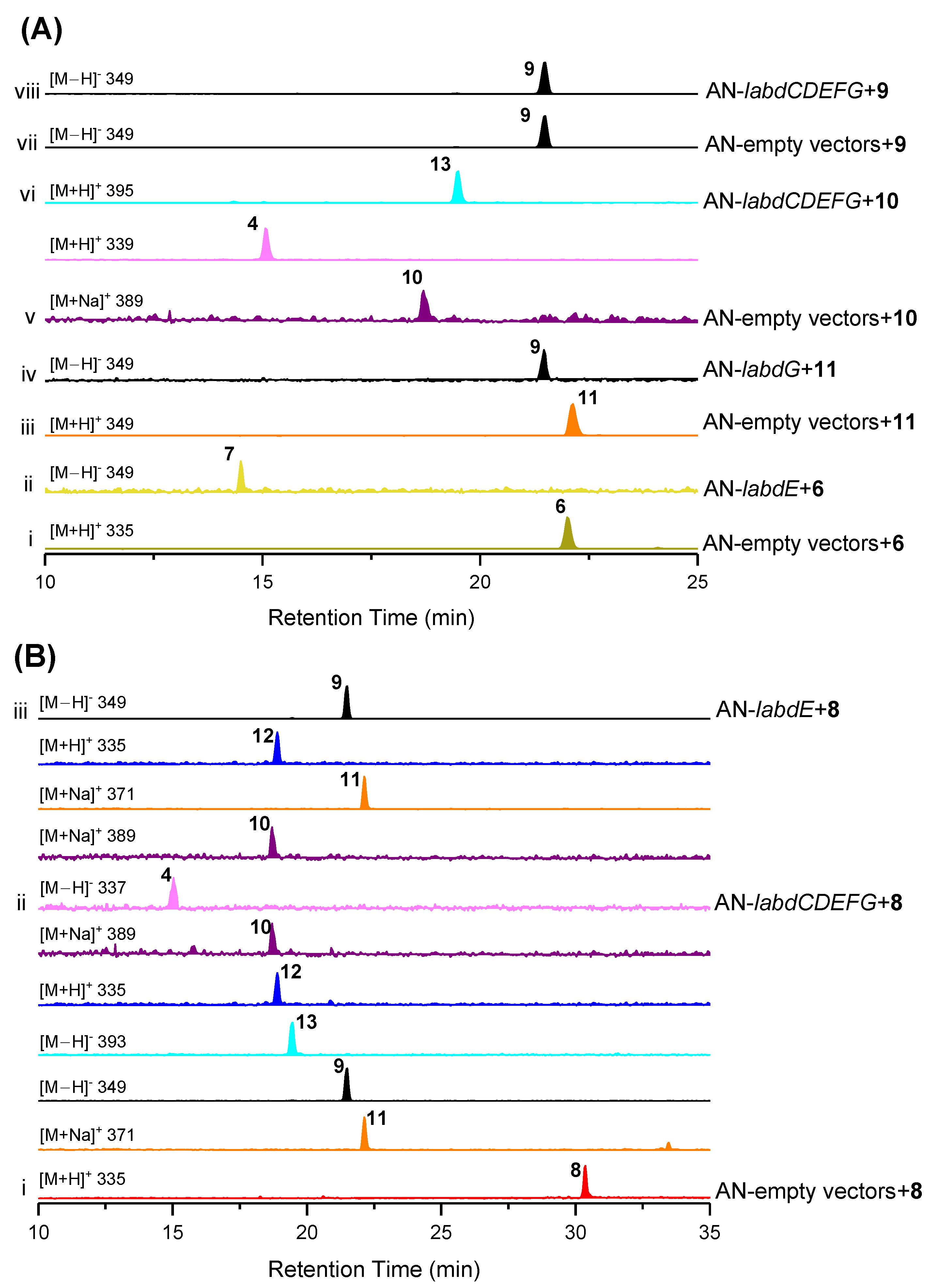
2.3. Heterologous Reconstitution of the Labdane Diterpenes Biosynthetic Pathway
2.4. The Proposed Biosynthetic Pathways
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Materials and Culture Conditions
3.3. Gene Cloning, Plasmid Construction, and Genetic Manipulation
3.4. RNA Extraction and Reverse Transcriptase PCR (RT-PCR)
3.5. Transformation of A. nidulans
3.6. Fermentation and LC/LC−MS Analysis
3.7. Extraction, Isolation, and Purification
3.8. A. nidulans Feeding Experiments
3.9. ECD Calculations
3.10. Antimicrobial Activity Assay
3.11. Cytotoxicity Assay
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chinou, I. Labdanes of natural origin-biological activities (1981–2004). Curr. Med. Chem. 2005, 12, 1295–1317. [Google Scholar] [CrossRef]
- De Souza, N.J.; Dohadwalla, A.N.; Reden, Ü. Forskolin: A labdane diterpenoid with antihypertensive, positive inotropic, platelet aggregation inhibitory, and adenylate cyclase activating properties. Med. Res. Rev. 1983, 3, 201–219. [Google Scholar] [CrossRef]
- Deng, Z.T.; Chen, J.J.; Geng, C.A. ent-Labdane and ent-kaurane diterpenoids from Chelonopsis odontochila with α-glucosidase inhibitory activity. Bioorg. Chem. 2020, 95, 103571. [Google Scholar] [CrossRef] [PubMed]
- Maurya, A.K.; Baliyan, N.; Kumar, R.; Agnihotri, V.K. Synthesis of sulfur-containing analogues of hedychenone, a labdane diterpenoid from Hedychium spicatum. J. Nat. Prod. 2022, 85, 1691–1696. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Pal, M.; Sharma, R.P. Biological activity of the labdane diterpenes. Planta Med. 1999, 65, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Tran, Q.T.N.; Wong, W.S.F.; Chai, C.L.L. Labdane diterpenoids as potential anti-inflammatory agents. Pharmacol. Res. 2017, 124, 43–63. [Google Scholar] [CrossRef]
- Acquaviva, R.; Malfa, G.A.; Loizzo, M.R.; Xiao, J.; Bianchi, S.; Tundis, R. Advances on natural abietane, labdane and clerodane diterpenes as anti-cancer agents: Sources and mechanisms of action. Molecules 2022, 27, 4791. [Google Scholar] [CrossRef]
- Voon, K.J.; Sivasothy, Y.; Sundralingam, U.; Lalmahomed, A.; Goh, A.P. Cytotoxic labdane diterpenes, norlabdane diterpenes and bis-labdanic diterpenes from the Zingiberaceae: A systematic review. Pharmaceuticals 2022, 15, 1517. [Google Scholar] [CrossRef]
- Zhao, L.; Xiang, K.L.; Liu, R.X.; Xie, Z.P.; Zhang, S.M.; Dai, S.J. Anti-inflammatory and anti-viral labdane diterpenoids from the fruits of Forsythia suspensa. Bioorg. Chem. 2020, 96, 103651. [Google Scholar] [CrossRef]
- Liu, Y.; Zhou, Y.Y.; Luo, S.H.; Guo, K.; Zhang, M.W.; Jing, S.X.; Li, C.H.; Hua, J.; Li, S.H. Labdane diterpenoids from the heartwood of Leucosceptrum canum that impact on root growth and seed germination of Arabidopsis thaliana. Phytochemistry 2023, 206, 113531. [Google Scholar] [CrossRef]
- Peters, R.J. Two rings in them all: The labdane-related diterpenoids. Nat. Prod. Rep. 2010, 27, 1521–1530. [Google Scholar] [CrossRef] [PubMed]
- Kildegaard, K.R.; Arnesen, J.A.; Adiego-Perez, B.; Rago, D.; Kristensen, M.; Klitgaard, A.K.; Hansen, E.H.; Hansen, J.; Borodina, I. Tailored biosynthesis of gibberellin plant hormones in yeast. Metab. Eng. 2021, 66, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Komatsu, M.; Ikeda, H. Chemical diversity of labdane-type bicyclic diterpene biosynthesis in Actinomycetales microorganisms. J. Antibiot. 2016, 69, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Rudolf, J.D.; Dong, L.B.; Cao, H.; Hatzos-Skintges, C.; Osipiuk, J.; Endres, M.; Chang, C.Y.; Ma, M.; Babnigg, G.; Joachimiak, A.; et al. Structure of the ent-copalyl diphosphate synthase PtmT2 from Streptomyces platensis CB00739, a bacterial type II diterpene synthase. J. Am. Chem. Soc. 2016, 138, 10905–10915. [Google Scholar] [CrossRef]
- Mitsuhashi, T.; Okada, M.; Abe, I. Identification of chimeric αβγ diterpene synthases possessing both type II terpene cyclase and prenyltransferase activities. ChemBioChem 2017, 18, 2104–2109. [Google Scholar] [CrossRef]
- Stowell, E.A.; Ehrenberger, M.A.; Lin, Y.L.; Chang, C.Y.; Rudolf, J.D. Structure-guided product determination of the bacterial type II diterpene synthase Tpn2. Commun. Chem. 2022, 5, 146. [Google Scholar] [CrossRef]
- Guzman-Trampe, S.M.; Ikeda, H.; Vinuesa, P.; Macias-Rubalcava, M.L.; Esquivel, B.; Centeno-Leija, S.; Tapia-Cabrera, S.M.; Mora-Herrera, S.I.; Ruiz-Villafan, B.; Rodriguez-Sanoja, R.; et al. Production of distinct labdane-type diterpenoids using a novel cryptic labdane-like cluster from Streptomyces thermocarboxydus K155. Appl. Microbiol. Biotechnol. 2020, 104, 741–750. [Google Scholar] [CrossRef]
- Li, R.; Morris-Natschke, S.L.; Lee, K.H. Clerodane diterpenes: Sources, structures, and biological activities. Nat. Prod. Rep. 2016, 33, 1166–1226. [Google Scholar] [CrossRef]
- Roncero, A.M.; Tobal, I.E.; Moro, R.F.; Diez, D.; Marcos, I.S. Isolation and biosynthesis Sources, structures, nomenclature and biological activities. Nat. Prod. Rep. 2018, 35, 955–991. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Zha, W.L.; Zhu, J.X.; Zheng, C.; Zi, J.C. A review: Biosynthesis of plant-derived labdane-related diterpenoids. Chin. J. Nat. Med. 2021, 19, 666–674. [Google Scholar] [CrossRef]
- Chiang, Y.M.; Lin, T.S.; Wang, C.C.C. Total heterologous biosynthesis of fungal natural products in Aspergillus nidulans. J. Nat. Prod. 2022, 85, 2484–2518. [Google Scholar] [CrossRef] [PubMed]
- Gilchrist, C.L.M.; Booth, T.J.; van Wersch, B.; van Grieken, L.; Medema, M.H.; Chooi, Y.-H.; Ouangraoua, A. cblaster: A remote search tool for rapid identification and visualization of homologous gene clusters. Bioinform. Adv. 2021, 1, vbab016. [Google Scholar] [CrossRef]
- Gilchrist, C.L.M.; Chooi, Y.-H. clinker & clustermap.js: Automatic generation of gene cluster comparison figures. Bioinformatics 2021, 37, 2473–2475. [Google Scholar] [CrossRef] [PubMed]
- Grigoriev, I.V.; Nikitin, R.; Haridas, S.; Kuo, A.; Ohm, R.; Otillar, R.; Riley, R.; Salamov, A.; Zhao, X.; Korzeniewski, F.; et al. MycoCosm portal: Gearing up for 1000 fungal genomes. Nucleic Acids Res. 2014, 42, D699–D704. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.W.; Cheng, Y.B.; Liaw, C.C.; Chen, C.H.; Guh, J.H.; Hwang, T.L.; Tsai, J.S.; Wang, W.B.; Shen, Y.C. Bioactive diterpenes from Callicarpa longissima. J. Nat. Prod. 2012, 75, 689–693. [Google Scholar] [CrossRef]
- Braun, S.; Breitenbach, H. Strukturaufklärung einer neuen diterpensäure aus Metasequoia glyptostroboides mit hilfe der 13C-NMR-spektroskopie. Tetrahedron 1977, 33, 145–150. [Google Scholar] [CrossRef]
- Frija, L.M.T.; Garcia, H.; Rodrigues, C.; Martins, I.; Candeias, N.R.; André, V.; Duarte, M.T.; Pereira, C.S.; Afonso, C.A.M. Short synthesis of the natural product 3β-hydroxy-labd-8(17)-en-15-oic acid via microbial transformation of labdanolic acid. Phytochem. Lett. 2013, 6, 165–169. [Google Scholar] [CrossRef]
- Bastard, J.; Duc, D.K.; Fetizon, M.; Francis, M.J.; Grant, P.K.; Weavers, R.T.; Kaneko, C.; Baddeley, G.V.; Bernassau, J.-M.; Burfitt, I.R.; et al. CMR spectroscopy of labdanic diterpenes and related substances. J. Nat. Prod. 1984, 47, 592–599. [Google Scholar] [CrossRef]
- Zhou, K.; Zhao, X.-L.; Han, L.-P.; Cao, M.-M.; Chen, C.; Shi, B.-Z.; Luo, D.-Q. Paecilomycines A and B, novel diterpenoids, isolated from insect-pathogenic fungi Paecilomyces sp. ACCC 37762. Helv. Chim. Acta 2015, 98, 642–649. [Google Scholar] [CrossRef]
- Qi, B.; Jia, F.; Luo, Y.; Ding, N.; Li, S.; Shi, F.; Hai, Y.; Wang, L.; Zhu, Z.X.; Liu, X.; et al. Two new diterpenoids from Penicillium chrysogenum MT-12, an endophytic fungus isolated from Huperzia serrata. Nat. Prod. Res. 2022, 36, 814–821. [Google Scholar] [CrossRef]
- Li, Y.; Liu, W.; Han, S.; Zhang, J.; Xu, W.; Li, Q.; Cheng, Z. Penitholabene, a rare 19-nor labdane-type diterpenoid from the deep-sea-derived fungus Penicillium thomii YPGA3. Fitoterapia 2020, 146, 104691. [Google Scholar] [CrossRef] [PubMed]
- Nuansri, S.; Rukachaisirikul, V.; Muanprasat, C.; Phongpaichit, S.; Preedanon, S.; Sakayaroj, J. Furanone, morpholinone and tetrahydrofuran derivatives from the marine-derived fungus Talaromyces sp. PSU-MF07. Nat. Prod. Res. 2022, 37, 3434–3442. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Z.-J.; Huang, J.; Yan, Y.; Wang, L.; Wang, Z.; Yang, J.; Luo, J.; Li, J.; Huang, S.-X. Isolation and biosynthesis of labdanmycins: Four new labdane diterpenes from endophytic Streptomyces. Org. Chem. Front. 2018, 5, 1272–1279. [Google Scholar] [CrossRef]
- Sun, C.; Liu, Q.; Shah, M.; Che, Q.; Zhang, G.; Zhu, T.; Zhou, J.; Rong, X.; Li, D. Talaverrucin A, Heterodimeric Oxaphenalenone from Antarctica Sponge-Derived Fungus Talaromyces sp. HDN151403, Inhibits Wnt/β-Catenin Signaling Pathway. Org. Lett. 2022, 24, 3993–3997. [Google Scholar] [CrossRef]
- Yee, D.A.; Tang, Y. Investigating fungal biosynthetic pathways using heterologous gene expression: Aspergillus nidulans as a heterologous host. In Engineering Natural Product Biosynthesis: Methods in Molecular Biology; Skellam, E., Ed.; Springer: New York, NY, USA, 2022; Volume 2489, pp. 41–52. [Google Scholar] [CrossRef]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Petersson, G.A.; Nakatsuji, H.; et al. Gaussian 16, Revision B.01; Gaussian, Inc.: Wallingford, CT, USA, 2016. [Google Scholar]
- Bruhn, T.; Schaumlöffel, A.; Hemberger, Y.; Bringmann, G. SpecDis: Quantifying the comparison of calculated and experimental electronic circular dichroism spectra. Chirality 2013, 25, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Dentinger, B.T.M.; Nielson, J.R.; Peterson, R.T.; Winter, J.M. Emerimicins V–X, 15-residue peptaibols discovered from an Acremonium sp. through integrated genomic and chemical approaches. J. Nat. Prod. 2021, 84, 1113–1126. [Google Scholar] [CrossRef]
- Zhang, A.; Qi, X.; Du, F.; Zhang, G.; Li, D.; Li, J. PNSA, a novel C-terminal inhibitor of HSP90, reverses epithelial–mesenchymal transition and suppresses metastasis of breast cancer cells in vitro. Mar. Drugs 2021, 19, 117. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Schueffler, A.; Anke, T. Fungal natural products in research and development. Nat. Prod. Rep. 2014, 31, 1425–1448. [Google Scholar] [CrossRef]
- Brakhage, A.A.; Schroeckh, V. Fungal secondary metabolites—Strategies to activate silent gene clusters. Fungal Genet. Biol. 2011, 48, 15–22. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | 3 | 4 | ||
|---|---|---|---|---|
| δC, Mult | δH (J in Hz) | δC, Mult | δH (J in Hz) | |
| 1a | 49.0, CH2 | 2.08, m | 42.9, CH2 | 2.49, d (15.7) |
| 1b | 1.00, m | 2.14, d (15.7) | ||
| 2 | 65.5, CH | 3.81, m | 174.3, C | – |
| 3a | 45.0, CH2 | 2.17, m | – | – |
| 3b | 0.89, m | |||
| 4 | 41.5, C | – | 215.1, C | – |
| 5 | 56.9, CH | 1.26, m | 63.6, CH | 3.39, d (11.0) |
| 6a | 25.2, CH2 | 1.85, m | 71.5, CH | 3.85, td (11.0, 5.0) |
| 6b | 1.34, m | |||
| 7a | 39.5, CH2 | 2.42, m | 47.6, CH2 | 2.65, dd (12.0, 5.0) |
| 7b | 1.96, m | 2.02, overlapped | ||
| 8 | 148.7, C | – | 145.3, C | – |
| 9 | 57.4, CH | 1.70, m | 49.5, CH | 2.27, m |
| 10 | 41.8, C | – | 42.0, C | – |
| 11a | 23.0, CH2 | 1.75, m | 23.8, CH2 | 1.82, m |
| 11b | 1.57, m | 1.58, m | ||
| 12a | 40.7, CH2 | 2.31, m | 40.7, CH2 | 2.29, m |
| 12b | 2.02, m | 2.02, overlapped | ||
| 13 | 161.9, C | – | 161.8, C | – |
| 14 | 116.8, CH | 5.62, d (1.2) | 116.8, CH | 5.64, d (1.5) |
| 15 | 170.2, C | – | 170.3, C | – |
| 16 | 18.9, CH3 | 2.14, d (1.2) | 19.0, CH3 | 2.15 d (1.5) |
| 17a | 107.8, CH2 | 4.90, s | 111.0, CH2 | 5.08, s |
| 17b | 4.57, s | 4.75, s | ||
| 18a | 65.5, CH2 | 3.61, d (11.5) | – | – |
| 18b | 3.29, d (11.5) | |||
| 19 | 28.1, CH3 | 1.01, s | 35.2, CH3 | 2.32, s |
| 20 | 16.7, CH3 | 0.74, s | 17.9, CH3 | 0.81, s |
| No. | 5 | 6 | 7 | |||
|---|---|---|---|---|---|---|
| δC, Mult | δH (J in Hz) | δC, Mult | δH (J in Hz) | δC, Mult | δH (J in Hz) | |
| 1a | 48.4, CH2 | 2.16, m | 39.6, CH2 | 1.91, m | 48.3, CH2 | 2.17, m |
| 1b | 1.22, m | 1.35, m | 1.25, m | |||
| 2a | 64.6, CH | 3.83 m | 18.8, CH2 | 1.57, m | 64.1, CH | 3.82, m |
| 2b | 1.50, m | |||||
| 3a | 52.6, CH2 | 1.65, m | 37.2, CH2 | 1.94, m | 45.6, CH2 | 2.25, m |
| 3b | 1.14, m | 0.96, m | 0.91, m | |||
| 4 | 35.1, C | – | 39.0, C | – | 41.0, C | – |
| 5 | 63.6, CH | 2.21, s | 65.5, CH | 2.43, s | 64.6, CH | 2.44, s |
| 6 | 202.0, C | – | 203.0, C | – | 202.1, C | – |
| 7 | 129.2, CH | 5.77, s | 128.8, CH | 5.76, s | 129.0, CH | 5.78, s |
| 8 | 161.9, C | – | 162.6, C | – | 162.4, C | – |
| 9 | 57.0, CH | 2.32, m | 57.2, CH | 2.27, m | 57.2, CH | 2.35, m |
| 10 | 45.8, C | – | 44.5, C | – | 45.7, C | – |
| 11a | 26.5, CH2 | 1.69, m | 26.5, CH2 | 1.66, m | 26.6, CH2 | 1.70, m |
| 11b | 1.61, m | |||||
| 12a | 43.9, CH2 | 2.49, m | 43.9, CH2 | 2.48, m | 43.9, CH2 | 2.49, m |
| 12b | 2.27, m | 2.25, m | 2.29, m | |||
| 13 | 160.5, C | – | 160.6, C | – | 160.5, C | – |
| 14 | 117.5, CH | 5.74, d (1.2) | 117.5, CH | 5.73 d (1.2) | 117.5, CH | 5.74, d (1.2) |
| 15 | 170.1, C | – | 170.1, C | – | 170.0, C | – |
| 16 | 19.0, CH3 | 2.20, d (1.2) | 19.0, CH3 | 2.18, d (1.2) | 19.0, CH3 | 2.20, d (1.2) |
| 17 | 22.3, CH3 | 2.02 s | 22.4, CH3 | 2.01, s | 22.3, CH3 | 2.02, s |
| 18a | 22.9, CH3 | 1.20, s | 63.7, CH2 | 4.10, d (11.0) | 64.3, CH2 | 4.11, d (11.0) |
| 18b | 3.63, d (11.0) | 3.57, d (11.0) | ||||
| 19 | 33.8, CH3 | 1.17, s | 27.3, CH3 | 1.15, s | 27.5, CH3 | 1.20, s |
| 20 | 15.9, CH3 | 0.91, s | 15.8, CH3 | 0.84, s | 16.7, CH3 | 0.88, s |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, F.; Ma, C.; Che, Q.; Zhu, T.; Zhang, G.; Li, D. Extending the Structural Diversity of Labdane Diterpenoids from Marine-Derived Fungus Talaromyces sp. HDN151403 Using Heterologous Expression. Mar. Drugs 2023, 21, 628. https://doi.org/10.3390/md21120628
Zhang F, Ma C, Che Q, Zhu T, Zhang G, Li D. Extending the Structural Diversity of Labdane Diterpenoids from Marine-Derived Fungus Talaromyces sp. HDN151403 Using Heterologous Expression. Marine Drugs. 2023; 21(12):628. https://doi.org/10.3390/md21120628
Chicago/Turabian StyleZhang, Falei, Chuanteng Ma, Qian Che, Tianjiao Zhu, Guojian Zhang, and Dehai Li. 2023. "Extending the Structural Diversity of Labdane Diterpenoids from Marine-Derived Fungus Talaromyces sp. HDN151403 Using Heterologous Expression" Marine Drugs 21, no. 12: 628. https://doi.org/10.3390/md21120628
APA StyleZhang, F., Ma, C., Che, Q., Zhu, T., Zhang, G., & Li, D. (2023). Extending the Structural Diversity of Labdane Diterpenoids from Marine-Derived Fungus Talaromyces sp. HDN151403 Using Heterologous Expression. Marine Drugs, 21(12), 628. https://doi.org/10.3390/md21120628