
In Vitro Interactions between Okadaic Acid and Rat Gut Microbiome


Abstract
:
1. Introduction
2. Results
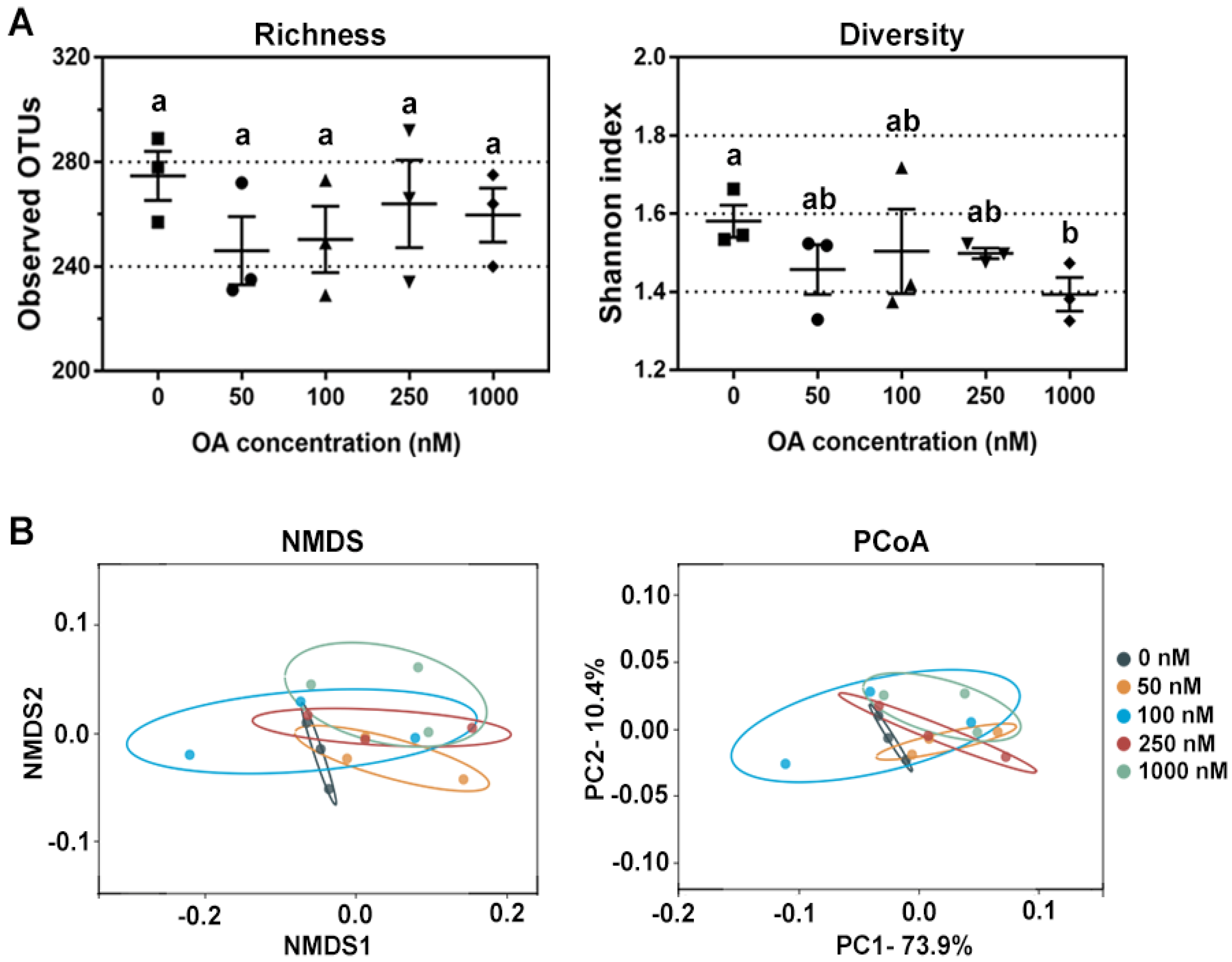
2.1. Effects of OA on Bacterial Diversity In Vitro
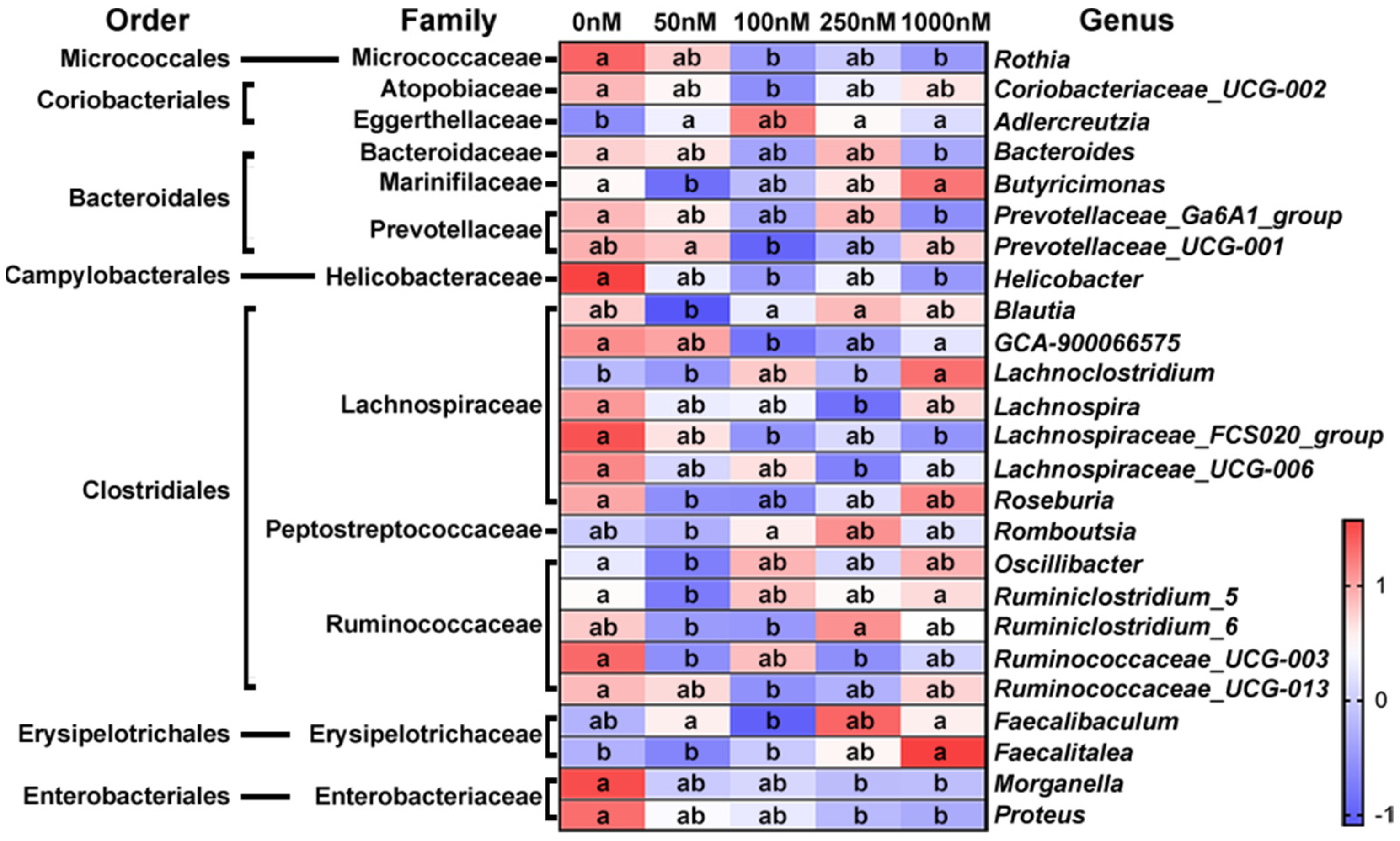
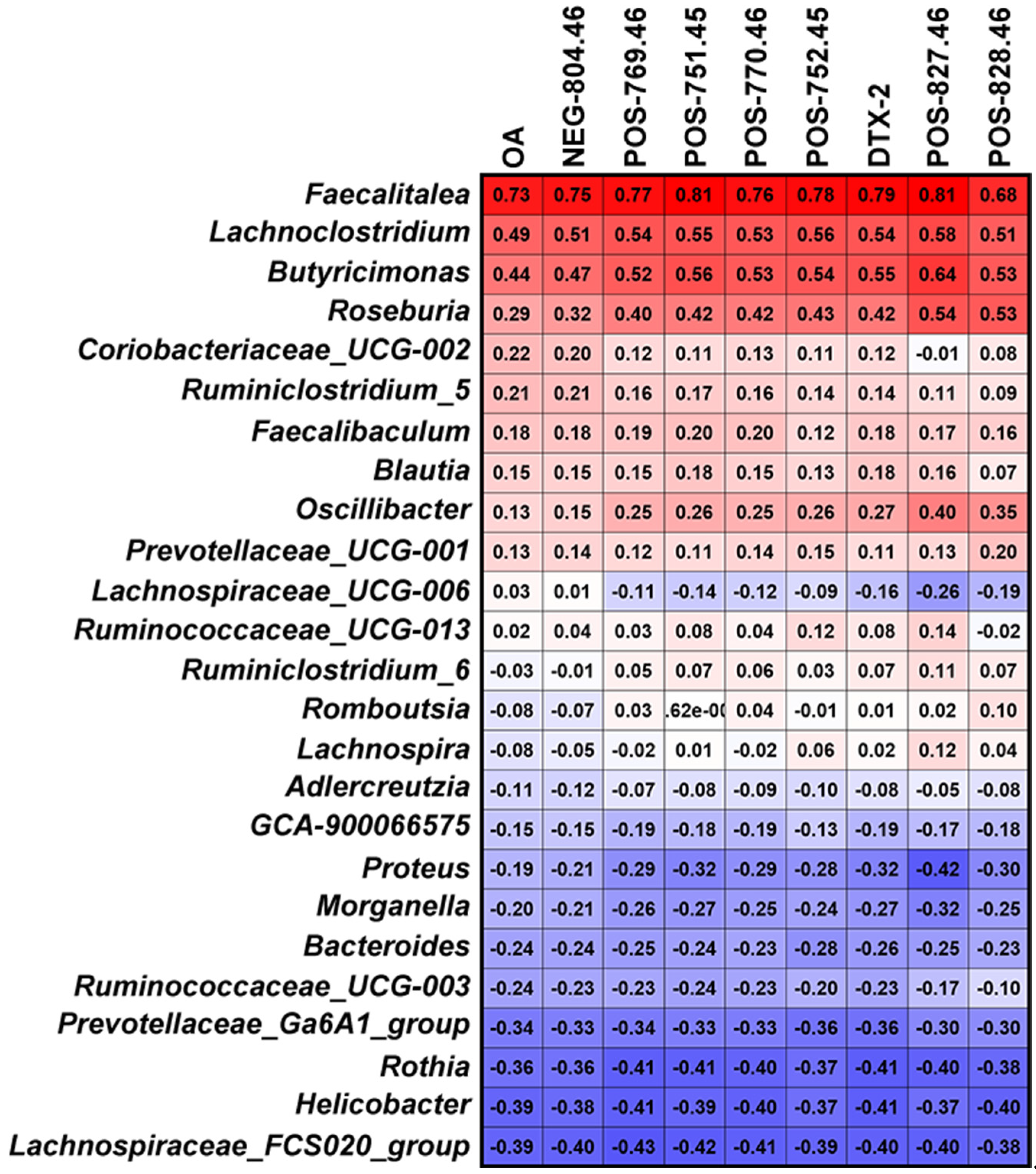
2.2. Changes in Bacterial Genera after OA Exposure
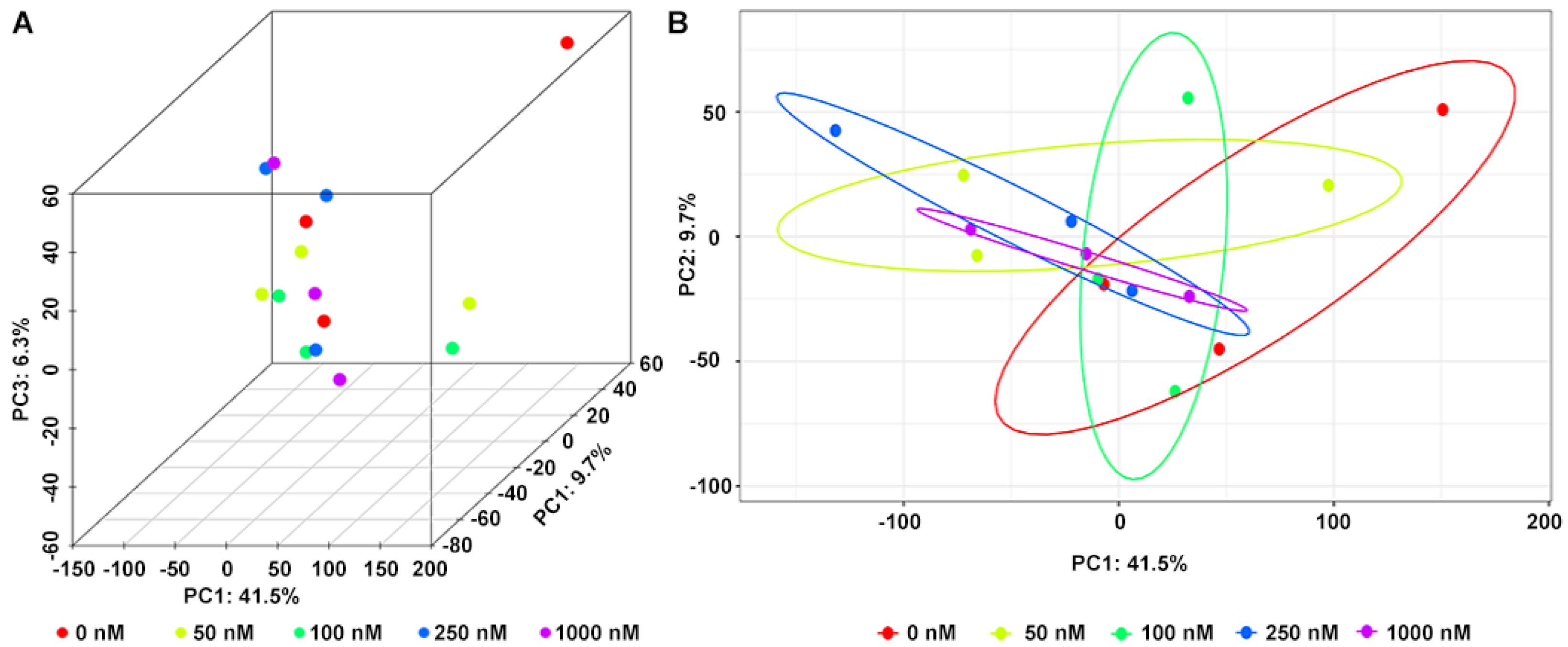
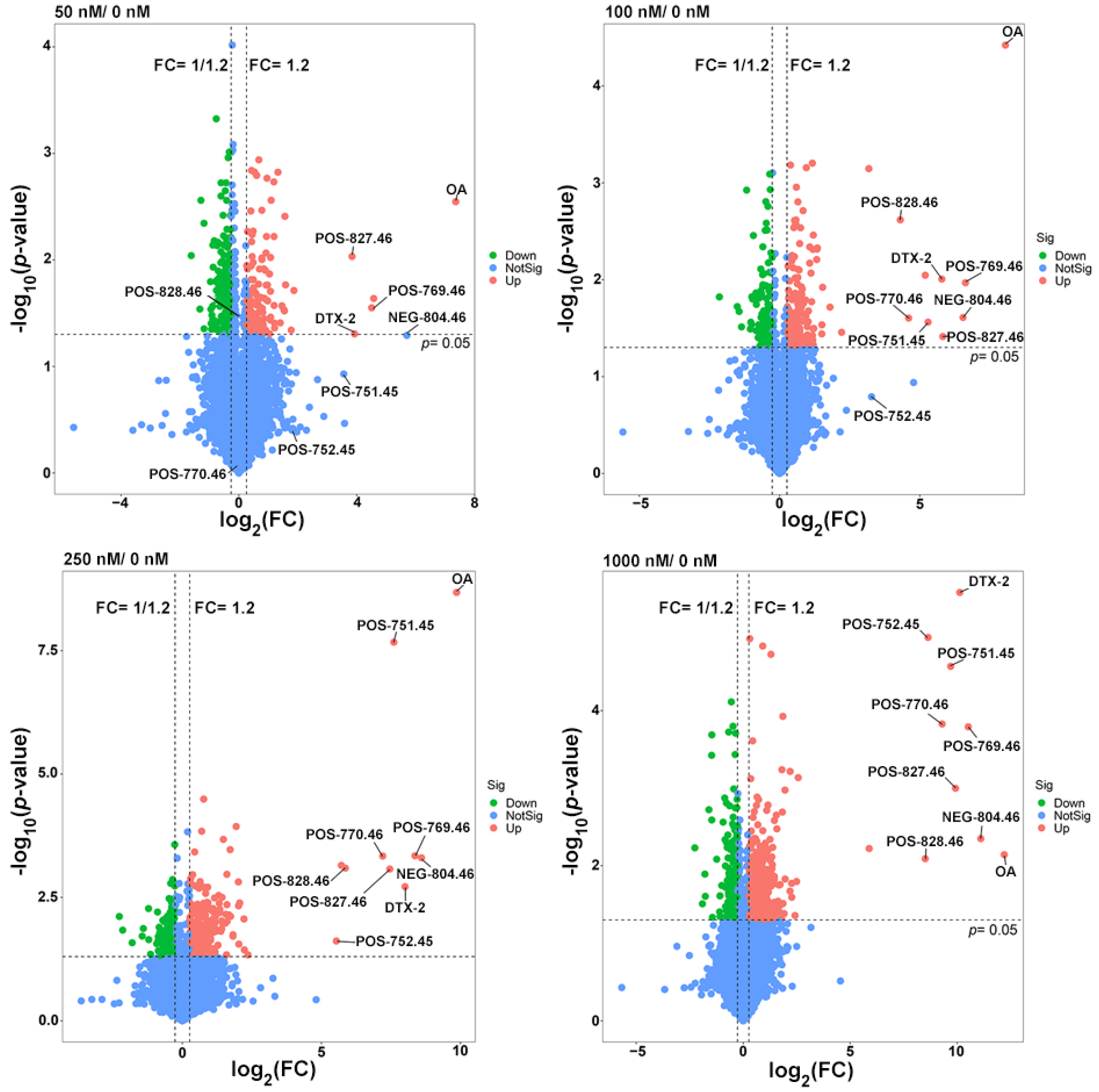
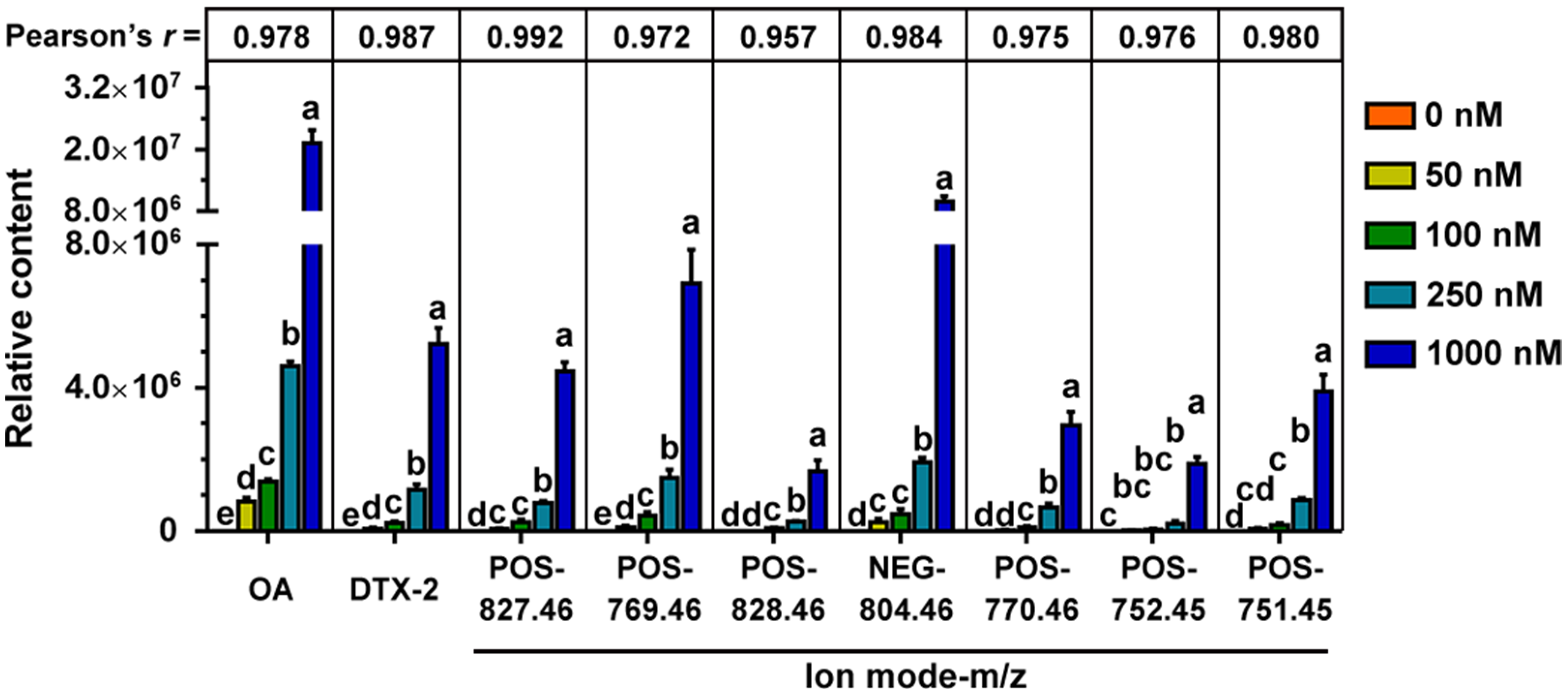
2.3. Metabolites of OA in the Fermentation System
3. Discussion
4. Materials and Methods
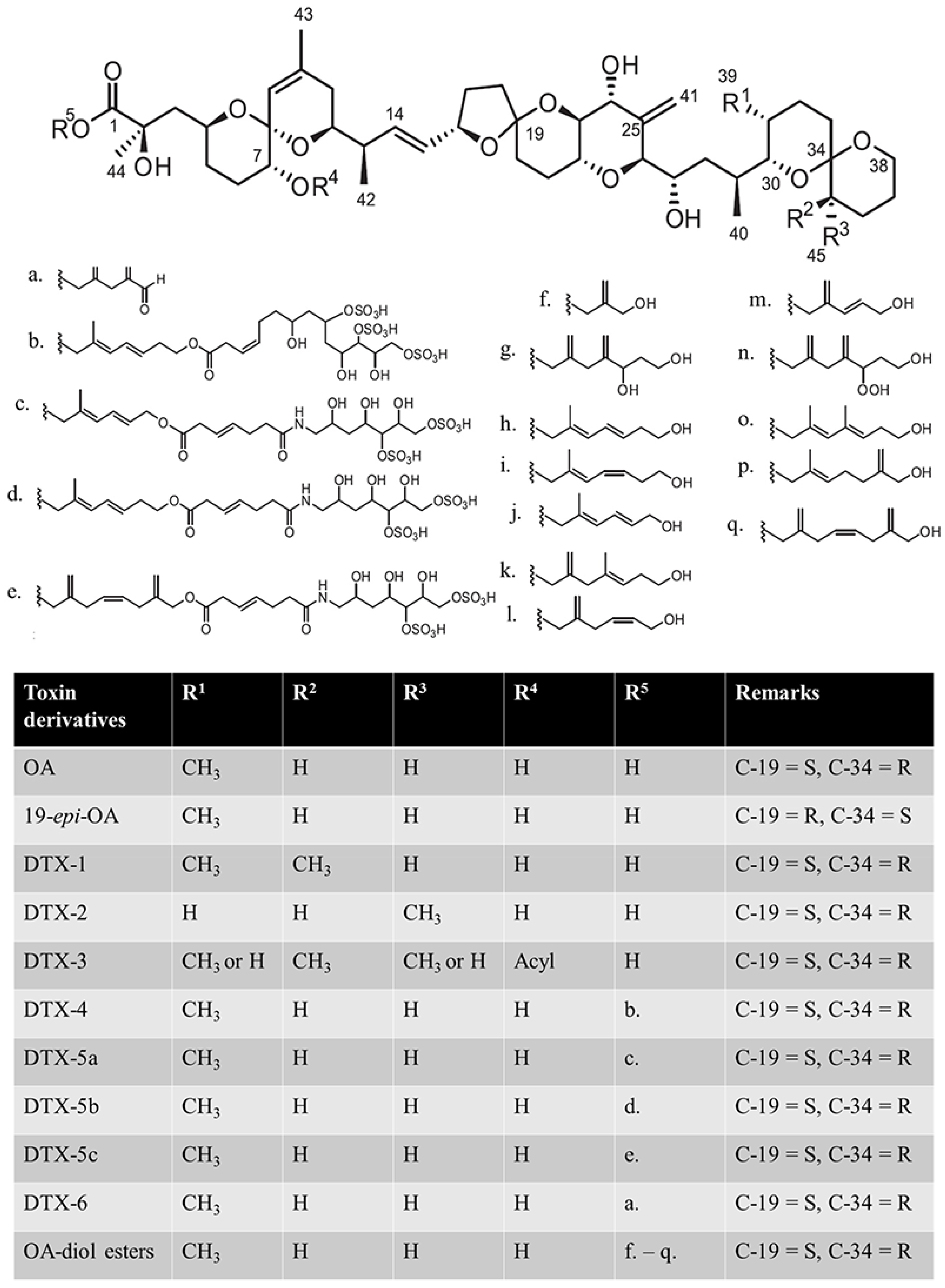
4.1. Material
4.2. Collection of Gut Microbiota
4.3. Fecal Microbial Culture Fermentation
4.4. Structure Analysis of Bacterial Community
4.5. Extraction of Metabolites from Fermentation System
4.6. LC-MS/MS Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- James, K.J.; Carey, B.; O’Halloran, J.; van Pelt, F.N.; Skrabakova, Z. Shellfish toxicity: Human health implications of marine algal toxins. Epidemiol. Infect. 2010, 138, 927–940. [Google Scholar] [PubMed]
- Chen, J.; Han, T.; Li, X.; He, X.; Wang, Y.; Chen, F.; Song, X.; Zhou, D.; Wang, X. Occurrence and distribution of marine natural organic pollutants: Lipophilic marine algal toxins in the Yellow Sea and the Bohai Sea, China. Sci. Total Environ. 2018, 612, 931–939. [Google Scholar] [CrossRef] [PubMed]
- Lü, Y.; Zhou, Z.; Chen, S.; Zhang, X. Progress on the metabolic rules and detection methods for okadaic acid related toxins in biological samples. Chin. J. Chromatogr. 2020, 38, 621–626. [Google Scholar]
- Munday, R. Is protein phosphatase inhibition responsible for the toxic effects of okadaic acid in animals? Toxins 2013, 5, 267–285. [Google Scholar] [CrossRef] [PubMed]
- Manerio, E.; Rodas, V.L.; Costas, E.; Hernandez, J.M. Shellfish consumption: A major risk factor for colorectal cancer. Med. Hypotheses 2008, 70, 409–412. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Rodas, V.; Maneiro, E.; Martínez, J.; Navarro, M.; Costas, E. Harmful algal blooms, red tides and human health: Diarrhetic shellfish poisoning and colorectal cancer. An. Real Acad. Nac. Farm. 2006, 72, 391. [Google Scholar]
- Lee, T.C.; Fong, F.L.; Ho, K.C.; Lee, F.W. The mechanism of diarrhetic shellfish poisoning toxin production in Prorocentrum spp.: Physiological and molecular perspectives. Toxins 2016, 8, 272. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). Marine biotoxins in shellfish-okadaic acid and analogues-scientific opinion of the panel on contaminants in the food chain. EFSA J. 2008, 6, 589. [Google Scholar] [CrossRef]
- Yasumoto, T.; Murata, M.; Oshima, Y.; Sano, M.; Matsumoto, G.; Clardy, J. Diarrhetic shellfish toxins. Tetrahedron 1985, 41, 1019–1025. [Google Scholar] [CrossRef]
- GARCêA, C.; Truan, D.; Lagos, M.; Santelices, J.P.; DêAZ, J.C.; Lagos, N. Metabolic transformation of dinophysistoxin-3 into dinophysistoxin-1 causes human intoxication by consumption of O-acyl-derivatives dinophysistoxins contaminated shellfish. J. Toxicol. Sci. 2005, 30, 287–296. [Google Scholar] [CrossRef]
- Botana, L.; Hess, P.; Munday, R.; Nathalie, A.; DeGrasse, S.; Feeley, M.; Suzuki, T.; Van den Berg, M.; Fattori, V.; Gamarro, E.; et al. Derivation of toxicity equivalency factors for marine biotoxins associated with bivalve molluscs. Trends Food Sci. Technol. 2017, 59, 15–24. [Google Scholar] [CrossRef]
- Aune, T.; Larsen, S.; Aasen, J.A.; Rehmann, N.; Satake, M.; Hess, P. Relative toxicity of dinophysistoxin-2 (DTX-2) compared with okadaic acid, based on acute intraperitoneal toxicity in mice. Toxicon 2007, 49, 1–7. [Google Scholar] [CrossRef]
- Tubaro, A.; Sosa, S.; Bornacin, A.; Jungerford, J. Pharmacology and toxicology of diarrheic shellfish toxins. In Seafood and Freshwater Toxins: Pharmacology, Physiology and Detection, 2nd ed.; Botana, L.M., Ed.; CRC Press: Boca Raton, FL, USA, 2008; pp. 229–253. [Google Scholar]
- Abal, P.; Louzao, M.C.; Suzuki, T.; Watanabe, R.; Vilariño, N.; Carrera, C.; Botana, A.M.; Vieytes, M.R.; Botana, L.M. Toxic action reevaluation of Okadaic Acid, Dinophysistoxin-1 and Dinophysistoxin-2: Toxicity Equivalency Factors based on the oral toxicity study. Cell. Physiol. Biochem. 2018, 49, 743–757. [Google Scholar] [CrossRef] [PubMed]
- Abal, P.; Louzao, M.C.; Cifuentes, J.M.; Vilarino, N.; Rodriguez, I.; Alfonso, A.; Vieytes, M.R.; Botana, L.M. Characterization of the dinophysistoxin-2 acute oral toxicity in mice to define the Toxicity Equivalency Factor. Food Chem. Toxicol. 2017, 102, 166–175. [Google Scholar] [CrossRef]
- Valdiglesias, V.; Prego-Faraldo, M.V.; Pasaro, E.; Mendez, J.; Laffon, B. Okadaic acid: More than a diarrheic toxin. Mar. Drugs 2013, 11, 4328–4349. [Google Scholar] [CrossRef]
- Dietrich, J.; Sommersdorf, C.; Gohlke, S.; Poetz, O.; Traenkle, B.; Rothbauer, U.; Hessel-Pras, S.; Lampen, A.; Braeuning, A. Okadaic acid activates Wnt/beta-catenin-signaling in human HepaRG cells. Arch. Toxicol. 2019, 93, 1927–1939. [Google Scholar] [CrossRef] [PubMed]
- Valdiglesias, V.; Laffon, B.; Pasaro, E.; Mendez, J. Okadaic acid induces morphological changes, apoptosis and cell cycle alterations in different human cell types. J. Environ. Monit. 2011, 13, 1831–1840. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Chen, B.; Xu, W.F.; Liu, R.F.; Yang, J.; Yu, C.X. Effects of PTEN inhibition on regulation of tau phosphorylation in an okadaic acid-induced neurodegeneration model. Int. J. Dev. Neurosci. 2012, 30, 411–419. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, C.; Hu, M.; Pan, J.; Chen, J.; Duan, P.; Zhai, T.; Ding, J.; Xu, C. Effects of ginkgolide A on okadaic acid-induced tau hyperphosphorylation and the PI3K-Akt signaling pathway in N2a cells. Planta Med. 2012, 78, 1337–1341. [Google Scholar] [CrossRef]
- Espina, B.; Louzao, M.C.; Cagide, E.; Alfonso, A.; Vieytes, M.R.; Yasumoto, T.; Botana, L.M. The methyl ester of okadaic acid is more potent than okadaic acid in disrupting the actin cytoskeleton and metabolism of primary cultured hepatocytes. Br. J. Pharmacol. 2010, 159, 337–344. [Google Scholar] [CrossRef]
- Louzao, M.C.; Fernandez, D.A.; Abal, P.; Fraga, M.; Vilarino, N.; Vieytes, M.R.; Botana, L.M. Diarrhetic effect of okadaic acid could be related with its neuronal action: Changes in neuropeptide Y. Toxicol. Lett. 2015, 237, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Guo, F.; Crain, S.; Quilliam, M.A.; Wang, X.; Rein, K.S. The structures of three metabolites of the algal hepatotoxin okadaic acid produced by oxidation with human cytochrome P450. Bioorg. Med. Chem. 2012, 20, 3742–3745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolrep, F.; Rein, K.; Lampen, A.; Hessel-Pras, S. Metabolism of okadaic acid by NADPH-dependent enzymes present in human or rat liver S9 fractions results in different toxic effects. Toxicol. In Vitro 2017, 42, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lu, Y.; Jiao, Y.H.; Li, D.W.; Li, H.Y.; Yang, W.D. Multi-omics analysis reveals metabolism of okadaic acid in gut lumen of rat. Arch. Toxicol. 2022, 96, 831–843. [Google Scholar] [CrossRef] [PubMed]
- Collins, S.L.; Patterson, A.D. The gut microbiome: An orchestrator of xenobiotic metabolism. Acta Pharm. Sin. B 2020, 10, 19–32. [Google Scholar] [CrossRef]
- Dethlefsen, L.; Relman, D.A. Incomplete recovery and individualized responses of the human distal gut microbiota to repeated antibiotic perturbation. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. S1), 4554–4561. [Google Scholar] [CrossRef]
- Feng, R.; Shou, J.W.; Zhao, Z.X.; He, C.Y.; Ma, C.; Huang, M.; Fu, J.; Tan, X.S.; Li, X.Y.; Wen, B.Y. Transforming berberine into its intestine-absorbable form by the gut microbiota. Sci. Rep. 2015, 5, 12155. [Google Scholar] [CrossRef]
- Ito, E.; Yasumoto, T.; Takai, A.; Imanishi, S.; Harada, K. Investigation of the distribution and excretion of okadaic acid in mice using immunostaining method. Toxicon 2002, 40, 159–165. [Google Scholar] [CrossRef]
- Aune, T.; Espenes, A.; Aasen, J.A.; Quilliam, M.A.; Hess, P.; Larsen, S. Study of possible combined toxic effects of azaspiracid-1 and okadaic acid in mice via the oral route. Toxicon 2012, 60, 895–906. [Google Scholar] [CrossRef] [PubMed]
- Emery, H.; Traves, W.; Rowley, A.F.; Coates, C.J. The diarrhetic shellfish-poisoning toxin, okadaic acid, provokes gastropathy, dysbiosis and susceptibility to bacterial infection in a non-rodent bioassay, Galleria mellonella. Arch. Toxicol. 2021, 95, 3361–3376. [Google Scholar] [CrossRef]
- Lagier, J.C.; Dubourg, G.; Million, M.; Cadoret, F.; Bilen, M.; Fenollar, F.; Levasseur, A.; Rolain, J.M.; Fournier, P.E.; Raoult, D. Culturing the human microbiota and culturomics. Nat. Rev. Microbiol. 2018, 16, 540–550. [Google Scholar] [CrossRef]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.R.; Nelson, K.E.; Relman, D.A. Diversity of the human intestinal microbial flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef] [Green Version]
- Martelli, F.; Cirlini, M.; Dellafiora, L.; Neviani, E.; Dall’Asta, C.; Bernini, V. Mitigation of marine toxins by interactions with bacteria: The case of okadaic acid and tetrodotoxin. Food Control 2022, 131, 108428. [Google Scholar] [CrossRef]
- Gotoh, A.; Nara, M.; Sugiyama, Y.; Sakanaka, M.; Yachi, H.; Kitakata, A.; Nakagawa, A.; Minami, H.; Okuda, S.; Katoh, T.; et al. Use of Gifu Anaerobic Medium for culturing 32 dominant species of human gut microbes and its evaluation based on short-chain fatty acids fermentation profiles. Biosci. Biotechnol. Biochem. 2017, 81, 2009–2017. [Google Scholar] [CrossRef]
- Reboreda, A.; Lago, J.; Chapela, M.J.; Vieites, J.M.; Botana, L.M.; Alfonso, A.; Cabado, A.G. Decrease of marine toxin content in bivalves by industrial processes. Toxicon 2010, 55, 235–243. [Google Scholar] [CrossRef]
- McCarron, P.; Kilcoyne, J.; Hess, P. Effects of cooking and heat treatment on concentration and tissue distribution of okadaic acid and dinophysistoxin-2 in mussels (Mytilus edulis). Toxicon 2008, 51, 1081–1089. [Google Scholar] [CrossRef]
- Matias, W.G.; Traore, A.; Creppy, E.E. Variations in the distribution of okadaic acid in organs and biological fluids of mice related to diarrhoeic syndrome. Hum. Exp. Toxicol. 1999, 18, 345–350. [Google Scholar] [CrossRef]
- Fu, L.L.; Zhao, X.Y.; Ji, L.D.; Xu, J. Okadaic acid (OA): Toxicity, detection and detoxification. Toxicon 2019, 160, 1–7. [Google Scholar] [CrossRef]
- Liu, Y.; Zhen, J.W.; Peng, X.C.; Li, H.Y.; Huang, L.; Li, D.W.; Liu, J.S.; Yang, W.D. Changes in colonic microbiotas in rat after long-term exposure to low dose of okadaic acid. Chemosphere 2020, 254, 126874. [Google Scholar] [CrossRef]
- Ravindran, J.; Gupta, N.; Agrawal, M.; Bala Bhaskar, A.S.; Lakshmana Rao, P.V. Modulation of ROS/MAPK signaling pathways by okadaic acid leads to cell death via, mitochondrial mediated caspase-dependent mechanism. Apoptosis 2011, 16, 145–161. [Google Scholar] [CrossRef]
- Dietrich, J.; Grass, I.; Gunzel, D.; Herek, S.; Braeuning, A.; Lampen, A.; Hessel-Pras, S. The marine biotoxin okadaic acid affects intestinal tight junction proteins in human intestinal cells. Toxicol. In Vitro 2019, 58, 150–160. [Google Scholar] [CrossRef]
- Ferron, P.J.; Hogeveen, K.; Fessard, V.; Le Hegarat, L. Comparative analysis of the cytotoxic effects of okadaic acid-group toxins on human intestinal cell lines. Mar. Drugs 2014, 12, 4616–4634. [Google Scholar] [CrossRef] [Green Version]
- Idle, J.R.; Gonzalez, F.J. Metabolomics. Cell Metab. 2007, 6, 348–351. [Google Scholar] [CrossRef]
- Chaleckis, R.; Meister, I.; Zhang, P.; Wheelock, C.E. Challenges, progress and promises of metabolite annotation for LC–MS-based metabolomics. Curr. Opin. Biotechnol. 2019, 55, 44–50. [Google Scholar] [CrossRef]
- De Maesschalck, C.; Van Immerseel, F.; Eeckhaut, V.; De Baere, S.; Cnockaert, M.; Croubels, S.; Haesebrouck, F.; Ducatelle, R.; Vandamme, P. Faecalicoccus acidiformans gen. nov., sp. nov., isolated from the chicken caecum, and reclassification of Streptococcus pleomorphus (Barnes et al. 1977), Eubacterium biforme (Eggerth 1935) and Eubacterium cylindroides (Cato et al. 1974) as Faecalicoccus pleomorphus comb. nov., Holdemanella biformis gen. nov., comb. nov. and Faecalitalea cylindroides gen. nov., comb. nov., respectively, within the family Erysipelotrichaceae. Int. J. Syst. Evol. Micr. 2014, 64, 3877–3884. [Google Scholar]
- Ma, Q.T.; Li, Y.Q.; Wang, J.K.; Li, P.F.; Duan, Y.H.; Dai, H.Y.; An, Y.C.; Cheng, L.; Wang, T.S.; Wang, C.G.; et al. Investigation of gut microbiome changes in type 1 diabetic mellitus rats based on high-throughput sequencing. Biomed. Pharmacother. 2020, 124, 109873. [Google Scholar] [CrossRef]
- Chen, G.; Xie, M.; Wan, P.; Chen, D.; Ye, H.; Chen, L.; Zeng, X.; Liu, Z. Digestion under saliva, simulated gastric and small intestinal conditions and fermentation in vitro by human intestinal microbiota of polysaccharides from Fuzhuan brick tea. Food Chem. 2018, 244, 331–339. [Google Scholar] [CrossRef]
- Huang, W.; Yin, H.; Yang, Y.; Jin, L.; Lu, G.; Dang, Z. Influence of the co-exposure of microplastics and tetrabromobisphenol A on human gut: Simulation in vitro with human cell Caco-2 and gut microbiota. Sci. Total Environ. 2021, 778, 146264. [Google Scholar] [CrossRef]
- Dai, T.J.; Wen, D.H.; Bates, C.T.; Wu, L.W.; Guo, X.; Liu, S.; Su, Y.F.; Lei, J.S.; Zhou, J.Z.; Yang, Y.F. Nutrient supply controls the linkage between species abundance and ecological interactions in marine bacterial communities. Nat. Commun. 2022, 13, 175. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Magoc, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Wang, J.L.; Zhang, T.; Shen, X.T.; Liu, J.; Zhao, D.L.; Sun, Y.W.; Wang, L.; Liu, Y.L.; Gong, Y.X.; Liu, Y.X.; et al. Serum metabolomics for early diagnosis of esophageal squamous cell carcinoma by UHPLC-QTOF/MS. Metabolomics 2016, 12, 116. [Google Scholar] [CrossRef]
- Smith, C.A.; Want, E.J.; O’Maille, G.; Abagyan, R.; Siuzdak, G. XCMS: Processing mass spectrometry data for metabolite profiling using nonlinear peak alignment, matching, and identification. Anal. Chem. 2006, 78, 779–787. [Google Scholar] [CrossRef]
- Dunn, W.B.; Broadhurst, D.; Begley, P.; Begley, P.; Zelena, E.; Francis-McIntyre, S.; Anderson, N.; Brown, M.; Knowles, J.D.; Halsall, A.; et al. Procedures for large-scale metabolic profiling of serum and plasma using gas chromatography and liquid chromatography coupled to mass spectrometry. Nat. Protoc. 2011, 6, 1060–1083. [Google Scholar] [CrossRef]
- Li, L.; Lin, L.; Deng, J.; Lin, X.L.; Li, Y.M.; Xia, B.H. The therapeutic effects of Prunella vulgaris against fluoride-induced oxidative damage by using the metabolomics method. Environ. Toxicol. 2021, 36, 1802–1816. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dissimilarity Test (Compared with 0 nM) | Adonis | Amova | ||
|---|---|---|---|---|
| R2 | p-Value | Fs | p-Value | |
| 50 nM | 0.315 | 0.200 | 0.033 | 0.182 |
| 100 nM | 0.036 | 0.900 | 0.006 | 0.919 |
| 250 nM | 0.306 | 0.200 | 0.028 | 0.214 |
| 1000 nM | 0.303 | 0.100 | 0.027 | 0.099 |
| Ion Mode -m/z | MS2 Name | Average Peak Area | ||||
|---|---|---|---|---|---|---|
| 0 nM | 50 nM | 100 nM | 250 nM | 1000 nM | ||
| POS-847.4 | 772,914.5 | 1,354,450.7 | 1,245,177.5 | 1,238,153.1 | 1,361,723.0 | |
| POS-827.46 | 0.0 | 64,009.6 | 244,445.6 | 779,112.4 | 4,457,365.8 | |
| NEG-803.46 | OA | 0.0 | 817,356.1 | 1,384,218.9 | 4,597,209.1 | 21,190,896.5 |
| POS-787.47 | DTX-2 | 0.0 | 71,343.8 | 235,165.0 | 1,146,066.1 | 5,208,008.2 |
| POS-769.46 | 0.0 | 108,862.8 | 431,789.1 | 1,489,167.6 | 6,906,719.2 | |
| POS-665.5 | 214,218.8 | 559,356.9 | 637,709.5 | 730,160.9 | 806,568.0 | |
| POS-492.86 | 625,484.9 | 904,944.3 | 817,592.2 | 873,855.3 | 879,275.2 | |
| POS-429.14 | 26,078.9 | 650,092.3 | 1,050,385.8 | 1,424,048.2 | 1,468,577.4 | |
| POS-422.15 | 222,449.8 | 625,832.1 | 647,563.7 | 853,352.8 | 1,167,906.2 | |
| POS-351.12 | 259,230.5 | 720,817.2 | 681,464.2 | 845,456.2 | 642,511.6 | |
| NEG-343.25 | Streptidine 6-phosphate | 1,695,420.0 | 341,608.0 | 3,286,003.0 | 4,142,678.0 | 4,398,131.0 |
| POS-331.25 | 9,802,187.2 | 18,301,392.0 | 16,095,423.7 | 24,730,228.0 | 23,451,383.2 | |
| POS-302.23 | 364,045.6 | 764,114.8 | 685,071.7 | 594,568.3 | 635,510.6 | |
| POS-286.01 | 1,765,617.3 | 4,276,114.8 | 5,063,965.6 | 6,421,677.2 | 7,146,981.4 | |
| POS-239.12 | 3,614,630.5 | 6,147,948.6 | 6,369,886.7 | 7,900,067.9 | 8,214,998.3 | |
| POS-238.12 | 48,583,494.9 | 73,257,098.7 | 76,581,091.2 | 96,541,315.3 | 96,654,785.7 | |
| POS-220.03 | 834,014.0 | 2,285,900.4 | 1,906,606.8 | 1,964,003.2 | 1,862,868.0 | |
| POS-198.04 | 15,112,106.4 | 23,673,256.5 | 25,004,665.9 | 23,302,260.8 | 26,790,744.5 | |
| NEG-190.11 | 13,633,947.1 | 22,980,736.4 | 24,228,301.7 | 26,926,848.1 | 28,425,445.8 | |
| NEG-184.07 | Phosphorylcholine | 18,489,454.0 | 18,368,797.2 | 16,243,717.3 | 16,906,562.7 | 15,754,843.3 |
| NEG-182.03 | 35,467,400.7 | 22,823,567.8 | 26,780,550.4 | 23,895,222.5 | 26,635,669.7 | |
| NEG-176.1 | 33,279,171.4 | 51,094,329.5 | 53,722,869.8 | 65,596,077.6 | 79,499,673.8 | |
| NEG-176.09 | 2-minoheptanedioic acid | 2,666,927.2 | 5,726,041.4 | 5,114,338.2 | 5,593,144.0 | 6,024,575.6 |
| NEG-172.01 | 3,495,412.5 | 6,126,153.9 | 6,731,904.1 | 10,033,830.4 | 10,796,430.5 | |
| NEG-161.58 | 47,066.5 | 193,948.2 | 241,584.7 | 244,839.5 | 308,397.3 | |
| NEG-159.08 | 6,661,437.9 | 6,275,405.0 | 6,012,287.2 | 5,536,234.3 | 5,565,532.8 | |
| NEG-154.05 | 27,805,547.6 | 43,294,413.2 | 43,421,409.9 | 43,761,555.1 | 48,446,617.0 | |
| NEG-140.03 | 11,521,804.8 | 23,546,688.8 | 24,415,464.2 | 26,846,274.0 | 26,580,788.2 | |
| NEG-128.02 | 48,306,703.7 | 106,620,905.9 | 121,809,279.9 | 151,223,200.9 | 164,878,532.6 | |
| NEG-88.05 | 18,047,252.3 | 29,484,126.5 | 31,837,001.7 | 38,950,682.7 | 37,983,269.3 | |
| NEG-83.06 | 22,865,390.9 | 9,642,657.3 | 10,409,259.5 | 10,924,049.3 | 6,359,238.0 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Xu, S.; Cai, Q.; Li, D.; Li, H.; Yang, W. In Vitro Interactions between Okadaic Acid and Rat Gut Microbiome. Mar. Drugs 2022, 20, 556. https://doi.org/10.3390/md20090556
Liu Y, Xu S, Cai Q, Li D, Li H, Yang W. In Vitro Interactions between Okadaic Acid and Rat Gut Microbiome. Marine Drugs. 2022; 20(9):556. https://doi.org/10.3390/md20090556
Chicago/Turabian StyleLiu, Yang, Siyuan Xu, Qiudie Cai, Dawei Li, Hongye Li, and Weidong Yang. 2022. "In Vitro Interactions between Okadaic Acid and Rat Gut Microbiome" Marine Drugs 20, no. 9: 556. https://doi.org/10.3390/md20090556
APA StyleLiu, Y., Xu, S., Cai, Q., Li, D., Li, H., & Yang, W. (2022). In Vitro Interactions between Okadaic Acid and Rat Gut Microbiome. Marine Drugs, 20(9), 556. https://doi.org/10.3390/md20090556