
Identification of PKS-NRPS Hybrid Metabolites in Marine-Derived Penicillium oxalicum

 , ,
, ,  ,
,  ,
, 
Abstract
:1. Introduction
2. Results
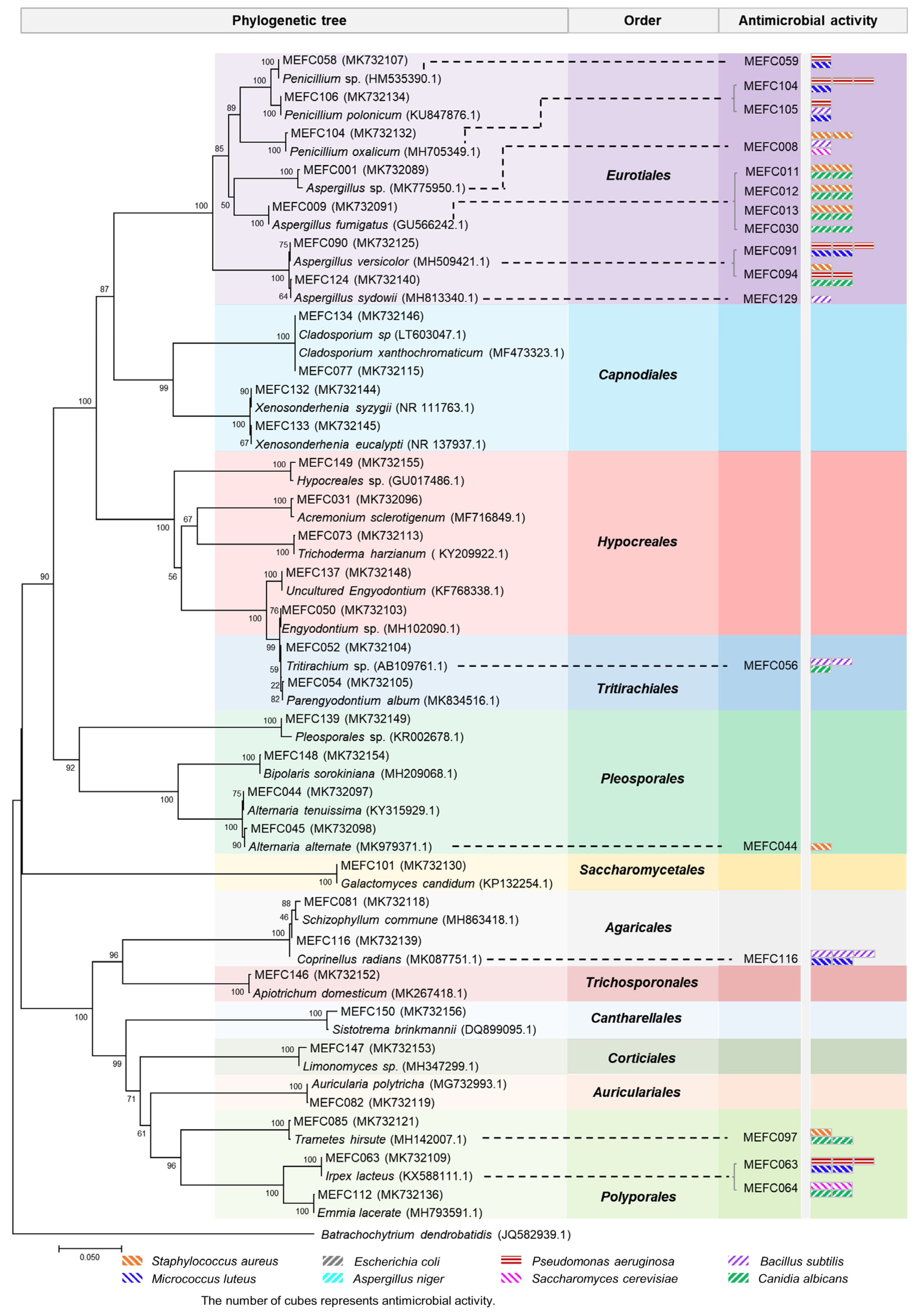
2.1. Isolation and Identification of Marine-Derived Fungi
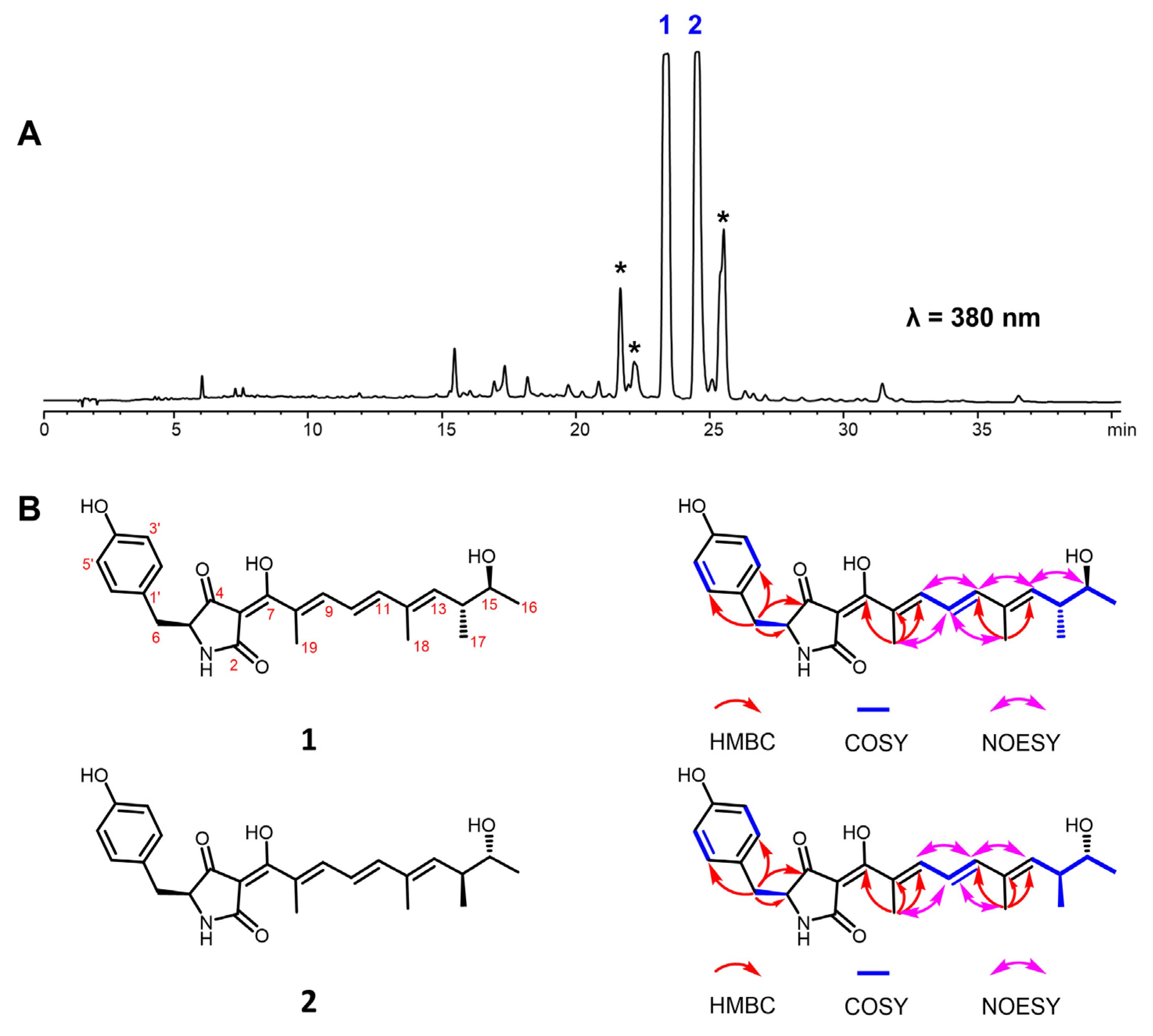
2.2. Structural Elucidation of Oxopyrrolidine A (1) and B (2)
2.3. Identification of the Biosynthetic Gene Cluster
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Isolation of Marine-Derived Fungi
3.3. Screening for Bioactive Marine-Derived Fungal Strains
3.4. Culture Conditions
3.5. Extraction and Isolation
3.6. Mosher’s Reactions of Oxopyrrolidine A (1) and B (2)
3.7. Computational Details
3.8. Antimicrobial Activity Analysis against P. aeruginosa
3.9. Protoplast Preparation and Transformation
3.10. Genome Sequencing
3.11. Construction of P. oxalicum Mutant Strains
3.12. HPLC and LC-HRMS Analysis of the Metabolites from P. oxalicum Mutant Strains
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lima, L.M.; da Silva, B.N.M.; Barbosa, G.; Barreiro, E.J. β-lactam antibiotics: An overview from a medicinal chemistry perspective. Eur. J. Med. Chem. 2020, 208, 112829. [Google Scholar] [CrossRef] [PubMed]
- Cacho, R.A.; Jiang, W.; Chooi, Y.H.; Walsh, C.T.; Tang, Y. Identification and characterization of the Echinocandin B biosynthetic gene cluster from Emericella rugulosa NRRL 11440. J. Am. Chem. Soc. 2012, 134, 16781–16790. [Google Scholar] [CrossRef]
- Kennedy, J.; Auclair, K.; Kendrew, S.G.; Park, C.; Vederas, J.C.; Hutchinson, C.R. Modulation of polyketide synthase activity by accessory proteins during lovastatin biosynthesis. Science 1999, 284, 1368–1372. [Google Scholar] [CrossRef] [PubMed]
- Survase, S.A.; Kagliwal, L.D.; Annapure, U.S.; Singhal, R.S. Cyclosporin A--A review on fermentative production, downstream processing and pharmacological applications. Biotechnol. Adv. 2011, 29, 418–435. [Google Scholar] [CrossRef] [PubMed]
- Rateb, M.E.; Ebel, R. Secondary metabolites of fungi from marine habitats. Nat. Prod. Rep. 2011, 28, 290–344. [Google Scholar] [CrossRef] [PubMed]
- Segeth, M.P.; Bonnefoy, A.; Broenstrup, M.; Knauf, M.; Schummer, D.; Toti, L.; Vertesy, L.; Wetzel-Raynal, M.C.; Wink, J.; Seibert, G. Coniosetin, a novel tetramic acid antibiotic from Coniochaeta ellipsoidea DSM 13856. J. Antibiot. 2003, 56, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Sata, N.U.; Wada, S.I.; Matsunaga, S.; Watabe, S.; van Soest, R.W.; Fusetani, N. Rubrosides A- H, new bioactive tetramic acid glycosides from the marine sponge Siliquariaspongia japonica. J. Org. Chem. 1999, 64, 2331–2339. [Google Scholar] [CrossRef]
- Holzapfel, C.W. The isolation and structure of cyclopiazonic acid, a toxic metabolite of Penicillium cyclopium westling. Tetrahedron 1968, 24, 2101–2119. [Google Scholar] [CrossRef]
- Jiang, M.H.; Chen, S.H.; Li, J.; Liu, L. The biological and chemical diversity of tetramic acid compounds from marine-derived microorganisms. Mar. Drugs 2020, 18, 114. [Google Scholar] [CrossRef] [PubMed]
- Kemami Wangun, H.V.; Hertweck, C. Epicoccarines A, B and epipyridone: Tetramic acids and pyridone alkaloids from an Epicoccum sp. associated with the tree fungus Pholiota squarrosa. Org. Biomol. Chem. 2007, 5, 1702–1705. [Google Scholar] [CrossRef] [PubMed]
- Matsumori, N.; Kaneno, D.; Murata, M.; Nakamura, H.; Tachibana, K. Stereochemical determination of acyclic structures based on carbon-proton spin-coupling constants. a method of configuration analysis for natural products. J. Org. Chem. 1999, 64, 866–876. [Google Scholar] [CrossRef] [PubMed]
- Neuhaus, G.F.; Adpressa, D.A.; Bruhn, T.; Loesgen, S. Polyketides from marine-derived Aspergillus porosus: Challenges and opportunities for determining absolute configuration. J. Nat. Prod. 2019, 82, 2780–2789. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Cai, X.L.; Jung, M.E.; Tang, Y. Analysis of intact and dissected fungal polyketide synthase-nonribosomal peptide synthetase in vitro and in Saccharomyces cerevisiae. J. Am. Chem. Soc. 2010, 132, 13604–13607. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, S.; Schumann, J.; Scherlach, K.; Lange, C.; Brakhage, A.A.; Hertweck, C. Genomics-driven discovery of PKS-NRPS hybrid metabolites from Aspergillus nidulans. Nat. Chem. Biol. 2007, 3, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.M.; Li, J.W.H.; Choi, J.W.; Zhou, H.; Lee, K.K.M.; Moorthie, V.A.; Xie, X.; Kealey, J.T.; Da Silva, N.A.; Vederas, J.C.; et al. Complete reconstitution of a highly reducing iterative polyketide synthase. Science 2009, 326, 589–592. [Google Scholar] [CrossRef] [PubMed]
- Takino, T.; Kotani, A.; Ozaki, T.; Peng, W.Q.; Yu, J.; Guo, Y.; Mochizuki, S.; Akimitsu, K.; Hashimoto, M.; Ye, T.; et al. Biochemistry-guided prediction of the absolute configuration of fungal reduced polyketides. Angew. Chem. Int. Ed. 2021, 60, 23403–23411. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhao, S.; Chen, X.X.; Deng, Q.P.; Li, C.X.; Feng, J.X. Secretory overproduction of a raw starch-degrading glucoamylase in Penicillium oxalicum using strong promoter and signal peptide. Appl. Microbiol. Biotechnol. 2018, 102, 9291–9301. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.N.; Chen, M.; Li, J.J.; Lu, X.F. Establishing an efficient gene-targeting system in an itaconic-acid producing Aspergillus terreus strain. Biotechnol. Lett. 2016, 38, 1603–1610. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1 | 2 | |||||
|---|---|---|---|---|---|---|
| Pos. | δC, Type | δH (J in Hz) | HMBC | δC, Type | δH (J in Hz) | HMBC |
| 2 | 176.9 C | 177.1 C | ||||
| 3 | 99.1 C | 98.5 C | ||||
| 4 | 192.0 C | 192.3 C | ||||
| 5 | 61.3 CH | 4.03 (1H, br s) | 1′; 6′ | 61.5 CH | 4.02 (1H, s) | |
| 6 | 36.0 CH2 | 2.85 (2H, m) | 5; 1′; 2′; 6′ | 35.9 CH2 | 2.84 (2H, t, J = 4.5 Hz) | 4; 5; 1′; 2′; 6′ |
| 7 | 183.7 C | 182.7 C | ||||
| 8 | 128.2 C | 127.5 C | ||||
| 9 | 142.0 CH | 7.68 (1H, s) | 141.7 CH | 7.65 (1H, d, J = 6.1 Hz) | ||
| 10 | 122.1 CH | 6.53 (1H, dd, J = 14.9, 11.3 Hz) | 8; 9; 11; 12 | 121.7 CH | 6.54 (1H, m) | 11 |
| 11 | 146.6 CH | 6.64 (1H, m) | 9; 10; 12; 13; 18 | 146.5 CH | 6.64 (1H, m) | |
| 12 | 133.4 C | 133.7 C | ||||
| 13 | 142.7 CH | 5.73 (1H, d, J = 9.9 Hz) | 11; 14; 15; 17; 18; 19 | 142.1 CH | 5.79 (1H, d, J = 9.1 Hz) | 11; 14; 15; 17; 18; 19 |
| 14 | 40.8 CH | 2.45 (1H, m) | 12; 13; 15; 16; 17 | 39.6 CH | 2.54 (1H, m) | 12; 13; 15; 16; 17 |
| 15 | 70.0 CH | 3.45 (1H, m) | 13; 14; 16; 17 | 69.3 CH | 3.57 (1H, m) | 13; 14; 16; 17 |
| 16 | 21.6 CH3 | 1.02 (3H, d, J = 6.1 Hz) | 14; 15 | 20.6 CH3 | 1.01 (3H, d, J = 6.3 Hz) | 14; 15 |
| 17 | 16.6 CH3 | 0.97 (3H, d, J = 6.6 Hz) | 13; 14; 15 | 16.4 CH3 | 0.96 (3H, d, J = 6.8 Hz) | 13; 14; 15 |
| 18 | 12.7 CH3 | 1.83 (3H, s) | 11; 12; 13; 15; 17 | 12.4 CH3 | 1.83 (3H, s) | 11; 12; 13; 14; 15; 17 |
| 19 | 12.3 CH3 | 1.89 (3H, s) | 7; 8; 9; 10; 11 | 12.2 CH3 | 1.91 (3H, s) | 7; 8; 9 |
| 1′ | 125.8 C | 125.7 C | ||||
| 2′ | 130.8 CH | 6.93 (1H, d, J = 7.9 Hz) | 6; 3′; 4′; 5′; 6′ | 130.6 CH | 6.94 (2H, d, J = 8.0 Hz) | 6; 3′; 4′; 5′; 6′ |
| 3′ | 114.8 CH | 6.61 (1H, d, J = 7.9 Hz) | 1′; 4′; 5′ | 114.6 CH | 6.62 (1H, d, J = 8.0 Hz) | 1′; 4′; 5′ |
| 4′ | 156.0 C | 155.8 C | ||||
| 5′ | 114.8 CH | 6.61 (1H, d, J = 7.9 Hz) | 1′; 3′; 4′ | 114.6 CH | 6.62 (1H, d, J = 8.0 Hz) | 1′; 3′; 4′ |
| 6′ | 130.8 CH | 6.93 (1H, d, J = 7.9 Hz) | 6; 2′; 3′; 4′; 5′ | 130.6 CH | 6.94 (2H, d, J = 8.0 Hz) | 6; 2′; 3′; 4′; 5′ |
| 15-OH | 3.51 (1H, s) | 15 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Zhang, W.; Zhang, X.; Tang, S.; Men, P.; Xiong, M.; Li, Z.; Zhang, Y.; Huang, X.; Lu, X. Identification of PKS-NRPS Hybrid Metabolites in Marine-Derived Penicillium oxalicum. Mar. Drugs 2022, 20, 523. https://doi.org/10.3390/md20080523
Li H, Zhang W, Zhang X, Tang S, Men P, Xiong M, Li Z, Zhang Y, Huang X, Lu X. Identification of PKS-NRPS Hybrid Metabolites in Marine-Derived Penicillium oxalicum. Marine Drugs. 2022; 20(8):523. https://doi.org/10.3390/md20080523
Chicago/Turabian StyleLi, Hongcheng, Wei Zhang, Xuan Zhang, Shen Tang, Ping Men, Mengyi Xiong, Zhimin Li, Yongyu Zhang, Xuenian Huang, and Xuefeng Lu. 2022. "Identification of PKS-NRPS Hybrid Metabolites in Marine-Derived Penicillium oxalicum" Marine Drugs 20, no. 8: 523. https://doi.org/10.3390/md20080523
APA StyleLi, H., Zhang, W., Zhang, X., Tang, S., Men, P., Xiong, M., Li, Z., Zhang, Y., Huang, X., & Lu, X. (2022). Identification of PKS-NRPS Hybrid Metabolites in Marine-Derived Penicillium oxalicum. Marine Drugs, 20(8), 523. https://doi.org/10.3390/md20080523