
UV/H2O2-Degraded Polysaccharides from Sargassum fusiforme: Purification, Structural Properties, and Anti-Inflammatory Activity



Abstract
:1. Introduction
2. Results and Discussion
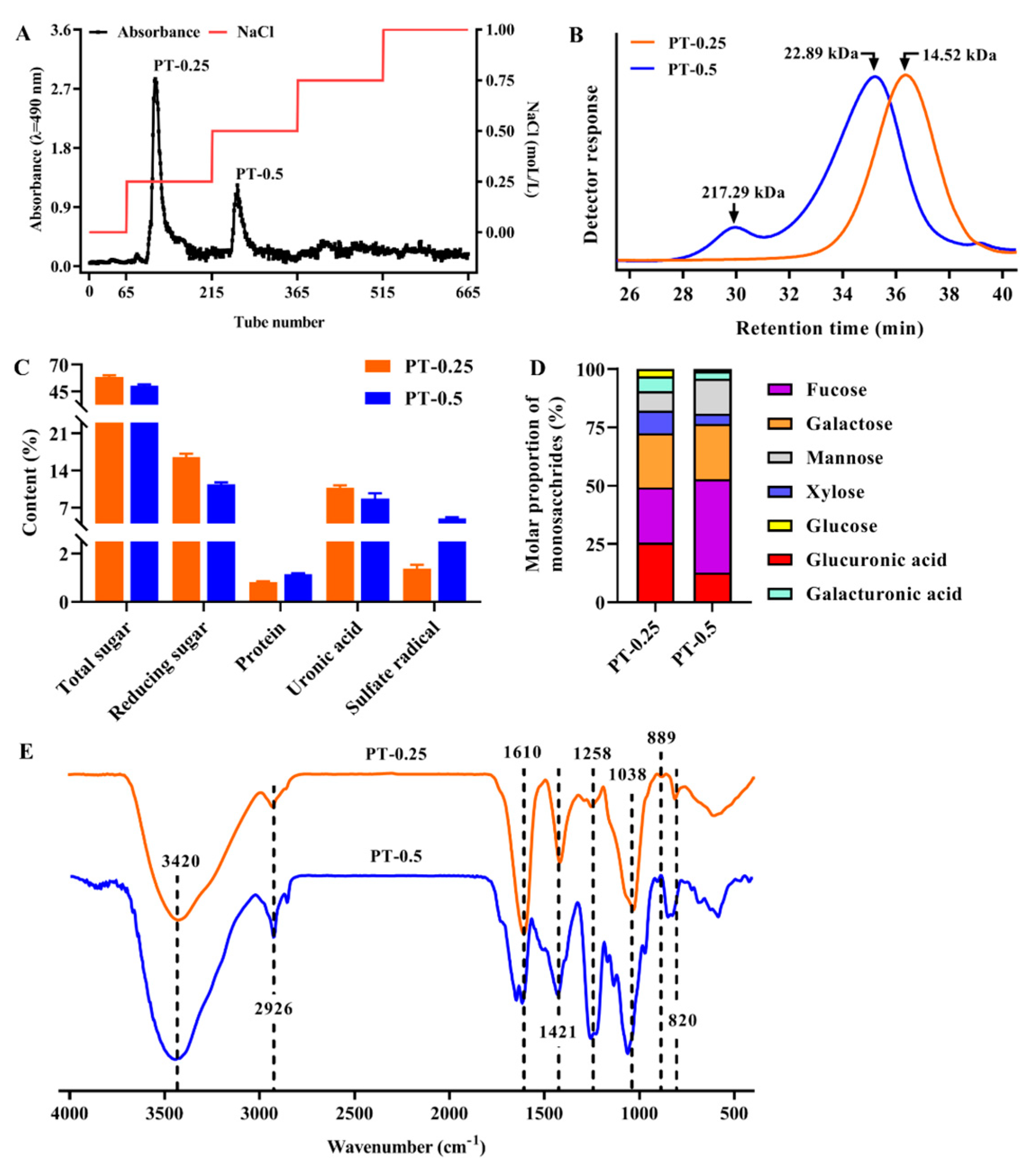
2.1. Isolation and Purification of Polysaccharides and Their Molecular Weight
2.2. Chemical Composition and Monosaccharide Composition of Purified Fractions
2.3. FT-IR Spectra of Purified Fractions
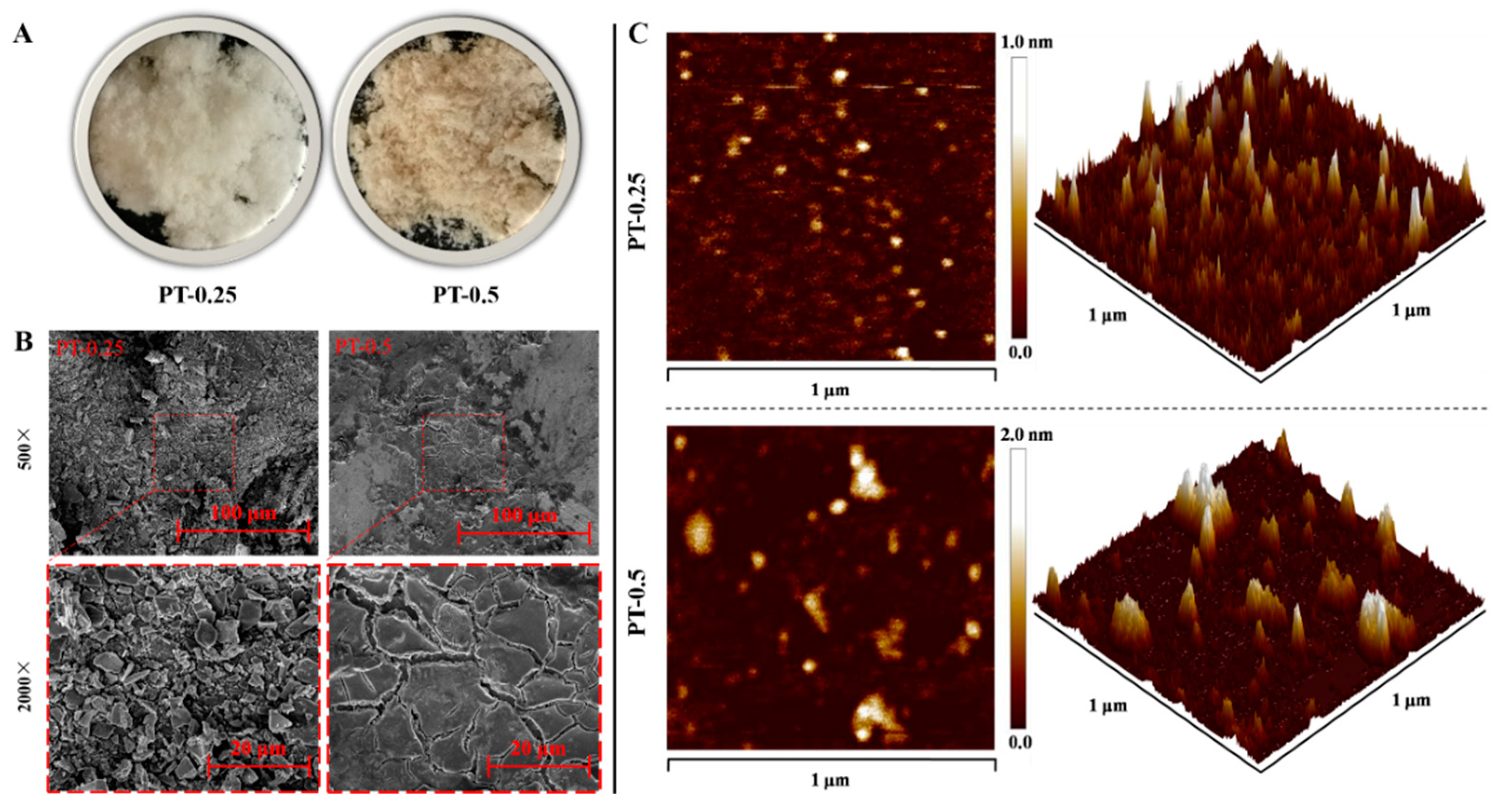
2.4. Morphological Characterization of Purified Fractions
2.5. Cell Viability of Purified Fractions
2.6. Purified Fractions Inhibited the Secretion of Pro-Inflammatory Mediators
2.7. Purified Fractions Inhibited the Expression of Related Pro-Inflammatory Genes
2.8. Methylation Analysis of PT-0.5 Fraction
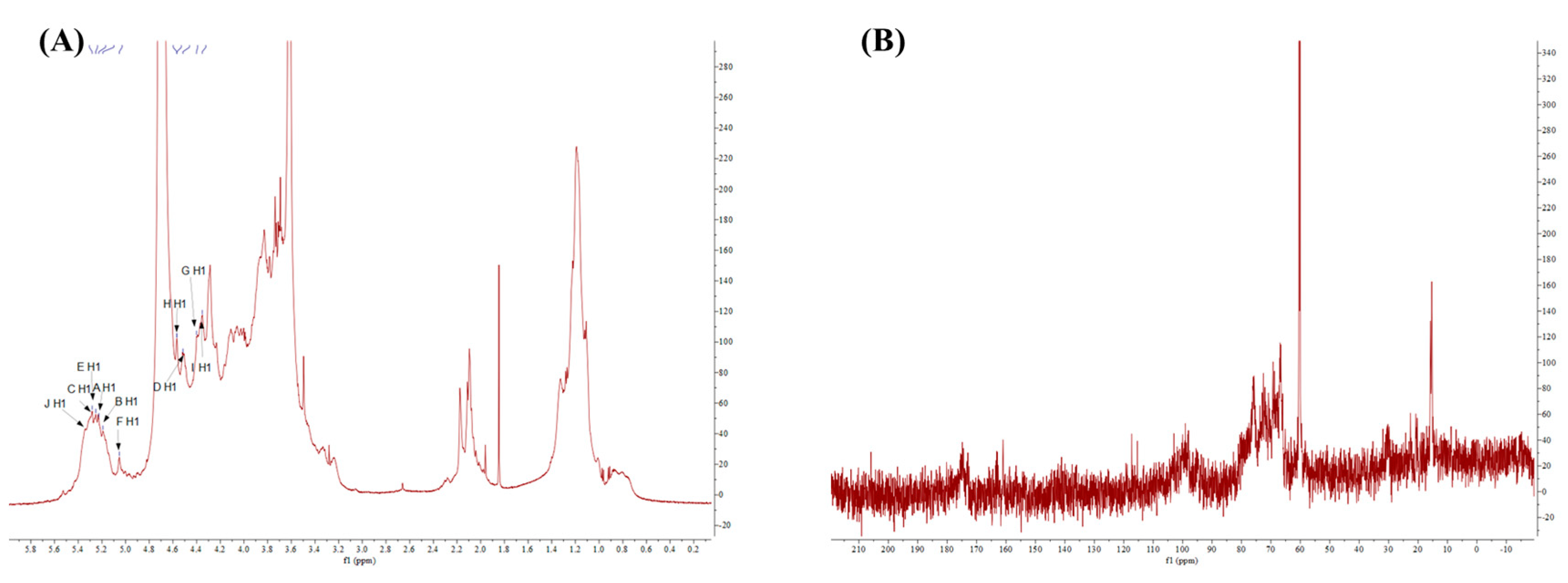
2.9. NMR Spectra of Purified Fraction with Best Anti-Inflammatory Activity
3. Materials and Methods
3.1. Preparation and Purification of PSF-T2
3.2. Chemical Composition and Molecular Weight Analysis
3.3. Monosaccharide Composition Analysis
3.4. Fourier Transform Infrared (FT-IR) Spectroscopy Analysis
3.5. Morphological Characterization
3.6. Anti-Inflammatory Activity Evaluation of Polysaccharide Fractions
3.6.1. Cell Culture and Cell Viability Analysis
3.6.2. Measurement of Pro-Inflammatory Mediators
3.6.3. qRT-PCR Analysis
3.6.4. Methylation Analysis
3.6.5. NMR Spectral Analysis
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Sun, H.; Gao, L.; Xue, C.; Mao, X. Marine-polysaccharide degrading enzymes: Status and prospects. Compr. Rev. Food Sci. Food Saf. 2020, 19, 2767–2796. [Google Scholar] [CrossRef]
- Xu, Y.; Wu, Y.-J.; Sun, P.-L.; Zhang, F.-M.; Linhardt, R.J.; Zhang, A.-Q. Chemically modified polysaccharides: Synthesis, characterization, structure activity relationships of action. Int. J. Biol. Macromol. 2019, 132, 970–977. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wang, J.; Fu, L.; Al-Wraikat, M.; Lin, S.; Lu, P.; Shan, L.; Fan, J.; Zhang, B. Degradation of polysaccharides from Lycium barbarum L. leaves improves bioaccessibility and gastrointestinal transport of endogenous minerals. Int. J. Biol. Macromol. 2020, 143, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Dou, Z.; Zhang, Y.; Tang, W.; Deng, Q.; Hu, B.; Chen, X.; Niu, H.; Wang, W.; Li, Z.; Zhou, H.; et al. Ultrasonic effects on the degradation kinetics, structural characteristics and protective effects on hepatocyte lipotoxicity induced by palmitic acid of Pueraria Lobata polysaccharides. Ultrason. Sonochem. 2023, 101, 106652. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.; Hu, J.; Zhu, B.; You, L.; Pedisić, S.; Hileuskaya, K. The effects of alginate lyase treatment on structural characteristics and anti-photoaging activity of alginate from Sargassum fusiforme. Food Front. 2023, 4, 807–817. [Google Scholar] [CrossRef]
- Chen, S.; Wang, X.; Guo, Y.; Song, X.; Yin, J.; Nie, S. Exploring the partial degradation of polysaccharides: Structure, mechanism, bioactivities, and perspectives. Compr. Rev. Food Sci. Food Saf. 2023. [Google Scholar] [CrossRef]
- Li, N.; Fu, X.; Xiao, M.; Wei, X.; Yang, M.; Liu, Z.; Mou, H. Enzymatic preparation of a low-molecular-weight polysaccharide rich in uronic acid from the seaweed Laminaria japonica and evaluation of its hypolipidemic effect in mice. Food Funct. 2020, 11, 2395–2405. [Google Scholar] [CrossRef]
- Xu, Y.; Niu, X.; Liu, N.; Gao, Y.; Wang, L.; Xu, G.; Li, X.; Yang, Y. Characterization, antioxidant and hypoglycemic activities of degraded polysaccharides from blackcurrant (Ribes nigrum L.) fruits. Food Chem. 2018, 243, 26–35. [Google Scholar] [CrossRef]
- Chen, B.-J.; Shi, M.-J.; Cui, S.; Hao, S.-X.; Hider, R.C.; Zhou, T. Improved antioxidant and anti-tyrosinase activity of poly-saccharide from Sargassum fusiforme by degradation. Int. J. Biol. Macromol. 2016, 92, 715–722. [Google Scholar] [CrossRef]
- Qian, W.-W.; Yang, S.-Q.; Hu, S.-M.; Wang, X.-L.; Zhu, Y.; Zhou, T. Enzymatic degradation, antioxidant and immunoregu-latory activities of polysaccharides from brown algae Sargassum fusiforme. J. Food Meas. Charact. 2021, 15, 1960–1972. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, R.; Li, Y.; Li, X.; You, L.; Kulikouskaya, V.; Hileuskaya, K. Degradation of polysaccharides from Sargassum fusiforme using UV/H2O2 and its effects on structural characteristics. Carbohydr. Polym. 2020, 230, 115647. [Google Scholar] [CrossRef]
- Chen, X.; Li, X.; Sun-Waterhouse, D.; Zhu, B.; You, L.; Hileuskaya, K. Polysaccharides from Sargassum fusiforme after UV/H2O2 degradation effectively ameliorate dextran sulfate sodium-induced colitis. Food Funct. 2021, 12, 11747–11759. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; You, L.; Ma, Y.; Zhao, Z.; Kulikouskaya, V. Influence of UV/H2O2 treatment on polysaccharides from Sargassum fusiforme: Physicochemical properties and RAW 264.7 cells responses. Food Chem. Toxicol. 2021, 153, 112246. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wang, W.; Huang, Y.; Liu, Z.; Zhang, J. Characterization of a novel polysaccharide purified from a herb of Cynomorium songaricum Rupr. Food Hydrocoll. 2015, 47, 79–86. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, X.; Huo, D.; Cao, C.; Li, Y.; Liang, Y.; Li, B.; Li, L. Preliminary characterization, antioxidant and α-glucosidase inhibitory activities of polysaccharides from Mallotus furetianus. Carbohydr. Polym. 2019, 215, 307–315. [Google Scholar] [CrossRef]
- Zhang, H.; Zou, P.; Zhao, H.; Qiu, J.; Mac Regenstein, J.; Yang, X. Isolation, purification, structure and antioxidant activity of polysaccharide from pinecones of Pinus koraiensis. Carbohydr. Polym. 2020, 251, 117078. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhang, S.; Wang, Q.; Lu, X.; Lin, L.; Tian, Y.; Xiao, J.; Zheng, B. Characterization and hypoglycemic activity of a β-pyran polysaccharides from bamboo shoot (Leleba oldhami Nakal) shells. Carbohydr. Polym. 2016, 144, 438–446. [Google Scholar] [CrossRef]
- Zhang, Y.; Dai, L.; Kong, X.; Chen, L. Characterization and in vitro antioxidant activities of polysaccharides from Pleurotus ostreatus. Int. J. Biol. Macromol. 2012, 51, 259–265. [Google Scholar] [CrossRef]
- Jia, R.-B.; Li, Z.-R.; Wu, J.; Ou, Z.-R.; Zhu, Q.; Sun, B.; Lin, L.; Zhao, M. Physicochemical properties of polysaccharide fractions from Sargassum fusiforme and their hypoglycemic and hypolipidemic activities in type 2 diabetic rats. Int. J. Biol. Macromol. 2020, 147, 428–438. [Google Scholar] [CrossRef]
- Ma, F.; Wang, D.; Zhang, Y.; Li, M.; Qing, W.; Tikkanen-Kaukanen, C.; Liu, X.; Bell, A.E. Characterisation of the mucilage polysaccharides from Dioscorea opposita Thunb. with enzymatic hydrolysis. Food Chem. 2017, 245, 13–21. [Google Scholar] [CrossRef]
- Gu, J.; Zhang, H.; Yao, H.; Zhou, J.; Duan, Y.; Ma, H. Comparison of characterization, antioxidant and immunological activities of three polysaccharides from Sagittaria sagittifolia L. Carbohydr. Polym. 2020, 235, 115939. [Google Scholar] [CrossRef] [PubMed]
- Liyanage, N.M.; Lee, H.-G.; Nagahawatta, D.P.; Jayawardhana, H.H.A.C.K.; Song, K.-M.; Choi, Y.-S.; Jeon, Y.-J.; Kang, M.-C. Fucoidan from Sargassum autumnale Inhibits Potential Inflammatory Responses via NF-κB and MAPK Pathway Suppression in Lipopolysaccharide-Induced RAW 264.7 Macrophages. Mar. Drugs 2023, 21, 374. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Oh, J.-Y.; Yang, H.-W.; Fu, X.; Kim, J.-I.; Jeon, Y.-J. Fucoidan isolated from the popular edible brown seaweed Sar-gassum fusiforme suppresses lipopolysaccharide-induced inflammation by blocking NF-κB signal pathway. J. Appl. Phycol. 2021, 33, 1845–1852. [Google Scholar] [CrossRef]
- Jayasinghe, A.M.K.; Kirindage, K.G.I.S.; Fernando, I.P.S.; Kim, K.-N.; Oh, J.-Y.; Ahn, G. The Anti-Inflammatory Effect of Low Molecular Weight Fucoidan from Sargassum siliquastrum in Lipopolysaccharide-Stimulated RAW 264.7 Macrophages via In-hibiting NF-κB/MAPK Signaling Pathways. Mar. Drugs 2023, 21, 347. [Google Scholar] [CrossRef]
- Sun, Q.-L.; Li, Y.; Ni, L.-Q.; Li, Y.-X.; Cui, Y.-S.; Jiang, S.-L.; Xie, E.-Y.; Du, J.; Deng, F.; Dong, C.-X. Structural characterization and antiviral activity of two fucoidans from the brown algae Sargassum henslowianum. Carbohydr. Polym. 2019, 229, 115487. [Google Scholar] [CrossRef]
- Zeb, M.; Tackaberry, L.E.; Massicotte, H.B.; Egger, K.N.; Reimer, K.; Lu, G.; Heiss, C.; Azadi, P.; Lee, C.H. Structural elucidation and immuno-stimulatory activity of a novel polysaccharide containing glucuronic acid from the fungus Echinodontium tincto-rium. Carbohydr. Polym. 2021, 258, 117700. [Google Scholar] [CrossRef]
- Hua, Y.-F.; Zhang, M.; Fu, C.-X.; Chen, Z.-H.; Chan, G.Y.S. Structural characterization of a 2-O-acetylglucomannan from Dendrobium officinale stem. Carbohydr. Res. 2004, 339, 2219–2224. [Google Scholar] [CrossRef]
- Perepelov, A.V.; Zabłotni, A.; Shashkov, A.S.; Knirel, Y.A.; Sidorczyk, Z. Structure of the O-polysaccharide and serological studies of the lipopolysaccharide of Proteus mirabilis 2002. Carbohydr. Res. 2005, 340, 2305–2310. [Google Scholar] [CrossRef]
- An, Y.; Liu, H.; Li, X.; Liu, J.; Chen, L.; Jin, X.; Chen, T.; Wang, W.; Liu, Z.; Zhang, M.; et al. Carboxymethylation modification, characterization, antioxidant activity and anti-UVC ability of Sargassum fusiforme polysaccharide. Carbohydr. Res. 2022, 515, 108555. [Google Scholar] [CrossRef]
- Hu, P.; Li, Z.X.; Chen, M.C.; Sun, Z.L.; Ling, Y.; Jiang, J.; Huang, C.G. Structural elucidation and protective role of a poly-saccharide from Sargassum fusiforme on ameliorating learning and memory deficiencies in mice. Carbohydr. Polym. 2016, 139, 150–158. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Blumenkrantz, N.; Asboe-Hansen, G. New method for quantitative determination of uronic acids. Anal. Biochem. 1973, 54, 484–489. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, H. A colorimetric estimation of reducing sugars in potatoes with 3,5-dinitrosalicylic acid. Potato Res. 1973, 16, 176–179. [Google Scholar] [CrossRef]
- Kawai, Y.; Seno, N.; Anno, K. A modified method for chondrosulfatase assay. Anal. Biochem. 1969, 32, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Ji, D.; You, L.; Zhou, L.; Zhao, Z.; Brennan, C. Structural properties and protective effect of Sargassum fusiforme poly-saccharides against ultraviolet B radiation in hairless Kun Ming mice. J. Funct. Foods 2018, 43, 8–16. [Google Scholar] [CrossRef]
- Xiao, H.; Fu, X.; Cao, C.; Li, C.; Chen, C.; Huang, Q. Sulfated modification, characterization, antioxidant and hypoglycemic activities of polysaccharides from Sargassum pallidum. Int. J. Biol. Macromol. 2018, 121, 407–414. [Google Scholar] [CrossRef]
- Wan, X.; Jin, X.; Wu, X.; Yang, X.; Lin, D.; Li, C.; Fu, Y.; Liu, Y.; Liu, X.; Lv, J.; et al. Structural characterisation and antitumor activity against non-small cell lung cancer of polysaccharides from Sanghuangporus vaninii. Carbohydr. Polym. 2021, 276, 118798. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Residue | Glycosidic Linkage | Methylation | w | Relative Molar Ratio (%) |
|---|---|---|---|---|
| A | t-Fucp | 1,5-di-O-acetyl-6-deoxy-2,3,4-tri-O-methyl fucitol | 293 | 11.520 |
| B | t-Manp | 1,5-di-O-acetyl-2,3,4,6-tetra-O-methyl mannitol | 323 | 13.887 |
| C | t-GalAp | 1,5-di-O-acetyl-2,3,4,6-tetra-O-methyl galactitol | 323 | 3.102 |
| D | 4-Xylp | 1,4,5-tri-O-acetyl-2,3-di-O-methyl xylitol | 307 | 5.600 |
| E | 2-Galp | 1,2,5-tri-O-acetyl-3,4,6-tri-O-methyl galactitol | 351 | 3.333 |
| F | 4-Manp | 1,4,5-tri-O-acetyl-2,3,6-tri-O-methyl mannitol | 351 | 4.483 |
| G | 6-Galp | 1,5,6-tri-O-acetyl-2,3,4-tri-O-methyl galactitol | 351 | 11.942 |
| H | 4-Galp | 1,4,5-tri-O-acetyl-2,3,6-tri-O-methyl galactitol | 351 | 3.294 |
| I | 4-Fucp | 1,4,5-tri-O-acetyl-6-deoxy-2,3-di-O-methyl fucitol | 351 | 31.852 |
| J | 3,4-GlcAp | 1,3,4,5-tetra-O-acetyl-2,6-di-O-methyl glucitol | 379 | 10.987 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, S.; Chen, X.; Chen, Y.; You, L.; Hileuskaya, K. UV/H2O2-Degraded Polysaccharides from Sargassum fusiforme: Purification, Structural Properties, and Anti-Inflammatory Activity. Mar. Drugs 2023, 21, 561. https://doi.org/10.3390/md21110561
Chang S, Chen X, Chen Y, You L, Hileuskaya K. UV/H2O2-Degraded Polysaccharides from Sargassum fusiforme: Purification, Structural Properties, and Anti-Inflammatory Activity. Marine Drugs. 2023; 21(11):561. https://doi.org/10.3390/md21110561
Chicago/Turabian StyleChang, Shiyuan, Xiaoyong Chen, Yifan Chen, Lijun You, and Kseniya Hileuskaya. 2023. "UV/H2O2-Degraded Polysaccharides from Sargassum fusiforme: Purification, Structural Properties, and Anti-Inflammatory Activity" Marine Drugs 21, no. 11: 561. https://doi.org/10.3390/md21110561
APA StyleChang, S., Chen, X., Chen, Y., You, L., & Hileuskaya, K. (2023). UV/H2O2-Degraded Polysaccharides from Sargassum fusiforme: Purification, Structural Properties, and Anti-Inflammatory Activity. Marine Drugs, 21(11), 561. https://doi.org/10.3390/md21110561