

Deep-Sea Sponges and Corals off the Western Coast of Florida—Intracellular Mechanisms of Action of Bioactive Compounds and Technological Advances Supporting the Drug Discovery Pipeline
 , , , ,
, , , , 
Abstract
1. Introduction
1.1. Marine Corals and Sponges, the Supply Issue, and Marine Ecosystem Preservation
1.2. Deep-Sea Marine Resources off the Western Coast of Florida and Purpose of this Study
2. Methods
2.1. Data Mining from the National Oceanic and Atmospheric Administration (NOAA) Portal
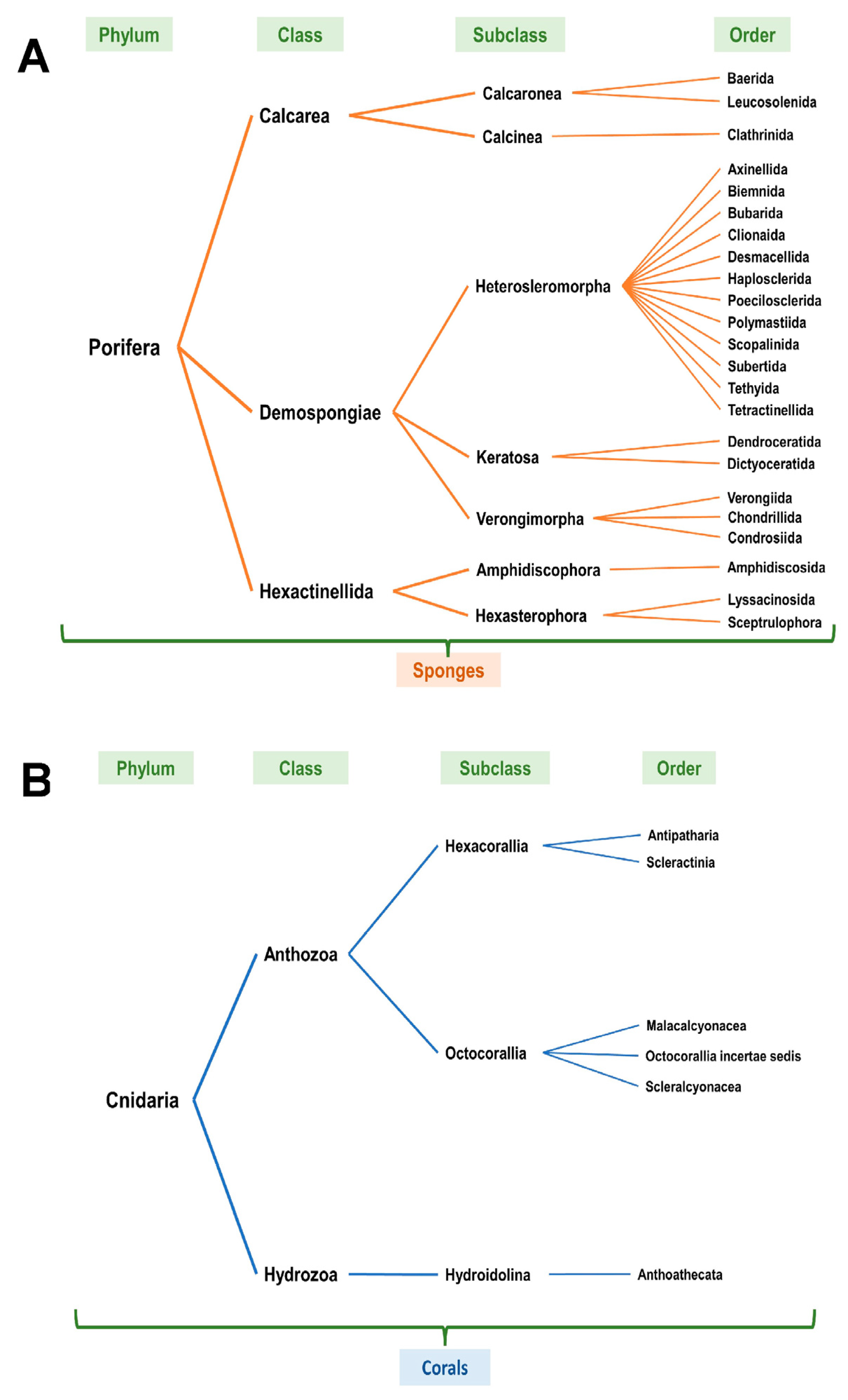
2.2. WoRMS Database and World Porifera Database
2.3. PubMed Literature Survey for Intracellular Mechanisms of Action
2.4. PubMed Literature Survey for Technological Advances Supporting the Drug Discovery Pipeline
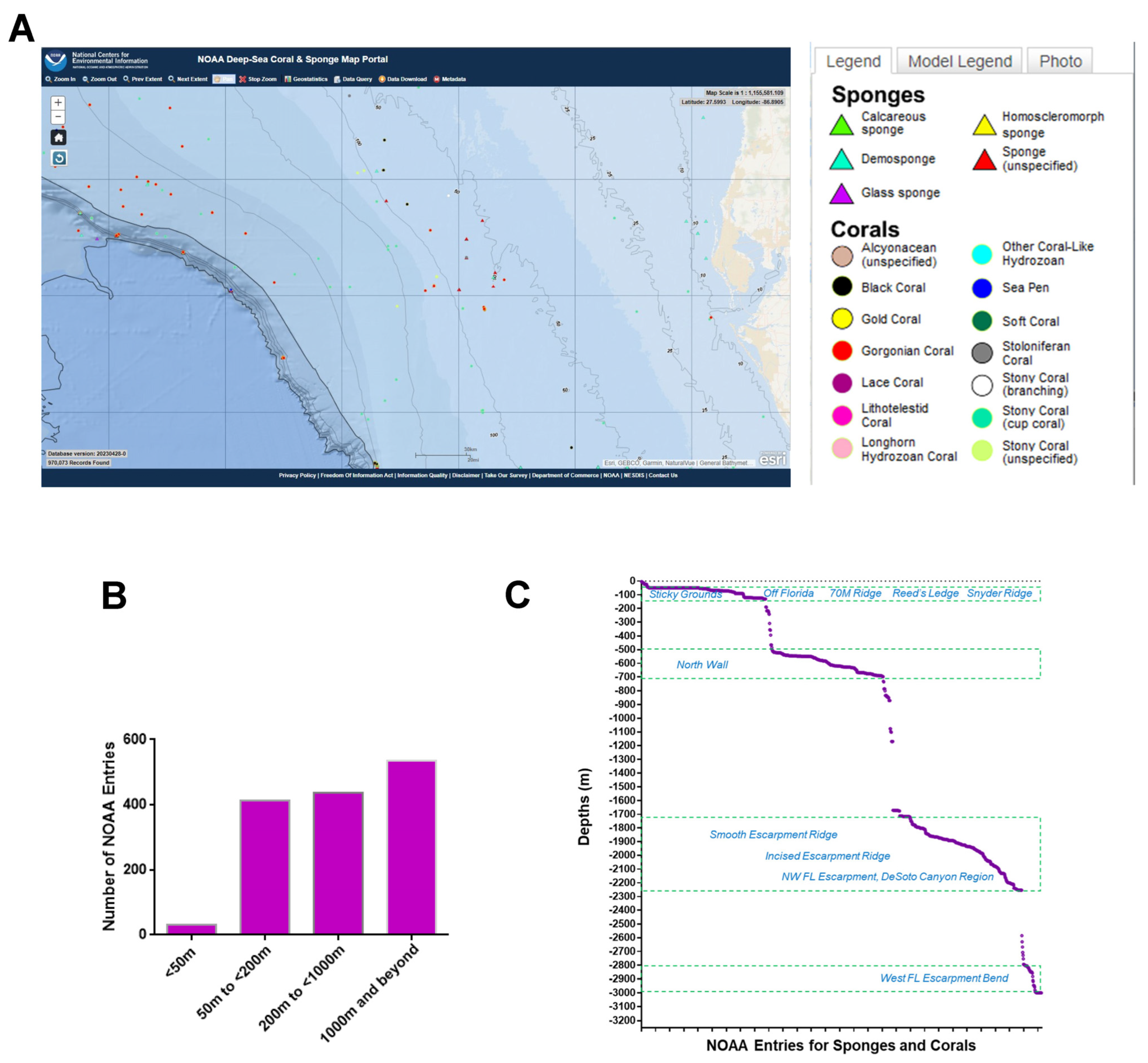
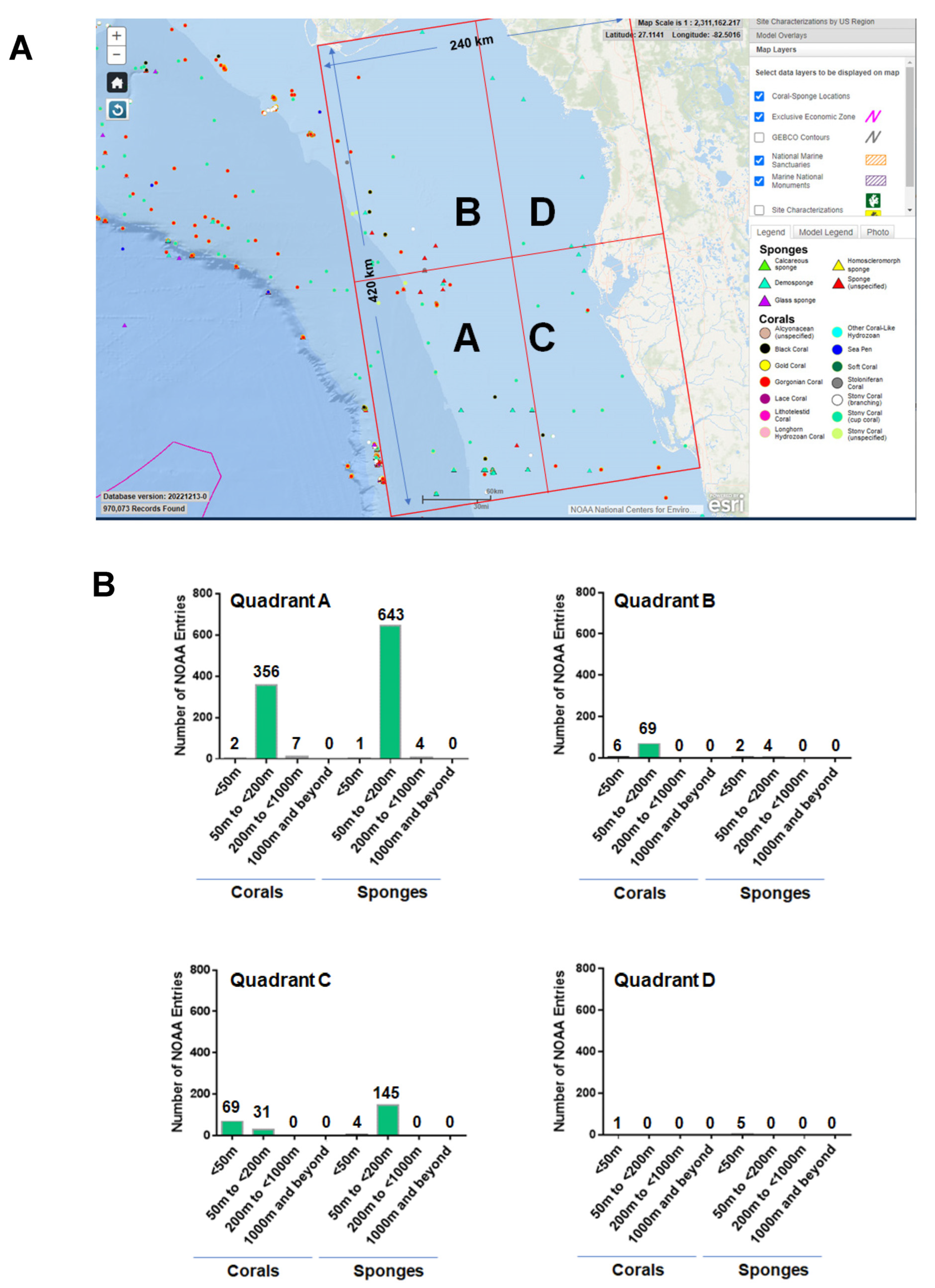
3. Off the Western Coast of Florida—NOAA Analyses
3.1. NOAA—Standard Data Download
3.2. NOAA—Manual Data Curation
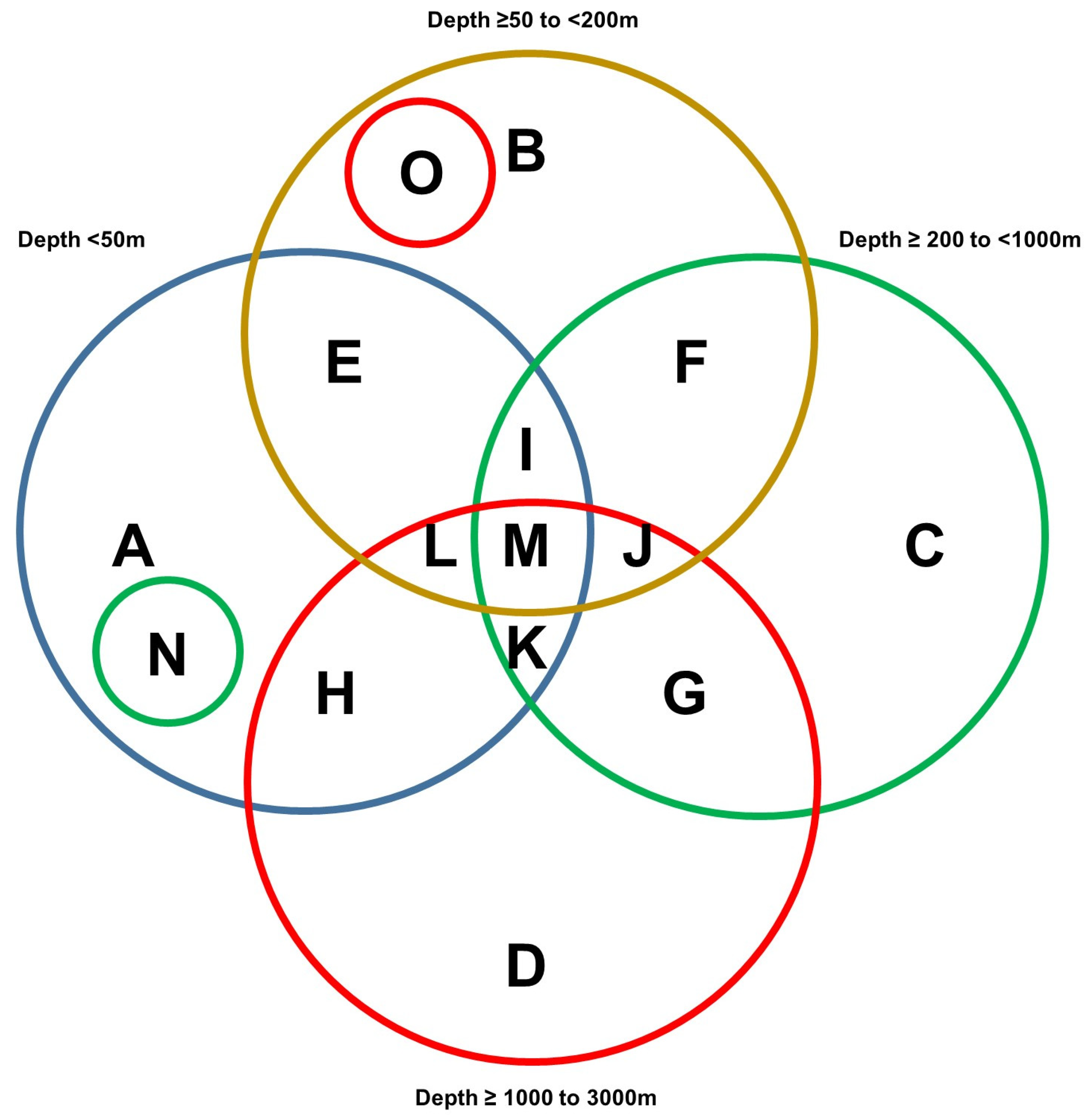
3.3. NOAA—Taxonomic Analyses
4. Intracellular Mechanisms of Action of Bioactive Compounds
4.1. Bioactive Compounds in Corals
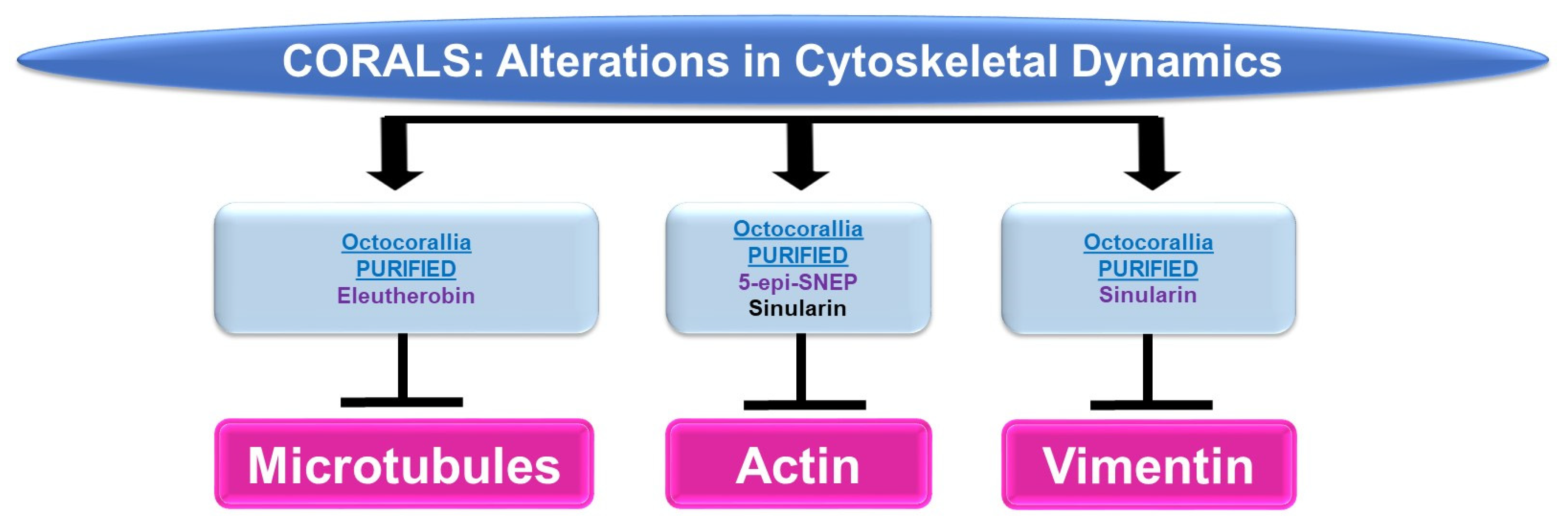
4.1.1. Cytoskeletal Alterations
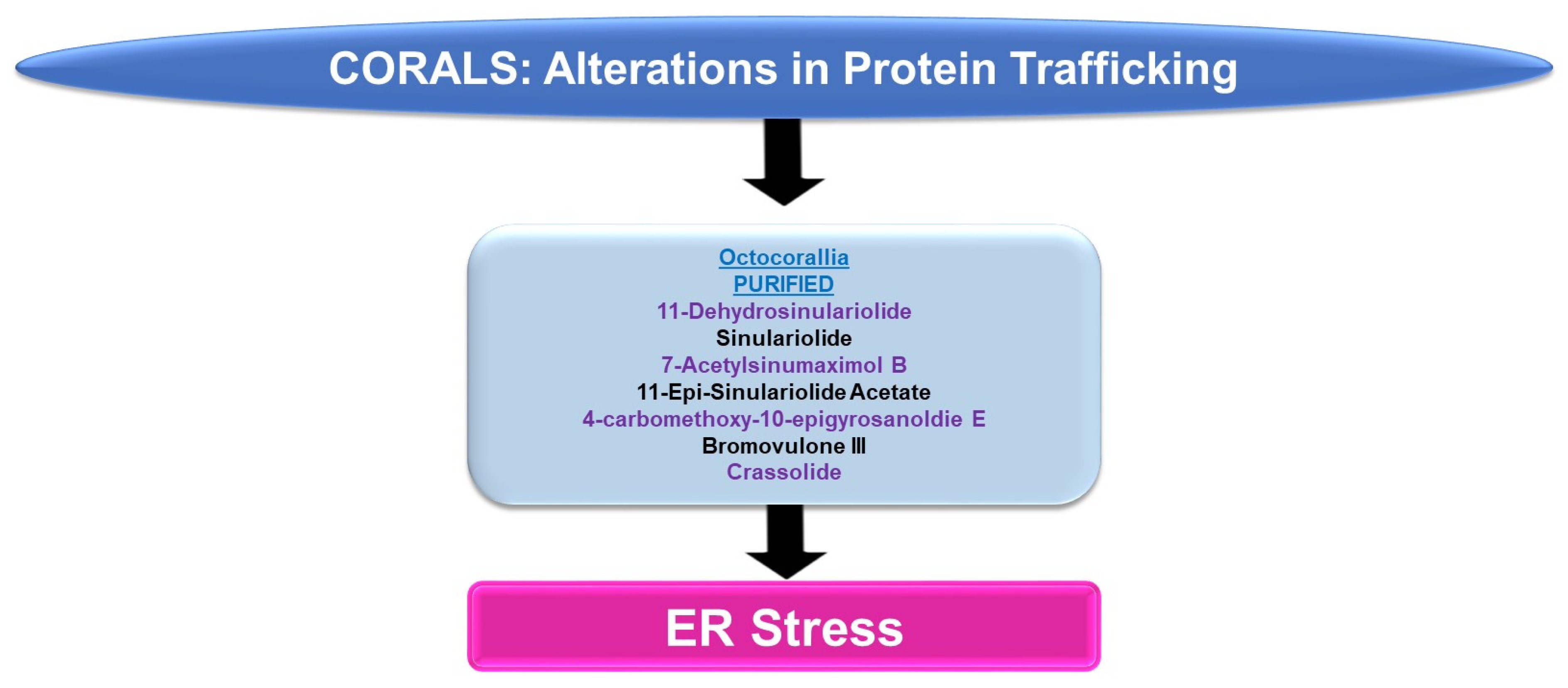
4.1.2. Protein Trafficking Alterations
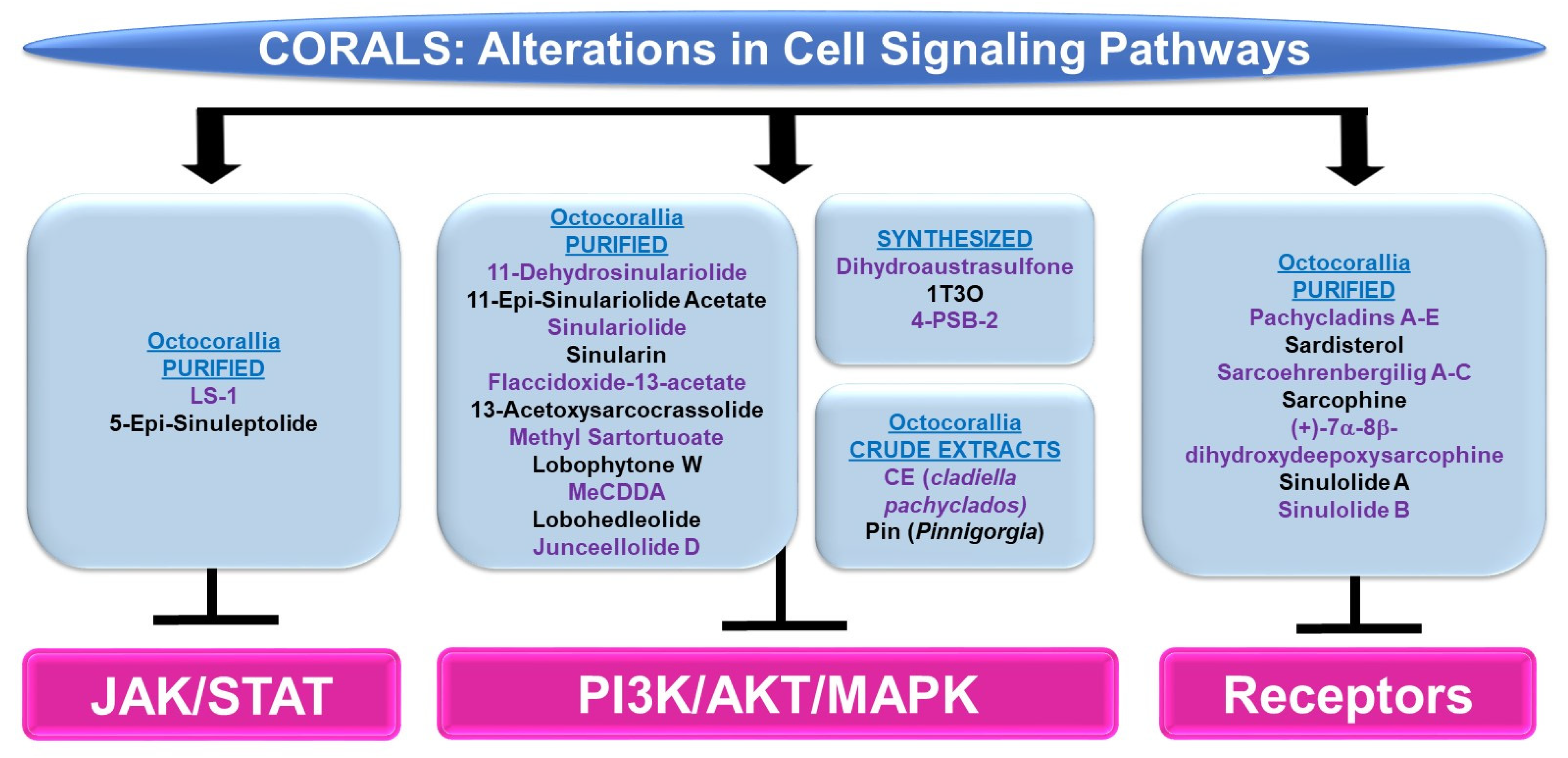
4.1.3. Cell Signaling Alterations
4.2. Bioactive Compounds in Sponges
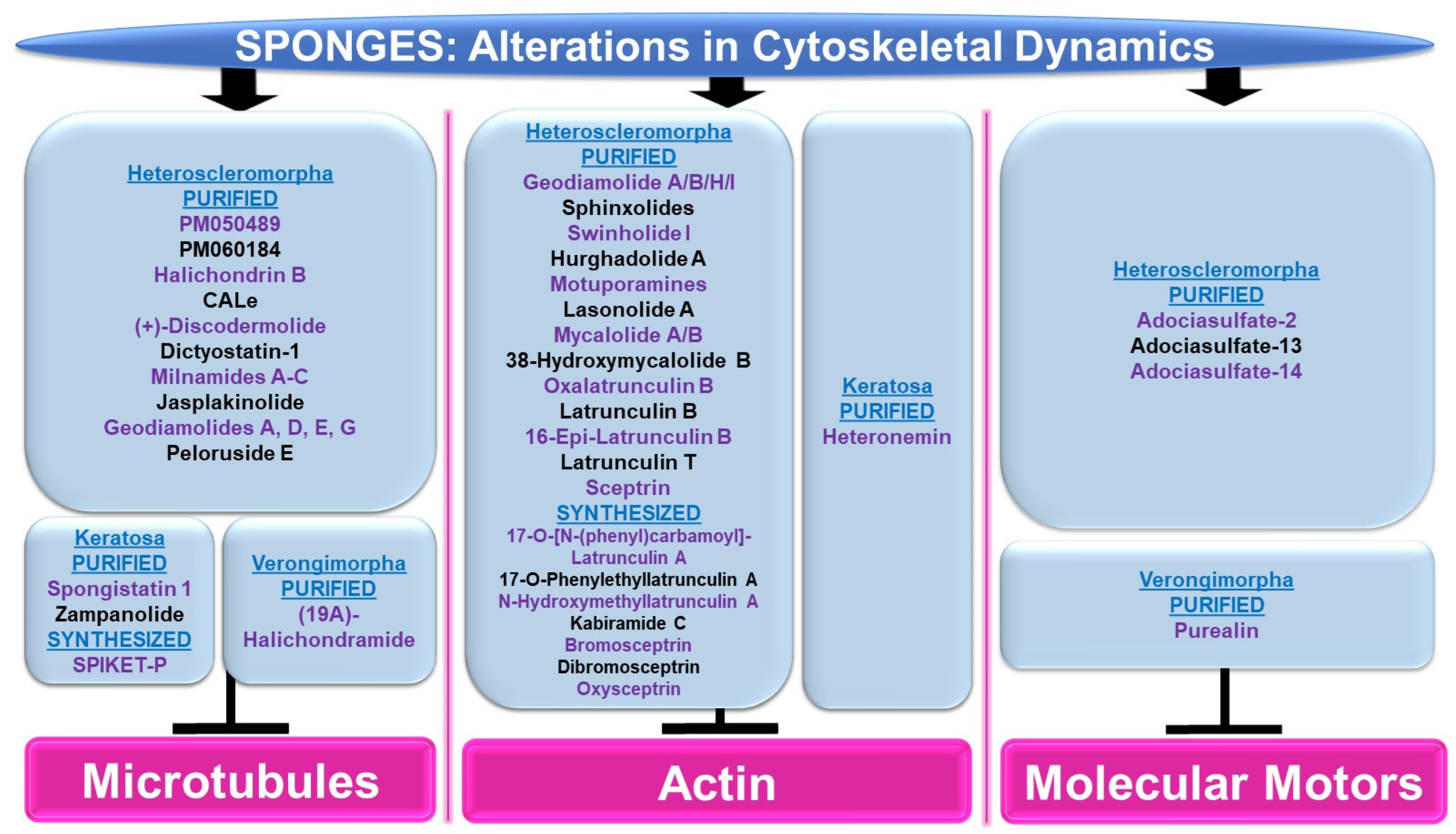
4.2.1. Cytoskeletal Alterations
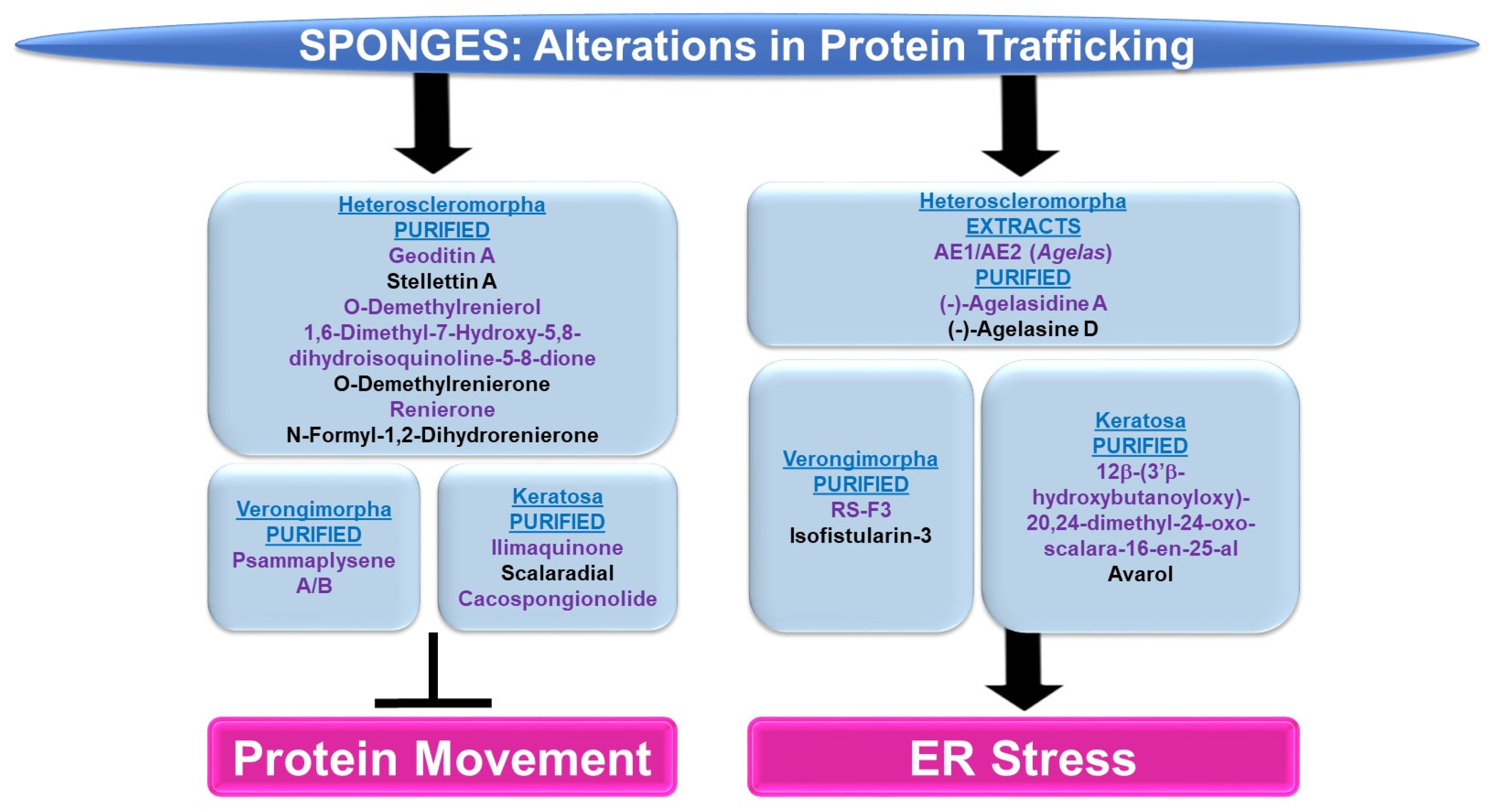
4.2.2. Protein Trafficking Alterations
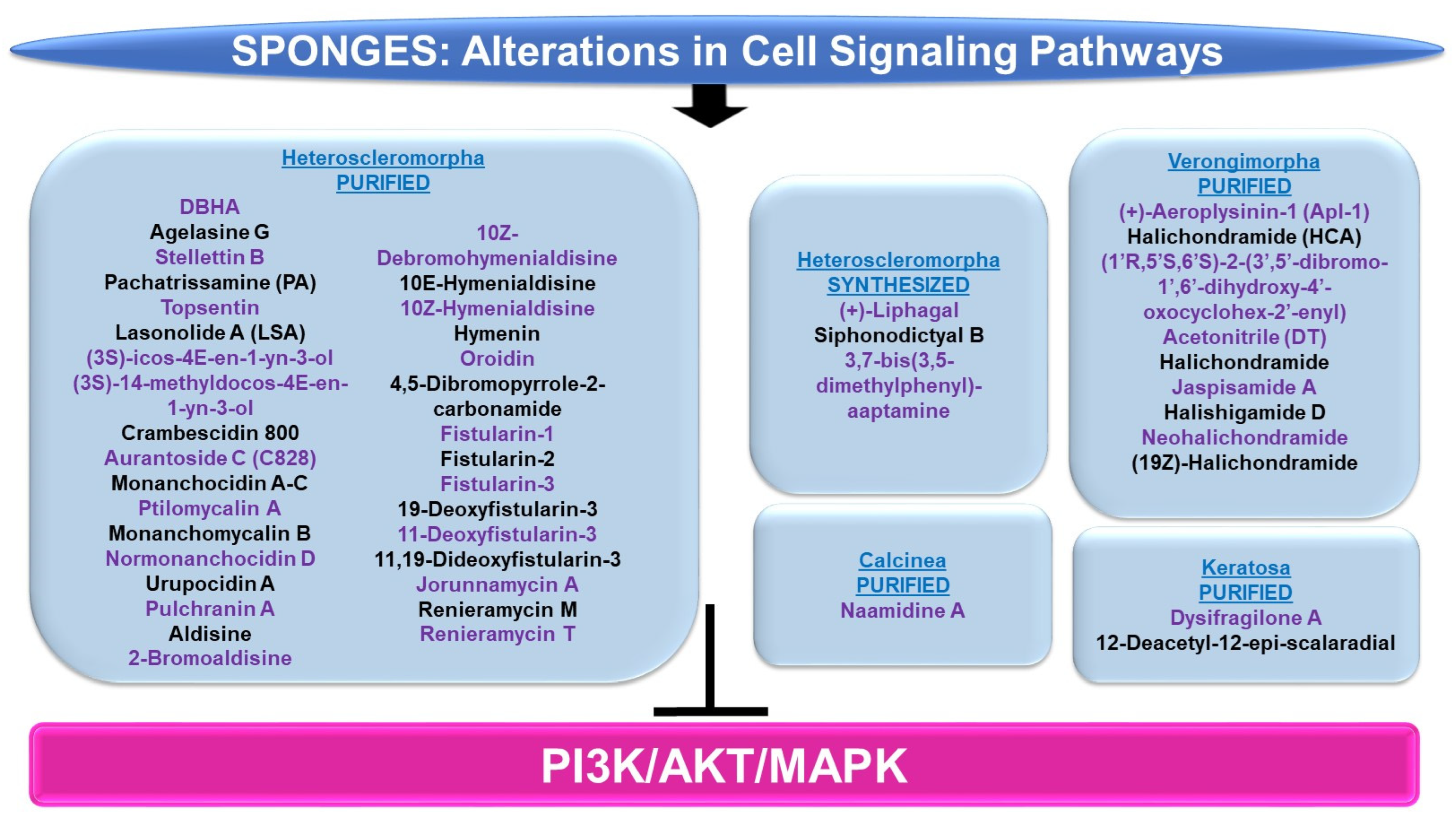
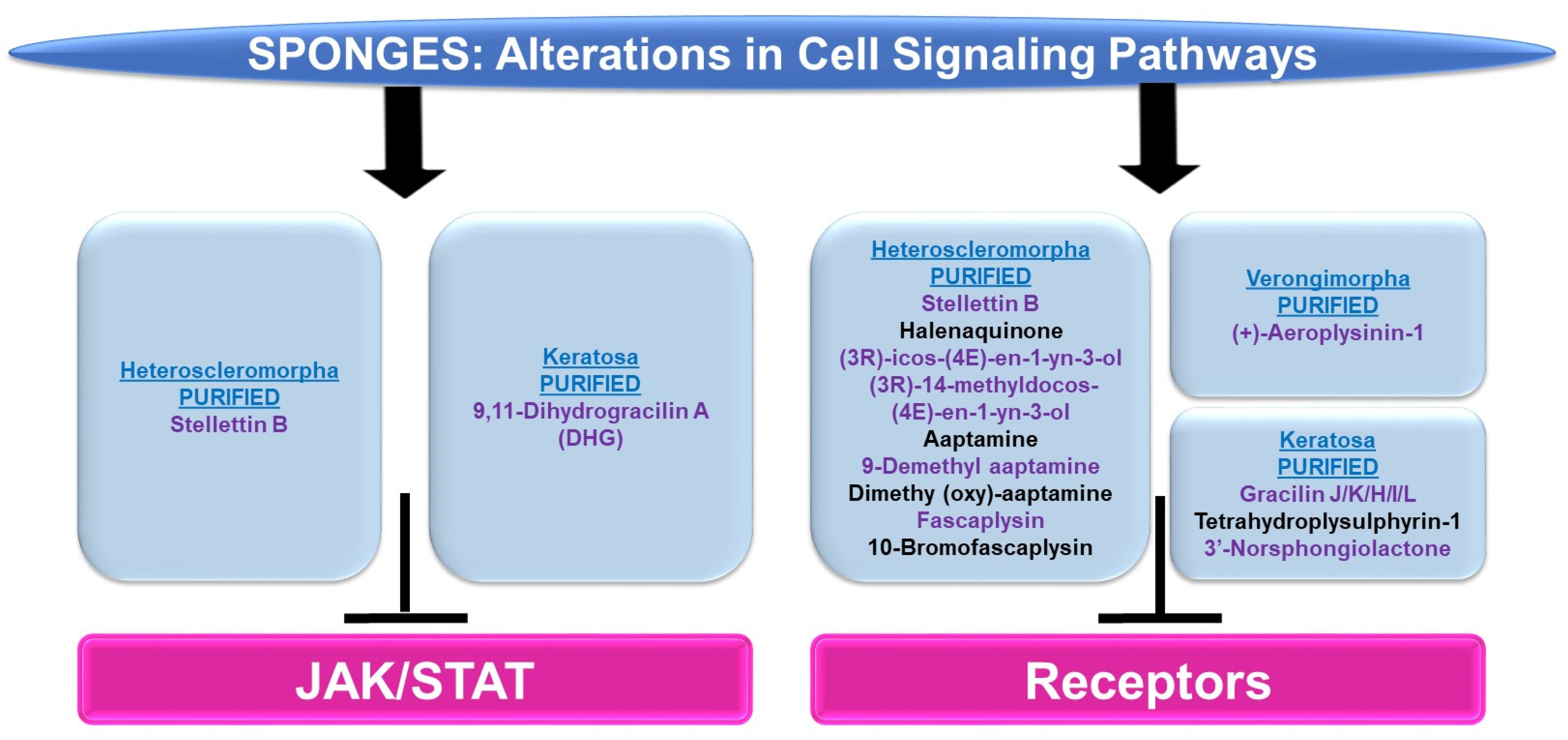
4.2.3. Cell Signaling Alterations
4.2.4. Calcarea Sponge
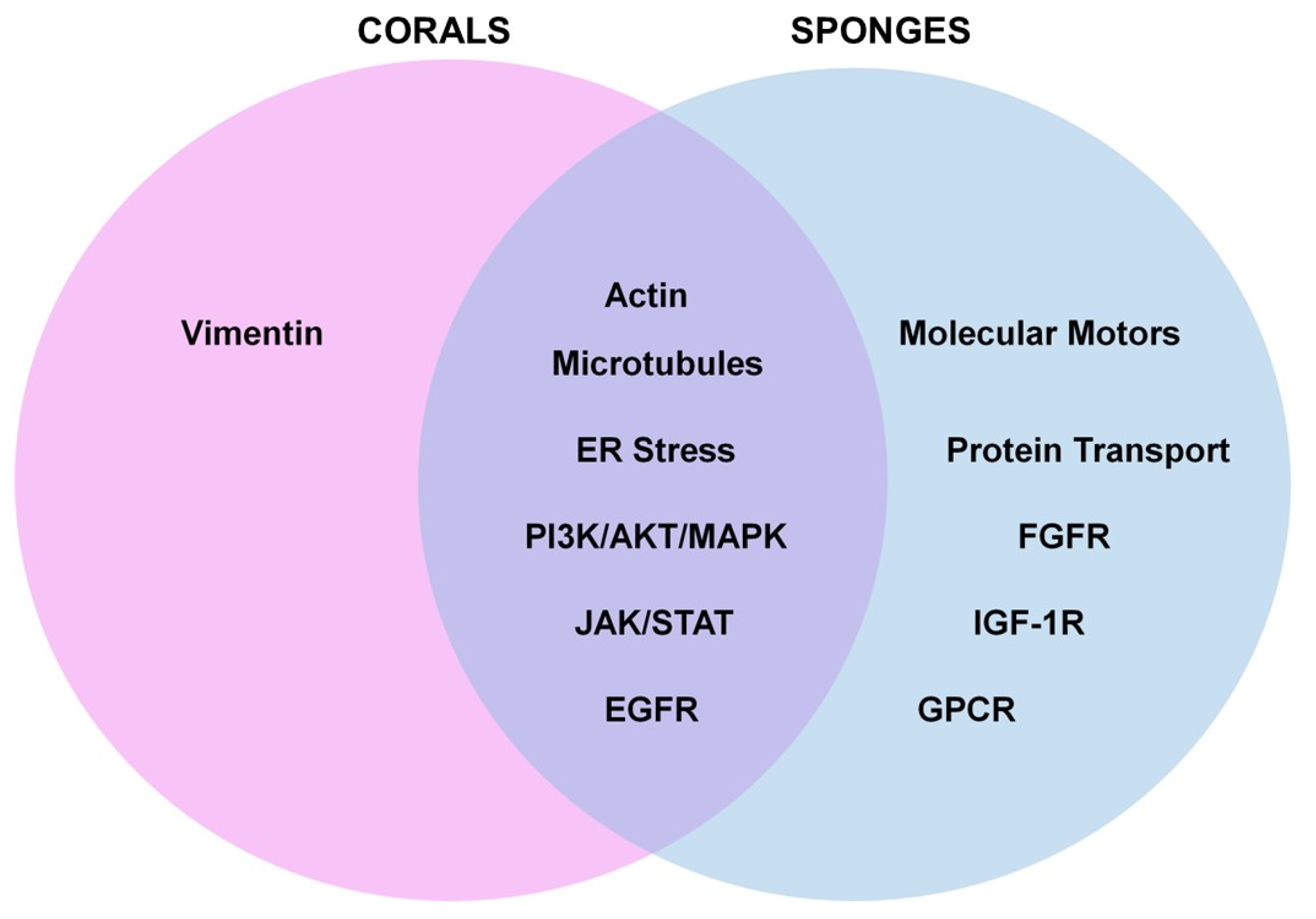
4.3. Gaps in Knowledge, Limitations, and Future Perspectives
5. Supporting the Drug Discovery Pipeline
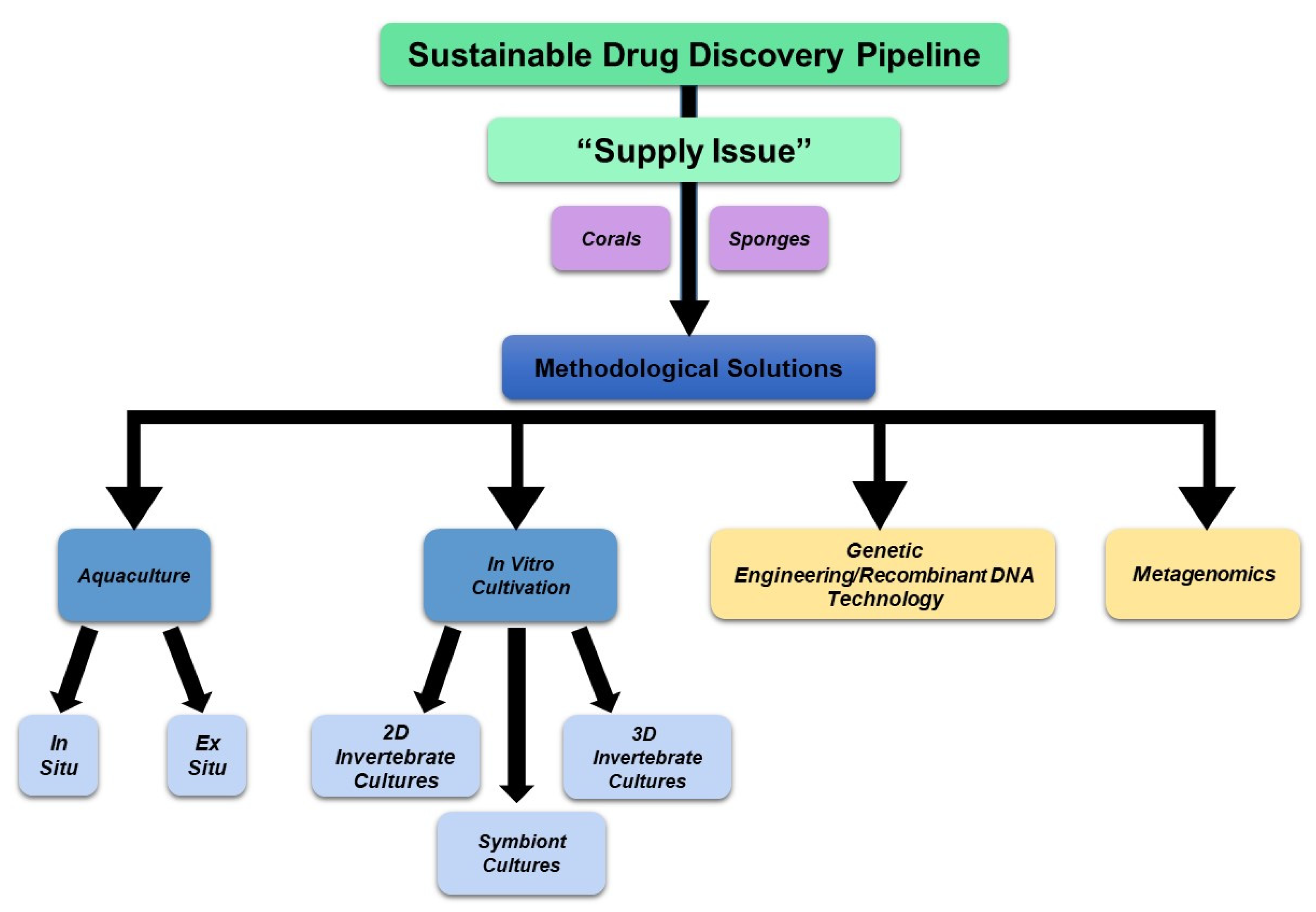
5.1. Overview—“Supply Issue” and Methodological Solutions
5.2. Aquaculture
5.3. In Vitro Cultivation
5.4. Future Considerations
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nguyen, N.B.A.; Chen, L.Y.; El-Shazly, M.; Peng, B.R.; Su, J.H.; Wu, H.C.; Lee, I.T.; Lai, K.H. Towards Sustainable Medicinal Resources through Marine Soft Coral Aquaculture: Insights into the Chemical Diversity and the Biological Potential. Mar. Drugs 2022, 20, 640. [Google Scholar] [CrossRef] [PubMed]
- Varijakzhan, D.; Loh, J.Y.; Yap, W.S.; Yusoff, K.; Seboussi, R.; Lim, S.E.; Lai, K.S.; Chong, C.M. Bioactive Compounds from Marine Sponges: Fundamentals and Applications. Mar. Drugs 2021, 19, 246. [Google Scholar] [CrossRef]
- Leal, M.C.; Calado, R.; Sheridan, C.; Alimonti, A.; Osinga, R. Coral aquaculture to support drug discovery. Trends Biotechnol. 2013, 31, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, C.; Valderrama, K.; Zea, S.; Castellanos, L. Mariculture and natural production of the antitumoural (+)-discodermolide by the Caribbean marine sponge Discodermia dissoluta. Mar. Biotechnol. 2013, 15, 571–583. [Google Scholar] [CrossRef] [PubMed]
- Roger, L.M.; Reich, H.G.; Lawrence, E.; Li, S.; Vizgaudis, W.; Brenner, N.; Kumar, L.; Klein-Seetharaman, J.; Yang, J.; Putnam, H.M.; et al. Applying model approaches in non-model systems: A review and case study on coral cell culture. PLoS ONE 2021, 16, e0248953. [Google Scholar] [CrossRef] [PubMed]
- Liebezeit, G. Aquaculture of “non-food organisms” for natural substance production. Adv. Biochem. Eng. Biotechnol. 2005, 97, 1–28. [Google Scholar] [PubMed]
- Leal, M.C.; Sheridan, C.; Osinga, R.; Dionisio, G.; Rocha, R.J.; Silva, B.; Rosa, R.; Calado, R. Marine microorganism-invertebrate assemblages: Perspectives to solve the “supply problem” in the initial steps of drug discovery. Mar. Drugs 2014, 12, 3929–3952. [Google Scholar] [CrossRef] [PubMed]
- Deep Sea Coral Research and Technology Program (DSCRTP). Observations of Deep-Sea Coral and Sponge Occurrences from the NOAA National Deep-Sea Coral and Sponge Database, 1842-Present, version 20230620-0 (NCEI Accession 0145037). [Western Coast of Florida]. NOAA National Centers for Environmental Information. Dataset. 2016. Available online: https://www.ncei.noaa.gov/archive/accession/0145037 (accessed on 26 May 2023 to 8 June 2023).
- Ahyong, S.; Boyko, C.B.; Bailly, N.; Bernot, J.; Bieler, R.; Brandão, S.N.; Daly, M.; De Grave, S.; Gofas, S.; Hernandez, F.; et al. World Register of Marine Species. 2023. Available online: https://www.marinespecies.org (accessed on 22 November 2023).
- de Voogd, N.J.; Alvarez, B.; Boury-Esnault, N.; Cárdenas, P.; Díaz, M.-C.; Dohrmann, M.; Downey, R.; Goodwin, C.; Hajdu, E.; Hooper, J.N.A.; et al. World Porifera Database. 2023. Available online: https://www.marinespecies.org/porifera (accessed on 22 November 2023).
- Lopp, A.; Pihlak, A.; Paves, H.; Samuel, K.; Koljak, R.; Samel, N. The effect of 9,11-secosterol, a newly discovered compound from the soft coral Gersemia fruticosa, on the growth and cell cycle progression of various tumor cells in culture. Steroids 1994, 59, 274–281. [Google Scholar] [CrossRef]
- Long, B.H.; Carboni, J.M.; Wasserman, A.J.; Cornell, L.A.; Casazza, A.M.; Jensen, P.R.; Lindel, T.; Fenical, W.; Fairchild, C.R. Eleutherobin, a novel cytotoxic agent that induces tubulin polymerization, is similar to paclitaxel (Taxol). Cancer Res. 1998, 58, 1111–1115. [Google Scholar] [PubMed]
- Morretta, E.; Esposito, R.; Festa, C.; Riccio, R.; Casapullo, A.; Monti, M.C. Discovering the Biological Target of 5-epi-Sinuleptolide Using a Combination of Proteomic Approaches. Mar. Drugs 2017, 15, 312. [Google Scholar] [CrossRef]
- Ko, C.Y.; Shih, P.C.; Huang, P.W.; Lee, Y.H.; Chen, Y.F.; Tai, M.H.; Liu, C.H.; Wen, Z.H.; Kuo, H.M. Sinularin, an Anti-Cancer Agent Causing Mitochondria-Modulated Apoptosis and Cytoskeleton Disruption in Human Hepatocellular Carcinoma. Int. J. Mol. Sci. 2021, 22, 3946. [Google Scholar] [CrossRef]
- Wu, Z.; Su, M.; Chen, H.; Chen, X.; Chen, C.Y.; An, L.; Shao, Z.; Liu, X.; Lin, Y.; OuYang, A.J.; et al. Sinularin Exerts Anti-Cancer Effects by Inducing Oxidative Stress-Mediated Ferroptosis, Apoptosis, and Autophagy in Prostate Cancer Cells. Anticancer Agents Med. Chem. 2023, 23, 1457–1468. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.I.; Wang, R.Y.; Lin, J.J.; Su, J.H.; Chiu, C.C.; Chen, J.C.; Chen, J.Y.; Wu, Y.J. Proteomic profiling of the 11-dehydrosinulariolide-treated oral carcinoma cells Ca9–22: Effects on the cell apoptosis through mitochondrial-related and ER stress pathway. J. Proteom. 2012, 75, 5578–5589. [Google Scholar] [CrossRef]
- Su, T.R.; Tsai, F.J.; Lin, J.J.; Huang, H.H.; Chiu, C.C.; Su, J.H.; Yang, Y.T.; Chen, J.Y.; Wong, B.S.; Wu, Y.J. Induction of apoptosis by 11-dehydrosinulariolide via mitochondrial dysregulation and ER stress pathways in human melanoma cells. Mar. Drugs 2012, 10, 1883–1898. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Su, J.H.; Tsao, C.Y.; Hung, C.T.; Chao, H.H.; Lin, J.J.; Liao, M.H.; Yang, Z.Y.; Huang, H.H.; Tsai, F.J.; et al. Sinulariolide induced hepatocellular carcinoma apoptosis through activation of mitochondrial-related apoptotic and PERK/eIF2α/ATF4/CHOP pathway. Molecules 2013, 18, 10146–10161. [Google Scholar] [CrossRef] [PubMed]
- Tsai, T.C.; Lai, K.H.; Su, J.H.; Wu, Y.J.; Sheu, J.H. 7-Acetylsinumaximol B Induces Apoptosis and Autophagy in Human Gastric Carcinoma Cells through Mitochondria Dysfunction and Activation of the PERK/eIF2α/ATF4/CHOP Signaling Pathway. Mar. Drugs 2018, 16, 104. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.J.; Wang, R.Y.; Chen, J.C.; Chiu, C.C.; Liao, M.H.; Wu, Y.J. Cytotoxicity of 11-epi-Sinulariolide Acetate Isolated from Cultured Soft Corals on HA22T Cells through the Endoplasmic Reticulum Stress Pathway and Mitochondrial Dysfunction. Int. J. Mol. Sci. 2016, 17, 1787. [Google Scholar] [CrossRef] [PubMed]
- She, Y.Y.; Lin, J.J.; Su, J.H.; Chang, T.S.; Wu, Y.J. 4-Carbomethoxyl-10-epigyrosanoldie E Extracted from Cultured Soft Coral Sinularia sandensis Induced Apoptosis and Autophagy via ROS and Mitochondrial Dysfunction and ER Stress in Oral Cancer Cells. Oxid. Med. Cell. Longev. 2022, 2022, 3017807. [Google Scholar] [CrossRef] [PubMed]
- Chiang, P.C.; Chien, C.L.; Pan, S.L.; Chen, W.P.; Teng, C.M.; Shen, Y.C.; Guh, J.H. Induction of endoplasmic reticulum stress and apoptosis by a marine prostanoid in human hepatocellular carcinoma. J. Hepatol. 2005, 43, 679–686. [Google Scholar] [CrossRef]
- Lai, K.M.; Wang, J.H.; Lin, S.C.; Wen, Y.; Wu, C.L.; Su, J.H.; Chen, C.C.; Lin, C.C. Crassolide Induces G2/M Cell Cycle Arrest, Apoptosis, and Autophagy in Human Lung Cancer Cells via ROS-Mediated ER Stress Pathways. Int. J. Mol. Sci. 2022, 23, 5624. [Google Scholar] [CrossRef]
- Chen, W.F.; Chakraborty, C.; Sung, C.S.; Feng, C.W.; Jean, Y.H.; Lin, Y.Y.; Hung, H.C.; Huang, T.Y.; Huang, S.Y.; Su, T.M.; et al. Neuroprotection by marine-derived compound, 11-dehydrosinulariolide, in an in vitro Parkinson’s model: A promising candidate for the treatment of Parkinson’s disease. Naunyn Schmiedebergs Arch. Pharmacol. 2012, 385, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.C.; Su, J.H.; Lin, S.C.; Chang, C.C.; Hsia, T.C.; Tung, Y.T.; Lin, C.C. A Soft Coral-Derived Compound, 11-Dehydrosinulariolide, Induces G2/M Cell Cycle Arrest and Apoptosis in Small Cell Lung Cancer. Mar. Drugs 2018, 16, 479. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.J.; Su, J.H.; Tsai, C.C.; Chen, Y.J.; Liao, M.H.; Wu, Y.J. 11-epi-Sinulariolide acetate reduces cell migration and invasion of human hepatocellular carcinoma by reducing the activation of ERK1/2, p38MAPK and FAK/PI3K/AKT/mTOR signaling pathways. Mar. Drugs 2014, 12, 4783–4798. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.J.; Neoh, C.A.; Tsao, C.Y.; Su, J.H.; Li, H.H. Sinulariolide Suppresses Human Hepatocellular Carcinoma Cell Migration and Invasion by Inhibiting Matrix Metalloproteinase-2/-9 through MAPKs and PI3K/Akt Signaling Pathways. Int. J. Mol. Sci. 2015, 16, 16469–16482. [Google Scholar] [CrossRef]
- Cheng, T.C.; Din, Z.H.; Su, J.H.; Wu, Y.J.; Liu, C.I. Sinulariolide Suppresses Cell Migration and Invasion by Inhibiting Matrix Metalloproteinase-2/-9 and Urokinase through the PI3K/AKT/mTOR Signaling Pathway in Human Bladder Cancer Cells. Mar. Drugs 2017, 15, 238. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.J.; Lin, S.H.; Din, Z.H.; Su, J.H.; Liu, C.I. Sinulariolide Inhibits Gastric Cancer Cell Migration and Invasion through Downregulation of the EMT Process and Suppression of FAK/PI3K/AKT/mTOR and MAPKs Signaling Pathways. Mar. Drugs 2019, 17, 668. [Google Scholar] [CrossRef]
- Ma, Q.; Meng, X.Y.; Wu, K.R.; Cao, J.Z.; Yu, R.; Yan, Z.J. Sinularin exerts anti-tumor effects against human renal cancer cells relies on the generation of ROS. J. Cancer 2019, 10, 5114–5123. [Google Scholar] [CrossRef]
- Wu, Y.J.; Wong, B.S.; Yea, S.H.; Lu, C.I.; Weng, S.H. Sinularin Induces Apoptosis through Mitochondria Dysfunction and Inactivation of the pI3K/Akt/mTOR Pathway in Gastric Carcinoma Cells. Mar. Drugs 2016, 14, 142. [Google Scholar] [CrossRef]
- Wu, Y.J.; Su, T.R.; Dai, G.F.; Su, J.H.; Liu, C.I. Flaccidoxide-13-Acetate-Induced Apoptosis in Human Bladder Cancer Cells is through Activation of p38/JNK, Mitochondrial Dysfunction, and Endoplasmic Reticulum Stress Regulated Pathway. Mar. Drugs 2019, 17, 287. [Google Scholar] [CrossRef]
- Wu, Y.J.; Wei, W.C.; Dai, G.F.; Su, J.H.; Tseng, Y.H.; Tsai, T.C. Exploring the Mechanism of Flaccidoxide-13-Acetate in Suppressing Cell Metastasis of Hepatocellular Carcinoma. Mar. Drugs 2020, 18, 314. [Google Scholar] [CrossRef]
- Neoh, C.A.; Wu, W.T.; Dai, G.F.; Su, J.H.; Liu, C.I.; Su, T.R.; Wu, Y.J. Flaccidoxide-13-Acetate Extracted from the Soft Coral Cladiella kashmani Reduces Human Bladder Cancer Cell Migration and Invasion through Reducing Activation of the FAK/PI3K/AKT/mTOR Signaling Pathway. Molecules 2017, 23, 58. [Google Scholar] [CrossRef] [PubMed]
- Su, C.C.; Chen, J.Y.; Din, Z.H.; Su, J.H.; Yang, Z.Y.; Chen, Y.J.; Wang, R.Y.; Wu, Y.J. 13-acetoxysarcocrassolide induces apoptosis on human gastric carcinoma cells through mitochondria-related apoptotic pathways: p38/JNK activation and PI3K/AKT suppression. Mar. Drugs 2014, 12, 5295–5315. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.M.; Lin, J.J.; Su, J.H.; Liu, C.I. 13-Acetoxysarcocrassolide induces apoptosis in human hepatocellular carcinoma cells through mitochondrial dysfunction and suppression of the PI3K/AKT/mTOR/p70S6K signalling pathway. Pharm. Biol. 2022, 60, 2276–2285. [Google Scholar] [CrossRef] [PubMed]
- Lan, Q.; Li, S.; Lai, W.; Xu, H.; Zhang, Y.; Zeng, Y.; Lan, W.; Chu, Z. Methyl Sartortuoate Inhibits Colon Cancer Cell Growth by Inducing Apoptosis and G2/M-Phase Arrest. Int. J. Mol. Sci. 2015, 16, 19401–19418. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.A.A.; Abdelrahman, A.H.M.; Atia, M.A.M.; Mohamed, T.A.; Moustafa, M.F.; Hakami, A.R.; Khalifa, S.A.M.; Alhumaydhi, F.A.; Alrumaihi, F.; Abidi, S.H.; et al. Blue Biotechnology: Computational Screening of Sarcophyton Cembranoid Diterpenes for SARS-CoV-2 Main Protease Inhibition. Mar. Drugs 2021, 19, 391. [Google Scholar] [CrossRef] [PubMed]
- Chung, T.W.; Su, J.H.; Lin, C.C.; Li, Y.R.; Chao, Y.H.; Lin, S.H.; Chan, H.L. 24-Methyl-Cholesta-5,24(28)-Diene-3beta,19-diol-7beta-Monoacetate Inhibits Human Small Cell Lung Cancer Growth In Vitro and In Vivo via Apoptosis Induction. Mar. Drugs 2017, 15, 210. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.K.; Tseng, C.K.; Liaw, C.C.; Huang, C.Y.; Wei, C.K.; Sheu, J.H.; Lee, J.C. Lobohedleolide suppresses hepatitis C virus replication via JNK/c-Jun-C/EBP-mediated down-regulation of cyclooxygenase-2 expression. Sci. Rep. 2018, 8, 8676. [Google Scholar] [CrossRef]
- Meng, J.; Zhang, X.; Guo, X.; Cheng, W.; Qi, X.; Huang, J.; Lin, W. Briarane-type diterpenoids suppress osteoclastogenisis by regulation of Nrf2 and MAPK/NF-kB signaling pathway. Bioorg. Chem. 2021, 112, 104976. [Google Scholar] [CrossRef]
- Chen, Y.C.; Wen, Z.H.; Lee, Y.H.; Chen, C.L.; Hung, H.C.; Chen, C.H.; Chen, W.F.; Tsai, M.C. Dihydroaustrasulfone alcohol inhibits PDGF-induced proliferation and migration of human aortic smooth muscle cells through inhibition of the cell cycle. Mar. Drugs 2015, 13, 2390–2406. [Google Scholar] [CrossRef]
- Kao, C.J.; Chen, W.F.; Guo, B.L.; Feng, C.W.; Hung, H.C.; Yang, W.Y.; Sung, C.S.; Tsui, K.H.; Chu, H.; Chen, N.F.; et al. The 1-Tosylpentan-3-one Protects against 6-Hydroxydopamine-Induced Neurotoxicity. Int. J. Mol. Sci. 2017, 18, 1096. [Google Scholar] [CrossRef]
- Tsai, M.K.; Tsai, M.L.; Wen, Z.H.; Liao, W.T.; Lin, Y.C.; Chiou, H.C.; Lin, M.H.; Hung, C.H. Suppressive Effects of 4-(Phenylsulfanyl) Butan-2-One on CCL-1 Production via Histone Acetylation in Monocytes. Curr. Issues Mol. Biol. 2022, 44, 4616–4625. [Google Scholar] [CrossRef] [PubMed]
- Alhadrami, H.A.; Alkhatabi, H.; Abduljabbar, F.H.; Abdelmohsen, U.R.; Sayed, A.M. Anticancer Potential of Green Synthesized Silver Nanoparticles of the Soft Coral Cladiella pachyclados Supported by Network Pharmacology and In Silico Analyses. Pharmaceutics 2021, 13, 1846. [Google Scholar] [CrossRef] [PubMed]
- Kuo, L.M.; Chen, P.J.; Sung, P.J.; Chang, Y.C.; Ho, C.T.; Wu, Y.H.; Hwang, T.L. The Bioactive Extract of Pinnigorgia sp. Induces Apoptosis of Hepatic Stellate Cells via ROS-ERK/JNK-Caspase-3 Signaling. Mar. Drugs 2018, 16, 19. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.Y.; Boo, H.J.; Kang, J.I.; Kim, M.K.; Yoo, E.S.; Hyun, J.W.; Koh, Y.S.; Kim, G.Y.; Maeng, Y.H.; Hyun, C.L.; et al. (1S,2S,3E,7E,11E)-3,7,11,15-Cembratetraen-17,2-olide, a cembrenolide diterpene from soft coral Lobophytum sp., inhibits growth and induces apoptosis in human colon cancer cells through reactive oxygen species generation. Biol. Pharm. Bull. 2012, 35, 1054–1063. [Google Scholar] [CrossRef]
- Tsai, W.C.; Wang, W.H.; Huang, B.C.; Huang, C.Y.; Sheu, J.H. 5-epi-Sinuleptolide from Soft Corals of the Genus Sinularia Exerts Cytotoxic Effects on Pancreatic Cancer Cell Lines via the Inhibition of JAK2/STAT3, AKT, and ERK Activity. Molecules 2021, 26, 6932. [Google Scholar] [CrossRef] [PubMed]
- Mohyeldin, M.M.; Akl, M.R.; Siddique, A.B.; Hassan, H.M.; El Sayed, K.A. The marine-derived pachycladin diterpenoids as novel inhibitors of wild-type and mutant EGFR. Biochem. Pharmacol. 2017, 126, 51–68. [Google Scholar] [CrossRef]
- Hegazy, M.F.; Elshamy, A.I.; Mohamed, T.A.; Hamed, A.R.; Ibrahim, M.A.A.; Ohta, S.; Pare, P.W. Cembrene Diterpenoids with Ether Linkages from Sarcophyton ehrenbergi: An Anti-Proliferation and Molecular-Docking Assessment. Mar. Drugs 2017, 15, 192. [Google Scholar] [CrossRef]
- Martin, M.J.; Coello, L.; Fernandez, R.; Reyes, F.; Rodriguez, A.; Murcia, C.; Garranzo, M.; Mateo, C.; Sanchez-Sancho, F.; Bueno, S.; et al. Isolation and first total synthesis of PM050489 and PM060184, two new marine anticancer compounds. J. Am. Chem. Soc. 2013, 135, 10164–10171. [Google Scholar] [CrossRef]
- Pera, B.; Barasoain, I.; Pantazopoulou, A.; Canales, A.; Matesanz, R.; Rodriguez-Salarichs, J.; Garcia-Fernandez, L.F.; Moneo, V.; Jimenez-Barbero, J.; Galmarini, C.M.; et al. New interfacial microtubule inhibitors of marine origin, PM050489/PM060184, with potent antitumor activity and a distinct mechanism. ACS Chem. Biol. 2013, 8, 2084–2094. [Google Scholar] [CrossRef]
- Bai, R.L.; Paull, K.D.; Herald, C.L.; Malspeis, L.; Pettit, G.R.; Hamel, E. Halichondrin B and homohalichondrin B, marine natural products binding in the vinca domain of tubulin. Discovery of tubulin-based mechanism of action by analysis of differential cytotoxicity data. J. Biol. Chem. 1991, 266, 15882–15889. [Google Scholar] [CrossRef]
- Margarucci, L.; Monti, M.C.; Esposito, R.; Tosco, A.; Hamel, E.; Riccio, R.; Casapullo, A. N-Formyl-7-amino-11-cycloamphilectene, a marine sponge metabolite, binds to tubulin and modulates microtubule depolymerization. Mol. Biosyst. 2014, 10, 862–867. [Google Scholar] [CrossRef] [PubMed][Green Version]
- ter Haar, E.; Kowalski, R.J.; Hamel, E.; Lin, C.M.; Longley, R.E.; Gunasekera, S.P.; Rosenkranz, H.S.; Day, B.W. Discodermolide, a cytotoxic marine agent that stabilizes microtubules more potently than taxol. Biochemistry 1996, 35, 243–250. [Google Scholar] [CrossRef]
- Isbrucker, R.A.; Cummins, J.; Pomponi, S.A.; Longley, R.E.; Wright, A.E. Tubulin polymerizing activity of dictyostatin-1, a polyketide of marine sponge origin. Biochem. Pharmacol. 2003, 66, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Sonnenschein, R.N.; Farias, J.J.; Tenney, K.; Mooberry, S.L.; Lobkovsky, E.; Clardy, J.; Crews, P. A further study of the cytotoxic constituents of a milnamide-producing sponge. Org. Lett. 2004, 6, 779–782. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.W.; Singh, A.J.; Patel, V.; Russell, E.R.; Field, J.J.; Miller, J.H.; Northcote, P.T. Peloruside E (22-Norpeloruside A), a Pelorusane Macrolide from the New Zealand Marine Sponge Mycale hentscheli, Retains Microtubule-Stabilizing Properties. J. Nat. Prod. 2018, 81, 2125–2128. [Google Scholar] [CrossRef] [PubMed]
- Uckun, F.M.; Mao, C.; Vassilev, A.O.; Huang, H.; Jan, S.T. Structure-based design of a novel synthetic spiroketal pyran as a pharmacophore for the marine natural product spongistatin 1. Bioorg. Med. Chem. Lett. 2000, 10, 541–545. [Google Scholar] [CrossRef] [PubMed]
- Pettit, R.K.; Woyke, T.; Pon, S.; Cichacz, Z.A.; Pettit, G.R.; Herald, C.L. In vitro and in vivo antifungal activities of the marine sponge constituent spongistatin. Med. Mycol. 2005, 43, 453–463. [Google Scholar] [CrossRef]
- Field, J.J.; Singh, A.J.; Kanakkanthara, A.; Halafihi, T.; Northcote, P.T.; Miller, J.H. Microtubule-stabilizing activity of zampanolide, a potent macrolide isolated from the Tongan marine sponge Cacospongia mycofijiensis. J. Med. Chem. 2009, 52, 7328–7332. [Google Scholar] [CrossRef]
- Bae, S.Y.; Kim, G.D.; Jeon, J.E.; Shin, J.; Lee, S.K. Anti-proliferative effect of (19Z)-halichondramide, a novel marine macrolide isolated from the sponge Chondrosia corticata, is associated with G2/M cell cycle arrest and suppression of mTOR signaling in human lung cancer cells. Toxicol. In Vitro 2013, 27, 694–699. [Google Scholar] [CrossRef]
- Rangel, M.; Prado, M.P.; Konno, K.; Naoki, H.; Freitas, J.C.; Machado-Santelli, G.M. Cytoskeleton alterations induced by Geodia corticostylifera depsipeptides in breast cancer cells. Peptides 2006, 27, 2047–2057. [Google Scholar] [CrossRef]
- Freitas, V.M.; Rangel, M.; Bisson, L.F.; Jaeger, R.G.; Machado-Santelli, G.M. The geodiamolide H, derived from Brazilian sponge Geodia corticostylifera, regulates actin cytoskeleton, migration and invasion of breast cancer cells cultured in three-dimensional environment. J. Cell. Physiol. 2008, 216, 583–594. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Minale, L.; Zampella, A.; Smith, C.D. Microfilament depletion and circumvention of multiple drug resistance by sphinxolides. Cancer Res. 1997, 57, 3751–3758. [Google Scholar] [PubMed]
- Youssef, D.T.; Mooberry, S.L. Hurghadolide A and swinholide I, potent actin-microfilament disrupters from the Red Sea sponge Theonella swinhoei. J. Nat. Prod. 2006, 69, 154–157. [Google Scholar] [CrossRef] [PubMed]
- Roskelley, C.D.; Williams, D.E.; McHardy, L.M.; Leong, K.G.; Troussard, A.; Karsan, A.; Andersen, R.J.; Dedhar, S.; Roberge, M. Inhibition of tumor cell invasion and angiogenesis by motuporamines. Cancer Res. 2001, 61, 6788–6794. [Google Scholar] [PubMed]
- Isbrucker, R.A.; Guzman, E.A.; Pitts, T.P.; Wright, A.E. Early effects of lasonolide a on pancreatic cancer cells. J. Pharmacol. Exp. Ther. 2009, 331, 733–739. [Google Scholar] [CrossRef]
- Hayashi-Takanaka, Y.; Kina, Y.; Nakamura, F.; Yamazaki, S.; Harata, M.; Soest, R.; Kimura, H.; Nakao, Y. Effect of mycalolides isolated from a marine sponge Mycale aff. nullarosette on actin in living cells. Sci. Rep. 2019, 9, 7540. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.A.; Odde, S.; Daga, P.R.; Bowling, J.J.; Mesbah, M.K.; Youssef, D.T.; Khalifa, S.I.; Doerksen, R.J.; Hamann, M.T. Latrunculin with a highly oxidized thiazolidinone ring: Structure assignment and actin docking. Org. Lett. 2007, 9, 4773–4776. [Google Scholar] [CrossRef] [PubMed]
- Helal, M.A.; Khalifa, S.; Ahmed, S. Differential binding of latrunculins to G-actin: A molecular dynamics study. J. Chem. Inf. Model. 2013, 53, 2369–2375. [Google Scholar] [CrossRef]
- Morton, W.M.; Ayscough, K.R.; McLaughlin, P.J. Latrunculin alters the actin-monomer subunit interface to prevent polymerization. Nat. Cell Biol. 2000, 2, 376–378. [Google Scholar] [CrossRef]
- Sayed, K.A.; Khanfar, M.A.; Shallal, H.M.; Muralidharan, A.; Awate, B.; Youssef, D.T.; Liu, Y.; Zhou, Y.D.; Nagle, D.G.; Shah, G. Latrunculin A and its C-17-O-carbamates inhibit prostate tumor cell invasion and HIF-1 activation in breast tumor cells. J. Nat. Prod. 2008, 71, 396–402. [Google Scholar] [CrossRef]
- Khanfar, M.A.; Youssef, D.T.; El Sayed, K.A. Semisynthetic latrunculin derivatives as inhibitors of metastatic breast cancer: Biological evaluations, preliminary structure-activity relationship and molecular modeling studies. ChemMedChem 2010, 5, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Petchprayoon, C.; Suwanborirux, K.; Miller, R.; Sakata, T.; Marriott, G. Synthesis and characterization of the 7-(4-aminomethyl-1H-1,2,3-triazol-1-yl) analogue of kabiramide C. J. Nat. Prod. 2005, 68, 157–161. [Google Scholar] [CrossRef]
- Cipres, A.; O’Malley, D.P.; Li, K.; Finlay, D.; Baran, P.S.; Vuori, K. Sceptrin, a marine natural compound, inhibits cell motility in a variety of cancer cell lines. ACS Chem. Biol. 2010, 5, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.C.; Lu, M.C.; El-Shazly, M.; Lai, K.H.; Wu, T.Y.; Hsu, Y.M.; Lee, Y.L.; Liu, Y.C. Breaking down Leukemia Walls: Heteronemin, a Sesterterpene Derivative, Induces Apoptosis in Leukemia Molt4 Cells through Oxidative Stress, Mitochondrial Dysfunction and Induction of Talin Expression. Mar. Drugs 2018, 16, 212. [Google Scholar] [CrossRef] [PubMed]
- Sakowicz, R.; Berdelis, M.S.; Ray, K.; Blackburn, C.L.; Hopmann, C.; Faulkner, D.J.; Goldstein, L.S. A marine natural product inhibitor of kinesin motors. Science 1998, 280, 292–295. [Google Scholar] [CrossRef] [PubMed]
- Brier, S.; Carletti, E.; DeBonis, S.; Hewat, E.; Lemaire, D.; Kozielski, F. The marine natural product adociasulfate-2 as a tool to identify the MT-binding region of kinesins. Biochemistry 2006, 45, 15644–15653. [Google Scholar] [CrossRef]
- Smith, T.E.; Hong, W.; Zachariah, M.M.; Harper, M.K.; Matainaho, T.K.; Van Wagoner, R.M.; Ireland, C.M.; Vershinin, M. Single-molecule inhibition of human kinesin by adociasulfate-13 and -14 from the sponge Cladocroce aculeata. Proc. Natl. Acad. Sci. USA 2013, 110, 18880–18885. [Google Scholar] [CrossRef]
- Zhu, G.; Yang, F.; Balachandran, R.; Hook, P.; Vallee, R.B.; Curran, D.P.; Day, B.W. Synthesis and biological evaluation of purealin and analogues as cytoplasmic dynein heavy chain inhibitors. J. Med. Chem. 2006, 49, 2063–2076. [Google Scholar] [CrossRef]
- Cheung, F.W.K.; Guo, J.; Ling, Y.H.; Che, C.T.; Liu, W.K. Anti-melanogenic property of geoditin A in murine B16 melanoma cells. Mar. Drugs 2012, 10, 465–476. [Google Scholar] [CrossRef]
- Liu, W.K.; Ling, Y.H.; Cheung, F.W.; Che, C.T. Stellettin A induces endoplasmic reticulum stress in murine B16 melanoma cells. J. Nat. Prod. 2012, 75, 586–590. [Google Scholar] [CrossRef]
- Kim, Y.N.; Ji, Y.K.; Kim, N.H.; Van Tu, N.; Rho, J.R.; Jeong, E.J. Isoquinolinequinone Derivatives from a Marine Sponge (Haliclona sp.) Regulate Inflammation in In Vitro System of Intestine. Mar. Drugs 2021, 19, 90. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, F.C.; Kau, T.R.; Silver, P.A.; Clardy, J. The psammaplysenes, specific inhibitors of FOXO1a nuclear export. J. Nat. Prod. 2005, 68, 574–576. [Google Scholar] [CrossRef] [PubMed]
- Takizawa, P.A.; Yucel, J.K.; Veit, B.; Faulkner, D.J.; Deerinck, T.; Soto, G.; Ellisman, M.; Malhotra, V. Complete vesiculation of Golgi membranes and inhibition of protein transport by a novel sea sponge metabolite, ilimaquinone. Cell 1993, 73, 1079–1090. [Google Scholar] [CrossRef]
- Lu, P.H.; Chueh, S.C.; Kung, F.L.; Pan, S.L.; Shen, Y.C.; Guh, J.H. Ilimaquinone, a marine sponge metabolite, displays anticancer activity via GADD153-mediated pathway. Eur. J. Pharmacol. 2007, 556, 45–54. [Google Scholar] [CrossRef]
- De Stefano, D.; Tommonaro, G.; Malik, S.A.; Iodice, C.; De Rosa, S.; Maiuri, M.C.; Carnuccio, R. Cacospongionolide and scalaradial, two marine sesterterpenoids as potent apoptosis-inducing factors in human carcinoma cell lines. PLoS ONE 2012, 7, e33031. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.; Son, A.; Lee, H.S.; Lee, Y.J.; Park, H.C. Radiosensitization by Marine Sponge Agelas sp. Extracts in Hepatocellular Carcinoma Cells with Autophagy Induction. Sci. Rep. 2018, 8, 6317. [Google Scholar] [CrossRef]
- Lu, I.T.; Lin, S.C.; Chu, Y.C.; Wen, Y.; Lin, Y.C.; Cheng, W.C.; Sheu, J.H.; Lin, C.C. (−)-Agelasidine A Induces Endoplasmic Reticulum Stress-Dependent Apoptosis in Human Hepatocellular Carcinoma. Mar. Drugs 2022, 20, 109. [Google Scholar] [CrossRef]
- Choi, C.; Cho, Y.; Son, A.; Shin, S.W.; Lee, Y.J.; Park, H.C. Therapeutic Potential of (−)-Agelamide D, a Diterpene Alkaloid from the Marine Sponge Agelas sp., as a Natural Radiosensitizer in Hepatocellular Carcinoma Models. Mar. Drugs 2020, 18, 500. [Google Scholar] [CrossRef]
- Florean, C.; Kim, K.R.; Schnekenburger, M.; Kim, H.J.; Moriou, C.; Debitus, C.; Dicato, M.; Al-Mourabit, A.; Han, B.W.; Diederich, M. Synergistic AML Cell Death Induction by Marine Cytotoxin (+)-1(R), 6(S), 1’(R), 6’(S), 11(R), 17(S)-Fistularin-3 and Bcl-2 Inhibitor Venetoclax. Mar. Drugs 2018, 16, 518. [Google Scholar] [CrossRef]
- Florean, C.; Schnekenburger, M.; Lee, J.Y.; Kim, K.R.; Mazumder, A.; Song, S.; Kim, J.M.; Grandjenette, C.; Kim, J.G.; Yoon, A.Y.; et al. Discovery and characterization of Isofistularin-3, a marine brominated alkaloid, as a new DNA demethylating agent inducing cell cycle arrest and sensitization to TRAIL in cancer cells. Oncotarget 2016, 7, 24027–24049. [Google Scholar] [CrossRef]
- Lai, K.H.; Liu, Y.C.; Su, J.H.; El-Shazly, M.; Wu, C.F.; Du, Y.C.; Hsu, Y.M.; Yang, J.C.; Weng, M.K.; Chou, C.H.; et al. Antileukemic Scalarane Sesterterpenoids and Meroditerpenoid from Carteriospongia (Phyllospongia) sp., Induce Apoptosis via Dual Inhibitory Effects on Topoisomerase II and Hsp90. Sci. Rep. 2016, 6, 36170. [Google Scholar] [CrossRef] [PubMed]
- Namba, T.; Kodama, R. Avarol induces apoptosis in pancreatic ductal adenocarcinoma cells by activating PERK-eIF2α-CHOP signaling. Mar. Drugs 2015, 13, 2376–2389. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.D.; Cheong, O.J.; Bae, S.Y.; Shin, J.; Lee, S.K. 6″-Debromohamacanthin A, a bis (indole) alkaloid, inhibits angiogenesis by targeting the VEGFR2-mediated PI3K/AKT/mTOR signaling pathways. Mar. Drugs 2013, 11, 1087–1103. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, H.; Kanno, S.I.; Abdjul, D.B.; Namikoshi, M. A bromopyrrole-containing diterpene alkaloid from the Okinawan marine sponge Agelas nakamurai activates the insulin pathway in Huh-7 human hepatoma cells by inhibiting protein tyrosine phosphatase 1B. Bioorg. Med. Chem. Lett. 2017, 27, 2207–2209. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Zhang, Q.; Peng, X.; Zhou, C.; Zhong, Y.; Chen, X.; Qiu, Y.; Jin, M.; Gong, M.; Kong, D. Stellettin B Induces G1 Arrest, Apoptosis and Autophagy in Human Non-small Cell Lung Cancer A549 Cells via Blocking PI3K/Akt/mTOR Pathway. Sci. Rep. 2016, 6, 27071. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.A.; Zhou, Q.; Guo, W.Z.; Qiu, Y.; Wang, R.; Jin, M.; Zhang, W.; Li, K.; Yamori, T.; Dan, S.; et al. In vitro antitumor activity of stellettin B, a triterpene from marine sponge Jaspis stellifera, on human glioblastoma cancer SF295 cells. Mar. Drugs 2014, 12, 4200–4213. [Google Scholar] [CrossRef]
- Feng, C.W.; Chen, N.F.; Wen, Z.H.; Yang, W.Y.; Kuo, H.M.; Sung, P.J.; Su, J.H.; Cheng, S.Y.; Chen, W.F. In Vitro and In Vivo Neuroprotective Effects of Stellettin B Through Anti-Apoptosis and the Nrf2/HO-1 Pathway. Mar. Drugs 2019, 17, 315. [Google Scholar] [CrossRef]
- Hwang, J.; Kim, D.; Park, J.S.; Park, H.J.; Shin, J.; Lee, S.K. Photoprotective Activity of Topsentin, A Bis(Indole) Alkaloid from the Marine Sponge Spongosorites genitrix, by Regulation of COX-2 and Mir-4485 Expression in UVB-Irradiated Human Keratinocyte Cells. Mar. Drugs 2020, 18, 87. [Google Scholar] [CrossRef]
- Josse, R.; Zhang, Y.W.; Giroux, V.; Ghosh, A.K.; Luo, J.; Pommier, Y. Activation of RAF1 (c-RAF) by the Marine Alkaloid Lasonolide A Induces Rapid Premature Chromosome Condensation. Mar. Drugs 2015, 13, 3625–3639. [Google Scholar] [CrossRef]
- Zovko, A.; Viktorsson, K.; Haag, P.; Kovalerchick, D.; Farnegardh, K.; Alimonti, A.; Ilan, M.; Carmeli, S.; Lewensohn, R. Marine sponge Cribrochalina vasculum compounds activate intrinsic apoptotic signaling and inhibit growth factor signaling cascades in non-small cell lung carcinoma. Mol. Cancer Ther. 2014, 13, 2941–2954. [Google Scholar] [CrossRef]
- Shrestha, S.; Sorolla, A.; Fromont, J.; Blancafort, P.; Flematti, G.R. Crambescidin 800, Isolated from the Marine Sponge Monanchora viridis, Induces Cell Cycle Arrest and Apoptosis in Triple-Negative Breast Cancer Cells. Mar. Drugs 2018, 16, 53. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, S.; Sorolla, A.; Fromont, J.; Blancafort, P.; Flematti, G.R. Aurantoside C Targets and Induces Apoptosis in Triple Negative Breast Cancer Cells. Mar. Drugs 2018, 16, 361. [Google Scholar] [CrossRef] [PubMed]
- Dyshlovoy, S.A.; Tabakmakher, K.M.; Hauschild, J.; Shchekaleva, R.K.; Otte, K.; Guzii, A.G.; Makarieva, T.N.; Kudryashova, E.K.; Fedorov, S.N.; Shubina, L.K.; et al. Guanidine Alkaloids from the Marine Sponge Monanchora pulchra Show Cytotoxic Properties and Prevent EGF-Induced Neoplastic Transformation In Vitro. Mar. Drugs 2016, 14, 133. [Google Scholar] [CrossRef] [PubMed]
- Tasdemir, D.; Mallon, R.; Greenstein, M.; Feldberg, L.R.; Kim, S.C.; Collins, K.; Wojciechowicz, D.; Mangalindan, G.C.; Concepcion, G.P.; Harper, M.K.; et al. Aldisine alkaloids from the Philippine sponge Stylissa massa are potent inhibitors of mitogen-activated protein kinase kinase-1 (MEK-1). J. Med. Chem. 2002, 45, 529–532. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.K.; Lee, S.M.; Kim, N.H.; Tu, N.V.; Kim, Y.N.; Heo, J.D.; Jeong, E.J.; Rho, J.R. Stereochemical Determination of Fistularins Isolated from the Marine Sponge Ecionemia acervus and Their Regulatory Effect on Intestinal Inflammation. Mar. Drugs 2021, 19, 170. [Google Scholar] [CrossRef] [PubMed]
- Ecoy, G.A.U.; Chamni, S.; Suwanborirux, K.; Chanvorachote, P.; Chaotham, C. Jorunnamycin A from Xestospongia sp. Suppresses Epithelial to Mesenchymal Transition and Sensitizes Anoikis in Human Lung Cancer Cells. J. Nat. Prod. 2019, 82, 1861–1873. [Google Scholar] [CrossRef] [PubMed]
- Tun, J.O.; Salvador-Reyes, L.A.; Velarde, M.C.; Saito, N.; Suwanborirux, K.; Concepcion, G.P. Synergistic Cytotoxicity of Renieramycin M and Doxorubicin in MCF-7 Breast Cancer Cells. Mar. Drugs 2019, 17, 536. [Google Scholar] [CrossRef]
- Chantarawong, W.; Chamni, S.; Suwanborirux, K.; Saito, N.; Chanvorachote, P. 5-O-Acetyl-Renieramycin T from Blue Sponge Xestospongia sp. Induces Lung Cancer Stem Cell Apoptosis. Mar. Drugs 2019, 17, 109. [Google Scholar] [CrossRef]
- Pereira, A.R.; Strangman, W.K.; Marion, F.; Feldberg, L.; Roll, D.; Mallon, R.; Hollander, I.; Andersen, R.J. Synthesis of phosphatidylinositol 3-kinase (PI3K) inhibitory analogues of the sponge meroterpenoid liphagal. J. Med. Chem. 2010, 53, 8523–8533. [Google Scholar] [CrossRef]
- Chikamatsu, S.; Saijo, K.; Imai, H.; Narita, K.; Kawamura, Y.; Katoh, T.; Ishioka, C. In Vitro and in Vivo antitumor activity and the mechanism of siphonodictyal B in human colon cancer cells. Cancer Med. 2019, 8, 5662–5672. [Google Scholar] [CrossRef]
- Zhao, H.M.; He, J.; Chang, Y.T.; Liu, L.Y.; Sun, F.; Lin, H.W.; Yang, F. Marine Sponge-Derived Alkaloid Induces Mitochondrial Dysfunction and Inhibits the PI3K/AKT/mTOR Signaling Pathway against Burkitt’s Lymphoma. J. Nat. Prod. 2023, 86, 45–51. [Google Scholar] [CrossRef]
- Vidal, I.; Castilla, L.; Marrero, A.D.; Bravo-Ruiz, I.; Bernal, M.; Manrique, I.; Quesada, A.R.; Medina, M.; Martínez-Poveda, B. The Sponge-Derived Brominated Compound Aeroplysinin-1 Impairs the Endothelial Inflammatory Response through Inhibition of the NF-κB Pathway. Mar. Drugs 2022, 20, 605. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.; Kim, G.D.; Jeon, J.E.; Shin, J.; Lee, S.K. Antimetastatic effect of halichondramide, a trisoxazole macrolide from the marine sponge Chondrosia corticata, on human prostate cancer cells via modulation of epithelial-to-mesenchymal transition. Mar. Drugs 2013, 11, 2472–2485. [Google Scholar] [CrossRef] [PubMed]
- Su, J.H.; Chen, Y.C.; El-Shazly, M.; Du, Y.C.; Su, C.W.; Tsao, C.W.; Liu, L.L.; Chou, Y.; Chang, W.B.; Su, Y.D.; et al. Towards the small and the beautiful: A small dibromotyrosine derivative from Pseudoceratina sp. sponge exhibits potent apoptotic effect through targeting IKK/NFκB signaling pathway. Mar. Drugs 2013, 11, 3168–3185. [Google Scholar] [CrossRef]
- Li, H.; Zhang, Q.; Jin, X.; Zou, X.; Wang, Y.; Hao, D.; Fu, F.; Jiao, W.; Zhang, C.; Lin, H.; et al. Dysifragilone A inhibits LPS-induced RAW264.7 macrophage activation by blocking the p38 MAPK signaling pathway. Mol. Med. Rep. 2018, 17, 674–682. [Google Scholar] [CrossRef]
- Zhou, M.; Peng, B.R.; Tian, W.; Su, J.H.; Wang, G.; Lin, T.; Zeng, D.; Sheu, J.H.; Chen, H. 12-Deacetyl-12-epi-Scalaradial, a Scalarane Sesterterpenoid from a Marine Sponge Hippospongia sp., Induces HeLa Cells Apoptosis via MAPK/ERK Pathway and Modulates Nuclear Receptor Nur77. Mar. Drugs 2020, 18, 375. [Google Scholar] [CrossRef]
- Chen, Y.; Zhou, Q.; Zhang, L.; Zhong, Y.; Fan, G.; Zhang, Z.; Wang, R.; Jin, M.; Qiu, Y.; Kong, D. Stellettin B induces apoptosis in human chronic myeloid leukemia cells via targeting PI3K and Stat5. Oncotarget 2017, 8, 28906–28921. [Google Scholar] [CrossRef]
- Ciaglia, E.; Malfitano, A.M.; Laezza, C.; Fontana, A.; Nuzzo, G.; Cutignano, A.; Abate, M.; Pelin, M.; Sosa, S.; Bifulco, M.; et al. Immuno-Modulatory and Anti-Inflammatory Effects of Dihydrogracilin A, a Terpene Derived from the Marine Sponge Dendrilla membranosa. Int. J. Mol. Sci. 2017, 18, 1643. [Google Scholar] [CrossRef]
- Chang, C.H.; Lin, B.J.; Chen, C.H.; Nguyen, N.L.; Hsieh, T.H.; Su, J.H.; Chen, M.C. Stellettin B Induces Cell Death in Bladder Cancer via Activating the Autophagy/DAPK2/Apoptosis Signaling Cascade. Mar. Drugs 2023, 21, 73. [Google Scholar] [CrossRef]
- Lee, R.H.; Slate, D.L.; Moretti, R.; Alvi, K.A.; Crews, P. Marine sponge polyketide inhibitors of protein tyrosine kinase. Biochem. Biophys. Res. Commun. 1992, 184, 765–772. [Google Scholar] [CrossRef]
- Zovko, A.; Novak, M.; Haag, P.; Kovalerchick, D.; Holmlund, T.; Farnegardh, K.; Ilan, M.; Carmeli, S.; Lewensohn, R.; Viktorsson, K. Compounds from the marine sponge Cribrochalina vasculum offer a way to target IGF-1R mediated signaling in tumor cells. Oncotarget 2016, 7, 50258–50276. [Google Scholar] [CrossRef] [PubMed]
- Luyao, H.; Luesch, H.; Uy, M. GPCR Pharmacological Profiling of Aaptamine from the Philippine Sponge Stylissa sp. Extends Its Therapeutic Potential for Noncommunicable Diseases. Molecules 2021, 26, 5618. [Google Scholar] [CrossRef] [PubMed]
- Johnson, T.A.; Milan-Lobo, L.; Che, T.; Ferwerda, M.; Lambu, E.; McIntosh, N.L.; Li, F.; He, L.; Lorig-Roach, N.; Crews, P.; et al. Identification of the First Marine-Derived Opioid Receptor “Balanced” Agonist with a Signaling Profile That Resembles the Endorphins. ACS Chem. Neurosci. 2017, 8, 473–485. [Google Scholar] [CrossRef] [PubMed]
- Kreuter, M.H.; Leake, R.E.; Rinaldi, F.; Muller-Klieser, W.; Maidhof, A.; Muller, W.E.; Schroder, H.C. Inhibition of intrinsic protein tyrosine kinase activity of EGF-receptor kinase complex from human breast cancer cells by the marine sponge metabolite (+)-aeroplysinin-1. Comp. Biochem. Physiol. B 1990, 97, 151–158. [Google Scholar] [CrossRef]
- Rateb, M.E.; Houssen, W.E.; Schumacher, M.; Harrison, W.T.; Diederich, M.; Ebel, R.; Jaspars, M. Bioactive diterpene derivatives from the marine sponge Spongionella sp. J. Nat. Prod. 2009, 72, 1471–1476. [Google Scholar] [CrossRef] [PubMed]
- James, R.D.; Jones, D.A.; Aalbersberg, W.; Ireland, C.M. Naamidine A intensifies the phosphotransferase activity of extracellular signal-regulated kinases causing A-431 cells to arrest in G1. Mol. Cancer Ther. 2003, 2, 747–751. [Google Scholar] [PubMed]
- Patel, A.; Rasheed, A.; Reilly, I.; Pareek, Z.; Hansen, M.; Haque, Z.; Simon-Fajardo, D.; Davies, C.; Tummala, A.; Reinhardt, K.; et al. Modulation of Cytoskeleton, Protein Trafficking, and Signaling Pathways by Metabolites from Cucurbitaceae, Ericaceae, and Rosaceae Plant Families. Pharmaceuticals 2022, 15, 1380. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Chen, J.; Hu, G.; Yu, J.; Zhu, X.; Lin, Y.; Chen, S.; Yuan, J. Statistical research on the bioactivity of new marine natural products discovered during the 28 years from 1985 to 2012. Mar Drugs 2015, 13, 202–221. [Google Scholar] [CrossRef]
- Giordano, D.; Costantini, M.; Coppola, D.; Lauritano, C.; Nunez Pons, L.; Ruocco, N.; di Prisco, G.; Ianora, A.; Verde, C. Biotechnological Applications of Bioactive Peptides From Marine Sources. Adv. Microb. Physiol. 2018, 73, 171–220. [Google Scholar] [CrossRef]
- Munro, M.H.; Blunt, J.W.; Dumdei, E.J.; Hickford, S.J.; Lill, R.E.; Li, S.; Battershill, C.N.; Duckworth, A.R. The discovery and development of marine compounds with pharmaceutical potential. J. Biotechnol. 1999, 70, 15–25. [Google Scholar] [CrossRef]
- Blunt, J.W.; Copp, B.R.; Munro, M.H.; Northcote, P.T.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2011, 28, 196–268. [Google Scholar] [CrossRef] [PubMed]
- Schippers, K.J.; Martens, D.E.; Pomponi, S.A.; Wijffels, R.H. Cell cycle analysis of primary sponge cell cultures. In Vitro Cell. Dev. Biol. Anim. 2011, 47, 302–311. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Khalifa, S.A.M.; Elias, N.; Farag, M.A.; Chen, L.; Saeed, A.; Hegazy, M.F.; Moustafa, M.S.; Abd El-Wahed, A.; Al-Mousawi, S.M.; Musharraf, S.G.; et al. Marine Natural Products: A Source of Novel Anticancer Drugs. Mar. Drugs 2019, 17, 491. [Google Scholar] [CrossRef]
- Proksch, P. Defensive roles for secondary metabolites from marine sponges and sponge-feeding nudibranchs. Toxicon 1994, 32, 639–655. [Google Scholar] [CrossRef]
- Hertiani, T.; Edrada-Ebel, R.; Ortlepp, S.; van Soest, R.W.; de Voogd, N.J.; Wray, V.; Hentschel, U.; Kozytska, S.; Muller, W.E.; Proksch, P. From anti-fouling to biofilm inhibition: New cytotoxic secondary metabolites from two Indonesian Agelas sponges. Bioorg. Med. Chem. 2010, 18, 1297–1311. [Google Scholar] [CrossRef]
- Dobretsov, S.; Coutinho, R.; Rittschof, D.; Salta, M.; Ragazzola, F.; Hellio, C. The oceans are changing: Impact of ocean warming and acidification on biofouling communities. Biofouling 2019, 35, 585–595. [Google Scholar] [CrossRef]
- Sipkema, D.; Osinga, R.; Schatton, W.; Mendola, D.; Tramper, J.; Wijffels, R.H. Large-scale production of pharmaceuticals by marine sponges: Sea, cell, or synthesis? Biotechnol. Bioeng. 2005, 90, 201–222. [Google Scholar] [CrossRef]
- Pomponi, S.A. The bioprocess-technological potential of the sea. J. Biotechnol. 1999, 70, 5–13. [Google Scholar] [CrossRef]
- Ortega, V.; Cortes, J. Potential clinical applications of halichondrins in breast cancer and other neoplasms. Breast Cancer Targets Ther. 2012, 4, 9–19. [Google Scholar] [CrossRef]
- McBride, A.; Butler, S.K. Eribulin mesylate: A novel halichondrin B analogue for the treatment of metastatic breast cancer. Am. J. Health-Syst. Pharm. 2012, 69, 745–755. [Google Scholar] [CrossRef]
- Wijffels, R.H. Potential of sponges and microalgae for marine biotechnology. Trends Biotechnol. 2008, 26, 26–31. [Google Scholar] [CrossRef]
- Schmidt, E.W.; Lin, Z. Translating Marine Symbioses toward Drug Development. mBio 2022, 13, e02499-22. [Google Scholar] [CrossRef] [PubMed]
- Schippers, K.J.; Sipkema, D.; Osinga, R.; Smidt, H.; Pomponi, S.A.; Martens, D.E.; Wijffels, R.H. Cultivation of sponges, sponge cells and symbionts: Achievements and future prospects. Adv. Mar. Biol. 2012, 62, 273–337. [Google Scholar] [CrossRef] [PubMed]
- Schroder, H.C.; Brummer, F.; Fattorusso, E.; Aiello, A.; Menna, M.; de Rosa, S.; Batel, R.; Muller, W.E. Sustainable production of bioactive compounds from sponges: Primmorphs as bioreactors. In Sponges (Porifera); Progress in Molecular and Subcellular Biology Book Series; Springer: Berlin/Heidelberg, Germany, 2003; Volume 37, pp. 163–197. [Google Scholar] [CrossRef]
- Koopmans, M.; Martens, D.; Wijffels, R.H. Towards commercial production of sponge medicines. Mar. Drugs 2009, 7, 787–802. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.S.; Johnston, S.A.; Riedy, M.; DeVit, M.J.; McElligott, S.G.; Sanford, J.C. Introduction of foreign genes into tissues of living mice by DNA-coated microprojectiles. Proc. Natl. Acad. Sci. USA 1991, 88, 2726–2730. [Google Scholar] [CrossRef] [PubMed]
- Fortman, J.L.; Sherman, D.H. Utilizing the power of microbial genetics to bridge the gap between the promise and the application of marine natural products. ChemBioChem 2005, 6, 960–978. [Google Scholar] [CrossRef]
- Storey, M.A.; Andreassend, S.K.; Bracegirdle, J.; Brown, A.; Keyzers, R.A.; Ackerley, D.F.; Northcote, P.T.; Owen, J.G. Metagenomic Exploration of the Marine Sponge Mycale hentscheli Uncovers Multiple Polyketide-Producing Bacterial Symbionts. mBio 2020, 11, e02997-19. [Google Scholar] [CrossRef]
- Song, Y.F.; Qu, Y.; Cao, X.P.; Zhang, W. Cellular localization of debromohymenialdisine and hymenialdisine in the marine sponge Axinella sp. using a newly developed cell purification protocol. Mar. Biotechnol. 2011, 13, 868–882. [Google Scholar] [CrossRef]
- Knapp, I.S.S.; Forsman, Z.H.; Greene, A.; Johnston, E.C.; Bardin, C.E.; Chan, N.; Wolke, C.; Gulko, D.; Toonen, R.J. Coral micro-fragmentation assays for optimizing active reef restoration efforts. PeerJ 2022, 10, e13653. [Google Scholar] [CrossRef]
- Duckworth, A. Farming sponges to supply bioactive metabolites and bath sponges: A review. Mar. Biotechnol. 2009, 11, 669–679. [Google Scholar] [CrossRef] [PubMed]
- Bahr, K.D.; Jokiel, P.L.; Toonen, R.J. The unnatural history of Kāne‘ohe Bay: Coral reef resilience in the face of centuries of anthropogenic impacts. PeerJ 2015, 3, e950. [Google Scholar] [CrossRef] [PubMed]
- Fabricius, K.E. Effects of terrestrial runoff on the ecology of corals and coral reefs: Review and synthesis. Mar. Pollut. Bull. 2005, 50, 125–146. [Google Scholar] [CrossRef] [PubMed]
- Williams, G.J.; Knapp, I.S.; Maragos, J.E.; Davy, S.K. Modeling patterns of coral bleaching at a remote Central Pacific atoll. Mar. Pollut. Bull. 2010, 60, 1467–1476. [Google Scholar] [CrossRef] [PubMed]
- Belarbi el, H.; Contreras Gomez, A.; Chisti, Y.; Garcia Camacho, F.; Molina Grima, E. Producing drugs from marine sponges. Biotechnol. Adv. 2003, 21, 585–598. [Google Scholar] [CrossRef] [PubMed]
- Bergman, O.; Mayzel, B.; Anderson, M.A.; Shpigel, M.; Hill, R.T.; Ilan, M. Examination of marine-based cultivation of three demosponges for acquiring bioactive marine natural products. Mar. Drugs 2011, 9, 2201–2219. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, P.M.; Alsaggaf, A.A.; Villela, H.M.; Peixoto, R.S. Inducing Polyp Bail-out in Coral Colonies to Obtain Individualized Micropropagates for Laboratory Experimental Use. J. Vis. Exp. 2022, 182, e63840. [Google Scholar] [CrossRef]
- Hughes, T.P.; Kerry, J.T.; Baird, A.H.; Connolly, S.R.; Chase, T.J.; Dietzel, A.; Hill, T.; Hoey, A.S.; Hoogenboom, M.O.; Jacobson, M.; et al. Global warming impairs stock-recruitment dynamics of corals. Nature 2019, 568, 387–390. [Google Scholar] [CrossRef]
- Webster, N.S.; Cobb, R.E.; Negri, A.P. Temperature thresholds for bacterial symbiosis with a sponge. ISME J. 2008, 2, 830–842. [Google Scholar] [CrossRef]
- Webster, N.S.; Webb, R.I.; Ridd, M.J.; Hill, R.T.; Negri, A.P. The effects of copper on the microbial community of a coral reef sponge. Environ. Microbiol. 2001, 3, 19–31. [Google Scholar] [CrossRef]
- Webster, N.S.; Xavier, J.R.; Freckelton, M.; Motti, C.A.; Cobb, R. Shifts in microbial and chemical patterns within the marine sponge Aplysina aerophoba during a disease outbreak. Environ. Microbiol. 2008, 10, 3366–3376. [Google Scholar] [CrossRef] [PubMed]
- Padiglia, A.; Ledda, F.D.; Padedda, B.M.; Pronzato, R.; Manconi, R. Long-term experimental in situ farming of Crambe crambe (Demospongiae: Poecilosclerida). PeerJ 2018, 6, e4964. [Google Scholar] [CrossRef] [PubMed]
- Webster, N.S.; Cobb, R.E.; Soo, R.; Anthony, S.L.; Battershill, C.N.; Whalan, S.; Evans-Illidge, E. Bacterial community dynamics in the marine sponge Rhopaloeides odorabile under in situ and ex situ cultivation. Mar. Biotechnol. 2011, 13, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.L.; Mayfield, A.B.; Fan, T.Y. Effects of feeding on the physiological performance of the stony coral Pocillopora acuta. Sci. Rep. 2020, 10, 19988. [Google Scholar] [CrossRef]
- Knobloch, S.; Johannsson, R.; Marteinsson, V. Co-cultivation of the marine sponge Halichondria panicea and its associated microorganisms. Sci. Rep. 2019, 9, 10403. [Google Scholar] [CrossRef]
- Mohamed, N.M.; Rao, V.; Hamann, M.T.; Kelly, M.; Hill, R.T. Monitoring bacterial diversity of the marine sponge Ircinia strobilina upon transfer into aquaculture. Appl. Environ. Microbiol. 2008, 74, 4133–4143. [Google Scholar] [CrossRef]
- Isaacs, L.T.; Kan, J.; Nguyen, L.; Videau, P.; Anderson, M.A.; Wright, T.L.; Hill, R.T. Comparison of the bacterial communities of wild and captive sponge Clathria prolifera from the Chesapeake Bay. Mar. Biotechnol. 2009, 11, 758–770. [Google Scholar] [CrossRef]
- Mohamed, N.M.; Enticknap, J.J.; Lohr, J.E.; McIntosh, S.M.; Hill, R.T. Changes in bacterial communities of the marine sponge Mycale laxissima on transfer into aquaculture. Appl. Environ. Microbiol. 2008, 74, 1209–1222. [Google Scholar] [CrossRef]
- Lippmann, R.B.; Helmstedt, K.J.; Gibbs, M.T.; Corry, P. Optimizing facility location, sizing, and growth time for a cultivated resource: A case study in coral aquaculture. PLoS ONE 2023, 18, e0282668. [Google Scholar] [CrossRef]
- Lecointe, A.; Cohen, S.; Geze, M.; Djediat, C.; Meibom, A.; Domart-Coulon, I. Scleractinian coral cell proliferation is reduced in primary culture of suspended multicellular aggregates compared to polyps. Cytotechnology 2013, 65, 705–724. [Google Scholar] [CrossRef][Green Version]
- Garcia Camacho, F.; Chileh, T.; Ceron Garcia, M.C.; Sanchez Miron, A.; Belarbi, E.H.; Contreras Gomez, A.; Molina Grima, E. Sustained growth of explants from Mediterranean sponge Crambe crambe cultured in vitro with enriched RPMI 1640. Biotechnol. Prog. 2006, 22, 781–790. [Google Scholar] [CrossRef]
- Grasela, J.J.; Pomponi, S.A.; Rinkevich, B.; Grima, J. Efforts to develop a cultured sponge cell line: Revisiting an intractable problem. In Vitro Cell. Dev. Biol. Anim. 2012, 48, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Munroe, S.; Sandoval, K.; Martens, D.E.; Sipkema, D.; Pomponi, S.A. Genetic algorithm as an optimization tool for the development of sponge cell culture media. In Vitro Cell. Dev. Biol. Anim. 2019, 55, 149–158. [Google Scholar] [CrossRef]
- Hesp, K.; van der Heijden, J.M.E.; Munroe, S.; Sipkema, D.; Martens, D.E.; Wijffels, R.H.; Pomponi, S.A. First continuous marine sponge cell line established. Sci. Rep. 2023, 13, 5766. [Google Scholar] [CrossRef]
- Puntin, G.; Sweet, M.; Fraune, S.; Medina, M.; Sharp, K.; Weis, V.M.; Ziegler, M. Harnessing the Power of Model Organisms to Unravel Microbial Functions in the Coral Holobiont. Microbiol. Mol. Biol. Rev. 2022, 86, e00053-22. [Google Scholar] [CrossRef] [PubMed]
- Huete-Stauffer, C.; Valisano, L.; Gaino, E.; Vezzulli, L.; Cerrano, C. Development of long-term primary cell aggregates from Mediterranean octocorals. In Vitro Cell. Dev. Biol. Anim. 2015, 51, 815–826. [Google Scholar] [CrossRef] [PubMed]
- Rinkevich, B. Cell cultures from marine invertebrates: New insights for capturing endless stemness. Mar. Biotechnol. 2011, 13, 345–354. [Google Scholar] [CrossRef]
- Hardoim, C.C.; Cardinale, M.; Cucio, A.C.; Esteves, A.I.; Berg, G.; Xavier, J.R.; Cox, C.J.; Costa, R. Effects of sample handling and cultivation bias on the specificity of bacterial communities in keratose marine sponges. Front. Microbiol. 2014, 5, 611. [Google Scholar] [CrossRef]
- Sun, L.; Song, Y.; Qu, Y.; Yu, X.; Zhang, W. Purification and in vitro cultivation of archaeocytes (stem cells) of the marine sponge Hymeniacidon perleve (Demospongiae). Cell Tissue Res. 2007, 328, 223–237. [Google Scholar] [CrossRef]
- Vizel, M.; Loya, Y.; Downs, C.A.; Kramarsky-Winter, E. A novel method for coral explant culture and micropropagation. Mar. Biotechnol. 2011, 13, 423–432. [Google Scholar] [CrossRef]
- Lu, Q.; Liu, T.; Tang, X.; Dong, B.; Guo, H. Reformation of tissue balls from tentacle explants of coral Goniopora lobata: Self-organization process and response to environmental stresses. In Vitro Cell. Dev. Biol. Anim. 2017, 53, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, K.; Nishitsuji, K.; Shoguchi, E.; Fujiwara, S.; Satoh, N. Establishing Sustainable Cell Lines of a Coral, Acropora tenuis. Mar. Biotechnol. 2021, 23, 373–388. [Google Scholar] [CrossRef] [PubMed]
- Domart-Coulon, I.; Ostrander, G.K. Coral Cell and Tissue Culture Methods. In Diseases of Coral; Wiley Online Library, John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015. [Google Scholar]
- Richelle-Maurer, E.; Gomez, R.; Braekman, J.C.; Van de Vyver, G.; Van Soest, R.W.; Devijver, C. Primary cultures from the marine sponge Xestospongia muta (Petrosiidae, Haplosclerida). J. Biotechnol. 2003, 100, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhang, X.; Cao, X.; Xu, J.; Zhao, Q.; Yu, X.; Jin, M.; Deng, M. Optimizing the formation of in vitro sponge primmorphs from the Chinese sponge Stylotella agminata (Ridley). J. Biotechnol. 2003, 100, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Le Pennec, G.; Perovic, S.; Ammar, M.S.; Grebenjuk, V.A.; Steffen, R.; Brummer, F.; Muller, W.E. Cultivation of primmorphs from the marine sponge Suberites domuncula: Morphogenetic potential of silicon and iron. J. Biotechnol. 2003, 100, 93–108. [Google Scholar] [CrossRef] [PubMed]
- Custodio, M.R.; Prokic, I.; Steffen, R.; Koziol, C.; Borojevic, R.; Brummer, F.; Nickel, M.; Muller, W.E. Primmorphs generated from dissociated cells of the sponge Suberites domuncula: A model system for studies of cell proliferation and cell death. Mech. Ageing Dev. 1998, 105, 45–59. [Google Scholar] [CrossRef]
- Muller, W.E.; Bohm, M.; Batel, R.; De Rosa, S.; Tommonaro, G.; Muller, I.M.; Schroder, H.C. Application of cell culture for the production of bioactive compounds from sponges: Synthesis of avarol by primmorphs from Dysidea avara. J. Nat. Prod. 2000, 63, 1077–1081. [Google Scholar] [CrossRef]
- Zhang, X.; Cao, X.; Zhang, W.; Yu, X.; Jin, M. Primmorphs from archaeocytes-dominant cell population of the sponge hymeniacidon perleve: Improved cell proliferation and spiculogenesis. Biotechnol. Bioeng. 2003, 84, 583–590. [Google Scholar] [CrossRef]
- Muller, W.E.; Grebenjuk, V.A.; Le Pennec, G.; Schroder, H.; Brummer, F.; Hentschel, U.; Muller, I.M.; Breter, H. Sustainable production of bioactive compounds by sponges—Cell culture and gene cluster approach: A review. Mar. Biotechnol. 2004, 6, 105–117. [Google Scholar] [CrossRef]
- Pozzolini, M.; Valisano, L.; Cerrano, C.; Menta, M.; Schiaparelli, S.; Bavestrello, G.; Benatti, U.; Giovine, M. Influence of rocky substrata on three-dimensional sponge cells model development. In Vitro Cell. Dev. Biol. Anim. 2010, 46, 140–147. [Google Scholar] [CrossRef]
- Hentschel, U.; Usher, K.M.; Taylor, M.W. Marine sponges as microbial fermenters. FEMS Microbiol. Ecol. 2006, 55, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, C.R. Net primary productivity in coral reef sponges. Science 1983, 219, 410–412. [Google Scholar] [CrossRef] [PubMed]
- Sipkema, D.; Franssen, M.C.; Osinga, R.; Tramper, J.; Wijffels, R.H. Marine sponges as pharmacy. Mar. Biotechnol. 2005, 7, 142–162. [Google Scholar] [CrossRef] [PubMed]
- Montalvo, N.F.; Davis, J.; Vicente, J.; Pittiglio, R.; Ravel, J.; Hill, R.T. Integration of culture-based and molecular analysis of a complex sponge-associated bacterial community. PLoS ONE 2014, 9, e90517. [Google Scholar] [CrossRef] [PubMed]
- Thinesh, T.; Meenatchi, R.; Lipton, A.N.; Anandham, R.; Jose, P.A.; Tang, S.L.; Seghal Kiran, G.; Selvin, J. Metagenomic sequencing reveals altered bacterial abundance during coral-sponge interaction: Insights into the invasive process of coral-killing sponge Terpios hoshinota. Microbiol. Res. 2020, 240, 126553. [Google Scholar] [CrossRef]
- Chaves-Fonnegra, A.; Riegl, B.; Zea, S.; Lopez, J.V.; Smith, T.; Brandt, M.; Gilliam, D.S. Bleaching events regulate shifts from corals to excavating sponges in algae-dominated reefs. Glob. Chang. Biol. 2018, 24, 773–785. [Google Scholar] [CrossRef]
- Van Oevelen, D.; Mueller, C.E.; Lundalv, T.; van Duyl, F.C.; de Goeij, J.M.; Middelburg, J.J. Niche overlap between a cold-water coral and an associated sponge for isotopically-enriched particulate food sources. PLoS ONE 2018, 13, e0194659. [Google Scholar] [CrossRef]
- Riley, K.L.; Wickliffe, L.C.; Jossart, J.A.; Morris, J.A., Jr.; Cockrell, M.C.; Culpepper, C.M.; Wander, S.; Sapp, P. Aquaculture Spatial Planning in Florida: A Pilot Study to Assess Potential Offshore Aquaculture Zones along Florida’s Gulf Coast. Technical Report Prepared by NCCOS and FDACS. 2023. Available online: https://repository.library.noaa.gov/view/noaa/55667/noaa_55667_DS1.pdf (accessed on 22 November 2023).














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scientific Name | Vernacular Name | Number of Records in NOAA | Venn Diagram Category |
|---|---|---|---|
| Depth < 50 m | |||
| Biemna | Demosponge | 1 | A |
| Cinachyra | Demosponge | 1 | A |
| Haliclona (Reniera) | Demosponge | 1 | A |
| Leptogorgia hebes | Gorgonian coral | 1 | A |
| Phacelocyathus flos | Stony coral (cup coral) | 1 | A |
| Phyllangia americana | Stony coral (cup coral) | 9 | A |
| Phyllangia americana americana | Stony coral (cup coral) | 1 | A |
| Phyllangia americana mouchezii | Stony coral (cup coral) | 4 | A |
| Stelletta kallitetilla | Demosponge | 1 | A |
| Depth ≥ 50 to <200 m | |||
| Aaptos pernucleata | Demosponge | 2 | B |
| Aiolochroia crassa | Demosponge | 2 | B |
| Antho (Acarnia) | Demosponge | 1 | B |
| Antipathes | Black coral | 16 | B |
| Antipathidae | Black coral | 3 | B |
| Aplysina | Demosponge | 1 | B |
| Aplysina cf. insularis | Demosponge | 1 | B |
| Aplysina fistularis | Demosponge | 1 | B |
| Asteropus | Demosponge | 1 | B |
| Axinella | Demosponge | 1 | B |
| Axinella corrugata | Demosponge | 3 | B |
| Axinella shoemakeri | Demosponge | 2 | B |
| Balanophyllia (Balanophyllia) floridana | Stony coral (cup coral) | 1 | B |
| Bebryce | Gorgonian coral | 3 | B |
| Bubaris | Demosponge | 2 | B |
| Callyspongia (Cladochalina) | Demosponge | 1 | B |
| Caryophyllia (Caryophyllia) horologium | Stony coral (cup coral) | 4 | B |
| cf. Leucetta | Calcareous sponge | 3 | B |
| cf. Leucosolenia | Calcareous sponge | 10 | B |
| Chelonaplysilla | Demosponge | 2 | B |
| Chondrilla | Demosponge | 1 | B |
| Chondrilla nucula | Demosponge | 1 | B |
| Chondrosia | Demosponge | 1 | B |
| Chondrosia reniformis | Demosponge | 2 | B |
| Cinachyrella alloclada | Demosponge | 7 | B |
| Cinachyrella arenosa | Demosponge | 1 | B |
| Cirrhipathes | Black coral | 2 | B |
| Cladocora arbuscula | Stony coral (branching) | 1 | B |
| Cladocora debilis | Stony coral (branching) | 1 | B |
| Clathria | Demosponge | 1 | B |
| Clathria (Microciona) | Demosponge | 5 | B |
| Cliona | Demosponge | 5 | B |
| Cliona varians | Demosponge | 14 | B |
| Dasmosmilia lymani | Stony coral (cup coral) | 1 | B |
| Desmacella | Demosponge | 1 | B |
| Diodogorgia nodulifera | Gorgonian coral | 2 | B |
| Dragmacidon reticulatum | Demosponge | 1 | B |
| Dysidea | Demosponge | 24 | B |
| Elatopathes | Black coral | 7 | B |
| Ellisella | Gorgonian coral | 6 | B |
| Ellisella atlantica | Gorgonian coral | 1 | B |
| Ellisella elongata | Gorgonian coral | 1 | B |
| Ellisellidae | Gorgonian coral | 1 | B |
| Epipolasis | Demosponge | 5 | B |
| Erylus trisphaerus | Demosponge | 2 | B |
| Eurypon | Demosponge | 1 | B |
| Flabellum (Flabellum) floridanum | Stony coral (cup coral) | 4 | B |
| Funiculina quadrangularis | Sea pen | 1 | B |
| Geodia | Demosponge | 2 | B |
| Geodia gibberosa | Demosponge | 2 | B |
| Halichondria | Demosponge | 6 | B |
| Halichondria (Halichondria) melanadocia | Demosponge | 3 | B |
| Haliclona | Demosponge | 2 | B |
| Haliclona (Gellius) | Demosponge | 1 | B |
| Higginsia | Demosponge | 1 | B |
| Higginsia strigilata | Demosponge | 1 | B |
| Hymedesmia | Demosponge | 1 | B |
| Hymeniacidon caerulea | Demosponge | 1 | B |
| Hypnogorgia pendula | Gorgonian coral | 12 | B |
| Igernella notabilis | Demosponge | 1 | B |
| Ircinia | Demosponge | 4 | B |
| Ircinia campana | Demosponge | 1 | B |
| Ircinia felix | Demosponge | 1 | B |
| Ircinia strobilina | Demosponge | 1 | B |
| Jaspis | Demosponge | 1 | B |
| Leptogorgia cardinalis | Gorgonian coral | 2 | B |
| Leptogorgia stheno | Gorgonian coral | 2 | B |
| Leucetta | Calcareous sponge | 1 | B |
| Leuconia | Calcareous sponge | 1 | B |
| Lissodendoryx (Anomodoryx) sigmata | Demosponge | 1 | B |
| Madracis | Stony coral (branching) | 4 | B |
| Madrepora oculata | Stony coral (branching) | 3 | B |
| Mycale (Mycale) laevis | Demosponge | 4 | B |
| Neofibularia nolitangere | Demosponge | 1 | B |
| Nicella | Gorgonian coral | 7 | B |
| Nicella guadalupensis | Gorgonian coral | 1 | B |
| Nicella toeplitzae | Gorgonian coral | 1 | B |
| Nidalia | Soft coral | 1 | B |
| Oculina diffusa | Stony coral (branching) | 3 | B |
| Oculina tenella | Stony coral (branching) | 3 | B |
| Oxysmilia rotundifolia | Stony coral (cup coral) | 1 | B |
| Placospongia melobesioides | Demosponge | 3 | B |
| Poecillastra | Demosponge | 3 | B |
| Riisea paniculata | Gorgonian coral | 1 | B |
| Scopalina ruetzleri | Demosponge | 1 | B |
| Siphonodictyon siphonum | Demosponge | 2 | B |
| Spheciospongia | Demosponge | 1 | B |
| Spheciospongia vesparium | Demosponge | 3 | B |
| Spirastrella | Demosponge | 1 | B |
| Spirastrella coccinea | Demosponge | 5 | B |
| Stelletta | Demosponge | 4 | B |
| Stelletta cf. grubii | Demosponge | 1 | B |
| Stichopathes luetkeni | Black coral | 1 | B |
| Stylaster | Lace coral | 1 | B |
| Swiftia exserta | Gorgonian coral | 2 | B |
| Telesto sanguinea | Stoloniferan coral | 8 | B |
| Terpios | Demosponge | 2 | B |
| Tethya | Demosponge | 2 | B |
| Tethya actinia | Demosponge | 1 | B |
| Tethya seychellensis | Demosponge | 4 | B |
| Thesea | Gorgonian coral | 2 | B |
| Timea | Demosponge | 3 | B |
| Timea mixta | Demosponge | 1 | B |
| Trochocyathus (Trochocyathus) rawsonii | Stony coral (cup coral) | 1 | B |
| Depth ≥ 200 to <1000 m | |||
| Acanella aurelia | Gorgonian coral | 1 | C |
| Acanthogorgiidae | Gorgonian coral | 1 | C |
| Aphrocallistes | Glass sponge | 2 | C |
| Balticina | Sea pen | 1 | C |
| Bathypathes pseudoalternata | Black coral | 2 | C |
| Bathypsammia | Stony coral (cup coral) | 1 | C |
| Caryophyllia (Caryophyllia) ambrosia ambrosia | Stony coral (cup coral) | 1 | C |
| Deltocyathoides stimpsonii | Stony coral (cup coral) | 1 | C |
| Deltocyathus | Stony coral (cup coral) | 1 | C |
| Desmophyllum pertusum | Stony coral (branching) | 7 | C |
| Flabellum (Ulocyathus) moseleyi | Stony coral (cup coral) | 13 | C |
| Fungiacyathus (Bathyactis) crispus | Stony coral (cup coral) | 1 | C |
| Fungiacyathus (Bathyactis) marenzelleri | Stony coral (cup coral) | 1 | C |
| Leiopathes glaberrima | Black coral | 3 | C |
| Paracyathus pulchellus | Stony coral (cup coral) | 1 | C |
| Plumarella | Gorgonian coral | 4 | C |
| Stylasteridae | Lace coral | 14 | C |
| Tethocyathus recurvatus | Stony coral (cup coral) | 1 | C |
| Depth ≥ 1000 to 3000 m | |||
| Anthomastinae | Soft coral | 4 | D |
| Caryophyllia (Caryophyllia) ambrosia | Stony coral (cup coral) | 1 | D |
| Chrysogorgiidae | Gorgonian coral | 1 | D |
| Cladorhizidae | Demosponge | 2 | D |
| Coralliidae | Gorgonian coral | 15 | D |
| Enallopsammia | Stony coral (branching) | 5 | D |
| Euplectella | Glass sponge | 9 | D |
| Euplectellidae | Glass sponge | 4 | D |
| Farreidae | Sponge (unspecified) | 1 | D |
| Heteropathes | Black coral | 3 | D |
| Hyalonema | Glass sponge | 1 | D |
| Iridogorgia | Gorgonian coral | 47 | D |
| Lepidisis | Gorgonian coral | 1 | D |
| Metallogorgia | Gorgonian coral | 6 | D |
| Narella | Gorgonian coral | 2 | D |
| Narella spectabilis | Gorgonian coral | 1 | D |
| Paragorgia | Gorgonian coral | 4 | D |
| Paragorgiidae | Gorgonian coral | 1 | D |
| Parantipathes | Black coral | 1 | D |
| Polymastia | Demosponge | 3 | D |
| Primnoidae | Gorgonian coral | 27 | D |
| Scleractinia | Stony coral (cup coral) | 5 | D |
| Stephanocyathus (Stephanocyathus) diadema | Stony coral (cup coral) | 3 | D |
| Tetilla | Demosponge | 1 | D |
| Umbellula | Sea pen | 4 | D |
| Victorgorgia | Gorgonian coral | 1 | D |
| Depth < 50 m and Depth ≥ 50 to <200 m | |||
| Clathria (Thalysias) juniperina | Demosponge | 3 | E |
| Leptogorgia euryale | Gorgonian coral | 7 | E |
| Leptogorgia medusa | Gorgonian coral | 4 | E |
| Lissodendoryx | Demosponge | 4 | E |
| Madrepora | Stony coral (branching) | 12 | E |
| Oculinidae | Stony coral (branching) | 2 | E |
| Depth ≥ 50 to <200 m and Depth ≥ 200 to <1000 m | |||
| Deltocyathus calcar | Stony coral (cup coral) | 6 | F |
| Dendrophylliidae | Stony coral (unspecified) | 4 | F |
| Mycale | Demosponge | 4 | F |
| Pennatulacea | Sea pen | 3 | F |
| Schizocyathus fissilis | Stony coral (cup coral) | 8 | F |
| Depth ≥ 200 to <1000 m and Depth ≥ 1000 to 3000 m | |||
| Acanella | Gorgonian coral | 14 | G |
| Acanella arbuscula | Gorgonian coral | 12 | G |
| Anthomastus | Soft coral | 10 | G |
| Bathypathes | Black coral | 77 | G |
| Caryophyllia (Caryophyllia) ambrosia caribbeana | Stony coral (cup coral) | 9 | G |
| Chondrocladia | Demosponge | 3 | G |
| Chrysogorgia | Gorgonian coral | 10 | G |
| Chrysogorgia elegans | Gorgonian coral | 21 | G |
| Deltocyathus italicus | Stony coral (cup coral) | 13 | G |
| Demospongiae | Demosponge | 41 | G |
| Hexactinellida | Glass sponge | 63 | G |
| Keratoisididae | Gorgonian coral | 125 | G |
| Muriceides | Gorgonian coral | 6 | G |
| Depth ≥ 50 to <200 m, Depth ≥ 200 to <1000 m, and Depth ≥ 1000 to 3000 m | |||
| Antipatharia | Black coral | 6 | J |
| Paramuricea | Gorgonian coral | 39 | J |
| Stichopathes | Black coral | 239 | J |
| Depth < 50 m, Depth ≥ 50 to <200 m, Depth ≥ 200 to <1000 m, and Depth ≥ 1000 to 3000 m | |||
| Porifera | Sponge (unspecified) | 100 | M |
| Depth ≥ 50 to <200 m and Depth ≥ 1000 to 3000 m | |||
| Alcyonacea | Gorgonian coral | 12 | O |
| Caryophylliidae | Stony coral (unspecified) | 2 | O |
| Plexauridae | Gorgonian coral | 12 | O |
| Swiftia | Gorgonian coral | 13 | O |
| Tanacetipathes | Black coral | 10 | O |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iskandar, M.; Ruiz-Houston, K.M.; Bracco, S.D.; Sharkasi, S.R.; Calabi Villarroel, C.L.; Desai, M.N.; Gerges, A.G.; Ortiz Lopez, N.A.; Xiao Barbero, M.; German, A.A.; et al. Deep-Sea Sponges and Corals off the Western Coast of Florida—Intracellular Mechanisms of Action of Bioactive Compounds and Technological Advances Supporting the Drug Discovery Pipeline. Mar. Drugs 2023, 21, 615. https://doi.org/10.3390/md21120615
Iskandar M, Ruiz-Houston KM, Bracco SD, Sharkasi SR, Calabi Villarroel CL, Desai MN, Gerges AG, Ortiz Lopez NA, Xiao Barbero M, German AA, et al. Deep-Sea Sponges and Corals off the Western Coast of Florida—Intracellular Mechanisms of Action of Bioactive Compounds and Technological Advances Supporting the Drug Discovery Pipeline. Marine Drugs. 2023; 21(12):615. https://doi.org/10.3390/md21120615
Chicago/Turabian StyleIskandar, Mina, Kira M. Ruiz-Houston, Steven D. Bracco, Sami R. Sharkasi, Cecilia L. Calabi Villarroel, Meghna N. Desai, Alexandra G. Gerges, Natalia A. Ortiz Lopez, Miguel Xiao Barbero, Amelia A. German, and et al. 2023. "Deep-Sea Sponges and Corals off the Western Coast of Florida—Intracellular Mechanisms of Action of Bioactive Compounds and Technological Advances Supporting the Drug Discovery Pipeline" Marine Drugs 21, no. 12: 615. https://doi.org/10.3390/md21120615
APA StyleIskandar, M., Ruiz-Houston, K. M., Bracco, S. D., Sharkasi, S. R., Calabi Villarroel, C. L., Desai, M. N., Gerges, A. G., Ortiz Lopez, N. A., Xiao Barbero, M., German, A. A., Moluguri, V. S., Walker, S. M., Silva Higashi, J., Palma, J. M., Medina, D. Z., Patel, M., Patel, P., Valentin, M., Diaz, A. C., ... Nanjundan, M. (2023). Deep-Sea Sponges and Corals off the Western Coast of Florida—Intracellular Mechanisms of Action of Bioactive Compounds and Technological Advances Supporting the Drug Discovery Pipeline. Marine Drugs, 21(12), 615. https://doi.org/10.3390/md21120615