
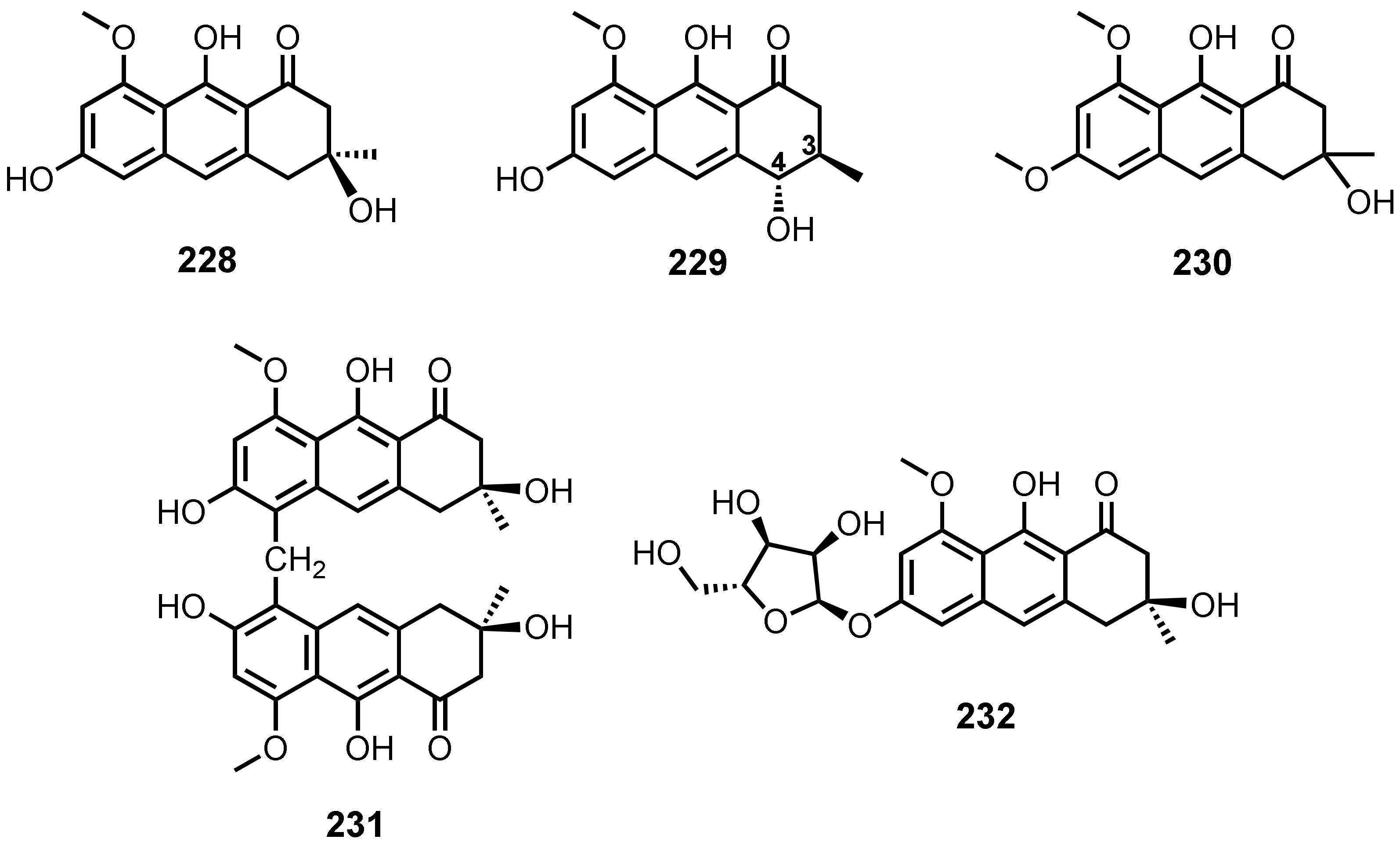
Anthraquinones and Their Analogues from Marine-Derived Fungi: Chemistry and Biological Activities





Abstract
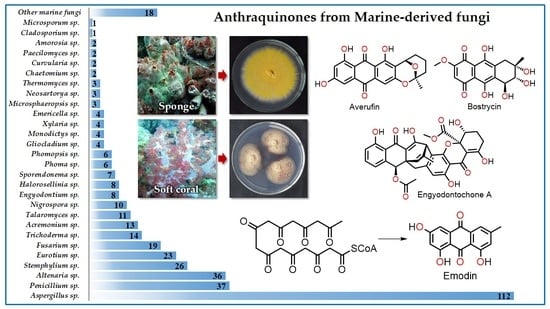
:
1. Introduction
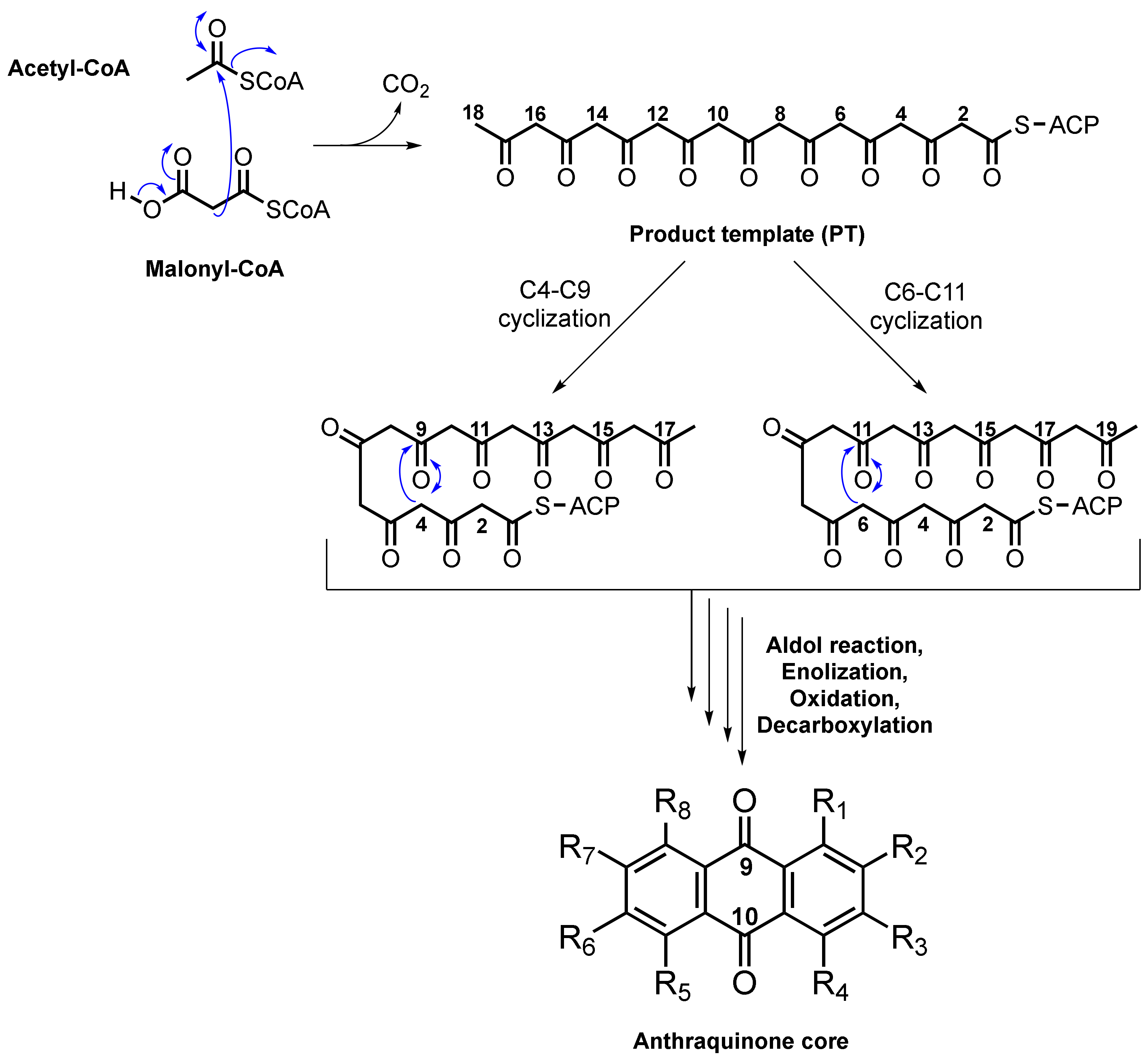
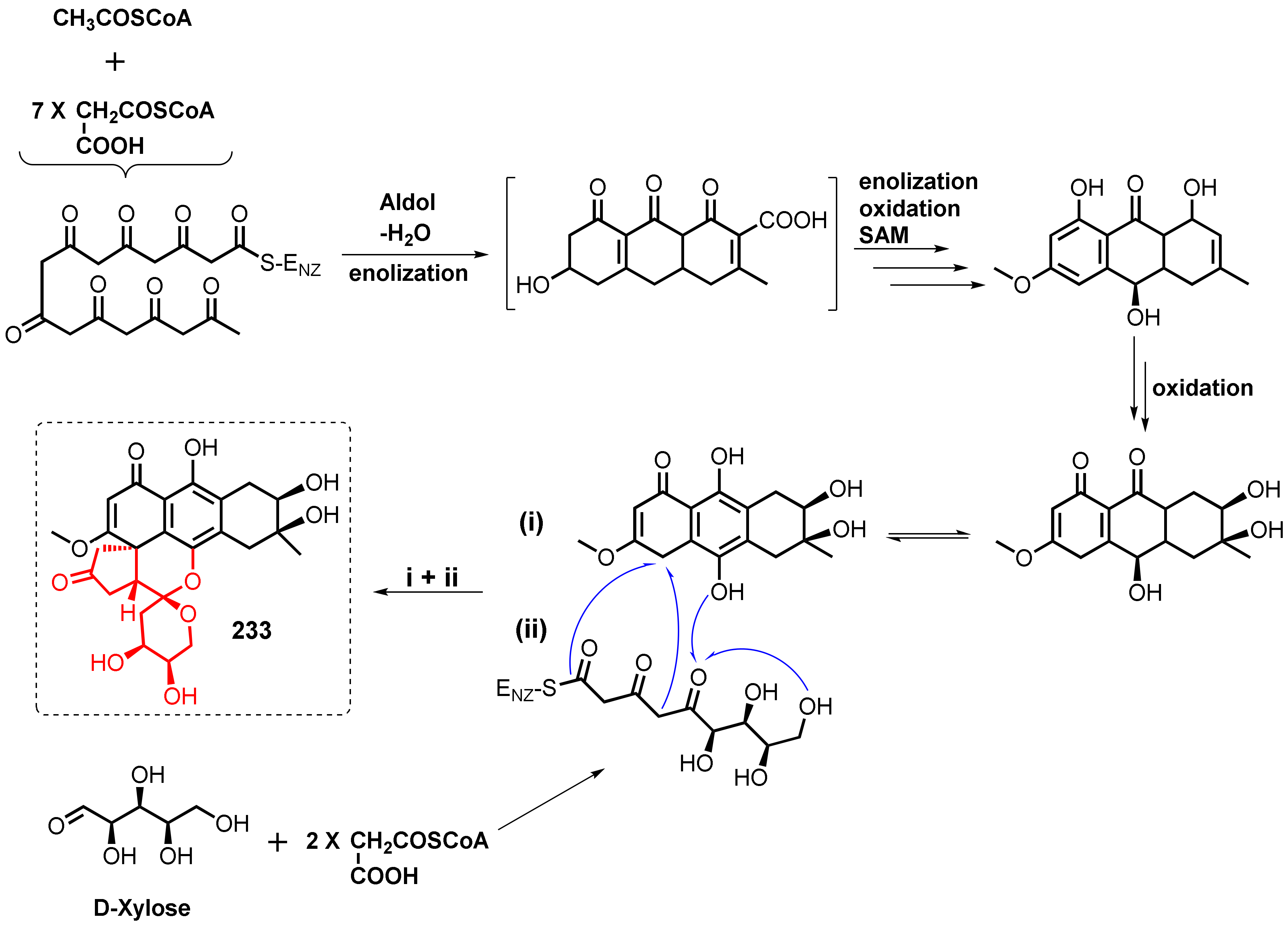
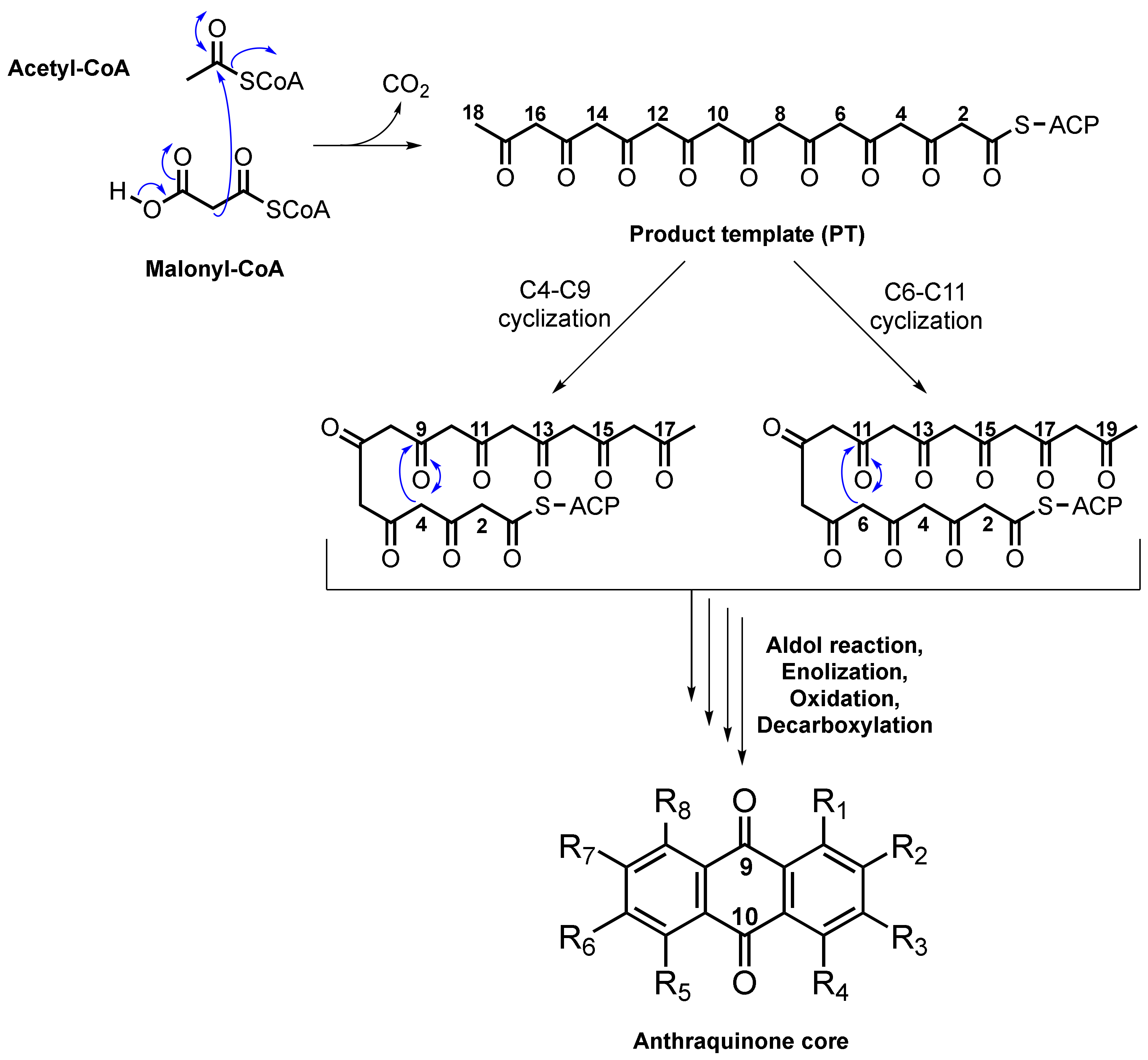
2. Biosynthesis of Anthraquinone Scaffold
3. Anthraquinoid Polyketides and Their Analogues from Marine-Derived Fungi
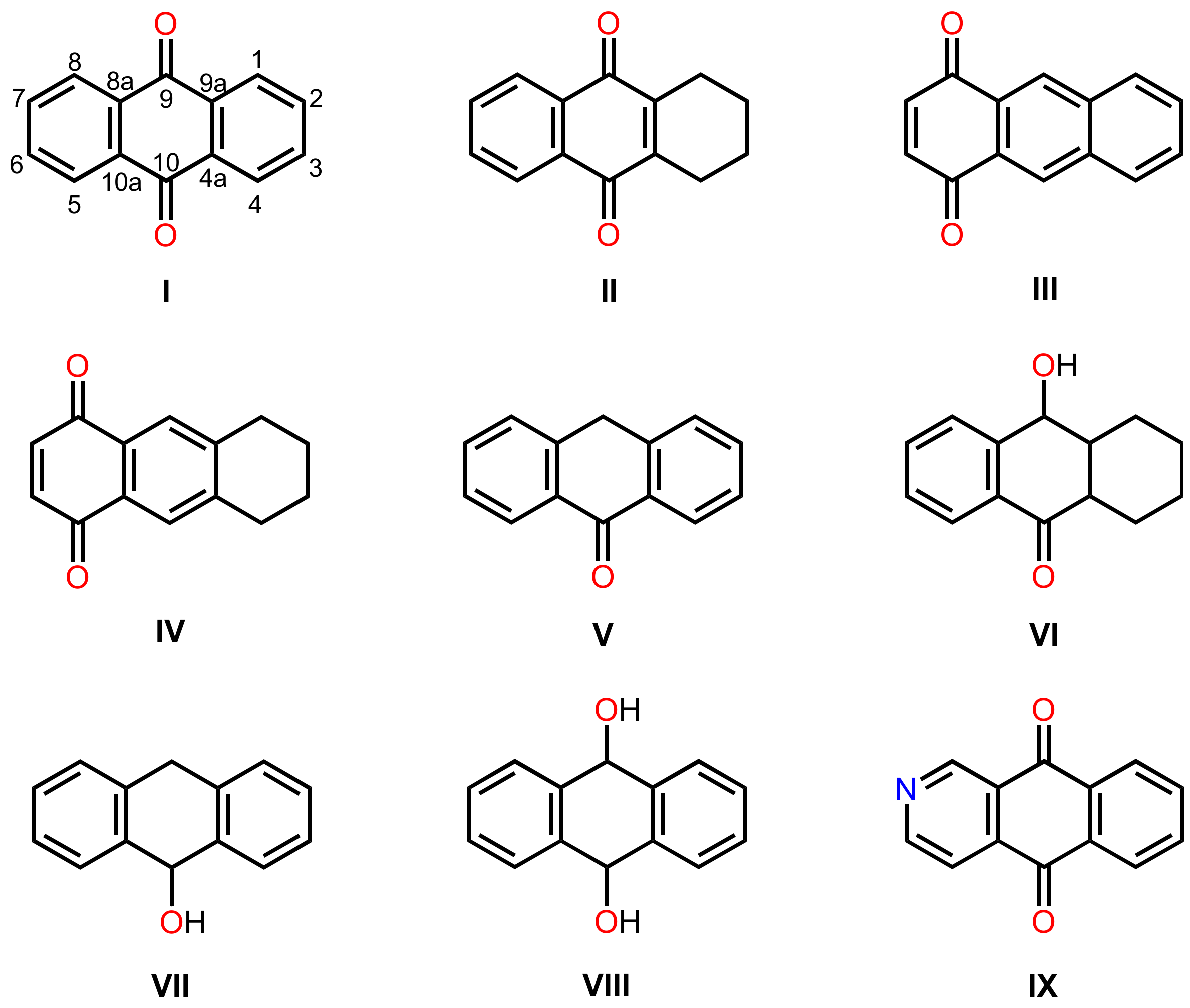
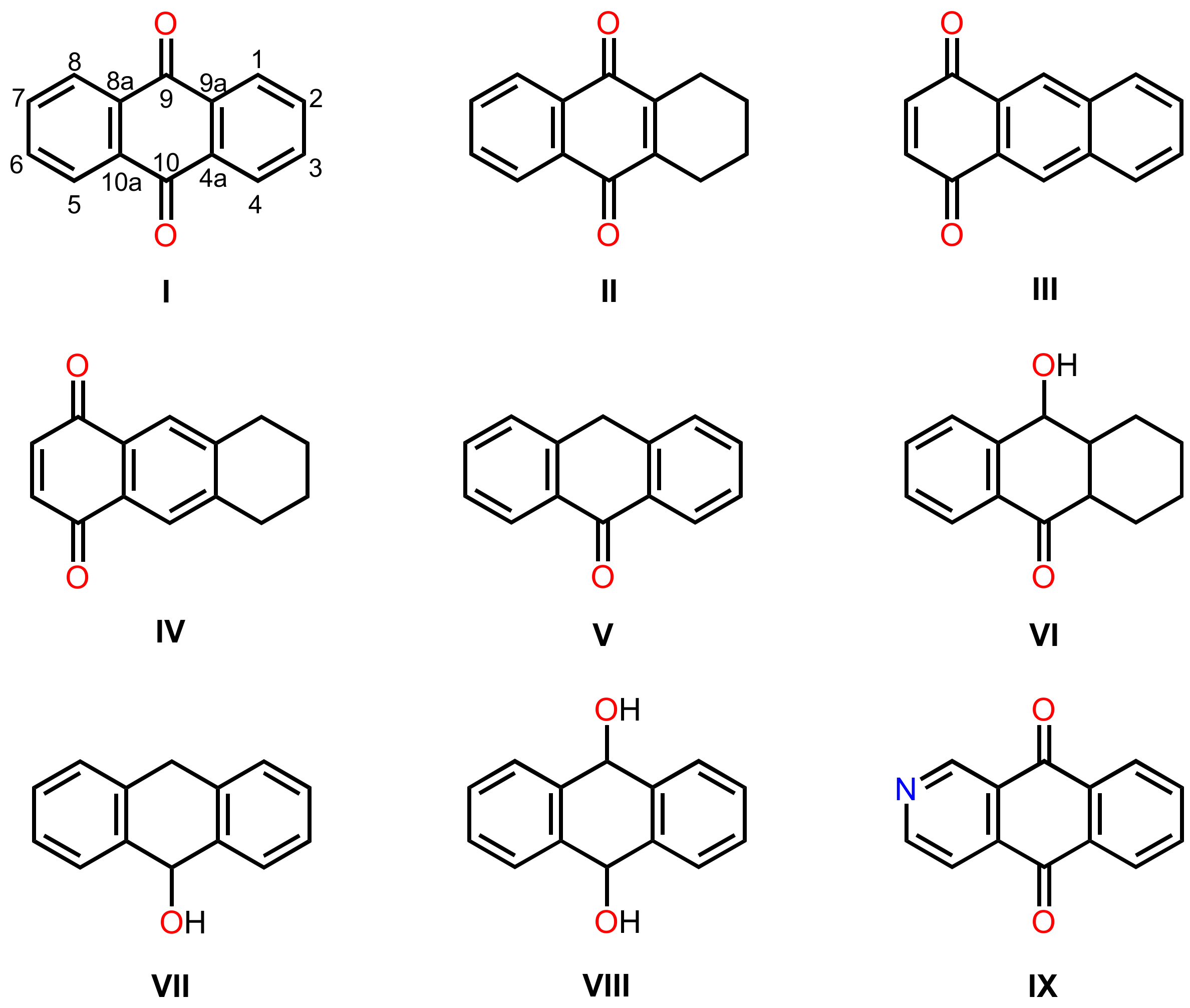
3.1. Anthraquinones
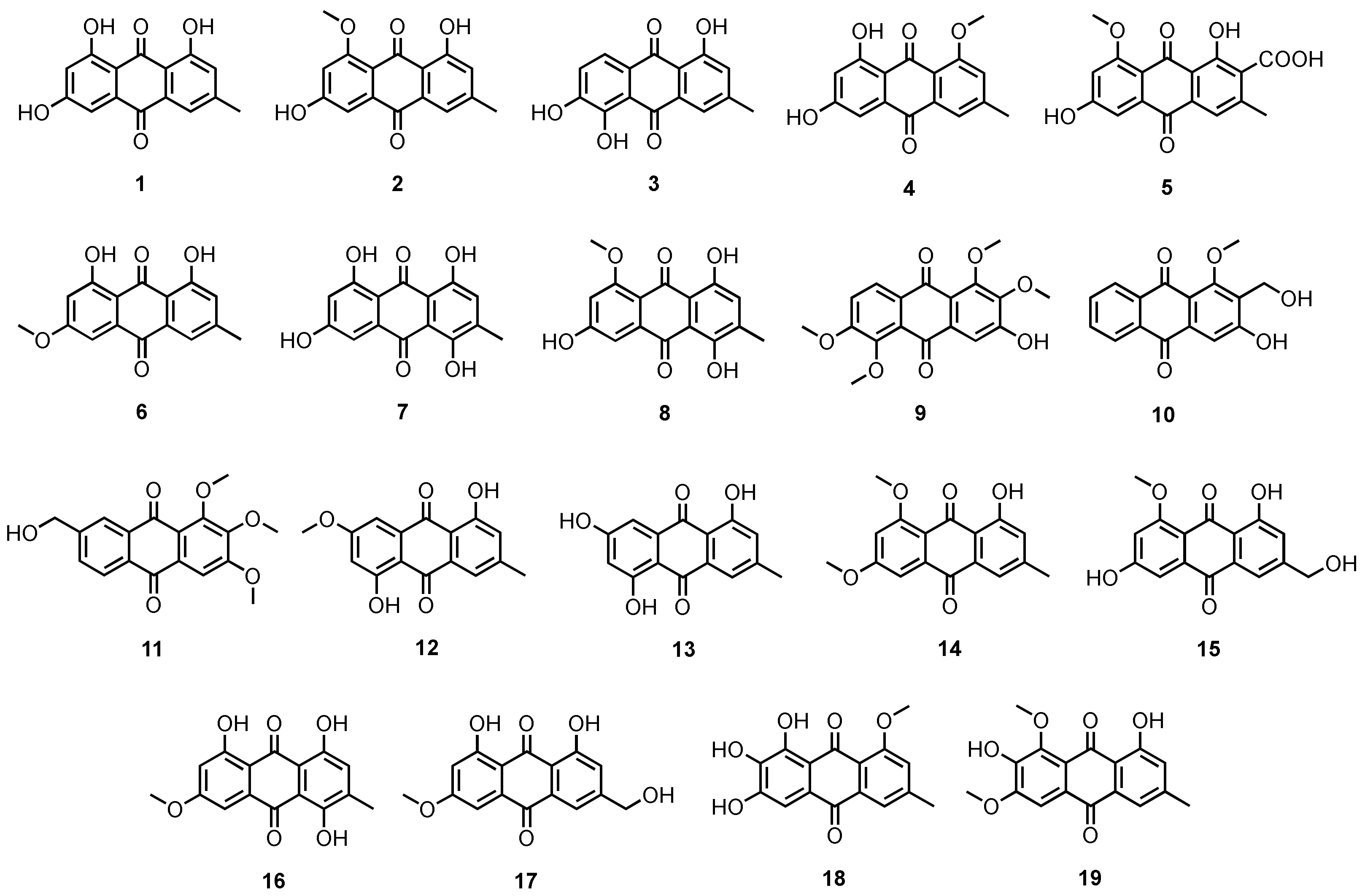
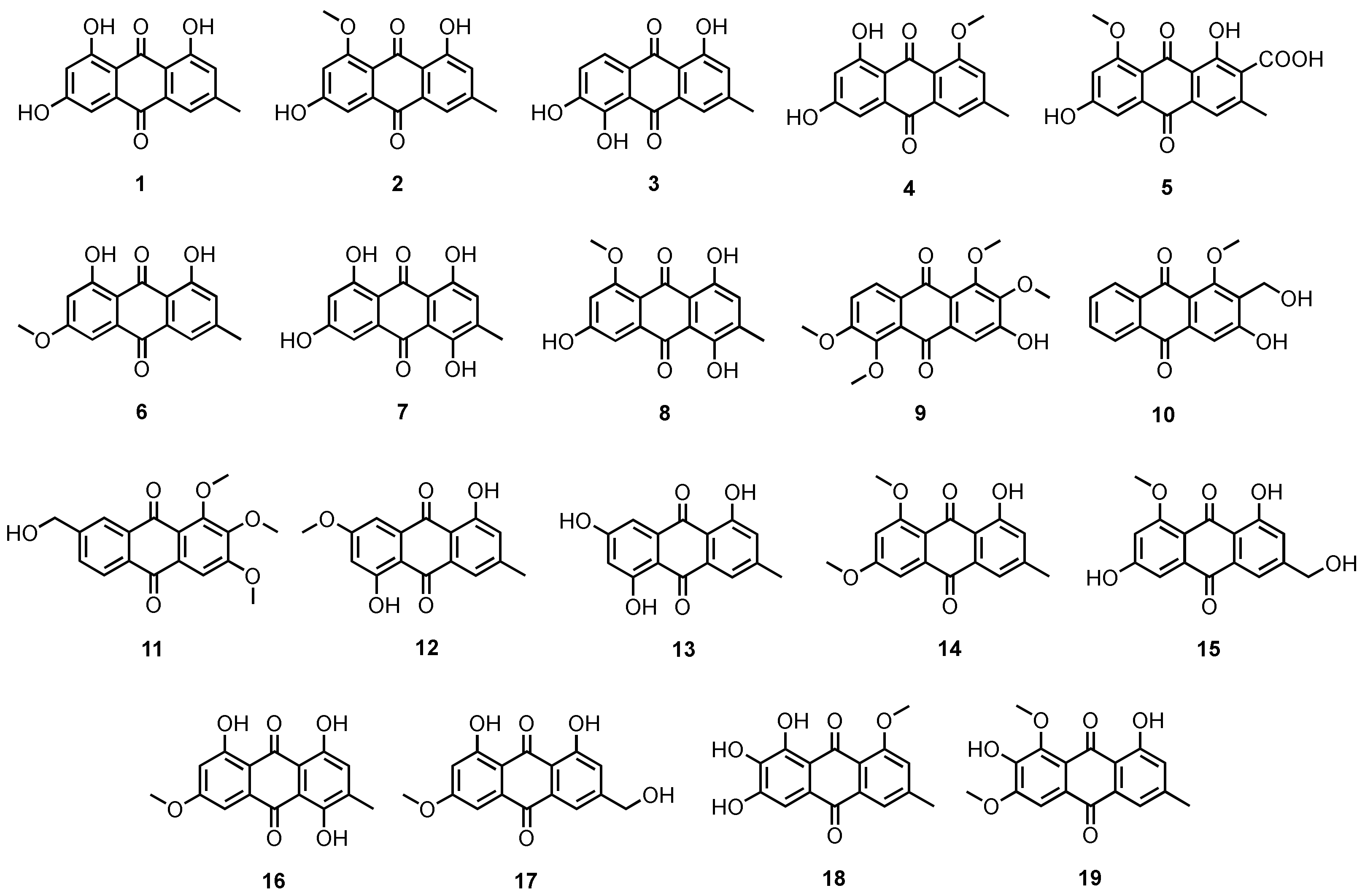
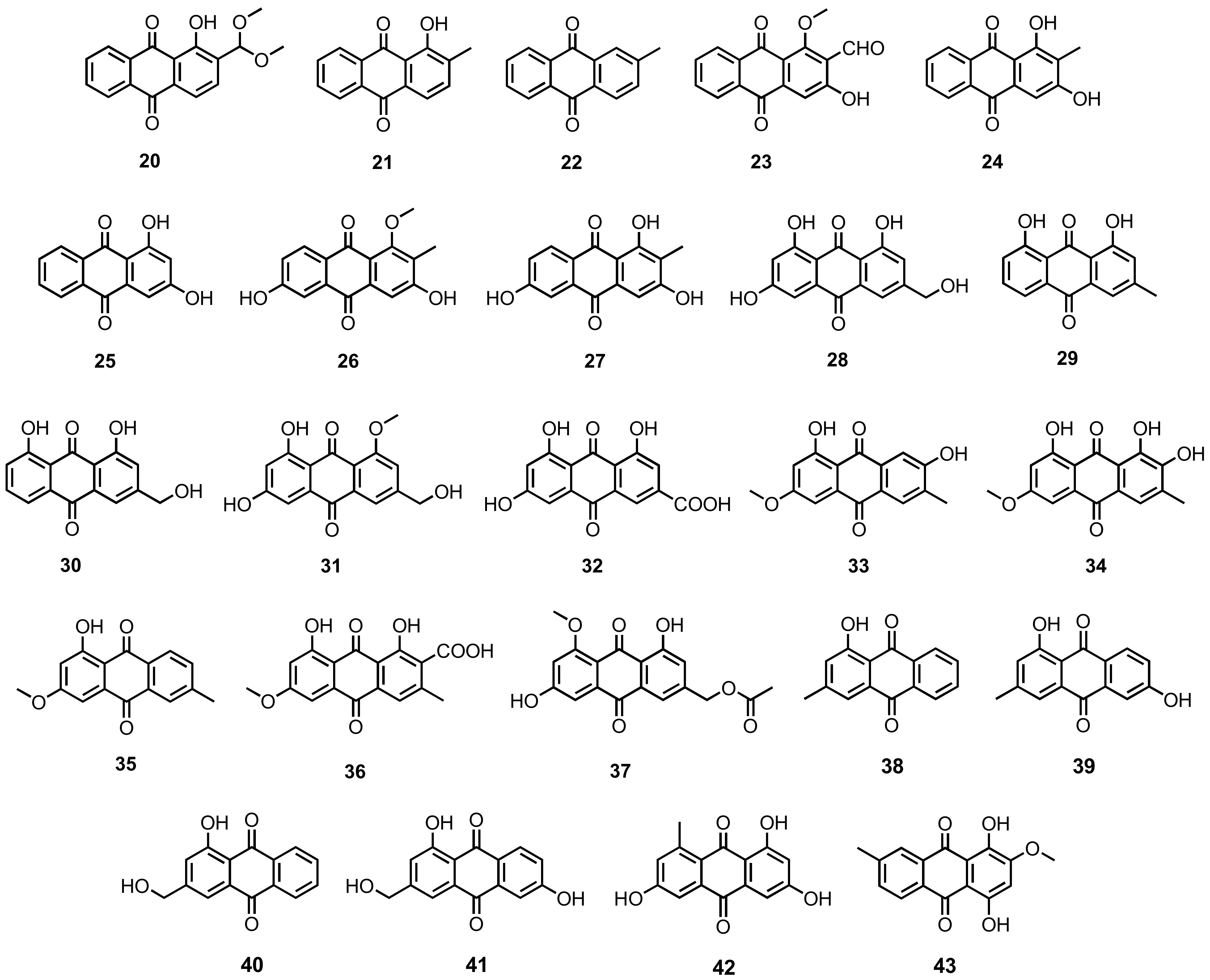
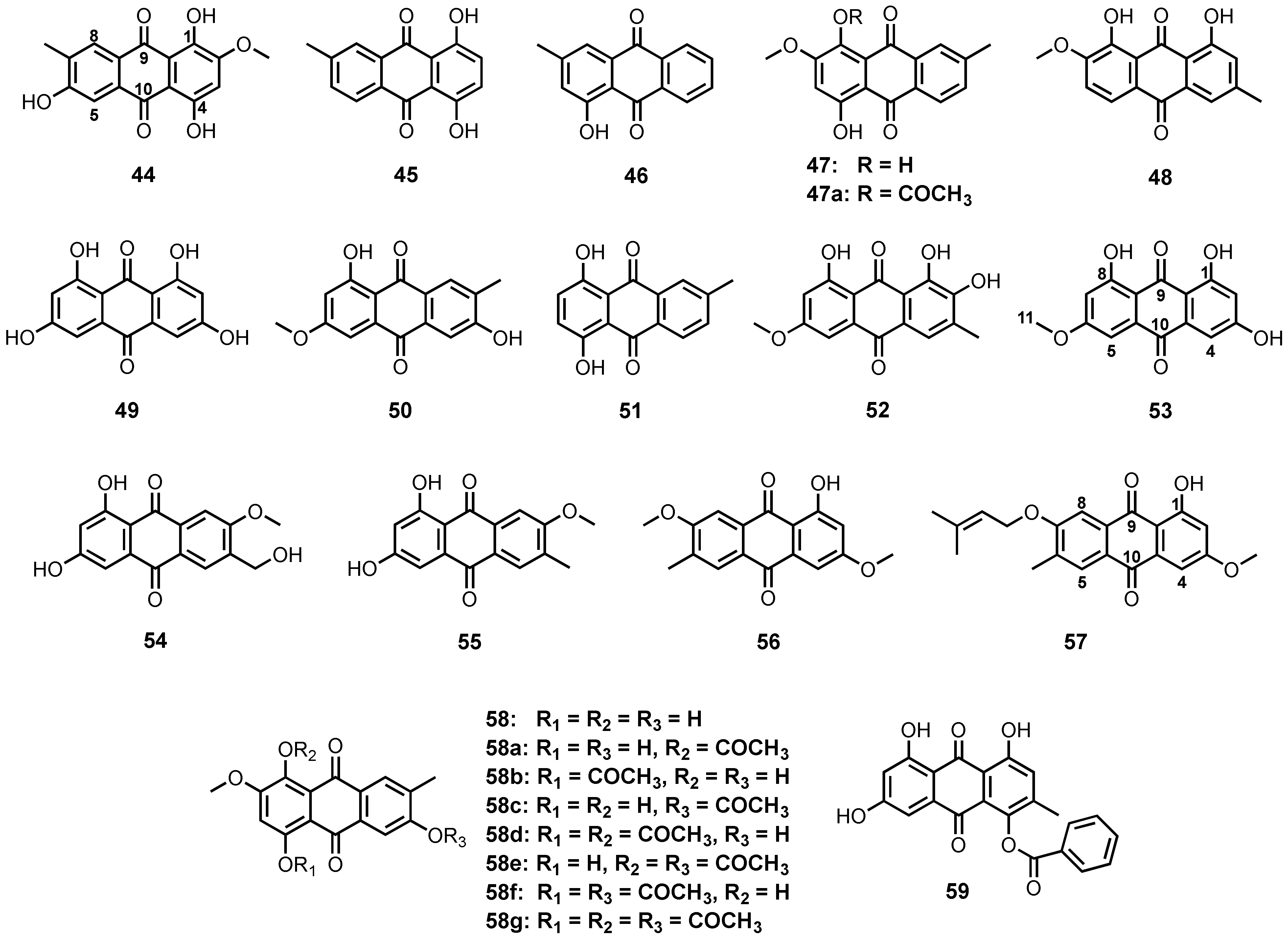
3.1.1. Simple Anthraquinones
3.1.2. Halogenated Anthraquinones
3.1.3. Sulphated Anthraquinones
3.1.4. Glycosylated Anthraquinones
3.1.5. Seco-Anthraquinones
3.2. Tetrahydroanthraquinones
3.3. Tetrahydro-5,8-anthraquinones
3.4. Anthrones
3.5. Tetrahydro-9-hydroxyanthrones
3.6. Tetrahydroanthrols
3.7. 9,10-Dihydroxyanthracenes
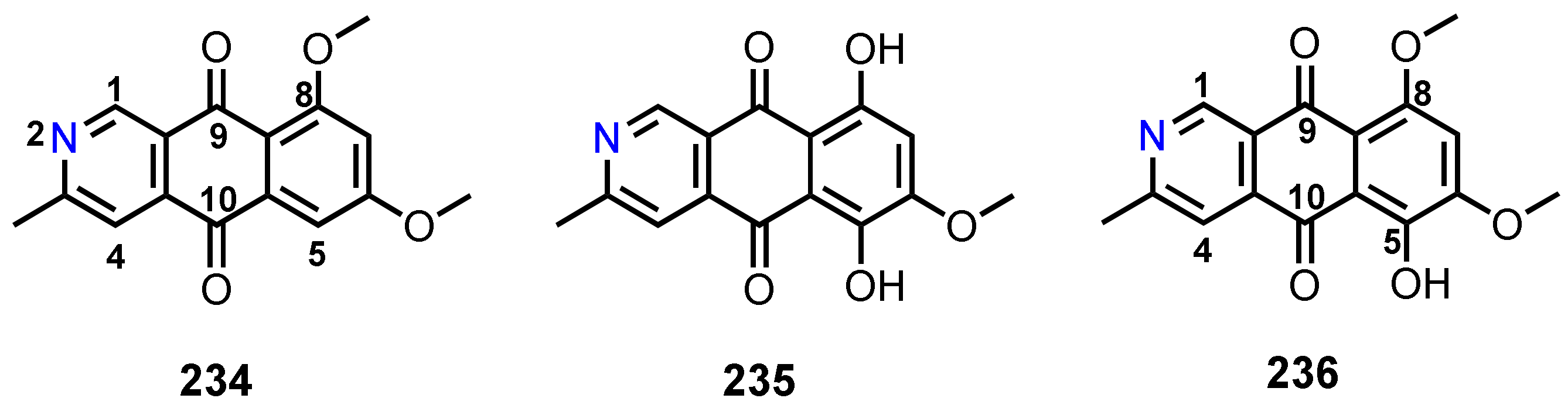
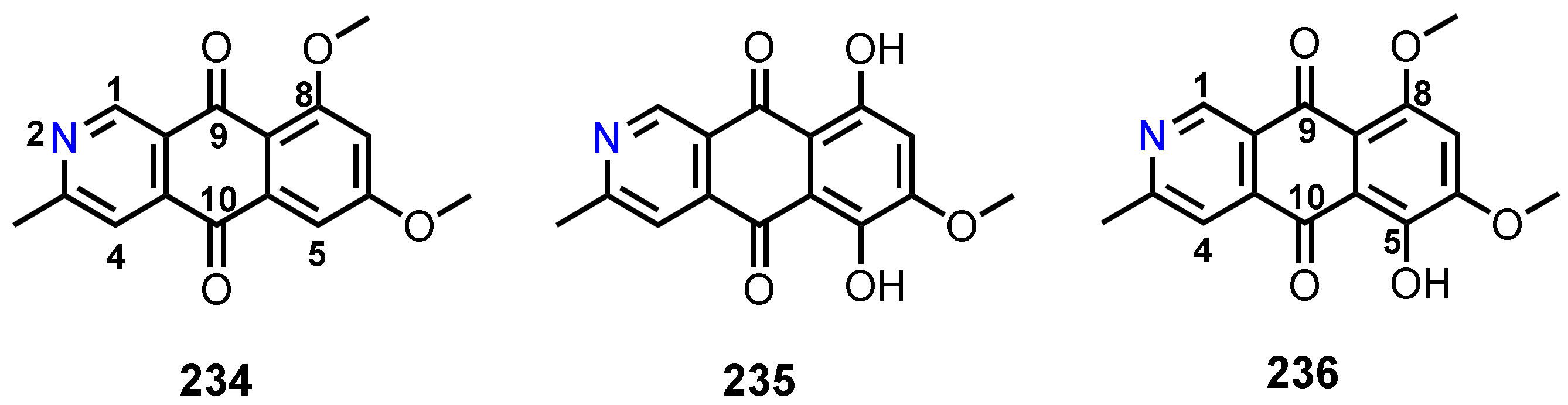
3.8. 2-Aza-anthraquinones
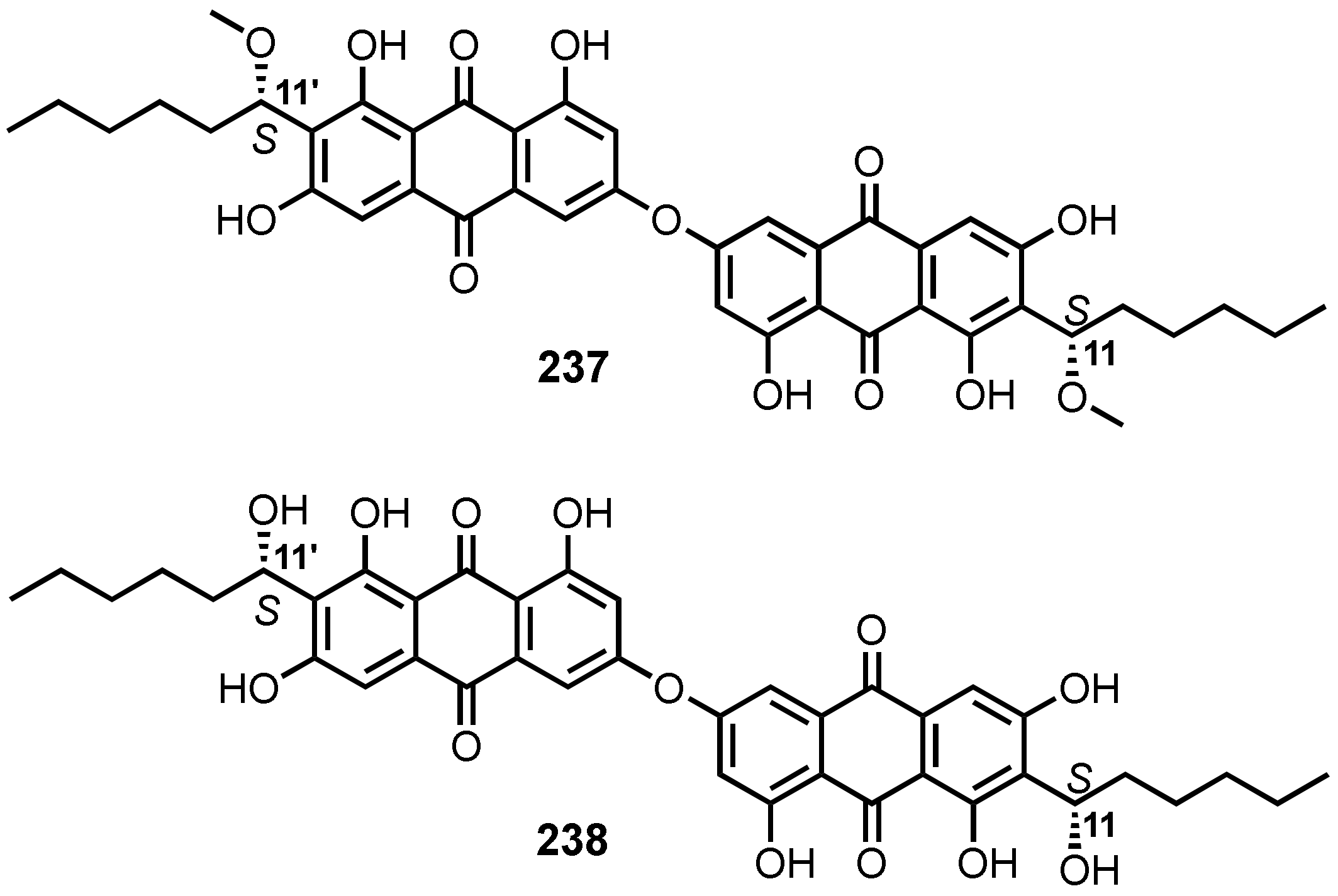
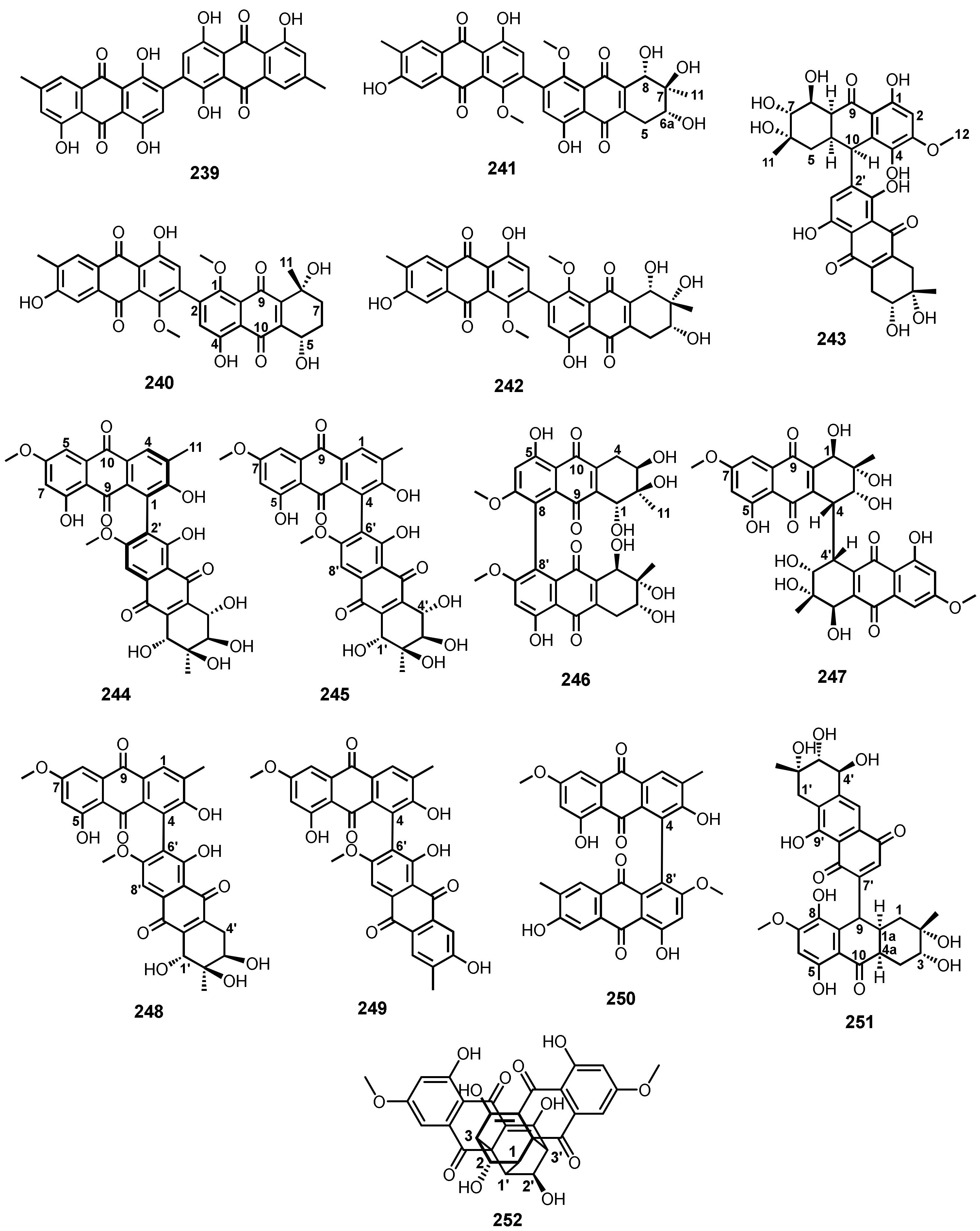
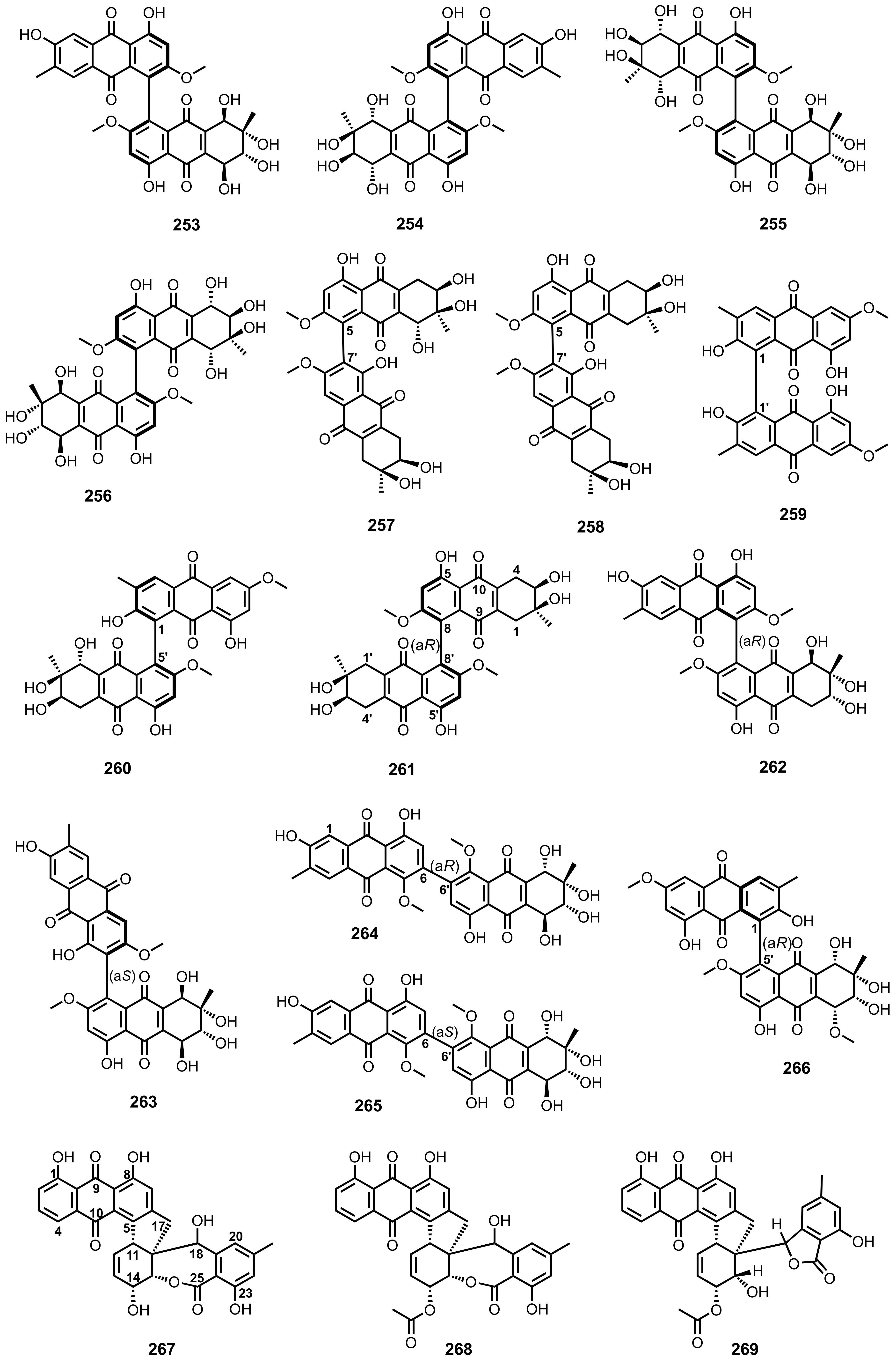
3.9. Dimeric anthraquinones
3.10. Bianthrones
3.11. Anthraquinone Analogues Fused with Xanthone and Chromone Derivatives
4. Biological Activitiesq
4.1. Antibacterial and Antibiofilm Activities
4.2. Antifungal Activity
4.3. Antiviral Activity
4.4. Antiparasitic Activity
4.5. Cytotoxic Activity
4.6. Enzyme Inhibitory Activity
4.6.1. Inhibition of α-Glucosidase Activity
4.6.2. Inhibition of Trypsin Activity
4.6.3. Inhibition of Tyrosinase Activity
4.6.4. Inhibition of Indoleamine 2,3-dioxygenase (IDO1) Activity
4.6.5. Inhibition of Protein Tyrosine Phosphatases and Protein Kinases Activity
4.6.6. Inhibition of Acetylcholinesterase (AChE) Activity
4.7. Anti-Inflammatory Activity
4.8. Anti-Obesity Activity
4.9. Anticoagulant Activity
4.10. Antiangiogenic Activity
4.11. Antifouling Activity
4.12. Algicidal Activity
4.13. Insecticidal Activity
4.14. Antioxidant Activity
4.15. Other Biological Activities
5. Concluding Remarks and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gladfelter, A.S.; James, T.Y.; Amend, A.S. Marine fungi. Curr. Biol. 2019, 29, R191–R195. [Google Scholar] [CrossRef] [PubMed]
- Amend, A.; Burgaud, G.; Cunliffe, M.; Edgcomb, V.P.; Ettinger, C.L.; Gutiérrez, M.; Heitman, J.; Hom, E.F.; Ianiri, G.; Jones, A.C. Fungi in the marine environment: Open questions and unsolved problems. MBio 2019, 10, e01189-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kis-papo, T. Marine fungal communities. In The Fungal Community: Its Organization and Role in the Ecosystem, 3rd ed.; Dighton, J., White, J.F., Oudemans, P., Eds.; CRC Press: Boca Raton, FL, USA; Taylor & Francis Group: Oxfordshire, UK, 2005; Volume 23, pp. 61–92. [Google Scholar]
- Kohlmeyer, J.; Kohlmeyer, E. Marine Mycology, the Higher Fungi; Academic Press: New York, NY, USA, 1979; pp. 1–3. [Google Scholar]
- Pang, K.-L.; Overy, D.P.; Jones, E.G.; da Luz Calado, M.; Burgaud, G.; Walker, A.K.; Johnson, J.A.; Kerr, R.G.; Cha, H.-J.; Bills, G.F. ‘Marine fungi’and ‘marine-derived fungi’in natural product chemistry research: Toward a new consensual definition. Fungal Biol. Rev. 2016, 30, 163–175. [Google Scholar] [CrossRef]
- Grossart, H.-P.; Van den Wyngaert, S.; Kagami, M.; Wurzbacher, C.; Cunliffe, M.; Rojas-Jimenez, K. Fungi in aquatic ecosystems. Nat. Rev. Microbiol. 2019, 17, 339–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bugni, T.S.; Ireland, C.M. Marine-derived fungi: A chemically and biologically diverse group of microorganisms. Nat. Prod. Rep. 2004, 21, 143–163. [Google Scholar] [CrossRef]
- Liu, Y.; Palaniveloo, K.; Alias, S.A.; Sathiya Seelan, J.S. Species diversity and secondary metabolites of Sarcophyton-associated marine fungi. Molecules 2021, 26, 3227. [Google Scholar] [CrossRef]
- Saleem, M.; Ali, M.S.; Hussain, S.; Jabbar, A.; Ashraf, M.; Lee, Y.S. Marine natural products of fungal origin. Nat. Prod. Rep. 2007, 24, 1142–1152. [Google Scholar] [CrossRef]
- Tao, L.Y.; Zhang, J.Y.; Liang, Y.J.; Chen, L.M.; Zheng, L.S.; Wang, F.; Mi, Y.J.; She, Z.G.; To, K.K.W.; Lin, Y.C. Anticancer effect and structure-activity analysis of marine products isolated from metabolites of mangrove fungi in the South China Sea. Mar. Drugs 2010, 8, 1094–1105. [Google Scholar] [CrossRef]
- Fouillaud, M.; Venkatachalam, M.; Girard-Valenciennes, E.; Caro, Y.; Dufossé, L. Anthraquinones and derivatives from marine-derived fungi: Structural diversity and selected biological activities. Mar. Drugs 2016, 14, 64. [Google Scholar] [CrossRef] [Green Version]
- Greco, G.; Turrini, E.; Catanzaro, E.; Fimognari, C. Marine anthraquinones: Pharmacological and toxicological issues. Mar. Drugs 2021, 19, 272. [Google Scholar] [CrossRef]
- Wang, X.; Mao, Z.-G.; Song, B.-B.; Chen, C.-H.; Xiao, W.-W.; Hu, B.; Wang, J.-W.; Jiang, X.-B.; Zhu, Y.-H.; Wang, H.-J. Advances in the study of the structures and bioactivities of metabolites isolated from mangrove-derived fungi in the South China Sea. Mar. Drugs 2013, 11, 3601–3616. [Google Scholar] [CrossRef] [Green Version]
- Gessler, N.; Egorova, A.; Belozerskaya, T. Fungal anthraquinones. Appl. Biochem. Microbiol. 2013, 49, 85–99. [Google Scholar] [CrossRef]
- Javidpour, P.; Korman, T.P.; Shakya, G.; Tsai, S.-C. Structural and biochemical analyses of regio-and stereospecificities observed in a type II polyketide ketoreductase. Biochemistry 2011, 50, 4638–4649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buttachon, S.; Ramos, A.A.; Inácio, Â.; Dethoup, T.; Gales, L.; Lee, M.; Costa, P.M.; Silva, A.; Sekeroglu, N.; Rocha, E. Bis-indolyl benzenoids, hydroxypyrrolidine derivatives and other constituents from cultures of the marine sponge-associated fungus Aspergillus candidus KUFA0062. Mar. Drugs 2018, 16, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eze, P.M.; Gao, Y.; Liu, Y.; Geelen, L.V.; Ejikeugwu, C.P.; Esimone, C.O.; Okoye, F.B.C.; Proksch, P.; Kalscheuer, R. Secondary metabolites of a marine-derived Penicillium ochrochloron. Not. Sci. Biol. 2021, 13, 11020. [Google Scholar] [CrossRef]
- Wang, C.; Guo, L.; Hao, J.; Wang, L.; Zhu, W. α-Glucosidase inhibitors from the marine-derived fungus Aspergillus flavipes HN4-13. J. Nat. Prod. 2016, 79, 2977–2981. [Google Scholar] [CrossRef]
- Liu, Y.; Ding, L.; He, J.; Zhang, Z.; Deng, Y.; He, S.; Yan, X. A new antibacterial chromone from a marine sponge-associated fungus Aspergillus sp. LS57. Fitoterapia 2021, 154, 105004. [Google Scholar] [CrossRef]
- Girich, E.V.; Yurchenko, A.N.; Smetanina, O.F.; Trinh, P.T.H.; Ngoc, N.T.D.; Pivkin, M.V.; Popov, R.S.; Pislyagin, E.A.; Menchinskaya, E.S.; Chingizova, E.A. Neuroprotective metabolites from vietnamese marine derived fungi of Aspergillus and Penicillium genera. Mar. Drugs 2020, 18, 608. [Google Scholar] [CrossRef]
- Du, X.; Liu, D.; Huang, J.; Zhang, C.; Proksch, P.; Lin, W. Polyketide derivatives from the sponge associated fungus Aspergillus europaeus with antioxidant and NO inhibitory activities. Fitoterapia 2018, 130, 190–197. [Google Scholar] [CrossRef]
- Du, L.; Zhu, T.; Liu, H.; Fang, Y.; Zhu, W.; Gu, Q. Cytotoxic polyketides from a marine-derived fungus Aspergillus glaucus. J. Nat. Prod. 2008, 71, 1837–1842. [Google Scholar] [CrossRef]
- Li, X.; Li, X.-M.; Xu, G.-M.; Li, C.-S.; Wang, B.-G. Antioxidant metabolites from marine alga-derived fungus Aspergillus wentii EN-48. Phytochem. Lett. 2014, 7, 120–123. [Google Scholar] [CrossRef]
- Smetanina, O.; Kalinovskii, A.; Khudyakova, Y.V.; Slinkina, N.; Pivkin, M.; Kuznetsova, T. Metabolites from the marine fungus Eurotium repens. Chem. Nat. Compd. 2007, 43, 395–398. [Google Scholar] [CrossRef]
- Almeida, A.P.; Dethoup, T.; Singburaudom, N.; Lima, R.; Vasconcelos, M.H.; Pinto, M.; Kijjoa, A. The in vitro anticancer activity of the crude extract of the sponge-associated fungus Eurotium cristatum and its secondary metabolites. J. Nat. Pharm. 2010, 1, 25–29. [Google Scholar] [CrossRef]
- Wijesekara, I.; Zhang, C.; Van Ta, Q.; Vo, T.-S.; Li, Y.-X.; Kim, S.-K. Physcion from marine-derived fungus Microsporum sp. induces apoptosis in human cervical carcinoma HeLa cells. Microbiol. Res. 2014, 169, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zhu, R.; Yi, W.; Chai, W.; Zhang, Z.; Lian, X.-Y. Peniciphenalenins A−F from the culture of a marine-associated fungus Penicillium sp. ZZ901. Phytochemistry 2018, 152, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Liao, Y.; Tang, C.; Huang, X.; Luo, Z.; Chen, J.; Cai, P. Cytotoxic and antibacterial compounds from the coral-derived fungus Aspergillus tritici SP2-8-1. Mar. Drugs 2017, 15, 348. [Google Scholar] [CrossRef] [Green Version]
- Miao, F.; Zuo, J.; Liu, X.; Ji, N. Algicidal activities of secondary metabolites of marine macroalgal-derived endophytic fungi. J. Oceanol. Limnol. 2019, 37, 112–121. [Google Scholar] [CrossRef]
- Sun, H.F.; Li, X.M.; Meng, L.H.; Cui, C.M.; Gao, S.S.; Li, C.S.; Wang, B.G. Two new secoanthraquinone derivatives from the marine-derived endophytic fungus Aspergillus wentii EN-48. Helv. Chim. Acta 2013, 96, 458–462. [Google Scholar] [CrossRef]
- Zin, W.W.M.; Buttachon, S.; Dethoup, T.; Pereira, J.A.; Gales, L.; Inácio, Â.; Costa, P.M.; Lee, M.; Sekeroglu, N.; Silva, A.M.; et al. Antibacterial and antibiofilm activities of the metabolites isolated from the culture of the mangrove-derived endophytic fungus Eurotium chevalieri KUFA 0006. Phytochemistry 2017, 141, 86–97. [Google Scholar] [CrossRef]
- Wang, S.; Li, X.-M.; Teuscher, F.; Li, D.-L.; Diesel, A.; Ebel, R.; Proksch, P.; Wang, B.-G. Chaetopyranin, a benzaldehyde derivative, and other related metabolites from Chaetomium globosum, an endophytic fungus derived from the marine red alga Polysiphonia urceolata. J. Nat. Prod. 2006, 69, 1622–1625. [Google Scholar] [CrossRef]
- Noinart, J.; Buttachon, S.; Dethoup, T.; Gales, L.; Pereira, J.A.; Urbatzka, R.; Freitas, S.; Lee, M.; Silva, A.; Pinto, M.M.A.; et al. A new ergosterol analog, a new bis-anthraquinone and anti-obesity activity of anthraquinones from the marine sponge-associated fungus Talaromyces stipitatus KUFA 0207. Mar. Drugs 2017, 15, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawas, U.W.; El-Beih, A.A.; El-Halawany, A.M. Bioactive anthraquinones from endophytic fungus Aspergillus versicolor isolated from red sea algae. Arch. Pharm. Res. 2012, 35, 1749–1756. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Chen, R.; Luo, Z.; Wang, W.; Chen, J. Antimicrobial activity and molecular docking studies of a novel anthraquinone from a marine-derived fungus Aspergillus versicolor. Nat. Prod. Res. 2018, 32, 558–563. [Google Scholar] [CrossRef]
- Wang, P.-L.; Li, D.-Y.; Xie, L.-R.; Wu, X.; Hua, H.-M.; Li, Z.-L. Two new compounds from a marine-derived fungus Penicillium oxalicum. Nat. Prod. Res. 2014, 28, 290–293. [Google Scholar] [CrossRef]
- Khamthong, N.; Rukachaisirikul, V.; Phongpaichit, S.; Preedanon, S.; Sakayaroj, J. Bioactive polyketides from the sea fan-derived fungus Penicillium citrinum PSU-F51. Tetrahedron 2012, 68, 8245–8250. [Google Scholar] [CrossRef]
- Ren, H.; Tian, L.; Gu, Q.; Zhu, W. Secalonic acid D; A cytotoxic constituent from marine lichen-derived fungus Gliocladium sp. T31. Arch. Pharm. Res. 2006, 29, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Bao, J.; Sun, Y.-L.; Zhang, X.-Y.; Han, Z.; Gao, H.-C.; He, F.; Qian, P.-Y.; Qi, S.-H. Antifouling and antibacterial polyketides from marine gorgonian coral-associated fungus Penicillium sp. SCSGAF 0023. J. Antibiot. 2013, 66, 219–223. [Google Scholar] [CrossRef] [Green Version]
- Hawas, U.W.; Al-Farawati, R. Chemical constituents and antiviral activity from marine endophytic fungi from Red Sea alga Padina pavonica. J. Chem. Soc. Pak. 2017, 39, 478. [Google Scholar]
- He, K.-Y.; Zhang, C.; Duan, Y.-R.; Huang, G.-L.; Yang, C.-Y.; Lu, X.-R.; Zheng, C.-J.; Chen, G.-Y. New chlorinated xanthone and anthraquinone produced by a mangrove-derived fungus Penicillium citrinum HL-5126. J. Antibiot. 2017, 70, 823–827. [Google Scholar] [CrossRef]
- Wen, L.; Lin, Y.-C.; She, Z.-G.; Du, D.-S.; Chan, W.-L.; Zheng, Z.-H. Paeciloxanthone, a new cytotoxic xanthone from the marine mangrove fungus Paecilomyces sp. (Tree1-7). J. Asian Nat. Prod. Res. 2008, 10, 133–137. [Google Scholar] [CrossRef]
- Qi, J.; Zhao, P.; Zhao, L.; Jia, A.; Liu, C.; Zhang, L.; Xia, X. Anthraquinone derivatives from a sea cucumber-derived Trichoderma sp. fungus with antibacterial activities. Chem. Nat. Compd. 2020, 56, 112–114. [Google Scholar] [CrossRef]
- Maarisit, W.; Ueda, K. Antimicrobial metabolites from a marine-derived fungus. Indones. J. Pharm. 2013, 24, 163–169. [Google Scholar] [CrossRef]
- Bugni, T.S.; Janso, J.E.; Williamson, R.T.; Feng, X.; Bernan, V.S.; Greenstein, M.; Carter, G.T.; Maiese, W.M.; Ireland, C.M. Dictyosphaeric acids A and B: New decalactones from an undescribed Penicillium sp. obtained from the alga Dictyosphaeria versluyii. J. Nat. Prod. 2004, 67, 1396–1399. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Cui, Z.; Huang, H.; Song, X.; Sun, A.; Dang, Y.; Lu, L.; Ju, J. Amino acid conjugated anthraquinones from the marine-derived fungus Penicillium sp. SCSIO sof101. J. Nat. Prod. 2017, 80, 1668–1673. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.J.; Li, X.M.; Li, C.S.; Wang, B.G. Alkaloid and anthraquinone derivatives produced by the marine-derived endophytic fungus Eurotium rubrum. Helv. Chim. Acta 2012, 95, 163–168. [Google Scholar] [CrossRef]
- Wang, F.; Li, H.; Lan, W. The metabolites of the marine fungus Penicillium sp. associated with soft coral Sarcophyton tortuosum. Acta Sci. Nat. Univ. Sunyatseni 2011, 50, 72–77. [Google Scholar]
- Zheng, C.-J.; Shao, C.-L.; Guo, Z.-Y.; Chen, J.-F.; Deng, D.-S.; Yang, K.-L.; Chen, Y.-Y.; Fu, X.-M.; She, Z.-G.; Lin, Y.-C. Bioactive hydroanthraquinones and anthraquinone dimers from a soft coral-derived Alternaria sp. fungus. J. Nat. Prod. 2012, 75, 189–197. [Google Scholar] [CrossRef]
- Zhou, X.-M.; Zheng, C.-J.; Chen, G.-Y.; Song, X.-P.; Han, C.-R.; Li, G.-N.; Fu, Y.-H.; Chen, W.-H.; Niu, Z.-G. Bioactive anthraquinone derivatives from the mangrove-derived fungus Stemphylium sp. 33231. J. Nat. Prod. 2014, 77, 2021–2028. [Google Scholar] [CrossRef]
- Li, J.; Zheng, Y.-B.; Kurtán, T.; Liu, M.-X.; Tang, H.; Zhuang, C.-L.; Zhang, W. Anthraquinone derivatives from a coral associated fungus Stemphylium lycopersici. Nat. Prod. Res. 2020, 34, 2116–2123. [Google Scholar] [CrossRef]
- Klaiklay, S.; Rukachaisirikul, V.; Phongpaichit, S.; Pakawatchai, C.; Saithong, S.; Buatong, J.; Preedanon, S.; Sakayaroj, J. Anthraquinone derivatives from the mangrove-derived fungus Phomopsis sp. PSU-MA214. Phytochem. Lett. 2012, 5, 738–742. [Google Scholar] [CrossRef]
- Bovio, E.; Fauchon, M.; Toueix, Y.; Mehiri, M.; Varese, G.C.; Hellio, C. The sponge-associated fungus Eurotium chevalieri MUT 2316 and its bioactive molecules: Potential applications in the field of antifouling. Mar. Biotechnol. 2019, 21, 743–752. [Google Scholar] [CrossRef] [PubMed]
- De Sá, J.D.; Pereira, J.A.; Dethoup, T.; Cidade, H.; Sousa, M.E.; Rodrigues, I.C.; Costa, P.M.; Mistry, S.; Silva, A.; Kijjoa, A. Anthraquinones, diphenyl ethers, and their derivatives from the culture of the marine sponge-associated fungus Neosartorya spinosa KUFA 1047. Mar. Drugs 2021, 19, 457. [Google Scholar] [CrossRef] [PubMed]
- Shi, T.; Hou, X.-M.; Li, Z.-Y.; Cao, F.; Zhang, Y.-H.; Yu, J.-Y.; Zhao, D.-L.; Shao, C.-L.; Wang, C.-Y. Harzianumnones A and B: Two hydroxyanthraquinones from the coral-derived fungus Trichoderma harzianum. RSC Adv. 2018, 8, 27596–27601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, X.; Lin, X.; Tian, Y.; Liang, R.; Wang, J.; Yang, B.; Zhou, X.; Kaliyaperumal, K.; Luo, X.; Tu, Z. Three new polyketides from the marine sponge-derived fungus Trichoderma sp. SCSIO41004. Nat. Prod. Res. 2018, 32, 105–111. [Google Scholar] [CrossRef]
- She, Z.-G.; Huang, H.-R.; Lin, Y.-C.; Vrijmoed, L.; Jones, E.G. 1,4-Dihydroxy-2-methoxy-7-methylanthracene-9,10-dione. Acta Crystallogr. Sect. E Struct. Rep. Online 2006, 62, o3737–o3738. [Google Scholar] [CrossRef]
- Xia, X.K.; Huang, H.R.; She, Z.G.; Shao, C.L.; Liu, F.; Cai, X.L.; Vrijmoed, L.; Lin, Y.C. 1H and 13C NMR assignments for five anthraquinones from the mangrove endophytic fungus Halorosellinia sp. (No. 1403). Magn. Reson. Chem. 2007, 45, 1006–1009. [Google Scholar] [CrossRef]
- Trisuwan, K.; Khamthong, N.; Rukachaisirikul, V.; Phongpaichit, S.; Preedanon, S.; Sakayaroj, J. Anthraquinone, cyclopentanone, and naphthoquinone derivatives from the sea fan-derived fungi Fusarium spp. PSU-F14 and PSU-F135. J. Nat. Prod. 2010, 73, 1507–1511. [Google Scholar] [CrossRef]
- El-Beih, A.A.; Kawabata, T.; Koimaru, K.; Ohta, T.; Tsukamoto, S. Monodictyquinone A: A new antimicrobial anthraquinone from a sea urchin-derived fungus Monodictys sp. Chem. Pharm. Bull. 2007, 55, 1097–1098. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.-H.; Pan, J.-H.; Chen, B.; Yu, M.; Huang, H.-B.; Zhu, X.; Lu, Y.-J.; She, Z.-G.; Lin, Y.-C. Three bianthraquinone derivatives from the mangrove endophytic fungus Alternaria sp. ZJ9-6B from the South China Sea. Mar. Drugs 2011, 9, 832–843. [Google Scholar] [CrossRef]
- Xia, G.; Li, J.; Li, H.; Long, Y.; Lin, S.E.; Lu, Y.; He, L.; Lin, Y.; Liu, L.; She, Z. Alterporriol-type dimers from the mangrove endophytic fungus, Alternaria sp. (SK11), and their MptpB inhibitions. Mar. Drugs 2014, 12, 2953–2969. [Google Scholar] [CrossRef] [Green Version]
- Pan, D.; Zhang, X.; Zheng, H.; Zheng, Z.; Nong, X.; Liang, X.; Ma, X.; Qi, S. Novel anthraquinone derivatives as inhibitors of protein tyrosine phosphatases and indoleamine 2,3-dioxygenase 1 from the deep-sea derived fungus Alternaria tenuissima DFFSCS013. Org. Chem. Front. 2019, 6, 3252–3258. [Google Scholar] [CrossRef]
- Jadulco, R.; Brauers, G.; Edrada, R.A.; Ebel, R.; Wray, V.; Sudarsono, A.; Proksch, P. New metabolites from sponge-derived fungi Curvularia lunata and Cladosporium herbarum. J. Nat. Prod. 2002, 65, 730–733. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Cao, X.; Gu, Q. Antitumor metabolites from marine-derived fungus Gliocladium catenulatum T31. J. Chin. Pharm. Sci. 2010, 45, 1720–1723. [Google Scholar]
- Sobolevskaya, M.P.; Berdyshev, D.V.; Zhuravleva, O.I.; Denisenko, V.A.; Dyshlovoy, S.A.; von Amsberg, G.; Khudyakova, Y.V.; Kirichuk, N.N.; Afiyatullov, S.S. Polyketides metabolites from the marine sediment-derived fungus Thermomyces lanuginosus Tsikl. KMM 4681. Phytochem. Lett. 2021, 41, 114–118. [Google Scholar] [CrossRef]
- Huang, S.; Xu, J.; Li, F.; Zhou, D.; Xu, L.; Li, C. Identification and antifungal activity of metabolites from the mangrove fungus Phoma sp. L28. Chem. Nat. Compd. 2017, 53, 237–240. [Google Scholar] [CrossRef]
- Yang, K.-L.; Wei, M.-Y.; Shao, C.-L.; Fu, X.-M.; Guo, Z.-Y.; Xu, R.-F.; Zheng, C.-J.; She, Z.-G.; Lin, Y.-C.; Wang, C.-Y. Antibacterial anthraquinone derivatives from a sea anemone-derived fungus Nigrospora sp. J. Nat. Prod. 2012, 75, 935–941. [Google Scholar] [CrossRef]
- Xu, W.-F.; Hou, X.-M.; Yang, K.-L.; Cao, F.; Yang, R.-Y.; Wang, C.-Y.; Shao, C.-L. Nigrodiquinone A, a hydroanthraquinone dimer containing a rare C-9–C-7′ linkage from a zoanthid-derived Nigrospora sp. fungus. Mar. Drugs 2016, 14, 51. [Google Scholar] [CrossRef] [Green Version]
- Zhong, W.-M.; Wang, J.-F.; Wei, X.-Y.; Zeng, Q.; Chen, X.-Y.; Xiang, Y.; Tian, X.-P.; Zhang, S.; Long, L.-J.; Wang, F.-Z. (+)- and (−)-Eurotone A: A pair of enantiomeric polyketide dimers from a marine-derived fungus Eurotium sp. SCSIO F452. Tetrahedron Lett. 2019, 60, 1600–1603. [Google Scholar] [CrossRef]
- Brauers, G.; Edrada, R.A.; Ebel, R.; Proksch, P.; Wray, V.; Berg, A.; Gräfe, U.; Schächtele, C.; Totzke, F.; Finkenzeller, G. Anthraquinones and betaenone derivatives from the sponge-associated fungus Microsphaeropsis species: Novel inhibitors of protein kinases. J. Nat. Prod. 2000, 63, 739–745. [Google Scholar] [CrossRef]
- Kjer, J.; Debbab, A.; Aly, A.H.; Proksch, P. Methods for isolation of marine-derived endophytic fungi and their bioactive secondary products. Nat. Protoc. 2010, 5, 479–490. [Google Scholar] [CrossRef]
- Zhao, D.-L.; Wang, D.; Tian, X.-Y.; Cao, F.; Li, Y.-Q.; Zhang, C.-S. Anti-phytopathogenic and cytotoxic activities of crude extracts and secondary metabolites of marine-derived fungi. Mar. Drugs 2018, 16, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.; Cui, Y.; Cai, C.; Chen, H.; Dai, Y.; Chen, P.; Kong, F.; Yuan, J.; Song, X.; Mei, W. Two new succinimide derivatives cladosporitins A and B from the mangrove-derived fungus Cladosporium sp. HNWSW-1. Mar. Drugs 2018, 17, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, C.; Wang, C.; Zheng, C.; She, Z.; Gu, Y.; Lin, Y. A new anthraquinone derivative from the marine endophytic fungus Fusarium sp. (No. b77). Nat. Prod. Res. 2010, 24, 81–85. [Google Scholar] [CrossRef]
- Jin, Y.; Qin, S.; Gao, H.; Zhu, G.; Wang, W.; Zhu, W.; Wang, Y. An anti-HBV anthraquinone from aciduric fungus Penicillium sp. OUCMDZ-4736 under low pH stress. Extremophiles 2018, 22, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Mandelare, P.; Adpressa, D.; Kaweesa, E.; Zakharov, L.; Loesgen, S. Coculture of two developmental stages of a marine-derived Aspergillus alliaceus results in the production of the cytotoxic bianthrone allianthrone A. J. Nat. Prod. 2018, 81, 1014–1022. [Google Scholar] [CrossRef]
- Fredimoses, M.; Zhou, X.; Ai, W.; Tian, X.; Yang, B.; Lin, X.; Liu, J.; Liu, Y. Emerixanthone E, a new xanthone derivative from deep sea fungus Emericella sp SCSIO 05240. Nat. Prod. Res. 2019, 33, 2088–2094. [Google Scholar] [CrossRef]
- Huang, Z.-J.; Yang, R.-Y.; Guo, Z.-Y.; She, Z.-G.; Lin, Y.-C. New anthraquinone derivative produced by cultivation of mangrove endophytic fungus Fusarium sp. ZZF60 from the South China Sea. Chin. J. Appl. Chem. 2010, 27, 394. [Google Scholar] [CrossRef]
- Yang, S.-Q.; Li, X.-M.; Xu, G.-M.; Li, X.; An, C.-Y.; Wang, B.-G. Antibacterial anthraquinone derivatives isolated from a mangrove-derived endophytic fungus Aspergillus nidulans by ethanol stress strategy. J. Antibiot. 2018, 71, 778–784. [Google Scholar] [CrossRef]
- Liang, X.; Huang, Z.-H.; Ma, X.; Zheng, Z.-H.; Zhang, X.-X.; Lu, X.-H.; Qi, S.-H. Mycotoxins as inhibitors of protein tyrosine phosphatases from the deep-sea-derived fungus Aspergillus puniceus SCSIO z021. Bioorg. Chem. 2021, 107, 104571. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, X.-M.; Wang, B.-G. Anthraquinone derivatives produced by marine-derived fungus Aspergillus versicolor EN-7. Biosci. Biotechnol. Biochem. 2012, 76, 1774–1776. [Google Scholar] [CrossRef] [Green Version]
- Shao, C.; She, Z.; Guo, Z.; Peng, H.; Cai, X.; Zhou, S.; Gu, Y.; Lin, Y. 1H and 13C NMR assignments for two anthraquinones and two xanthones from the mangrove fungus (ZSUH-36). Magn. Reson. Chem. 2007, 45, 434–438. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.M.; Li, H.; Hong, J.; Cho, H.Y.; Bae, K.S.; Kim, M.; Kim, D.-K.; Jung, J.H. Bioactive metabolites from the sponge-derived fungus Aspergillus versicolor. Arch. Pharm. Res. 2010, 33, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Gu, Q.-Q.; Cui, C.-B. Anthraquinone derivatives produced by marine-derived Penicillium flavidorsum SHK1-27 and their antitumor activities. Chin. J. Med. Chem. 2007, 17, 148–154. [Google Scholar]
- Shao, C.; Wang, C.; Wei, M.; Li, S.; She, Z.; Gu, Y.; Lin, Y. Structural and spectral assignments of six anthraquinone derivatives from the mangrove fungus (ZSUH-36). Magn. Reson. Chem. 2008, 46, 886–889. [Google Scholar] [CrossRef]
- Kim, K.-W.; Kim, H.J.; Sohn, J.H.; Yim, J.H.; Kim, Y.-C.; Oh, H. Anti-neuroinflammatory effect of 6,8,1′-tri-O-methylaverantin, a metabolite from a marine-derived fungal strain Aspergillus sp., via upregulation of heme oxygenase-1 in lipopolysaccharide-activated microglia. Neurochem. Int. 2018, 113, 8–22. [Google Scholar] [CrossRef]
- Li, J.L.; Jiang, X.; Liu, X.; He, C.; Di, Y.; Lu, S.; Huang, H.; Lin, B.; Wang, D.; Fan, B. Antibacterial anthraquinone dimers from marine derived fungus Aspergillus sp. Fitoterapia 2019, 133, 1–4. [Google Scholar] [CrossRef]
- Ze-Hong, W.; Dong, L.; Ying, X.; Jian-Liang, C.; Wen-Han, L. Antioxidant xanthones and anthraquinones isolated from a marine-derived fungus Aspergillus versicolor. Chin. J. Nat. Med. 2018, 16, 219–224. [Google Scholar] [CrossRef]
- Xiao, Z.E.; Lin, S.E.; She, Z.; Lu, Y.; Tan, Z.; Liu, Y. A new anthraquinone from mangrove endophytic fungus Aspergillus sp. 16-5C. Nat. Prod. Res. 2021, 1–6. [Google Scholar] [CrossRef]
- Huang, H.; Wang, F.; Luo, M.; Chen, Y.; Song, Y.; Zhang, W.; Zhang, S.; Ju, J. Halogenated anthraquinones from the marine-derived fungus Aspergillus sp. SCSIO F063. J. Nat. Prod. 2012, 75, 1346–1352. [Google Scholar] [CrossRef]
- Huang, Z.; Nong, X.; Ren, Z.; Wang, J.; Zhang, X.; Qi, S. Anti-HSV-1, antioxidant and antifouling phenolic compounds from the deep-sea-derived fungus Aspergillus versicolor SCSIO 41502. Bioorg. Med. Chem. Lett. 2017, 27, 787–791. [Google Scholar] [CrossRef]
- Tian, Y.-Q.; Lin, S.-T.; Kumaravel, K.; Zhou, H.; Wang, S.-Y.; Liu, Y.-H. Polyketide-derived metabolites from the sponge-derived fungus Aspergillus sp. F40. Phytochem. Lett. 2018, 27, 74–77. [Google Scholar] [CrossRef]
- Wang, W.; Zhu, T.; Tao, H.; Lu, Z.; Fang, Y.; Gu, Q.; Zhu, W. Two new cytotoxic quinone type compounds from the halotolerant fungus Aspergillus variecolor. J. Antibiot. 2007, 60, 603–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, F.Y.; Li, X.M.; Song, J.Y.; Li, C.S.; Wang, B.G. Anthraquinone derivatives and an orsellinic acid ester from the marine alga-derived endophytic fungus Eurotium cristatum EN-220. Helv. Chim. Acta 2014, 97, 973–978. [Google Scholar] [CrossRef]
- Xing, C.-P.; Wu, J.; Xia, J.-M.; Fan, S.-Q.; Yang, X.-W. Steroids and anthraquinones from the deep-sea-derived fungus Aspergillus nidulans MCCC 3A00050. Biochem. Syst. Ecol. 2019, 83, 103–105. [Google Scholar] [CrossRef]
- Song, F.; Ren, B.; Chen, C.; Yu, K.; Liu, X.; Zhang, Y.; Yang, N.; He, H.; Liu, X.; Dai, H. Three new sterigmatocystin analogues from marine-derived fungus Aspergillus versicolor MF359. Appl. Microbiol. Biotechnol. 2014, 98, 3753–3758. [Google Scholar] [CrossRef]
- Wu, Z.J.; Ouyang, M.A.; Su, R.K.; Guo, Y.X. Two new cerebrosides and anthraquinone derivatives from the marine fungus Aspergillus niger. Chin. J. Chem. 2008, 26, 759–764. [Google Scholar] [CrossRef]
- Ren, H.; Liu, W.-W. Nidurufin as a new cell cycle inhibitor from marine-derived fungus Penicillium flavidorsum SHK1-27. Arch. Pharm. Res. 2011, 34, 901–905. [Google Scholar] [CrossRef]
- Zhu, F.; Chen, G.; Chen, X.; Yuan, Y.; Huang, M.; Xiang, W.; Sun, H. Structural elucidation of three anthraquinones from a marine-derived mangrove endophytic fungus (isolate 1850). In Proceedings of the 2008 International Conference on BioMedical Engineering and Informatics, Sanya, China, 27–30 May 2008; pp. 664–667. [Google Scholar] [CrossRef]
- Hu, J.; Li, Z.; Gao, J.; He, H.; Dai, H.; Xia, X.; Liu, C.; Zhang, L.; Song, F. New diketopiperazines from a marine-derived fungus strain Aspergillus versicolor MF180151. Mar. Drugs 2019, 17, 262. [Google Scholar] [CrossRef] [Green Version]
- Luo, X.-W.; Lu, H.-M.; Chen, X.-Q.; Zhou, X.-F.; Gao, C.-H.; Liu, Y.-H. Secondary metabolites and their biological activities from the sponge derived fungus Aspergillus versicolor. Chem. Nat. Compd. 2020, 56, 716–719. [Google Scholar] [CrossRef]
- Chen, M.; Shao, C.-L.; Kong, C.-J.; She, Z.-G.; Wang, C.-Y. A new anthraquinone derivative from a gorgonian-derived fungus Aspergillus sp. Chem. Nat. Compd. 2014, 50, 617–620. [Google Scholar] [CrossRef]
- Yuan, X.-L.; Wang, X.-F.; Xu, K.; Li, W.; Chen, D.; Zhang, P. Characterization of a new insecticidal anthraquinone derivative from an endophyte of Acremonium vitellinum against Helicoverpa armigera. J. Agric. Food Chem. 2020, 68, 11480–11487. [Google Scholar] [CrossRef]
- Song, X.; Tu, R.; Mei, X.; Wu, S.; Lan, B.; Zhang, L.; Luo, X.; Liu, J.; Luo, M. A mycophenolic acid derivative from the fungus Penicillium sp. SCSIO sof101. Nat. Prod. Res. 2020, 34, 1206–1212. [Google Scholar] [CrossRef] [PubMed]
- Li, D.-L.; Li, X.-M.; Wang, B.-G. Natural anthraquinone derivatives from a marine mangrove plant-derived endophytic fungus Eurotium rubrum: Structural elucidation and DPPH radical scavenging activity. J. Microbiol. Biotechnol. 2009, 19, 675–680. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.-L.; Wu, Y.-C.; Cheng, F.; Guo, Z.-Y.; Chen, J.-F. Anti-PRRSV effect and mechanism of tetrahydroaltersolanol C in vitro. J. Asian Nat. Prod. Res. 2016, 18, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Xie, G.E.; Zhu, X.; Li, Q.; Gu, M.; He, Z.; Wu, J.; Li, J.; Lin, Y.; Li, M.; She, Z.; et al. SZ-685C, a marine anthraquinone, is a potent inducer of apoptosis with anticancer activity by suppression of the Akt/FOXO pathway. Br. J. Pharmacol. 2010, 159, 689–697. [Google Scholar] [CrossRef] [Green Version]
- Ge, X.; Sun, C.; Feng, Y.; Wang, L.; Peng, J.; Che, Q.; Gu, Q.; Zhu, T.; Li, D.; Zhang, G. Anthraquinone derivatives from a marine-derived fungus Sporendonema casei HDN16-802. Mar. Drugs 2019, 17, 334. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Nakazawa, T.; Ukai, K.; Kobayashi, H.; Mangindaan, R.E.; Wewengkang, D.S.; Rotinsulu, H.; Namikoshi, M. Tetrahydrobostrycin and 1-deoxytetrahydrobostrycin, two new hexahydroanthrone derivatives, from a marine-derived fungus Aspergillus sp. J. Antibiot. 2008, 61, 415–419. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Wang, J.; Gao, Y.; Lin, H.; Du, L.; Yang, S.; Long, S.; She, Z.; Cai, X.; Zhou, S. The anthracenedione compound bostrycin induces mitochondria-mediated apoptosis in the yeast Saccharomyces cerevisiae. FEMS Yeast Res. 2010, 10, 297–308. [Google Scholar] [CrossRef] [Green Version]
- Xia, X.; Li, Q.; Li, J.; Shao, C.; Zhang, J.; Zhang, Y.; Liu, X.; Lin, Y.; Liu, C.; She, Z. Two new derivatives of griseofulvin from the mangrove endophytic fungus Nigrospora sp. (strain No. 1403) from Kandelia candel (L.) Druce. Planta Med. 2011, 77, 1735–1738. [Google Scholar] [CrossRef]
- Zhang, J.-Y.; Wu, H.-Y.; Xia, X.-K.; Liang, Y.-J.; Yan, Y.-Y.; She, Z.-G.; Lin, Y.-C.; Fu, L.-W. Anthracenedione derivative 1403P-3 induces apoptosis in KB and KBv200 cells via reactive oxygen species-independent mitochondrial pathway and death receptor pathway. Cancer Biol. Ther. 2007, 6, 1409–1417. [Google Scholar] [CrossRef] [Green Version]
- Du, L.; Zhu, T.; Fang, Y.; Liu, H.; Gu, Q.; Zhu, W. Aspergiolide A, a novel anthraquinone derivative with naphtho [1,2,3-de] chromene-2,7-dione skeleton isolated from a marine-derived fungus Aspergillus glaucus. Tetrahedron 2007, 63, 1085–1088. [Google Scholar] [CrossRef]
- Du, L.; Ai, J.; Li, D.; Zhu, T.; Wang, Y.; Knauer, M.; Bruhn, T.; Liu, H.; Geng, M.; Gu, Q. Aspergiolides C and D: Spirocyclic aromatic polyketides with potent protein kinase c-Met inhibitory effects. Chem. Eur. J. 2011, 17, 1319–1326. [Google Scholar] [CrossRef]
- Zhong, W.; Wang, J.; Wei, X.; Chen, Y.; Fu, T.; Xiang, Y.; Huang, X.; Tian, X.; Xiao, Z.; Zhang, W. Variecolortins A–C, three pairs of spirocyclic diketopiperazine enantiomers from the marine-derived fungus Eurotium sp. SCSIO F452. Org. Lett. 2018, 20, 4593–4596. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-L.; Li, X.-M.; Li, X.; Wang, C.-Y.; Liu, H.; Kassack, M.U.; Meng, L.-H.; Wang, B.-G. Antioxidant hydroanthraquinones from the marine algal-derived endophytic fungus Talaromyces islandicus EN-501. J. Nat. Prod. 2017, 80, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Cai, X.; Pan, J.; Gao, J.; Li, J.; Yuan, J.; Fu, L.; She, Z.; Lin, Y. Structure elucidation and NMR assignments for three anthraquinone derivatives from the marine fungus Fusarium sp. (No. ZH-210). Magn. Reson. Chem. 2009, 47, 362–365. [Google Scholar] [CrossRef]
- Huang, X.; Sun, X.; Lin, S.E.; Xiao, Z.E.; Li, H.; Bo, D.; She, Z. Xylanthraquinone, a new anthraquinone from the fungus Xylaria sp. 2508 from the South China Sea. Nat. Prod. Res. 2014, 28, 111–114. [Google Scholar] [CrossRef]
- Van Wagoner, R.M.; Mantle, P.G.; Wright, J.L. Biosynthesis of scorpinone, a 2-azaanthraquinone from Amorosia littoralis, a fungus from marine sediment. J. Nat. Prod. 2008, 71, 426–430. [Google Scholar] [CrossRef]
- Mantle, P.G.; Hawksworth, D.L.; Pazoutova, S.; Collinson, L.M.; Rassing, B.R. Amorosia littoralis gen. sp. nov., a new genus and species name for the scorpinone and caffeine-producing hyphomycete from the littoral zone in The Bahamas. Mycol. Res. 2006, 110, 1371–1378. [Google Scholar] [CrossRef]
- Miljkovic, A.; Mantle, P.G.; Williams, D.J.; Rassing, B. Scorpinone: A new natural azaanthraquinone produced by a Bispora-like tropical fungus. J. Nat. Prod. 2001, 64, 1251–1253. [Google Scholar] [CrossRef]
- Deng, C.-M.; Liu, S.-X.; Huang, C.-H.; Pang, J.-Y.; Lin, Y.-C. Secondary metabolites of a mangrove endophytic fungus Aspergillus terreus (No. GX7-3B) from the South China Sea. Mar. Drugs 2013, 11, 2616–2624. [Google Scholar] [CrossRef] [Green Version]
- Hwang, J.-Y.; Park, S.C.; Byun, W.S.; Oh, D.-C.; Lee, S.K.; Oh, K.-B.; Shin, J. Bioactive bianthraquinones and meroterpenoids from a marine-derived Stemphylium sp. fungus. Mar. Drugs 2020, 18, 436. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Wiese, J.; Wenzel-Storjohann, A.; Malien, S.; Schmaljohann, R.; Imhoff, J.F. Engyodontochones, antibiotic polyketides from the marine fungus Engyodontium album strain LF069. Chem. Eur. J. 2016, 22, 7452–7462. [Google Scholar] [CrossRef] [PubMed]
- Martins, T.; Schinke, C.; Queiroz, S.C.; de C Braga, P.A.; Silva, F.S.; Melo, I.S.; Reyes, F.G. Role of bioactive metabolites from Acremonium camptosporum associated with the marine sponge Aplysina fulva. Chemosphere 2021, 274, 129753. [Google Scholar] [CrossRef]
- Ayers, S.; Graf, T.N.; Adcock, A.F.; Kroll, D.J.; Shen, Q.; Swanson, S.M.; Matthew, S.; Carcache de Blanco, E.J.; Wani, M.C.; Darveaux, B.A. Cytotoxic xanthone–anthraquinone heterodimers from an unidentified fungus of the order Hypocreales (MSX 17022). J. Antibiot. 2012, 65, 3–8. [Google Scholar] [CrossRef]
- Wang, J.; Nong, X.-H.; Zhang, X.-Y.; Xu, X.-Y.; Amin, M.; Qi, S.-H. Screening of anti-biofilm compounds from marine-derived fungi and the effects of secalonic acid D on Staphylococcus aureus biofilm. J. Microbiol. Biotechnol. 2017, 27, 1078–1089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, A.; Islam, M.H.; Fatima, N.; Upadhyay, T.K.; Khan, M.K.A.; Dwivedi, U.N.; Sharma, R. Elucidation of marine fungi derived anthraquinones as mycobacterial mycolic acid synthesis inhibitors: An in silico approach. Mol. Biol. Rep. 2019, 46, 1715–1725. [Google Scholar] [CrossRef]
- Wang, C.; Wang, J.; Huang, Y.; Chen, H.; Li, Y.; Zhong, L.; Chen, Y.; Chen, S.; Wang, J.; Kang, J. Anti-mycobacterial activity of marine fungus-derived 4-deoxybostrycin and nigrosporin. Molecules 2013, 18, 1728–1740. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.-Y.; Tao, L.-Y.; Liang, Y.-J.; Chen, L.-M.; Mi, Y.-J.; Zheng, L.-S.; Wang, F.; She, Z.-G.; Lin, Y.-C.; To, K.K.W. Anthracenedione derivatives as anticancer agents isolated from secondary metabolites of the mangrove endophytic fungi. Mar. Drugs 2010, 8, 1469–1481. [Google Scholar] [CrossRef]
- Chen, C.H.; Xiao, W.W.; Jiang, X.B.; Wang, J.W.; Mao, Z.G.; Lei, N.; Fan, X.; Song, B.B.; Liao, C.X.; Wang, H.J. A novel marine drug, SZ–685C, induces apoptosis of MMQ pituitary tumor cells by downregulating miR–200c. Curr. Med. Chem. 2013, 20, 2145–2154. [Google Scholar] [CrossRef]
- Wang, X.; Tan, T.; Mao, Z.-G.; Lei, N.; Wang, Z.-M.; Hu, B.; Chen, Z.-Y.; She, Z.-G.; Zhu, Y.-H.; Wang, H.-J. The marine metabolite SZ-685C induces apoptosis in primary human nonfunctioning pituitary adenoma cells by inhibition of the Akt pathway in vitro. Mar. Drugs 2015, 13, 1569–1580. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Zhu, X.; Zhong, L.-L.; Yang, B.; Li, J.; Wu, J.-H.; Chen, S.-P.; Lin, Y.-C.; Long, Y.; She, Z.-G. Synthesis and antitumor activities of derivatives of the marine mangrove fungal metabolite deoxybostrycin. Mar. Drugs 2012, 10, 2715–2728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.; Jin, H.; Song, B.; Zhu, X.; Zhao, H.; Cai, J.; Lu, Y.; Chen, B.; Lin, Y. The cytotoxicity and anticancer mechanisms of alterporriol L, a marine bianthraquinone, against MCF-7 human breast cancer cells. Appl. Microbiol. Biotechnol. 2012, 93, 777–785. [Google Scholar] [CrossRef] [PubMed]



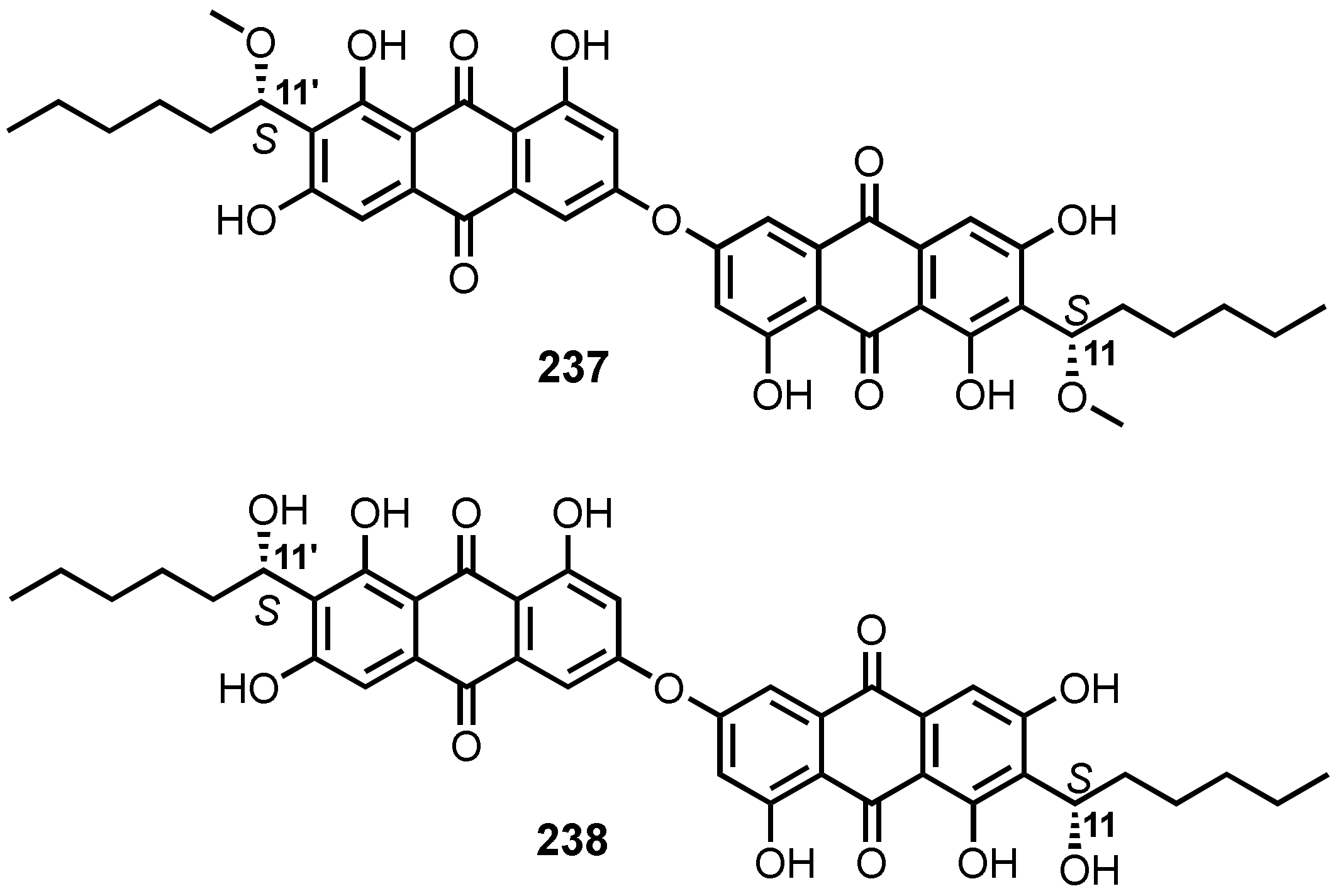
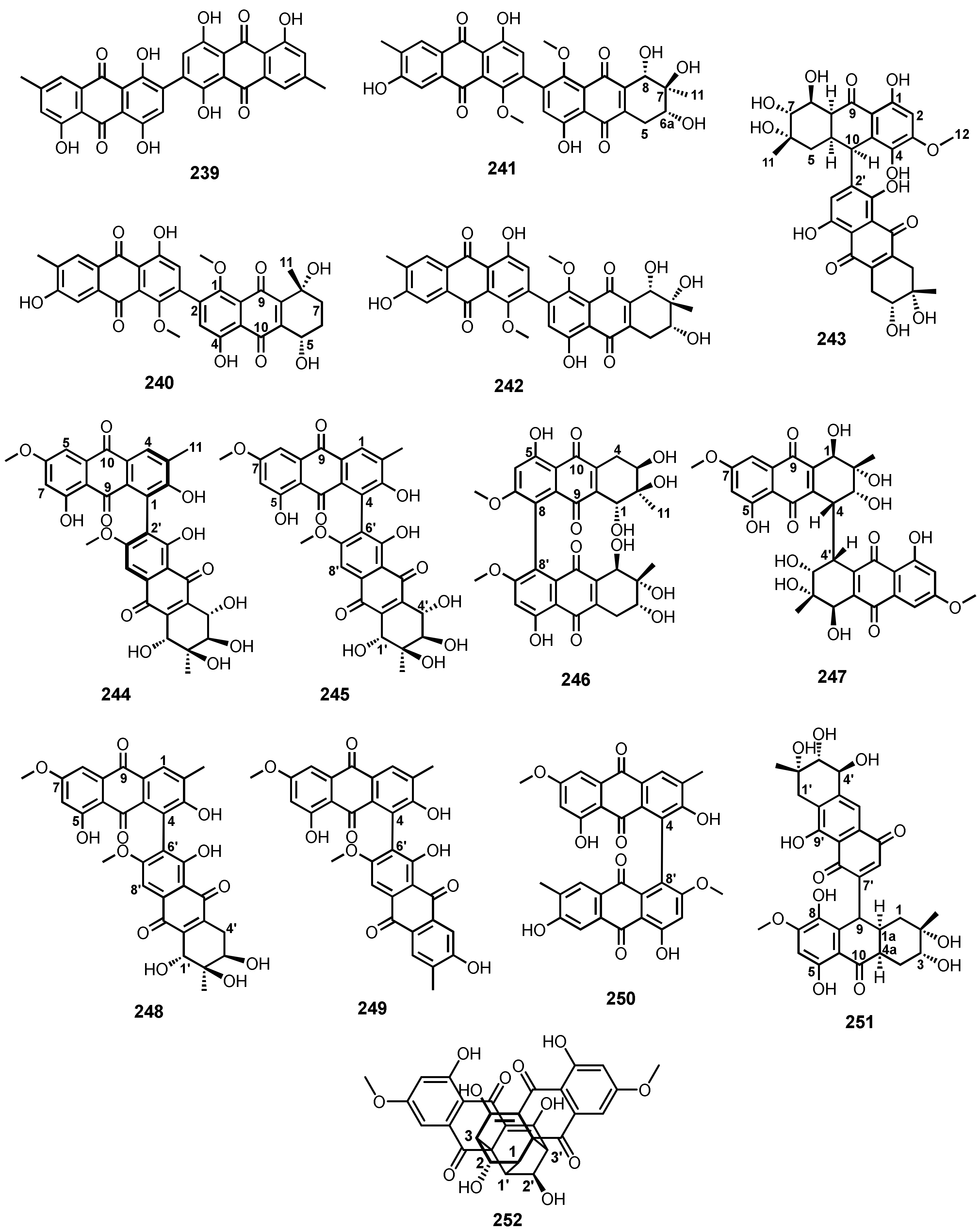
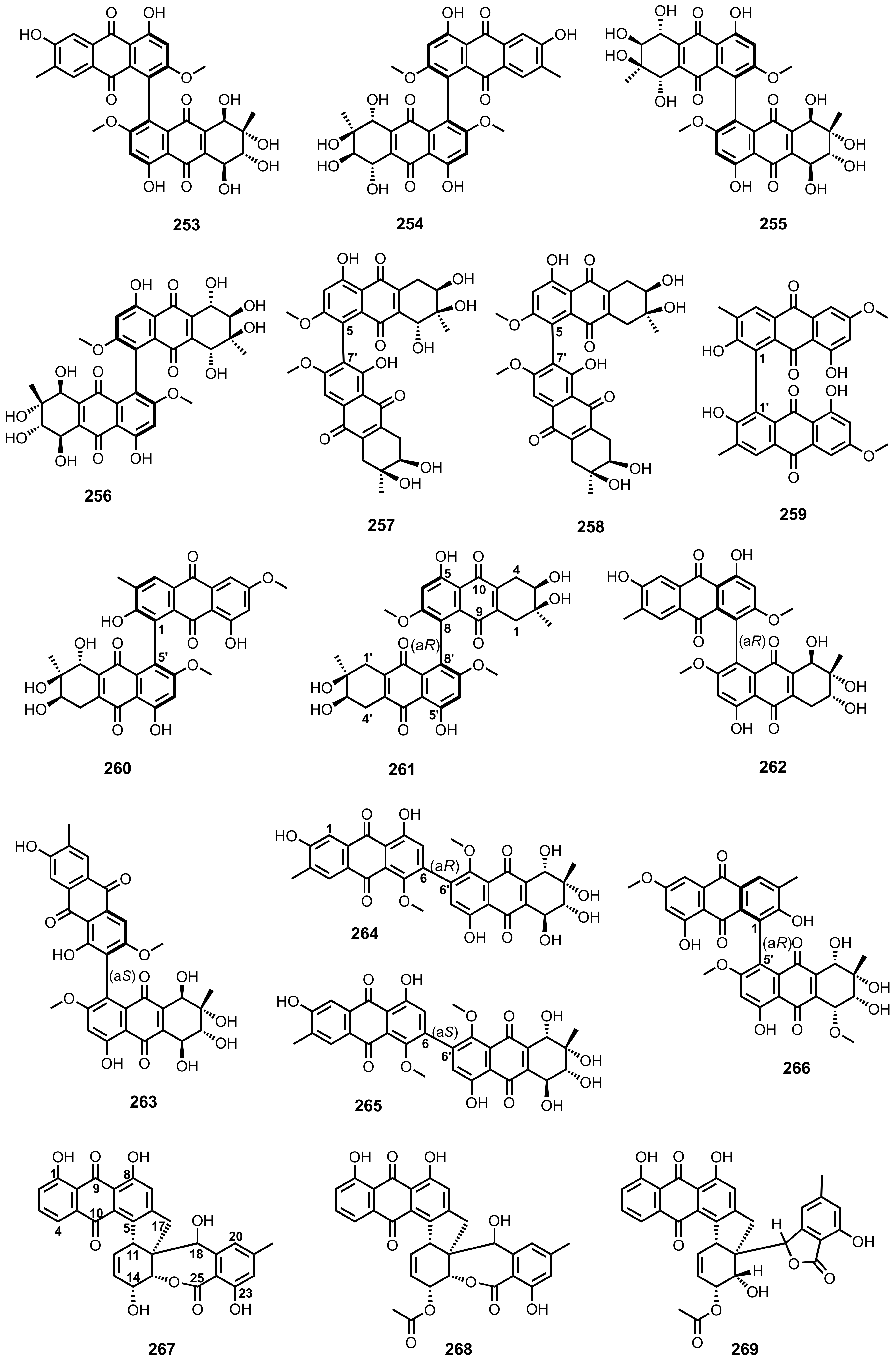
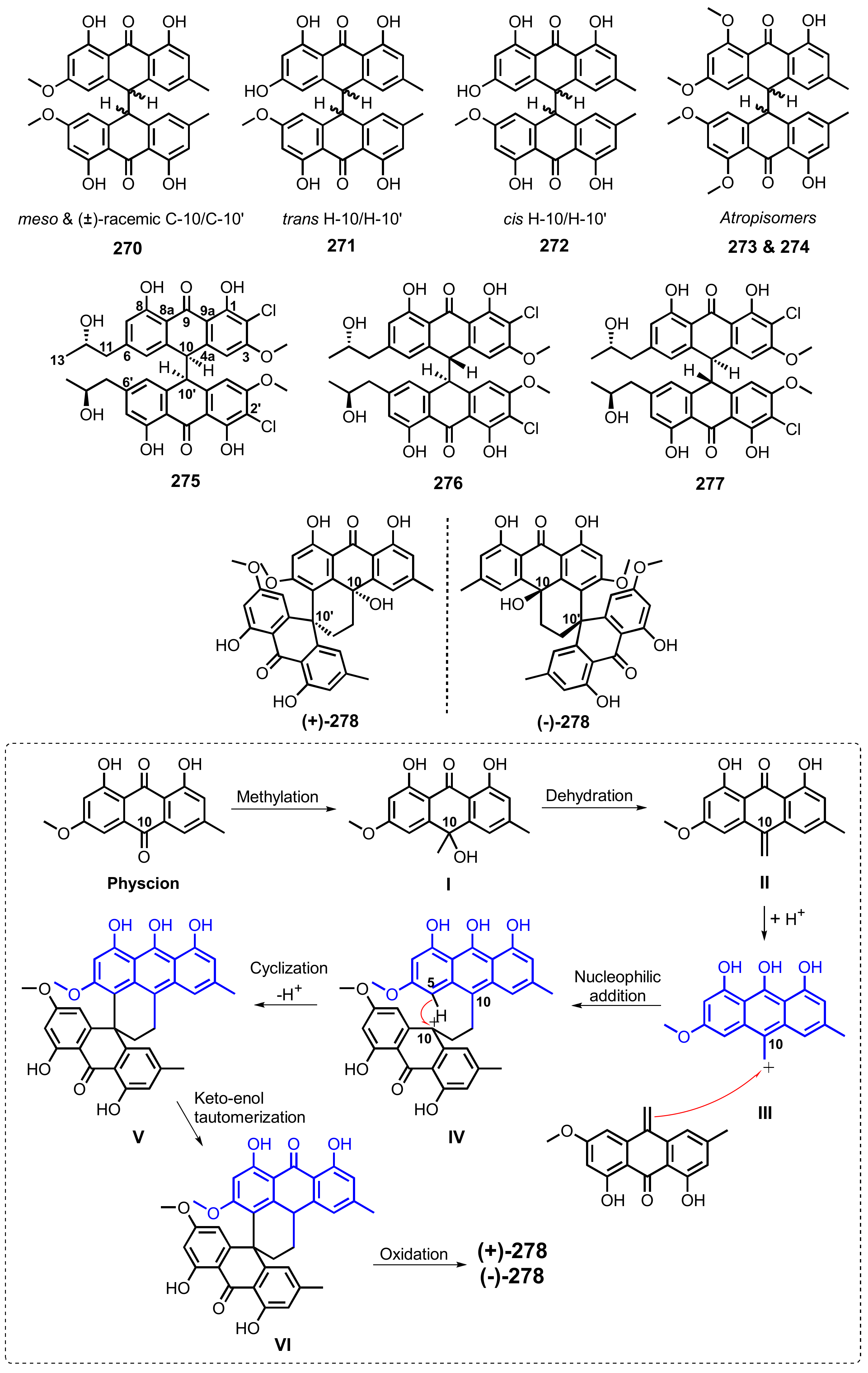
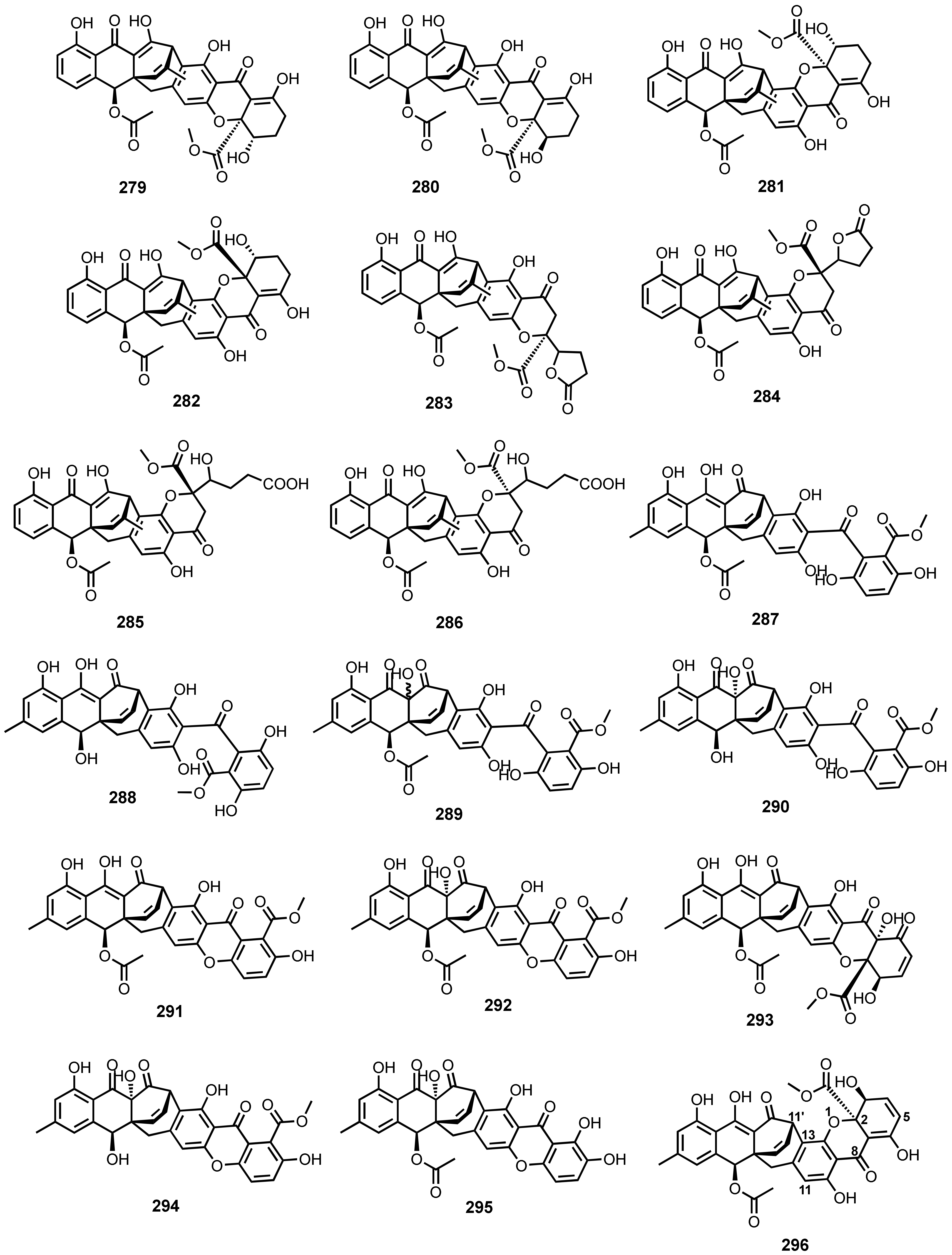
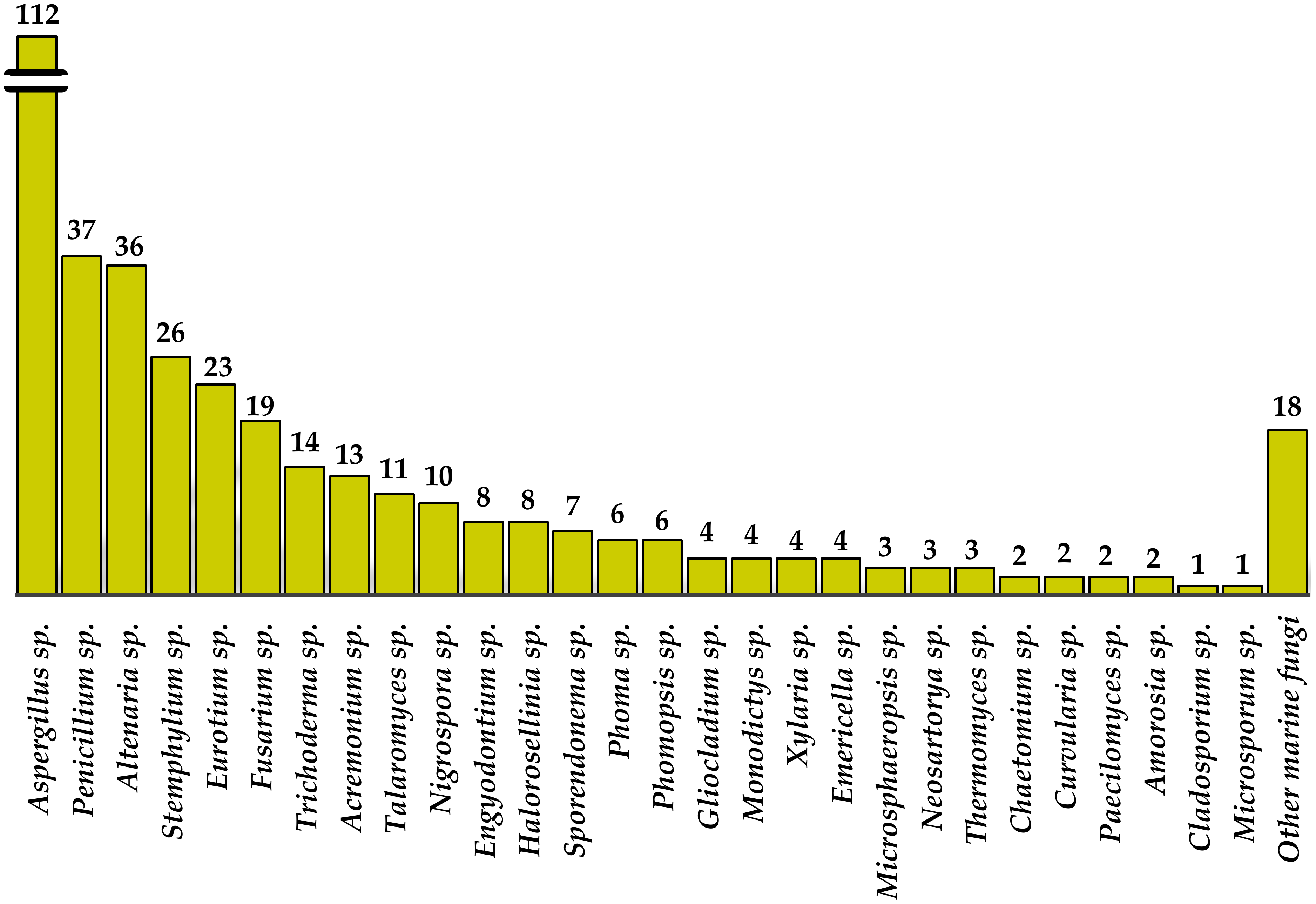

{kind=link}
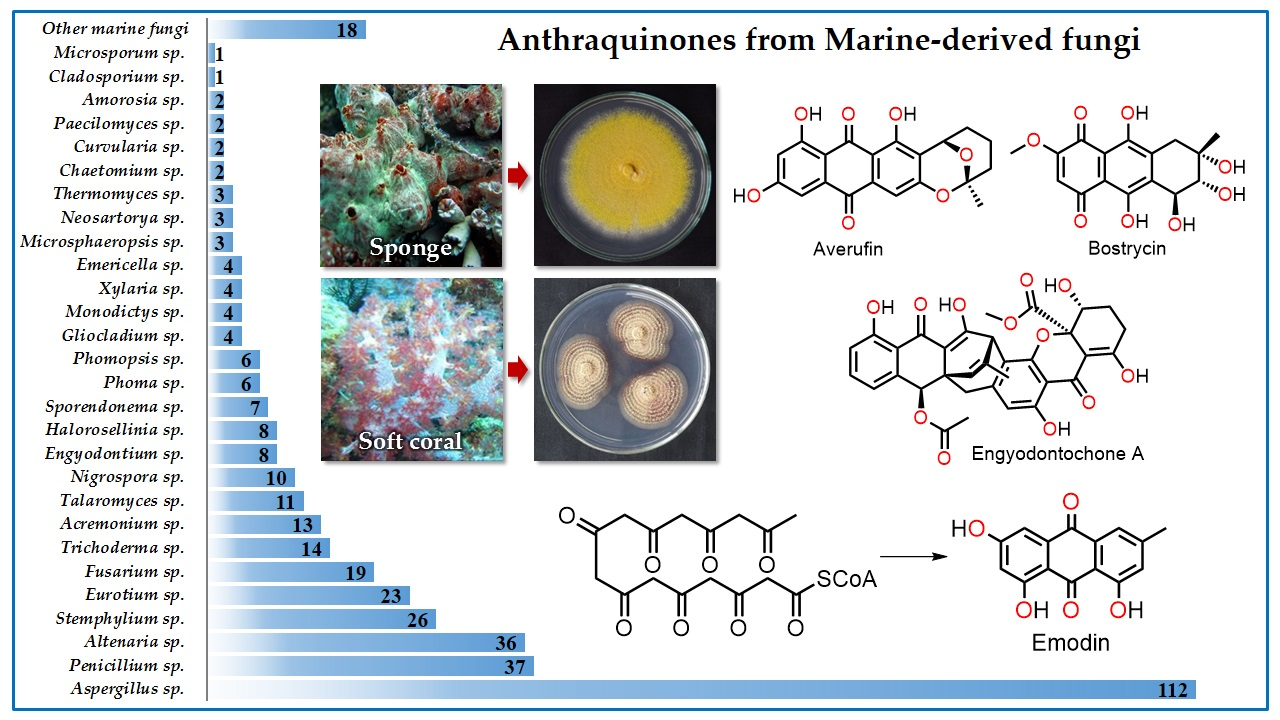
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
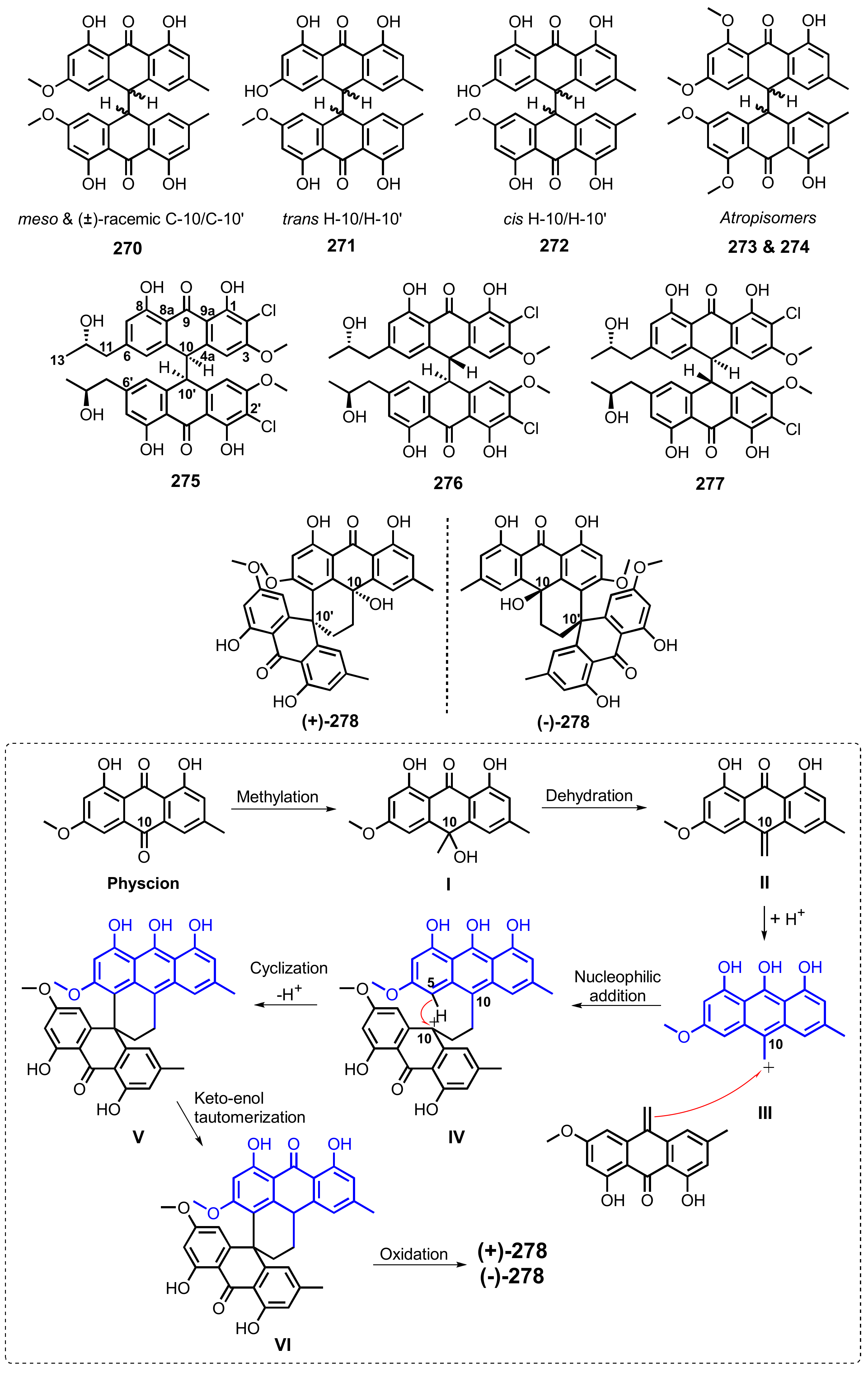{kind=link}
{kind=link}
{kind=link}
| Compound | Fungus Species/Strain No. | Source of Marine-Derived Fungi | Bioactivity | Ref. |
|---|---|---|---|---|
| Emodin (1) | Aspergillus candidus KUFA0062 | -Marine sponge Epipolasis sp. | - | [16] |
| A. flavipes HN4-13 | -Coastal sediment. | -Non-competitive α-glucosidase inhibitor. | [18] | |
| Aspergillus sp. LS57 | -Marine sponge Haliclona sp. | - | [19] | |
| A. glaucus HB1-19 | -Marine sediment. | -Antibacterial and cytotoxic activities. | [22] | |
| A. tritici SP2-8-1 | -Coral Galaxea fascicularis. | -Antibacterial and cytotoxic activities. | [28] | |
| A. versicolor | -Green alga Halimeda opuntia. | -Antiviral activity; inhibition of human trypsin activity. | [34] | |
| A. versicolor | -Deep-sea sediment. | -Antibacterial activity. | [35] | |
| Penicillium oxalicum 2HL-M-6 | -Sea mud. | - | [36] | |
| P. ochrochloron | -Sea mud. | - | [17] | |
| P. citrinum PSU-F51 | -Gorgonian Sea fan (Annella sp.) | -Antifouling activity. | [37] | |
| Penicillium sp. SCSGAF0023 | -Gorgonian coral. | -Antifouling activity. | [39] | |
| Eurotium rubrum | -Inner tissue of semi-mangrove plant Hibiscus tiliaceus. | -Antibacterial, reduction of biofilm formation and cytotoxic activities. | [47] | |
| E. chevalieri KUFA0006 | -Inner twig of mangrove plant Rhizophora mucronata Poir. | -Antibacterial activity. | [31] | |
| Talaromyces stipitatus KUFA0207 | -Marine sponge Stylissa flabelliformis. | -Anti-obesity activity. | [33] | |
| Paecilomyces sp. (Tree1-7) | -Mangrove saprophytic bark. | -Antibacterial activity. | [42] | |
| Trichoderma harzianum (XS-20090075) | -Inner tissue of soft coral. | -Antibacterial activity. | [55] | |
| Trichoderma sp. (H-1) | -Sea cucumber. | -Antibacterial activity. | [43] | |
| Gliocladium sp. T31 | -Marine lichen. | - | [38] | |
| G. catenulatum T31 | -Marine sediment. | -Cytotoxic activity. | [65] | |
| Monodictys sp. | -Sea urchin Anthocidaris crassispina. | -Antitumor activity. | [60] | |
| - | - | -Antibiofilm formation. | [128] | |
| Questin (MT-1; 2) | Aspergillus flavipes HN4-13 | -Coastal sediment. | - | [18] |
| A. terreus DTO 403-C9 | -Leaves of an unidentified mangrove tree. | - | [20] | |
| A. glaucus HB1-19 | -Marine sediment. | - | [22] | |
| Penicillium citrinum HL-5126 | -Mangrove Bruguiera sexangula var. rhynchopetala. | - | [41] | |
| Eurotium chevalieri KUFA0006 | -Inner twig of mangrove plant Rhizophora mucronata Poir. | - | [31] | |
| Eurotium sp. SCSIO F452 | -Marine sediment. | - | [70] | |
| E. rubrum | -Inner tissue of mangrove plant Hibiscus tiliaceus. | -DPPH• radicals scavenging activity. | [106] | |
| 1,2,5-Trihydroxy-7-methyl-9,10-anthraquinone (3) | Aspergillus terreus DTO 403-C9 | -Leaves of an unidentified mangrove tree. | - | [20] |
| 1-Methyl emodin (4) | A. europaeus WZXY-SX-4-1 | -Marine sponge Xestospongia testudinaria. | -Down-regulation of NF-κB. | [21] |
| A. vesicolor | -Green alga Halimeda opuntia. | -Antiviral activity; inhibition of human trypsin activity. | [34] | |
| Dermolutein (5) | A. europaeus WZXY-SX-4-1 | -Marine sponge X. testudinaria. | -Down-regulation of NF-κB. | [21] |
| Physcion (or parietin; 6) | A. glaucus HB1-19 | -Marine sediment. | - | [22] |
| A. wentii EN-48 | -Brown alga Sargassum sp. | -DPPH• radicals scavenging activity. | [23] | |
| Penicillium sp. ZZ901 | -Wild bivalve of Scapharca broughtonii (Schrenck). | -Anti-proliferative activity. | [27] | |
| Eurotium chevalieri MUT2316 | -Marine sponge Grantia compressa. | -Antifouling and antibacterial activities. | [53] | |
| E. chevalieri KUFA0006 | -Inner twig of mangrove plant Rhizophora mucronata Poir. | -Reduction of biofilm formation. | [31] | |
| Eurotium sp. SCSIO F452 | -Marine sediment. | - | [70] | |
| E. repens | -Marine sponge Suberites domuncula. | -Cytotoxicity against sex cells. | [24] | |
| E. cristatum | -Marine sponge Mycale sp. | - | [25] | |
| Altenaria sp. ZJ9-6B | -Mangrove tree Aegiceras corniculatum fruits. | - | [61] | |
| Chaetomium globosum | -Inner tissue of the marine red alga Polysiphonia urceolata. | - | [32] | |
| Microsporum sp. MFS-YL | -Marine red alga Lomentaria catenata. | -Anti-proliferative and cytotoxic activities. | [26] | |
| Catenarin (7) | Aspergillus glaucus HB1-19 | -Marine sediment. | - | [22] |
| Eurotium sp. SCSIO F452 | -Marine sediment. | - | [70] | |
| Rubrocristin (8) | A. glaucus HB1-19 | -Marine sediment. | - | [22] |
| 3-Hydroxy-1,2,5,6-tetramethoxy anthracene-9,10-dione (9) | A. tritici SP2-8-1 | -Soft coral Galaxea fascicularis. | -Antibacterial activity. | [28] |
| 3-Hydroxy-2-hydroxymethyl-1-methoxy anthracene-9,10-dione (10) | A. tritici SP2-8-1 | -Soft coral G. fascicularis. | -Antibacterial and cytotoxic activities. | [28] |
| 1,2,3-Trimethoxy-7-hydroxy methylanthracene-9,10-dione (11) | A. tritici SP2-8-1 | -Soft coral G. fascicularis. | -Antibacterial and cytotoxic activities. | [28] |
| 1,5-Dihydroxy-3-methoxy-7-methylanthraquinone (12) | A. wentii (pt-1), A. ustus (cf-42), A. versicolor (dl-29 and pt-20) | - | -Algicidal activity. | [29] |
| 1,3,5-Trihydroxy-7-methylanthraquinone (13) | A. wentii (pt-1), A. ustus (cf-42), A. versicolor (dl-29 and pt-20) | - | -Algicidal activity. | [29] |
| 5-Hydroxy-2,4-dimethoxy-7-methylanthraquinone (or emodin-6,8-dimethyl ether; 14) | A. wentii (pt-1), A. ustus (cf-42), A. versicolor (dl-29 and pt-20) | - | -Algicidal activity. | [29] |
| A. wentii EN-48 | -Brown alga Sargassum sp. | - | [30] | |
| Emericella sp. SCSIO 05240 | -Marine sediment. | - | [78] | |
| Questinol (15) | Eurotium chevalieri KUFA0006 | -Inner twig of mangrove plant Rhizophora mucronata Poir. | -Reduction of biofilm formation. | [31] |
| Talaromyces stipitatus KUFA0207 | -Marine sponge Stylissa flabelliformis. | -Anti-obesity activity. | [33] | |
| Erythroglaucin (16) | Chaetomium globosum | -Inner tissue of the marine red alga Polysiphonia urceolata. | -DPPH• radicals scavenging activity. | [32] |
| Fallacinol (17) | Talaromyces stipitatus KUFA0207 | -Marine sponge Stylissa flabelliformis. | -Anti-obesity activity. | [33] |
| Evariquinone (18) | A. vesicolor | -Green alga Halimeda opuntia. | - | [34] |
| 7-Hydroxyemodin-6,8-dimethyl ether (19) | A. vesicolor | -Green alga H. opuntia | - | [34] |
| Emericella sp. SCSIO 05240 | -Marine sediment. | - | [78] | |
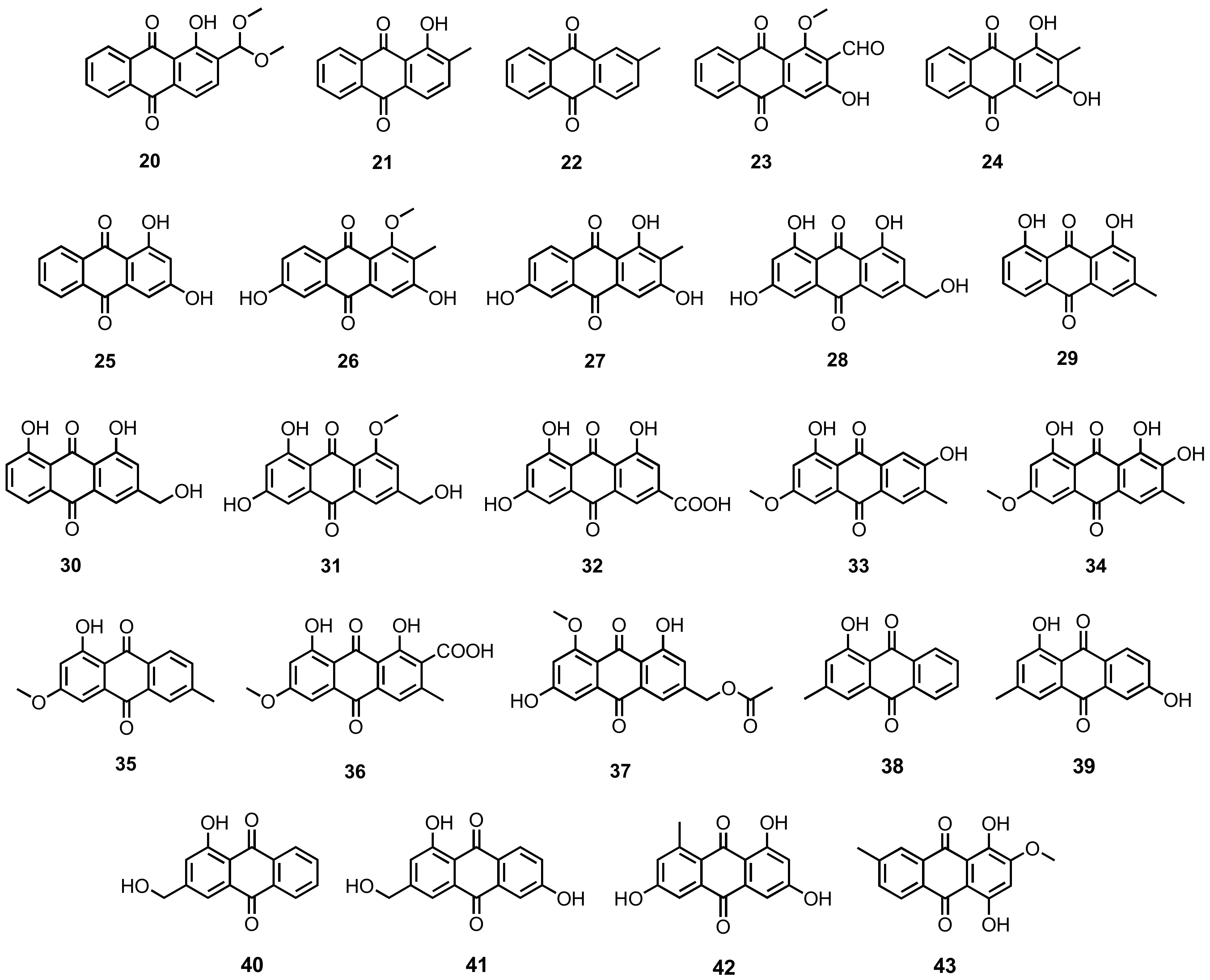
| 2-(dimethoxymethyl)-1-hydroxy anthracene-9,10-dione (20) | A. versicolor | -Deep-sea sediment. | -Antibacterial activity. | [35] |
| 1-Hydroxy-2-methylanthracene-9,10-dione (21) | A. versicolor | -Deep-sea sediment. | - | [35] |
| 2-Methylanthracene-9,10-dione (22) | A. versicolor | -Deep-sea sediment. | - | [35] |
| Damnacanthal (23) | A. versicolor | -Deep-sea sediment. | - | [35] |
| Rubiadin (24) | A. versicolor | -Deep-sea sediment. | - | [35] |
| Xanthopurpurin (25) | A. versicolor | -Deep-sea sediment. | - | [35] |
| Rubianthraquinone (26) | A. versicolor | -Deep-sea sediment. | - | [35] |
| 6-Hydroxyrubiadin (27) | A. versicolor | -Deep-sea sediment. | - | [35] |
| Citreorosein (or ω-hydroxyemodin; 28) | Penicillium oxalicum 2HL-M-6 | -Sea mud. | - | [36] |
| P. citrinum PSU-F51 | -Gorgonian Sea fan (Annella sp.) | - | [37] | |
| Penicillium sp. SCSGAF0023 | -Gorgonian coral. | -Antifouling activity. | [39] | |
| P. citrinum HL-5126 | -Mangrove Bruguiera sexangula var. rhynchopetala. | -Antibacterial activity. | [41] | |
| Talaromyces stipitatus KUFA0207 | -Marine sponge Stylissa flabelliformis. | -Anti-obesity activity. | [33] | |
| Emericella sp. SCSIO 05240 | -Marine sediment. | -Antibacterial activity. | [78] | |
| Fusarium equiseti | -Marine brown alga Padina pavonica. | -Antiviral activity; inhibition of human trypsin activity. | [40] | |
| Gliocladium sp. T31 | -Marine lichen. | - | [38] | |
| G. catenulatum T31 | -Marine sediment. | -Anti-tumor activity. | [65] | |
| - | - | -Anti-biofilm formation. | [135] | |
| Chrysophanol (or chrysophanic acid; 29) | Aspergillus candidus KUFA0062 | -Marine sponge Epipolasis sp. | -Anti-biofilm formation. | [16] |
| Penicillium oxalicum 2HL-M-6 | -Sea mud. | - | [36] | |
| P. citrinum PSU-F51 | -Gorgonian Sea fan (Annella sp.) | - | [37] | |
| Paecilomyces sp. (Tree1-7) | -Mangrove saprophytic bark. | - | [42] | |
| Fusarium equiseti | -Marine brown alga Padina pavonica. | -Antiviral activity; inhibition of human trypsin activity. | [40] | |
| Trichoderma harzianum (XS-20090075) | -Inner tissue of soft coral. | -Anti-acetylcholinesterase activity. | [55] | |
| Trichoderma sp. (H-1) | -Sea cucumber. | -Antibacterial activity. | [43] | |
| Monodictys sp. | -Sea urchin Anthocidaris crassispina. | - | [60] | |
| Strain F-F-3C | -Unidentified marine red alga. | -Antibacterial activity. | [44] | |
| Aloe-emodin (30) | Penicillium oxalicum 2HL-M-6 | -Sea mud. | - | [36] |
| Carviolin (31) | Penicillium sp. strain F01V25 | -Marine alga Dictyosphaeria versluyii. | - | [45] |
| Emodic acid (32) | Penicillium sp. SCSIOsof101 | -Deep-sea sediment. | - | [46] |
| Eurotium rubrum | -Inner tissue of semi-mangrove plant Hibiscus tiliaceus. | - | [47] | |
| Macrosporin (33) | Penicillium sp. | -Soft coral Sarcophyton tortuosum. | - | [48] |
| Altenaria sp. ZJ-2008003 | -Soft coral Sarcophyton sp. | -Antibacterial activity. | [49] | |
| Stemphylium sp. 33231 | -Mangrove tree Bruguiera sexangula var. rhynchopetala. | -Antibacterial activity. | [50] | |
| S. lycopersici | -Inner tissue of gorgonian soft coral Dichotella gammacea. | - | [51] | |
| Phoma sp. L28 | -Mangrove plant Myoporum bontioides A. Gray. | -Anti-fungal activity. | [67] | |
| Phomopsis sp. PSU-MA214 | -Leaves of mangrove plant Rhizophora apiculata Griff. Ex T. Anderson. | - | [52] | |
| 1,7,8-Tri-hydroxy-3-methoxy-6-methyl anthraquinone (34) | Penicillium sp. | -Soft coral Sarcophyton tortuosum. | - | [48] |
| 1-Hydroxy-3-methoxy-6-methylanthraquinone (35) | Penicillium sp. | -Soft coral Sarcophyton tortuosum. | - | [48] |
| Phomopsis sp. PSU-MA214 | -Leaves of mangrove plant Rhizophora apiculata Griff. Ex T. Anderson. | - | [52] | |
| Cinnalutein (36) | Eurotium chevalieri MUT2316 | -Marine sponge Grantia compressa. | -Antifouling and algicidal activities. | [53] |
| Acetylquestinol (37) | E. chevalieri KUFA0006 | -Inner twig of mangrove plant Rhizophora mucronata Poir. | -Reduction of biofilm formation. | [31] |
| Neosartorya spinosa KUFA1047 | -Marine sponge Mycale sp. | - | [54] | |
| Pachybasin (38) | Trichoderma harzianum (XS20090075) | -Inner tissue of a soft coral. | -Anti-acetylcholinesterase activity. | [55] |
| Monodictys sp. | -Sea urchin Anthocidaris crassispina. | - | [60] | |
| Phomarin (39) | T. harzianum (XS-20090075) | -Inner tissue of a soft coral. | - | [55] |
| 1-Hydroxy-3-hydroxy methylanthraquinone (40) | T. harzianum (XS-20090075) | -Inner tissue of a soft coral. | -Antibacterial and cytotoxic activities. | [55] |
| ω-Hydroxydigitoemodin (41) | T. harzianum (XS-20090075) | - Inner tissue of a soft coral. | -Anti-acetylcholinesterase activity. | [55] |
| 1,3,6-Trihydroxy-8-methylanthraquinone (42) | Trichoderma sp. strain SCSIO41004 | -Marine sponge Callyspongia sp. | - | [56] |
| 1,4-Dihydroxy-2-methoxy-7-methylanthracene-9,10-dione (43) | Halorosellinia sp. (no. 1403) | -Estuarine. | - | [57] |
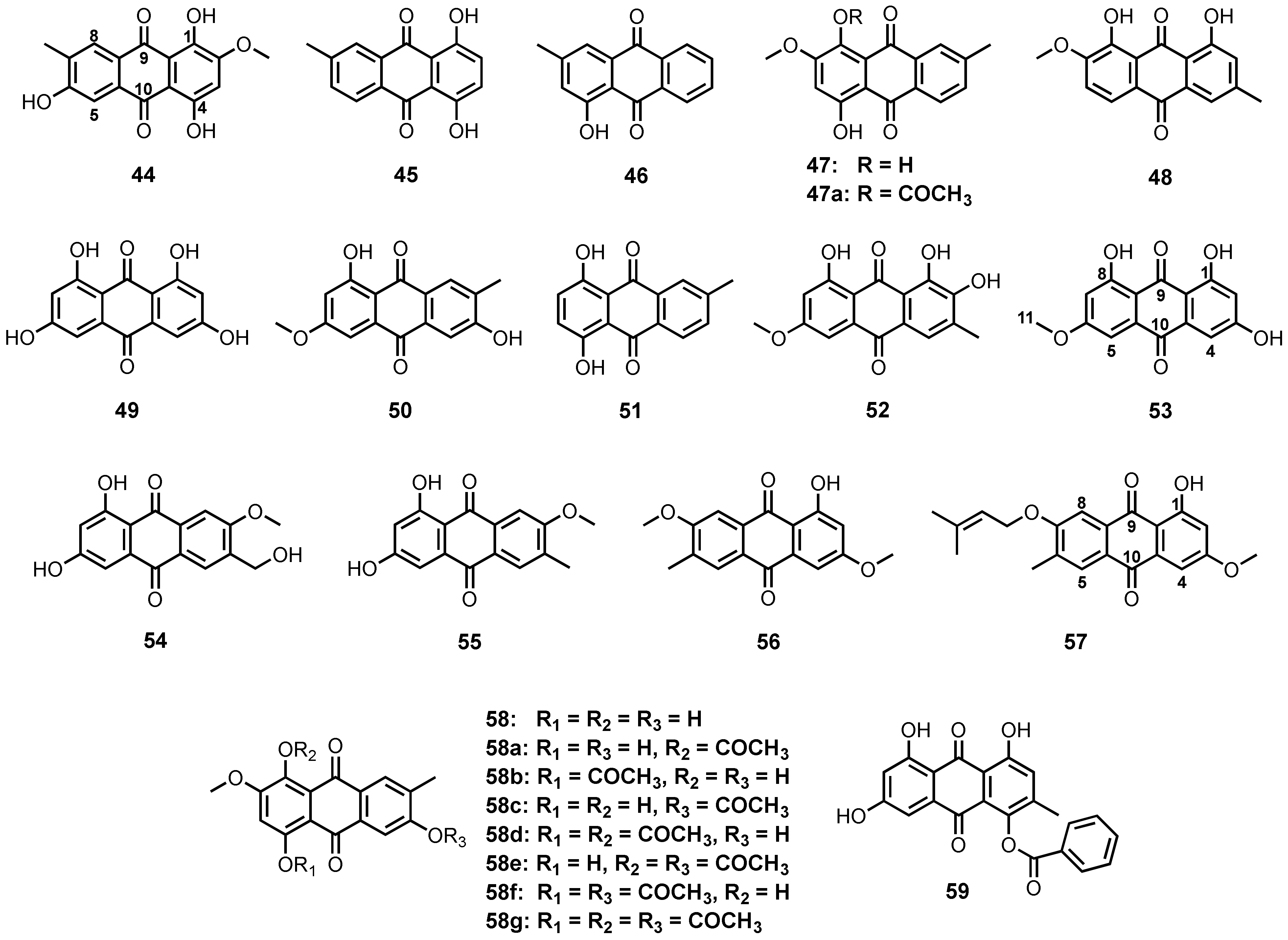
| 1,4,6-Trihydroxy-2-methoxy-7-methylanthracene-9,10-dione (44) | Halorosellinia sp. (no. 1403) | -Decayed Kandelia candel (L.) Druce. | - | [58] |
| Demethoxyaustrocortirubin (45) | Halorosellinia sp. (no. 1403) | -Decayed K. candel (L.) Druce. | - | [58] |
| Hydroxy-9,10-anthraquinone (46) | Halorosellinia sp. (no. 1403) | -Decayed Kandelia candel (L.) Druce. | -Cytotoxic activity. | [58,131] |
| Austrocortinin (47) | Altenaria sp. (SK11) | -Root of mangrove tree Excoecaria agallocha. | - | [62] |
| Fusarium sp. PSU-F14 | -Gorgonian sea fan. | -Antibacterial and cytotoxic activities. | [59] | |
| Nigrospora sp. ZJ-2010006 | -Unidentified sea anemone. | - | [68] | |
| Nigrospora sp. ZJ-2010006 | -Inner tissue of the zoathid Palythoa haddoni (GX-WZ-20100026). | -Antiviral activity. | [69] | |
| Halorosellinia sp. (no. 1403) | -Decayed Kandelia candel (L.) Druce. | - | [58] | |
| Monodictyquinone A (48) | Monodictys sp. | -Sea urchin Anthocidaris crassispina. | -Antibacterial activity. | [60] |
| Rheoemodin (49) | Talaromyces stipitatus KUFA0207 | -Marine sponge Stylissa flabelliformis. | -Antiobesity activity. | [33] |
| Marcrospin (50) | Altenaria sp. ZJ9-6B | -Mangrove tree Aegiceras corniculatum fruits. | - | [61] |
| 6-Methylquinizarin (51) | Altenaria sp. (SK11) | -Root of mangrove tree Excoecaria agallocha. | - | [62] |
| 6-O-Methylalaternin (52) | Altenaria tenuissima DFFSCS013 | -Marine sediment. | -Inhibition of human protein tyrosine phosphatases and inhibition of indoleamine 2,3-dioxygenase activity. | [63] |
| Lunatin (53) | Curvularia lunata | -Marine sponge Niphates olemda. | -Antibacterial activity. | [64] |
| Gliocladium catenulatum T31 | -Marine sediment. | -Anti-tumor activity. | [65] | |
| 1,3-Dihydroxy-6-hydroxymethyl-7-methoxyanthraquinone (54) | Thermomyces lanuginosus Tsikl KMM 4681 | -Marine sediment. | - | [66] |
| 1,3-Dihydroxy-6-methyl-7-methoxyanthraquinone (55) | T. lanuginosus Tsikl KMM 4681 | -Marine sediment. | -Cytotoxic activity. | [66] |
| 7-Methoxymacrosporin (56) | Phoma sp. L28 | -Mangrove plant Myoporum bontioides A. Gray. | -Antifungal activity. | [67] |
| 7-(γ,γ)-Dimethylallyloxymacrosporin (57) | Phoma sp. L28 | -Mangrove plant M. bontioides A. Gray. | -Antifungal activity. | [67] |
| 3,5,8-Tri-hydroxy-7-methoxy-2-methylanthracene-9,10-dione (58) | Nigrospora sp. ZJ-2010006 | -Unidentified sea anemone. | -Antibacterial activity. | [68] |
| Nigrospora sp. ZJ-2010006 | -Inner tissue of the zoathid Palythoa haddoni (GX-WZ-20100026). | -Antiviral activity. | [69] | |
| 1,6,8-Trihydroxy-4-benzoyloxy-3-methylanthraquinone (59) | Eurotium sp. SCSIO F452 | -Marine sediment. | - | [70] |
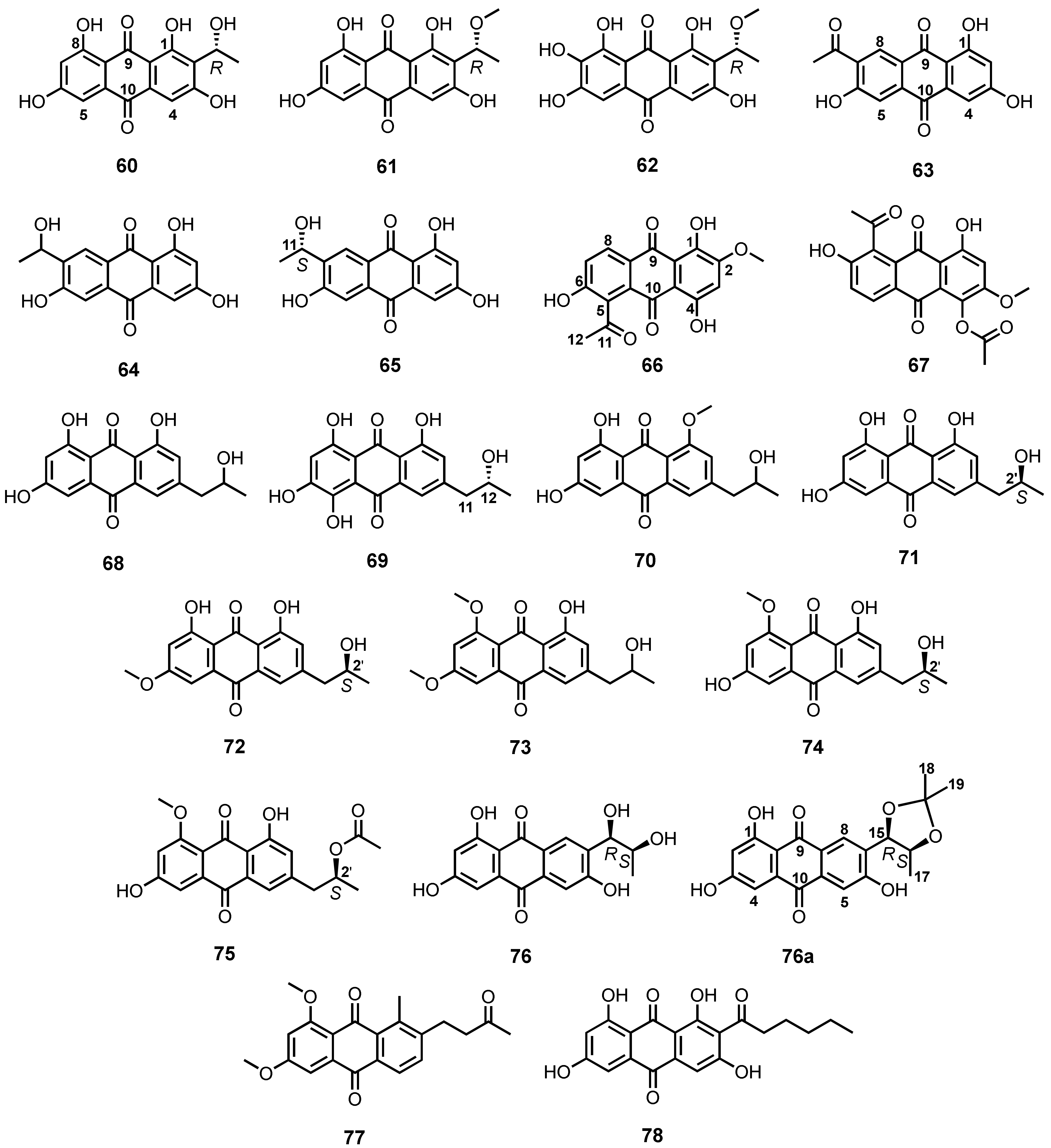
| 1,3,6,8-Tetrahydroxyanthraquinone analogues (60-62) | Microsphaeropsis sp. | -Marine sponge Aplysina aerophoba. | -Inhibition of protein kinases. | [71,72] |
| 7-Acetyl-1,3,6-trihydroxyanthracene-9,10-dione (63) | Trichoderma sp. strain SCSIO41004 | -Marine sponge Callyspongia sp. | - | [56] |
| Fusarium equiseti | -Intertidal marine plants. | -Antibacterial activity. | [73] | |
| ZSU-H85 (64) | Trichoderma sp. strain SCSIO41004 | -Marine sponge Callyspongia sp. | -Antiviral activity. | [56] |
| (11S)-1,3,6-Trihydroxy-7-(1-hydroxyethyl)anthracene-9,10-dione (65) | F. equiseti | -Intertidal marine plants. | -Antibacterial activity. | [73] |
| Cladosporium sp. HNWSW-1 | -Fresh roots of Ceriops tagal. | -Inhibition of α-glucosidase activity. | [74] | |
| 5-Acetyl-2-methoxy-1,4,6-trihydroxy-anthraquinone (66) | Fusarium sp. (no. b77) | -Costal environment. | - | [75] |
| 1-Acetoxy-5-acetyl-2-methoxy-4,6- trihydroxy-anthraquinone (67) | Fusarium sp. (no. b77) | -Costal environment. | - | [75] |
| Isorhodoptilometrin (68) | Penicillium oxalicum 2HL-M-6 | -Sea mud. | - | [36] |
| Penicillium sp. SCSGAF0023 | -Gorgonian coral. | -Antifouling activity. | [39] | |
| Gliocladium sp. T31 | -Marine lichen. | - | [38] | |
| G. catenulatum T31 | -Marine sediment. | -Antitumor activity. | [65] | |
| (−)-2′R-1-Hydroxyisorhodopilometrin (69) | Penicillium sp. OUCMDZ-4736 | -Mangrove roots of Acanthus ilicifolius. | -Antiviral activity. | [76] |
| Isorodoptilometrin-1-methyl ether (70) | Aspergillus vesicolor | -Green alga Halimeda opuntia. | -Antibacterial activity. | [34] |
| (+)-2′S-isorhodoptilometrin (71) | Trichoderma harzianum (XS-20090075) | -Inner tissue of a soft coral. | -Antibacterial, cytotoxic and anti-acetylcholinesterase activities. | [55] |
| Nalgiovensin (72) | A. alliaceus | - | - | [77] |
| 1-Methylether nalgiovensin (73) | Emericella sp. SCSIO 05240 | -Marine sediment. | -Antibacterial activity. | [78] |
| Penipurdin A (74) | Neosartorya spinosa KUFA 1047 | -Marine sponge Mycale sp. | -Anti-tyrosinase activity. | [54] |
| Acetylpenipurdin A (75) | N. spinosa KUFA 1047 | -Marine sponge Mycale sp. | - | [54] |
| 1,3,6-trihydroxy-7-(dihydroxypropyl)-anthraquinone (76) | Thermomyces lanuginosus Tsikl KMM 4681 | -Marine sediment. | - | [66] |
| 6,8-Dimethoxy-1-methyl-2-(3-oxobutyl)-anthrakunthone (77) | Fusarium sp. ZZF60 | -Marine mangrove plant. | -Cytotoxic activity. | [79] |
| Norsolorinic acid (78) | A. nidulans MA-143 | -Leaves of mangrove plant Rhizophora stylosa. | - | [80] |
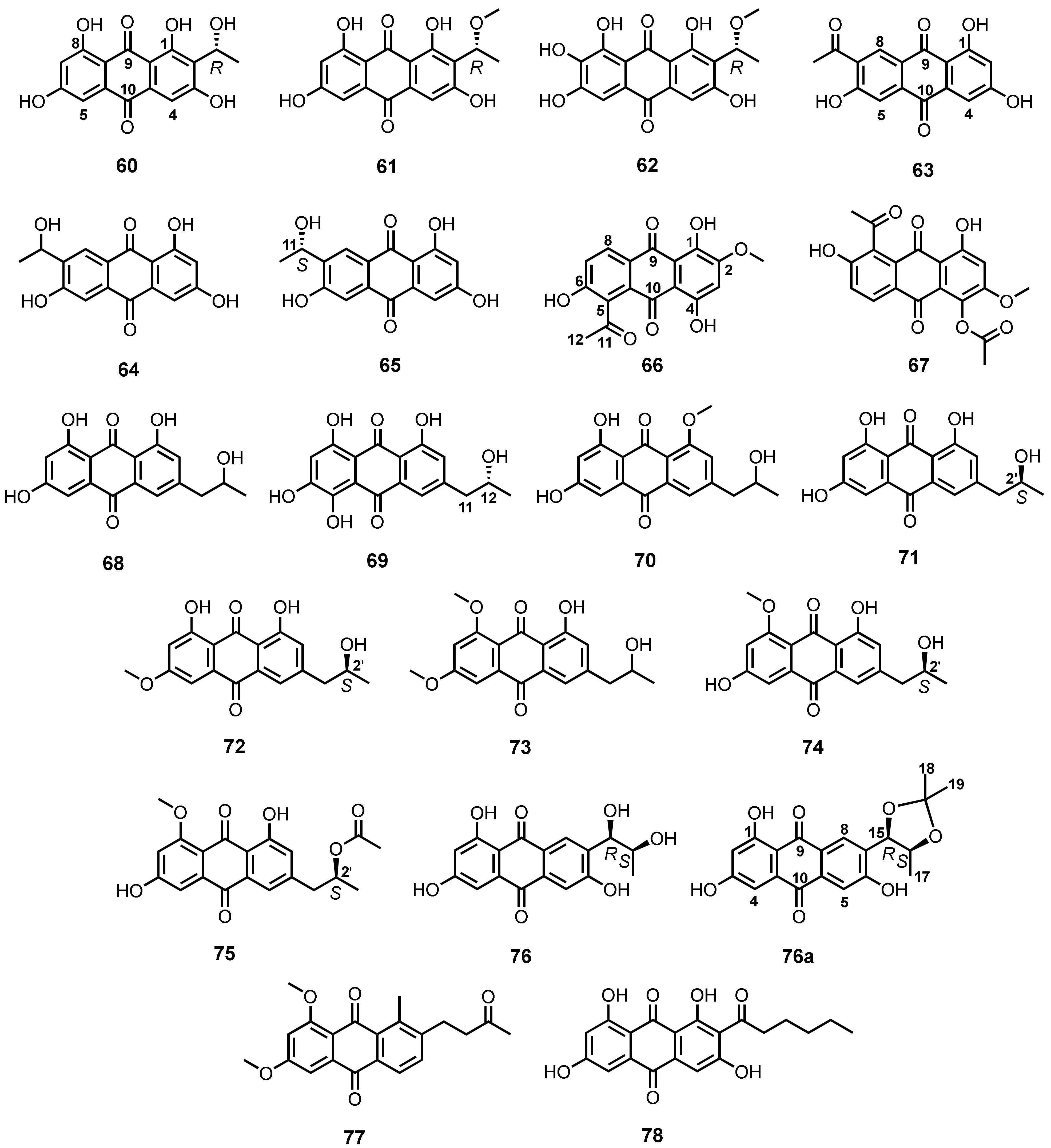
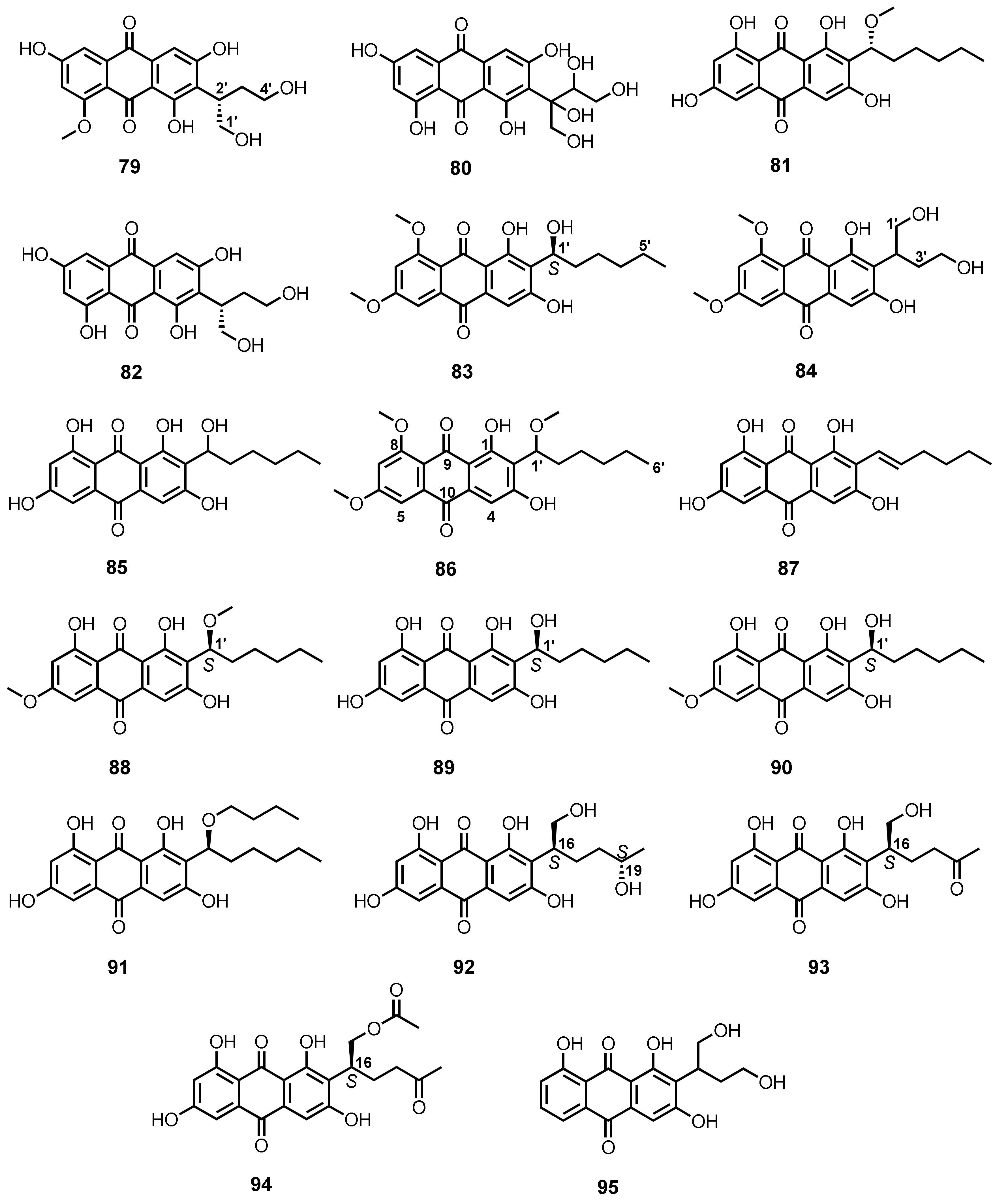
| 8-O-Methyl versiconol (79) | A. puniceus SCSIO z021 | -Deep-sea sediment. | -Inhibition of human protein tyrosine phosphatases. | [81] |
| 2′,3′-Dihydorxy versiconol (80) | A. puniceus SCSIO z021 | -Deep-sea sediment. | - | [81] |
| Methyl averantin (81) | A. puniceus SCSIO z021 | -Deep-sea sediment. | -Inhibition of human protein tyrosine phosphatases. | [81] |
| A. versicolor | -Marine sponge Petrosia sp. | -Antibacterial and cytotoxic activities. | [84] | |
| A. versicolor INF 16-17 | -Inner tissue of an unidentified marine clam. | - | [88] | |
| A. versicolor A-21-2-7 | -Deep-sea sediment. | - | [89] | |
| Aspergillus sp. SCSIO F063 | -Deep-sea sediment. | -Cytotoxic activity. | [91] | |
| A. versicolor SCSIO-41502 | -Deep-sea sediment. | - | [92] | |
| Versiconol (82) | A. puniceus SCSIO z021 | -Deep-sea sediment. | -Inhibition of human protein tyrosine phosphatases. | [81] |
| A. versicolor | -Marine sponge Petrosia sp. | -Cytotoxic activity. | [84] | |
| A. versicolor SCSIO-41502 | -Deep-sea sediment. | - | [92] | |
| Aspergillus sp. F40 | -Marine sponge Callyspongia sp. | -Antibacterial activity. | [93] | |
| Penicillium flavidorsum SHK1-27 | - | -Anti-proliferative activity. | [85] | |
| Strain ZSUH-36 | -Mangrove Acanthus ilicifolius Linn. | - | [83] | |
| 6,8-Di-O-methylaverantin (83) | A. versicolor EN-7 | -Brown algae Saragassum thunbergii. | -Antimicrobial activity. | [82] |
| 6,8-Di-O-methylversiconol (84) | A. versicolor EN-7 | -Brown algae Saragassum thunbergii. | -Antimicrobial activity. | [82] |
| Strain ZSUH-36 | -Mangrove Acanthus ilicifolius Linn. | - | [83] | |
| Averantin (85) | A. versicolor | -Marine sponge Petrosia sp. | -Antibacterial and cytotoxic activities. | [84] |
| A. versicolor INF 16-17 | -Inner tissue of an unidentified marine clam. | - | [88] | |
| A. versicolor A-21-2-7 | -Deep-sea sediment. | - | [89] | |
| Aspergillus sp. SCSIO F063 | - | -Cytotoxic activity. | [91] | |
| P. flavidorsum SHK1-27 | - | -Anti-proliferative activity. | [85] | |
| 6,8,1′-Tri-O-methylaverantin (86) | Aspergillus sp. SF-6796 | - | -Anti-neuroinflammatory activity. | [87] |
| Strain ZSUH-36 | -Mangrove Acanthus ilicifolius Linn. | - | [86] | |
| Averythrin (87) | A. versicolor INF 16-17 | -Inner tissue of an unidentified marine clam. | - | [88] |
| A. versicolor A-21-2-7 | -Deep-sea sediment. | -Antioxidant activity. | [89] | |
| Aspergillus sp. 16-5C | -Leaves of Sonneratia apetala. | -Anti-Mycobacterium tuberculosis activity. | [90] | |
| Aspergillus sp. SCSIO F063 | -Deep-sea sediment. | -Cytotoxic activity. | [91] | |
| (1′S)-6,1′-O,O-Dimethylaverantin (88) | Aspergillus sp. SCSIO F063 | -Deep-sea sediment. | -Cytotoxic activity. | [91] |
| (S)-(−)-Averantin (89) | Aspergillus sp. SCSIO F063 | -Deep-sea sediment. | - | [91] |
| 6,O-Methylaverantin (90) | Aspergillus sp. SCSIO F063 | -Deep-sea sediment. | - | [91] |
| Averantin-1′-butyl ether (91) | Aspergillus sp. SCSIO F063 | -Deep-sea sediment. | -Cytotoxic activity. | [91] |
| Aspergilol I (92) | A. versicolor SCSIO-41502 | -Deep-sea sediment. | -Antiviral activity. | [92] |
| SC3-22-3 (93) | A. versicolor SCSIO-41502 | -Deep-sea sediment. | - | [92] |
| Coccoquinone A (94) | A. versicolor SCSIO-41502 | -Deep-sea sediment. | -Antiviral activity. | [92] |
| Versiconol B (95) | Aspergillus sp. F40 | -Marine sponge Callyspongia sp. | -Antibacterial activity. | [93] |
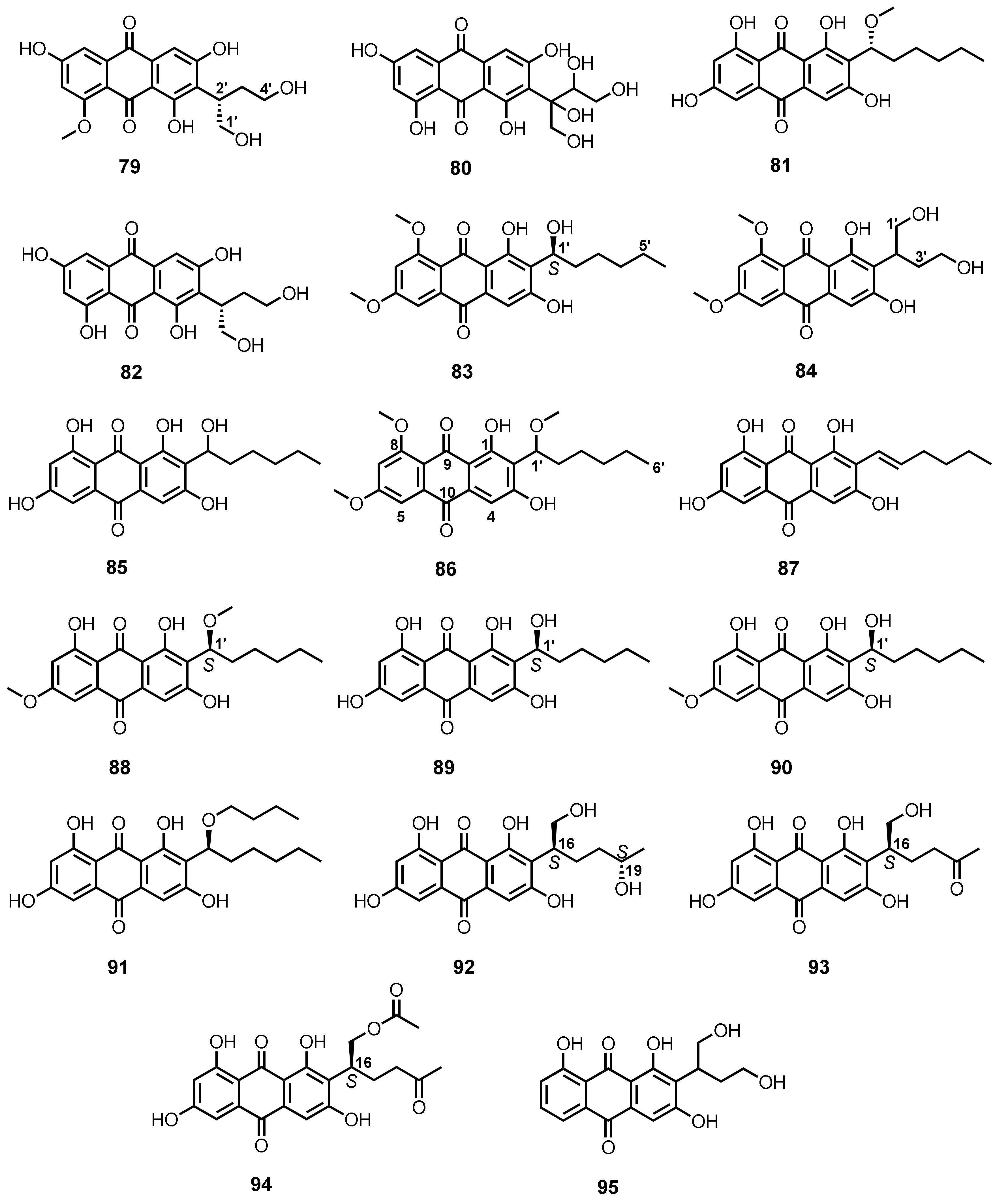
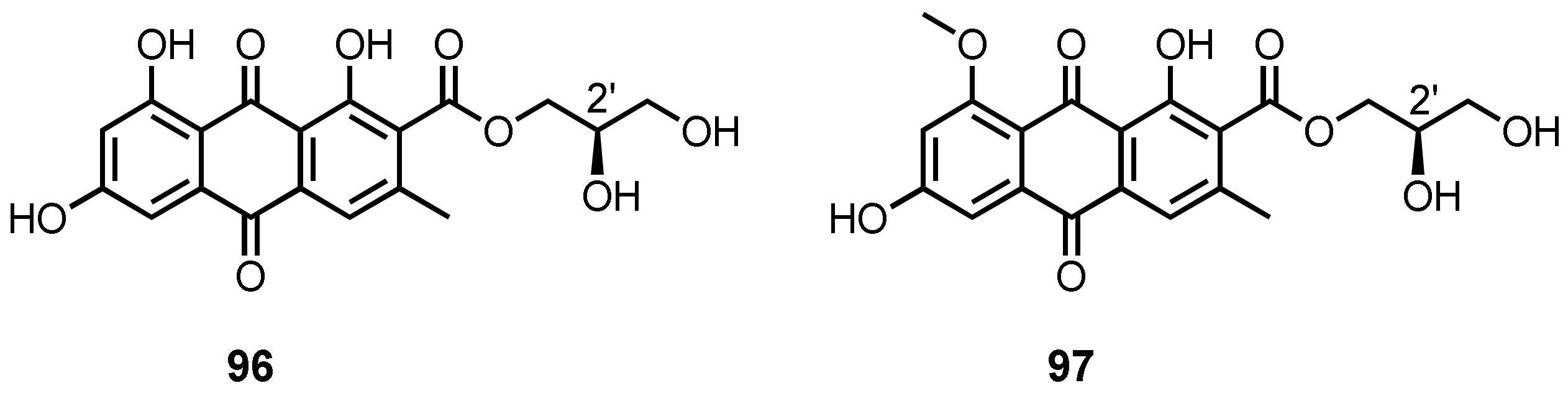
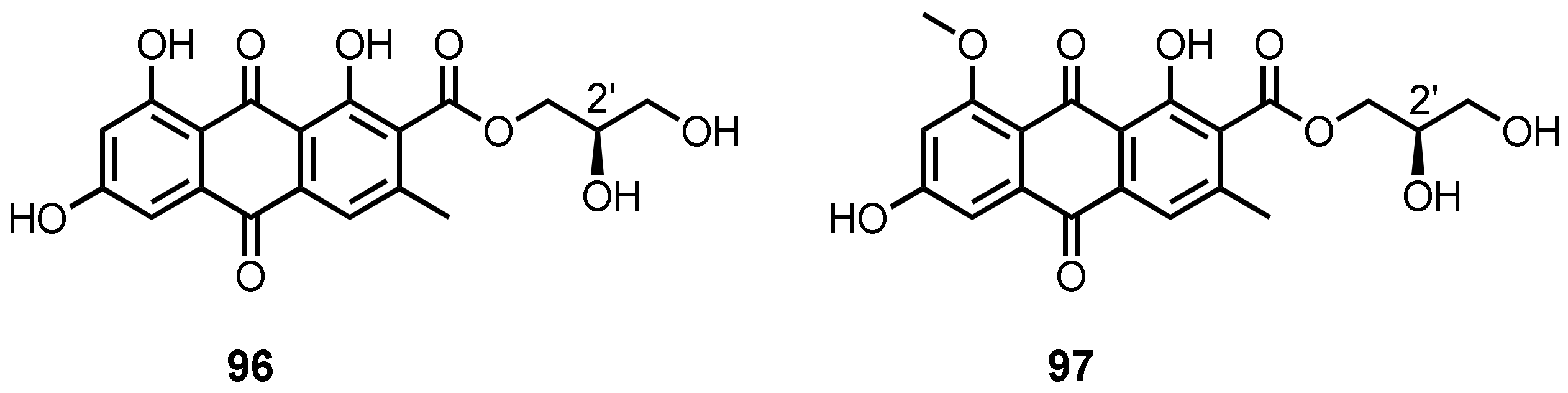
| (+)-1-O-Demethylvariecolorquinone A (96) | A. europaeus WZXY-SX-4-1 | -Marine sponge Xestospongia testudinaria. | - | [21] |
| (+)-Variecolorquinone A (97) | A. europaeus WZXY-SX-4-1 | -Marine sponge X. testudinaria. | -DPPH• radical scavenging activity. | [21] |
| A. glaucus HB1-19 | -Deep-sea sediment. | - | [22] | |
| Eurotium cristatum EN-220 | -Marine brown alga Sargassum thunbergii. | - | [95] | |
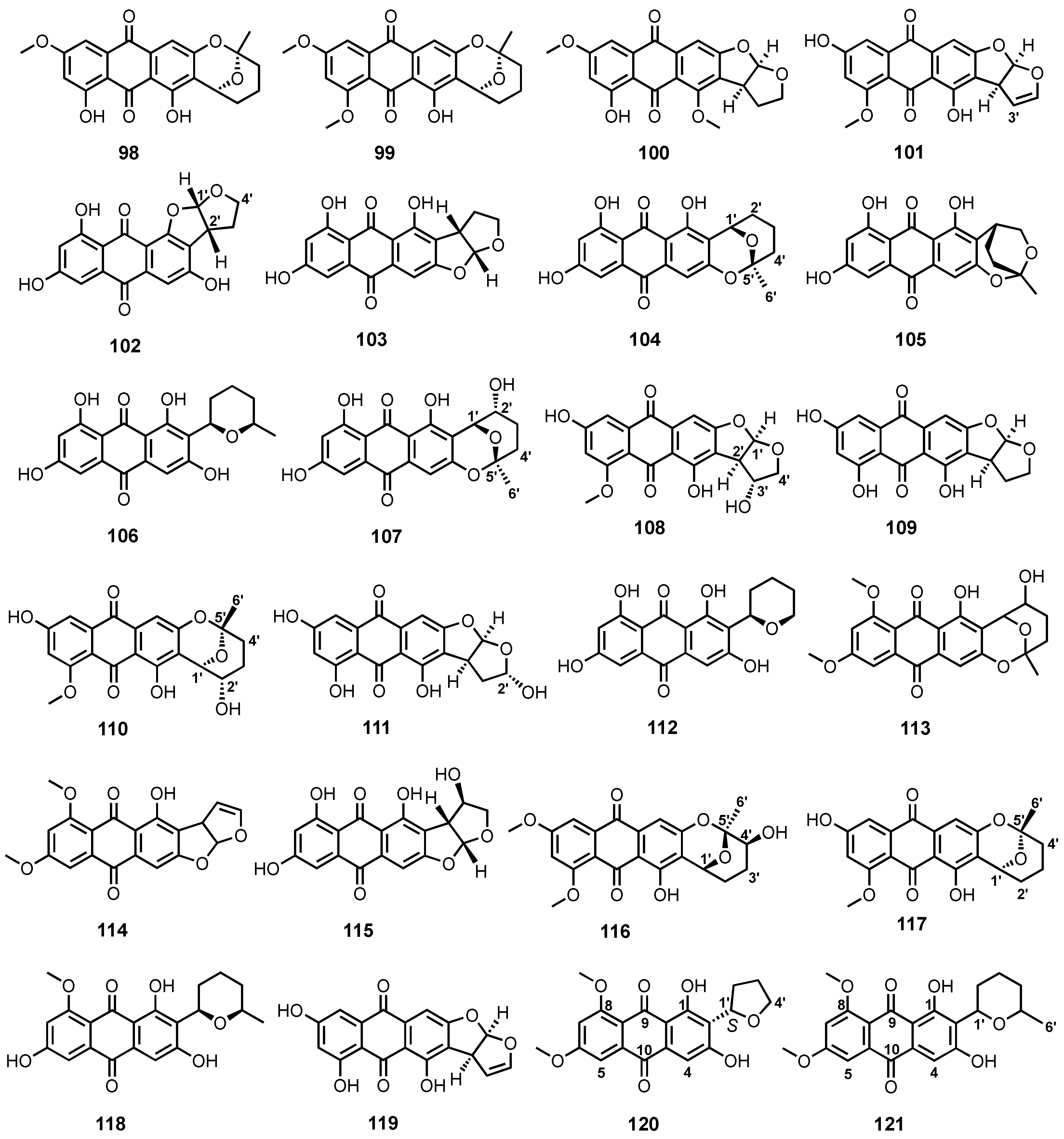
| 6-O-Methylaverufin (98) | A. nidulans MCCC 3A00050 | -Deep-sea sediment. | - | [96] |
| A. versicolor EN-7 | -Brown alga Sargassum thunbergii. | - | [82] | |
| 6,8-Di-O-methylaverufin (99) | A. nidulans MCCC 3A00050 | -Deep-sea sediment. | - | [96] |
| Aspergillus sp. SF-6796 | - | - | [87] | |
| A. versicolor EN-7 | -Brown alga Sargassum thunbergii. | - | [82] | |
| Aspergillus sp. 16-5C | -Leaves of Sonneratia apetala. | - | [90] | |
| P. flavidorsum SHK1-27 | - | -Anti-proliferative activity. | [85] | |
| Acremonium vitellinum | -Inner tissue of an unidentified marine red alga. | -Insecticidal activity. | [104] | |
| Strain ZSUH-36 | -Mangrove Acanthus ilicifolius Linn. | - | [86] | |
| Aversin (100) | A. nidulans MCCC 3A00050 | -Deep-sea sediment. | - | [96] |
| A. versicolor MF359 | -Marine sponge Hymeniacidon perleve. | - | [97] | |
| A. versicolor EN-7 | -Brown alga Sargassum thunbergii. | - | [82] | |
| Aspergillus sp. 16-5C | -Leaves of Sonneratia apetala | - | [90] | |
| Acremonium vitellinum | -Inner tissue of an unidentified marine red alga. | -Insecticidal activity. | [104] | |
| P. flavidorsum SHK1-27 | - | -Anti-proliferative activity. | [85] | |
| Strain ZSUH-36 | -Mangrove Acanthus ilicifolius Linn. | - | [83] | |
| 8-O-Methylversicolorin A (101) | A. nidulans MCCC 3A00050 | -Deep-sea sediment. | - | [96] |
| Isoversicolorin C (102) | A. nidulans MA-143 | -Leaves of mangrove plant Rhizophora stylosa. | -Antibacterial activity. | [80] |
| Versicolorin C (103) | A. nidulans MA-143 | -Leaves of mangrove plant Rhizophora stylosa. | -Antibacterial activity. | [80] |
| Strain ZSUH-36 | -Mangrove Acanthus ilicifolius Linn. | - | [86] | |
| Isolate 1850 | -Leaves of a mangrove tree Kandelia candel. | - | [100] | |
| Averufin (104) | A. nidulans MA-143 | -Leaves of mangrove plant Rhizophora stylosa. | - | [80] |
| A. versicolor SCSIO-41502 | -Deep-sea sediment. | - | [92] | |
| - | - | -Anti-Mycobacterium tuberculosis activity. | [129] | |
| A. niger (MF-16) | -Sea water. | -Antiviral activity. | [98] | |
| A. versicolor | -Marine sponge Petrosia sp. | -Antibacterial and cytotoxic activities. | [84] | |
| A. puniceus SCSIO z021 | -Deep-sea sediment. | -Inhibition of protein tyrosine phosphatases and toxicity against Brine shrimps. | [81] | |
| A. versicolor MF18051 | -Marine sediment. | -Antibacterial activity. | [101] | |
| Aspergillus sp. F40 | -Marine sponge Callyspongia sp. | - | [93] | |
| A. versicolor SCSIO 41016 | -Marine sponge. | - | [102] | |
| A. versicolor A-21-2-7 | -Deep-sea sediment. | -Antioxidant activity. | [89] | |
| Aspergillus sp. | -Gorgonian Dichotella gemmacea. | -Antibacterial activity. | [103] | |
| P. flavidorsum SHK1-27 | - | -Anti-proliferative activity. | [85] | |
| Strain ZSUH-36 | -Mangrove Acanthus ilicifolius Linn. | - | [86] | |
| Isolate 1850 | -Leaves of a mangrove tree Kandelia candel. | - | [100] | |
| Paeciloquinone E (105) | A. nidulans MA-143 | -Leaves of mangrove plant Rhizophora stylosa. | - | [80] |
| Averufanin (106) | A. nidulans MA-143 | -Leaves of mangrove plant Rhizophora stylosa. | - | [80] |
| A. puniceus SCSIO z021 | -Deep-sea sediment. | -Inhibition of protein tyrosine phosphatases and toxicity against Brine shrimps. | [81] | |
| A. versicolor A-21-2-7 | -Deep-sea sediment. | - | [89] | |
| Aspergillus sp. | -Gorgonian Dichotella gemmacea. | -Antibacterial activity. | [103] | |
| Nidurufin (107) | A. niger (MF-16) | -Sea water. | -Antiviral activity. | [98] |
| A. versicolor | -Marine sponge Petrosia sp. | -Antibacterial and cytotoxic activities. | [84] | |
| A. puniceus SCSIO z021 | -Deep-sea sediment. | -Inhibition of protein tyrosine phosphatases. | [81] | |
| A. versicolor SCSIO 41016 | -Marine sponge. | -Antiviral activity. | [102] | |
| A. versicolor A-21-2-7 | -Deep-sea sediment. | -Antioxidant and cytotoxic activities. | [89] | |
| Aspergillus sp. | -Gorgonian Dichotella gemmacea. | -Antibacterial and cytotoxic activities. | [103] | |
| P. flavidorsum SHK1-27 | - | -Anti-proliferative activity and cell cycle inhibitor. | [85,99] | |
| Isolate 1850 | -Leaves of a mangrove tree Kandelia candel. | - | [100] | |
| 3′-Hydroxy-8-O-methyl verscicolorin B (108) | A. puniceus SCSIO z021 | -Deep-sea sediment. | -Inhibition of protein tyrosine phosphatases. | [81] |
| Versicolorin B (109) | A. puniceus SCSIO z021 | -Deep-sea sediment. | -Inhibition of protein tyrosine phosphatases and toxicity against Brine shrimps. | [81] |
| A. versicolor MF18051 | -Marine sediment. | -Antibacterial activity. | [101] | |
| Aspergillus sp. F40 | -Marine sponge Callyspongia sp. | - | [93] | |
| A. versicolor SCSIO 41016 | -Marine sponge. | - | [102] | |
| A. versicolor A-21-2-7 | -Deep-sea sediment. | -Antioxidant and cytotoxic activities. | [89] | |
| P. flavidorsum SHK1-27 | - | -Anti-proliferative activity. | [85] | |
| 8-O-Methylnidurufin (110) | A. puniceus SCSIO z021 | -Deep-sea sediment. | -Inhibition of protein tyrosine phosphatases. | [81] |
| Aspergillus sp. | -Gorgonian Dichotella gemmacea. | -Antibacterial activity. | [103] | |
| 2′-Hydroxyversicolorin B (111) | A. versicolor SCSIO 41016 | -Marine sponge. | - | [102] |
| Noraverufanin (112) | Aspergillus versicolor SCSIO 41016 | -Marine sponge. | -Antiviral activity. | [102] |
| 6,8-Di-O-methylnidurufin (113) | A. versicolor EN-7 | -Brown alga Sargassum thunbergii. | -Antibacterial activity. | [82] |
| Aspergillus sp. 16-5C | -Leaves of Sonneratia apetala. | - | [90] | |
| Acremonium vitellinum | -Inner tissue of an unidentified marine red alga. | -Insecticidal activity. | [104] | |
| 6,8-Di-O-methylversicolorin A (114) | A. versicolor EN-7 | -Brown alga Sargassum thunbergii. | - | [82] |
| UCT1072M1 (115) | A. versicolor A-21-2-7 | -Deep-sea sediment. | -Antioxidant activity. | [89] |
| Asperquinone A (116) | Aspergillus sp. 16-5C | -Leaves of Sonneratia apetala. | - | [90] |
| 8-O-Methylaverufin (117) | Aspergillus sp. | -Gorgonian Dichotella gemmacea. | -Antibacterial activity | [103] |
| 8-O-Methylaverufanin (118) | Aspergillus sp. | -Gorgonian D. gemmacea. | - | [103] |
| P. flavidorsum SHK1-27 | - | -Anti-proliferative activity. | [85] | |
| Versicolorin A (119) | P. flavidorsum SHK1-27 | - | -Anti-proliferative activity. | [85] |
| 6,8-Di-O-methylbipolarin (120) | A. vitellinum | -Inner tissue of an unidentified marine red alga. | -Insecticidal activity. | [104] |
| 6,8-Di-O-methyl-averufinan (121) | Strain ZSUH-36 | -Mangrove Acanthus ilicifolius Linn. | - | [86] |
| Aspergilol (±)-A (122) | A. versicolor SCSIO-41502 | -Deep-sea sediment. | -Antioxidant activity. | [92] |
| Aspergilol (±)-B (123) | A. versicolor SCSIO-41502 | -Deep-sea sediment. | -Antioxidant activity. | [92] |
| Aspergilol (±)-G (124) | A. versicolor SCSIO-41502 | -Deep-sea sediment. | -Antioxidant activity. | [92] |
| Aspergilol (±)-H (125) | A. versicolor SCSIO-41502 | -Deep-sea sediment. | -Antioxidant and antiviral activities. | [92] |
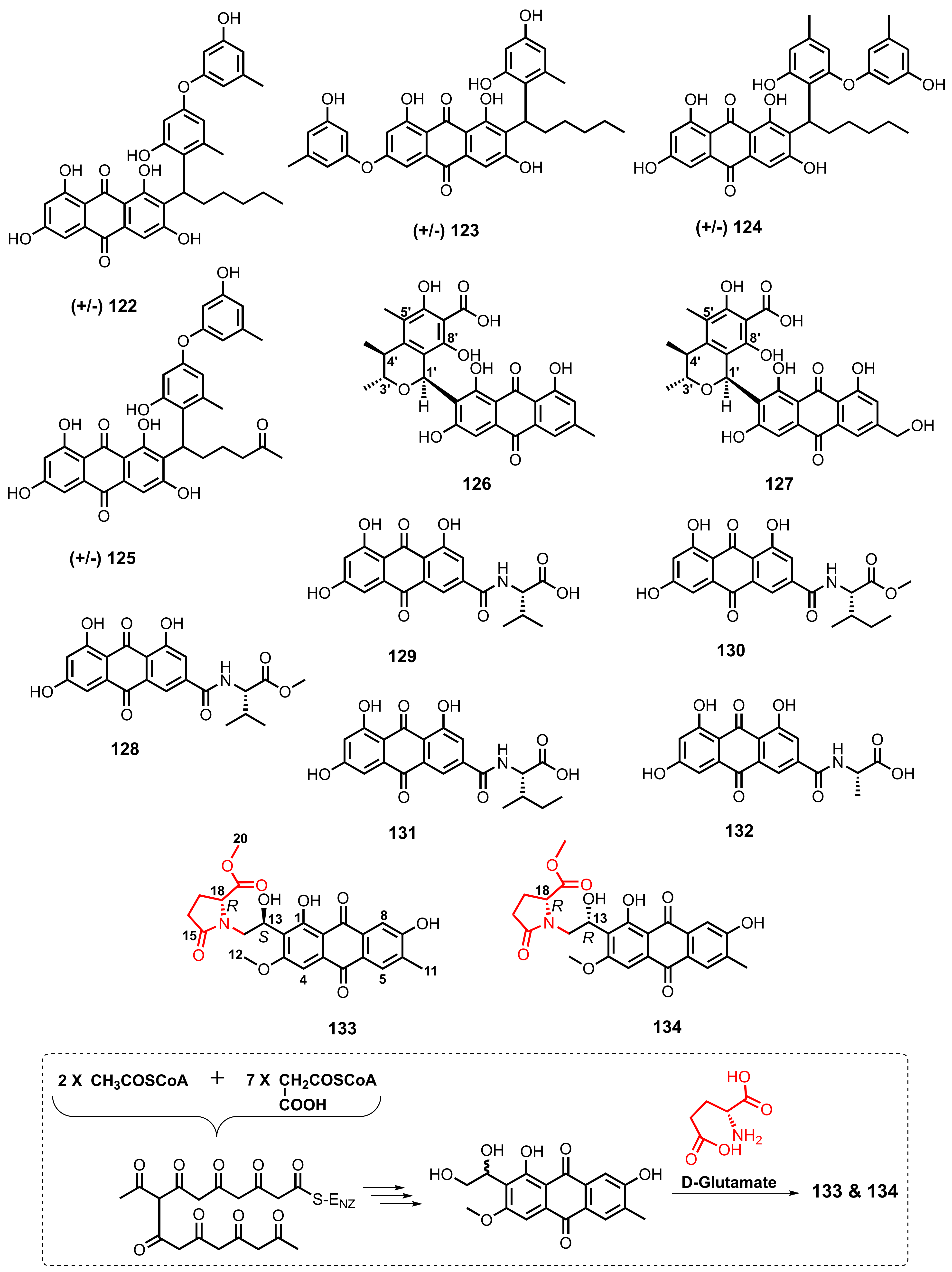
| Penicillanthranin A (126) | Penicillium citrinum PSU-F51 | -Gorgonian Sea fan (Annella sp.) | -Antibacterial and cytotoxic activities. | [37] |
| Penicillanthranin B (127) | P. citrinum PSU-F51 | -Gorgonian Sea fan (Annella sp.) | - | [37] |
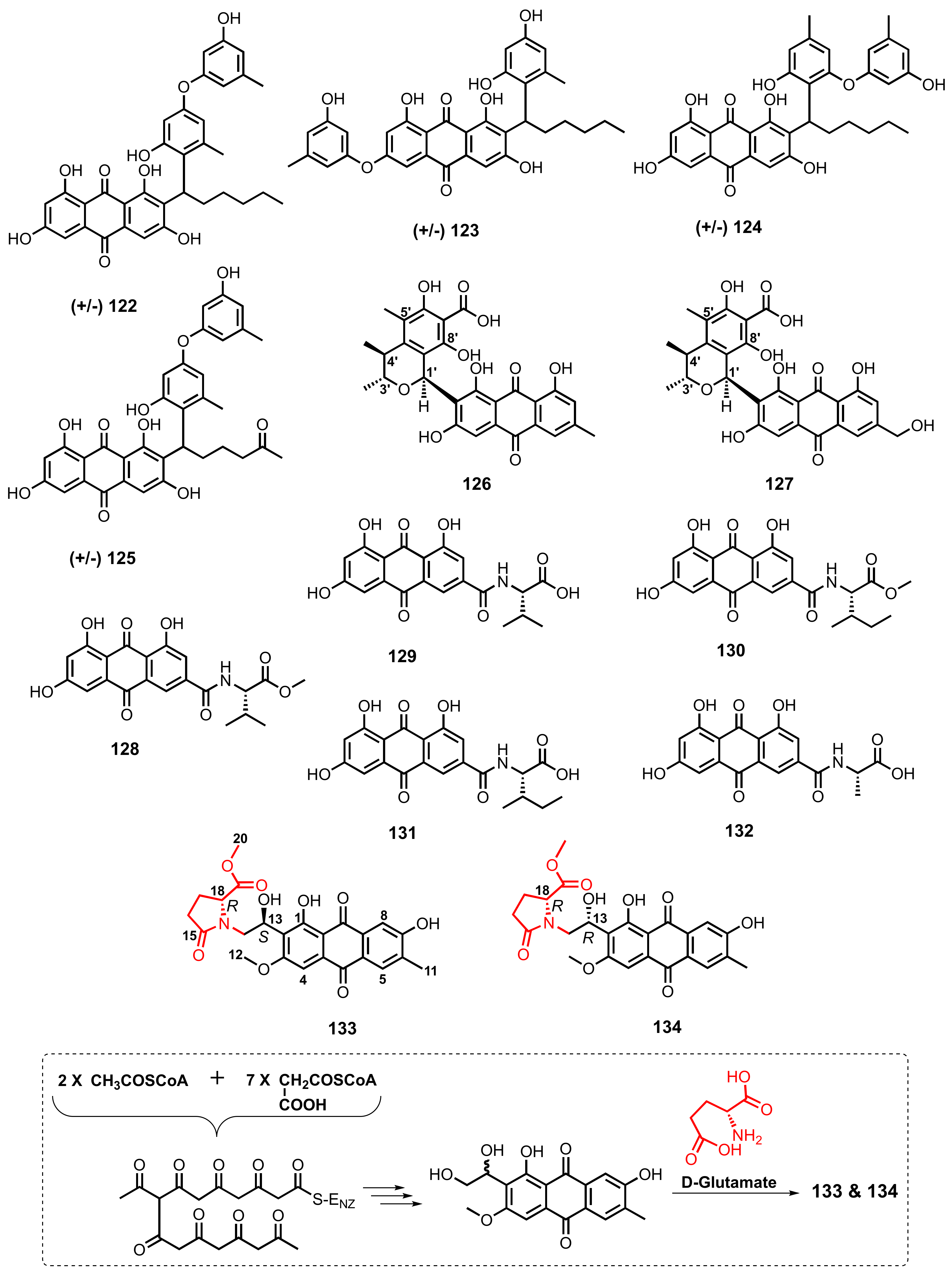
| Emodacidamide A (128) | Penicillium sp. SCSIOsof101 | -Marine sediment. | -Anti-inflammatory activity. | [46] |
| Emodacidamide B (129) | Penicillium sp. SCSIOsof101 | -Marine sediment. | - | [46] |
| Emodacidamide D (130) | Penicillium sp. SCSIOsof101 | -Marine sediment. | -Anti-inflammatory activity. | [46] |
| Emodacidamide E (131) | Penicillium sp. SCSIOsof101 | -Marine sediment. | -Anti-inflammatory activity. | [46] |
| Emodacidamide H (132) | Penicillium sp. SCSIOsof101 | -Marine sediment. | - | [46] |
| Anthrininone B (133) | Altenaria tenuissima DFFSCS013 | -Marine sediment. | -Inhibition of human protein tyrosine phosphatases and inhibition of indoleamine 2,3-dioxygenase activity. | [63] |
| Anthrininone C (134) | A. tenuissima DFFSCS013 | -Marine sediment. | -Inhibition of human protein tyrosine phosphatases and inhibition of indoleamine 2,3-dioxygenase activity. | [63] |
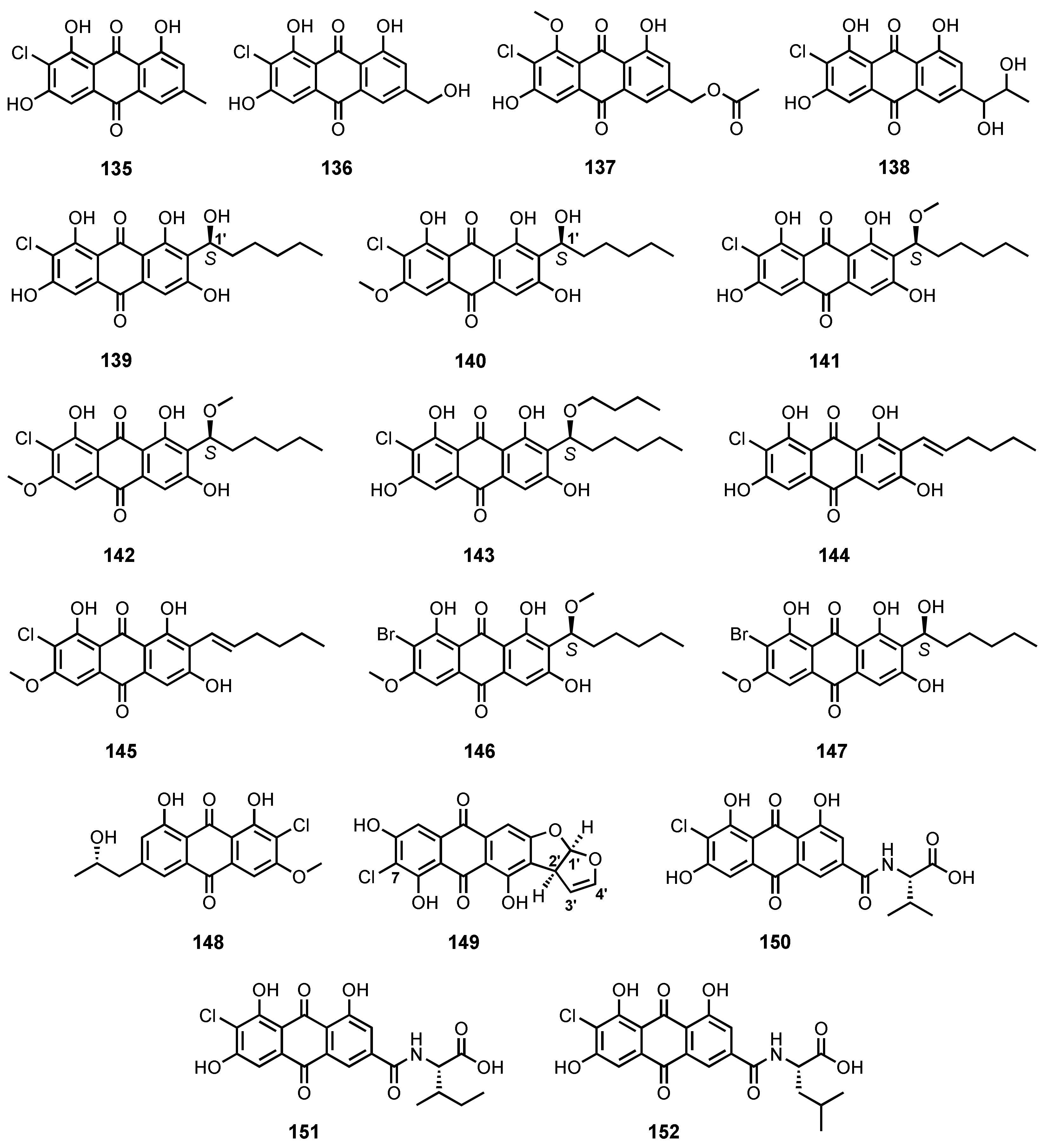
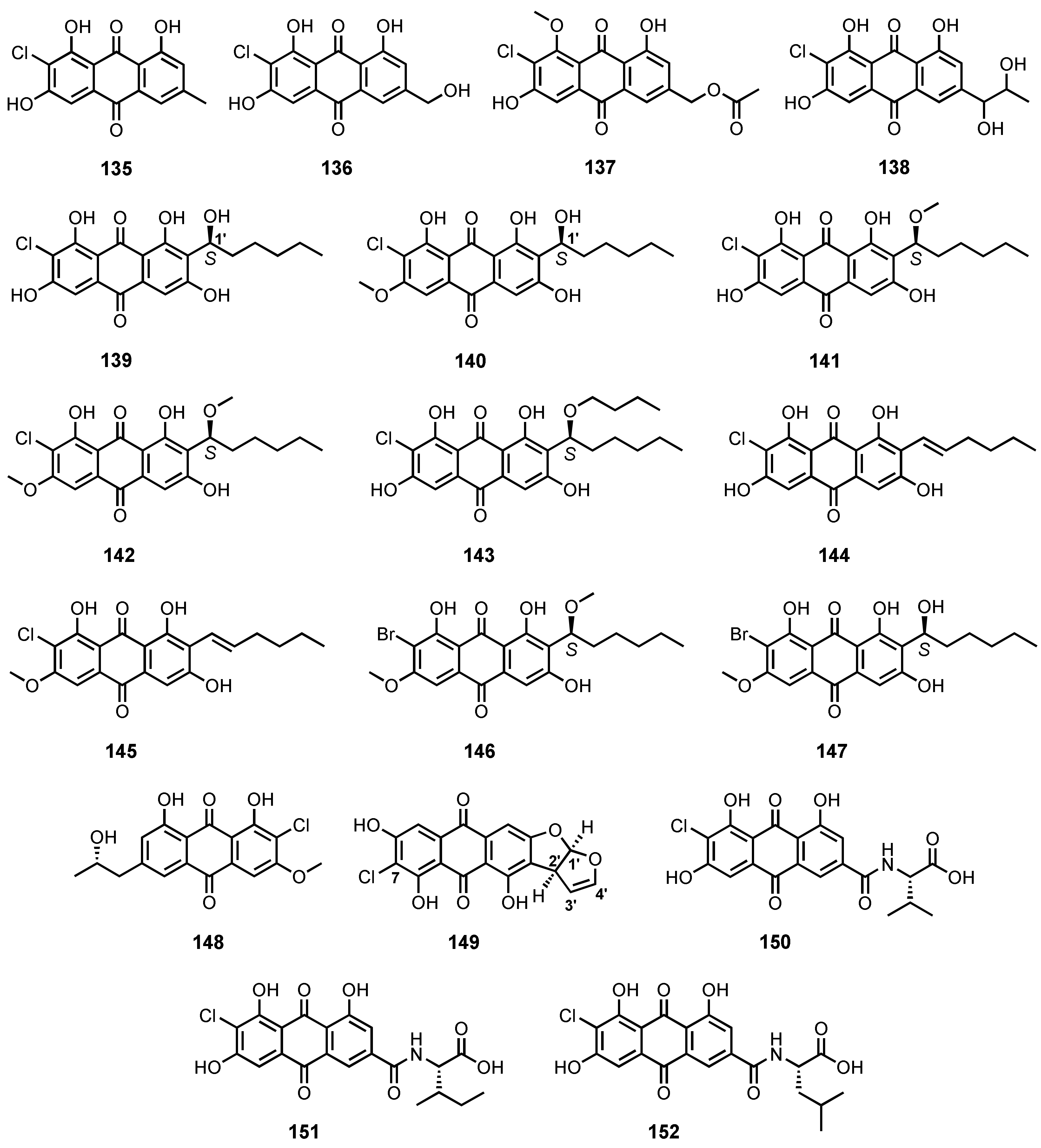
| 7-Chloroemodin (135) | Penicillium ochrochloron | -Sea sand. | - | [17] |
| 2-Chloro-1,3,8-trihydroxy-6-(hydroxy methyl)anthracene-9,10-dione (136) | Penicillium sp. SCSIOsof101 | -Marine sediment. | - | [46] |
| 2′-Acetoxy-7-chlorocitreorosein (137) | P. citrinum HL-5126 | -Mangrove Bruguiera sexangula var. rhynchopetala. | -Antibacterial activity. | [41] |
| 7-Chloro-1′-hydroxyisorhodoptilometrin (138) | Penicillium sp. SCSIO sof101 | -Marine sediment. | - | [105] |
| (1′S)-7-Chloroaverantin (139) | Aspergillus sp. SCSIO F063 | -Marine sediment. | -Cytotoxic activity. | [91] |
| (1′S)-6-O-Methyl-7-chloroaverantin (140) | Aspergillus sp. SCSIO F063 | -Marine sediment. | -Cytotoxic activity. | [91] |
| (1′S)-1′-O-Methyl-7-chloroaverantin (141) | Aspergillus sp. SCSIO F063 | -Marine sediment. | -Cytotoxic activity. | [91] |
| (1′S)-6,1′-O,O-Dimethyl-7-chloroaverantin (142) | Aspergillus sp. SCSIO F063 | -Marine sediment. | -Cytotoxic activity. | [91] |
| (1′S)-7-Chloroaverantin-1′-butyl ether (143) | Aspergillus sp. SCSIO F063 | -Marine sediment. | -Cytotoxic activity. | [91] |
| 7-Chloroaverythrin (144) | Aspergillus sp. SCSIO F063 | -Marine sediment. | -Cytotoxic activity. | [91] |
| 6-O-Methyl-7-chloroaverythrin (145) | Aspergillus sp. SCSIO F063 | -Marine sediment. | -Cytotoxic activity. | [91] |
| (1′S)-6,1′-O,O-Dimethyl-7-bromoaverantin (146) | Aspergillus sp. SCSIO F063 | -Marine sediment. | -Cytotoxic activity. | [91] |
| (1′S)-6-O-Dimethyl-7-bromoaverantin (147) | Aspergillus sp. SCSIO F063 | -Marine sediment. | -Cytotoxic activity. | [91] |
| Nalgiolaxin (148) | A. alliaceus | -Marine algae. | - | [77] |
| 7-Chloro-versicolorin A (149) | A. puniceus SCSIO z021 | -Deep-sea sediment. | -Inhibition of protein tyrosine phosphatases and toxicity against Brine shrimps. | [81] |
| Emodacidamide C (150) | Penicillium sp. SCSIOsof101 | -Marine sediment. | -Anti-inflammatory activity. | [46] |
| Emodacidamide F (151) | Penicillium sp. SCSIOsof101 | -Marine sediment. | - | [46] |
| Emodacidamide G (152) | Penicillium sp. SCSIOsof101 | -Marine sediment. | - | [46] |
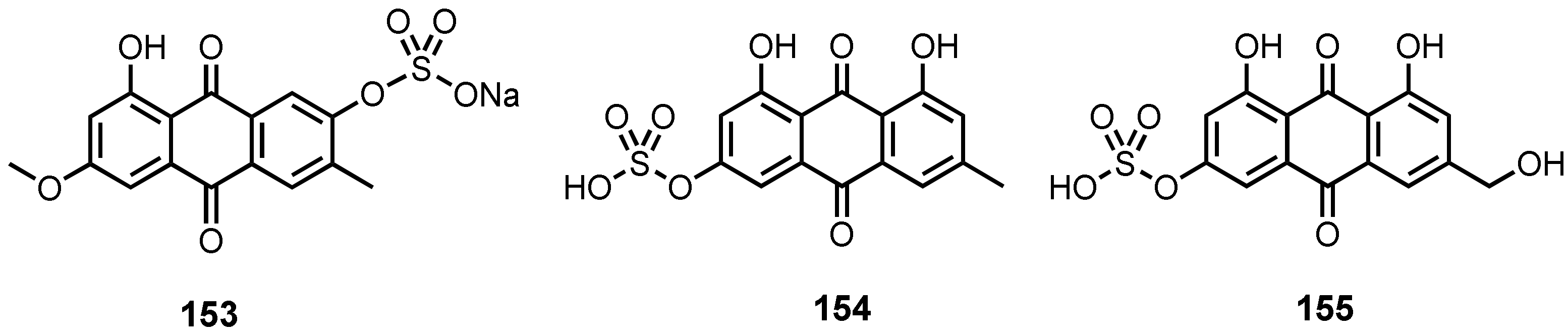
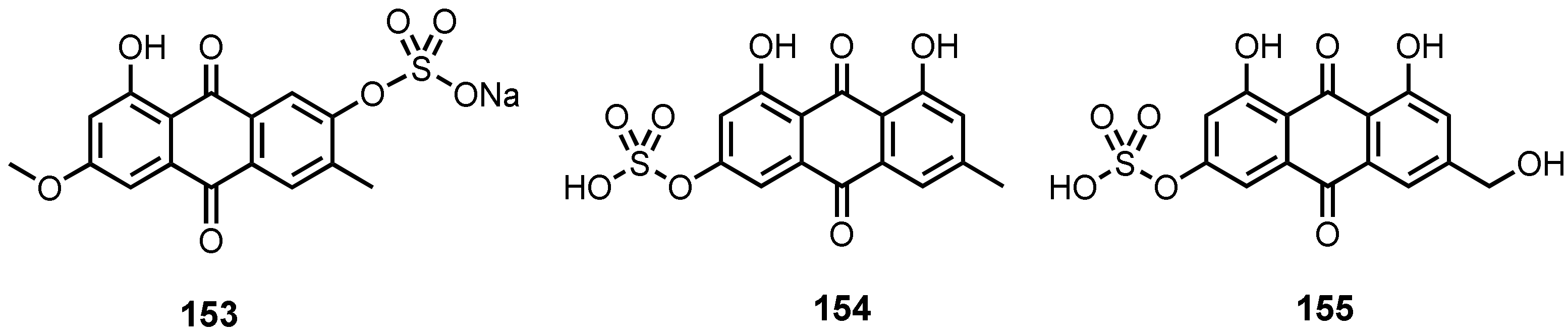
| Macrosposrin-7-O-sulfate (153) | Stemphylium sp. 33231 | -Mangrove tree Bruguiera sexangula var. rhynchopetala. | - | [50] |
| Emodin-3-O-sulphate (154) | Penicillium P. oxalicum 2HL-M-6 | -Sea mud. | - | [36] |
| Citreorosein-3-O-sulphate (155) | P. oxalicum 2HL-M-6 | -Sea mud. | - | [36] |
| Eurotium rubrum | -Inner tissue of semi-mangrove plant Hibiscus tiliaceus. | -DPPH• radicals scavenging activity. | [106] | |
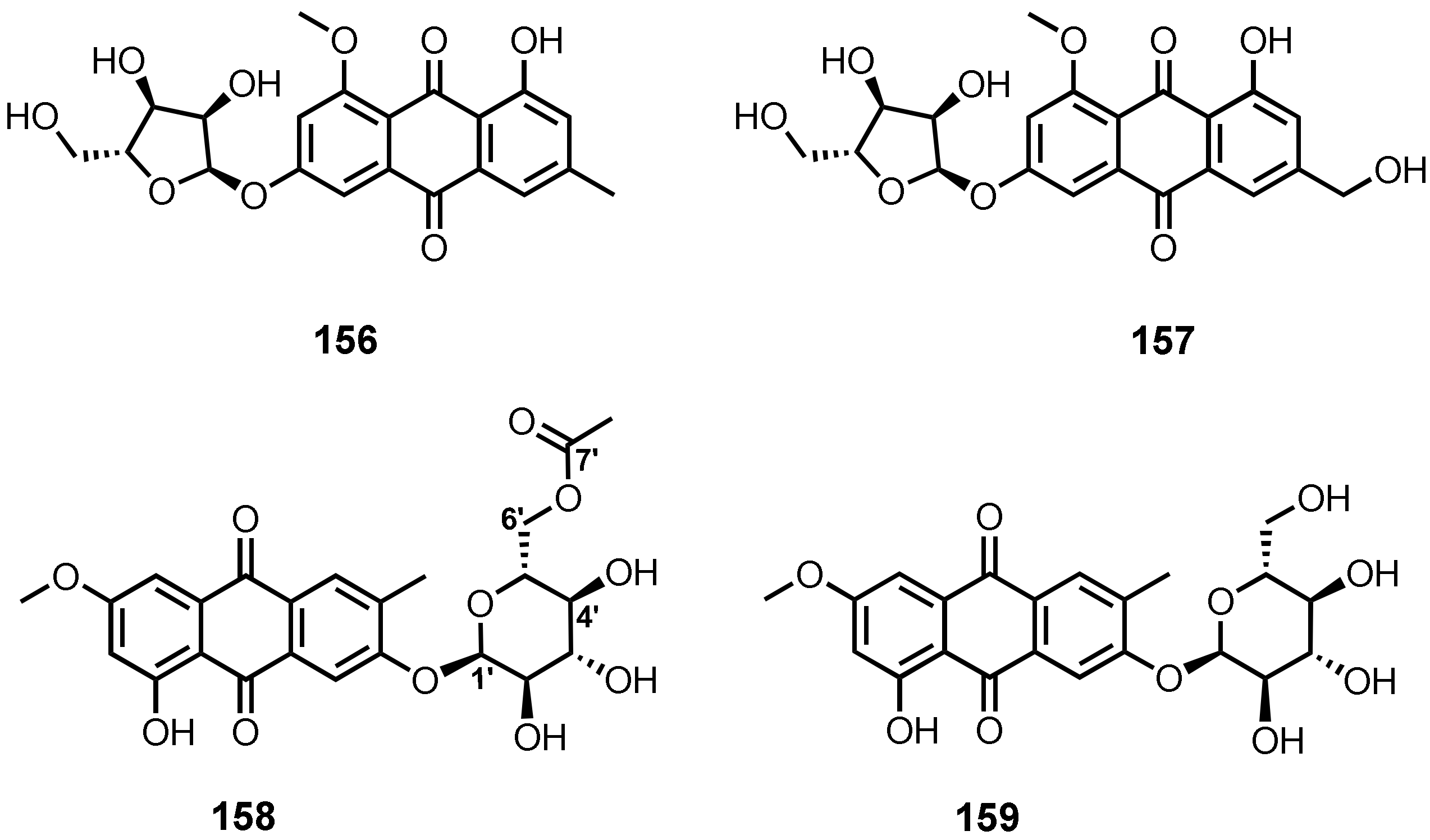
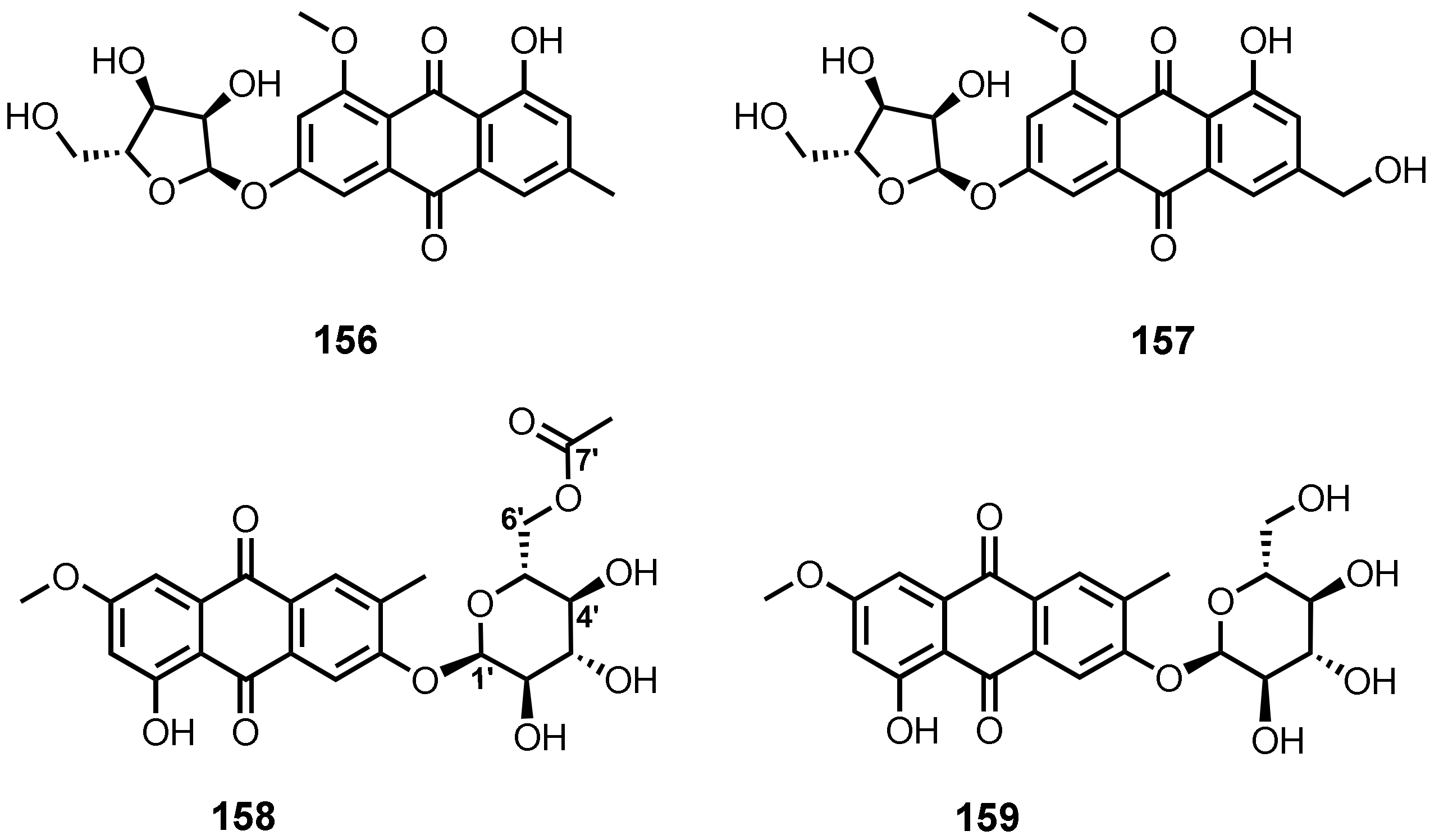
| 6-O-(α-D-ribofuranosyl)-questin (156) | Eurotium rubrum | -Inner tissue of mangrove plant H. tiliaceus. | -DPPH• radicals scavenging activity. | [106] |
| E. cristatum EN-220 | -Marine brown alga Sargassum thunbergii. | -Antibacterial activity. | [95] | |
| 6-O-(α-D-ribofuranosyl)-questinol (157) | E. cristatum EN-220 | -Marine brown alga S. thunbergii. | -Antibacterial activity. | [95] |
| 2-O-(6′-Acetyl)-α-D-glucopyranoside (158) | Stemphylium sp. 33231 | -Mangrove tree Bruguiera sexangula var. rhynchopetala. | -Cytotoxic activity. | [50] |
| Macrosporin 2-O-α-D-glucopyranoside (159) | S. lycopersici | -Inner tissue of gorgonian soft coral Dichotella gammacea. | - | [51] |
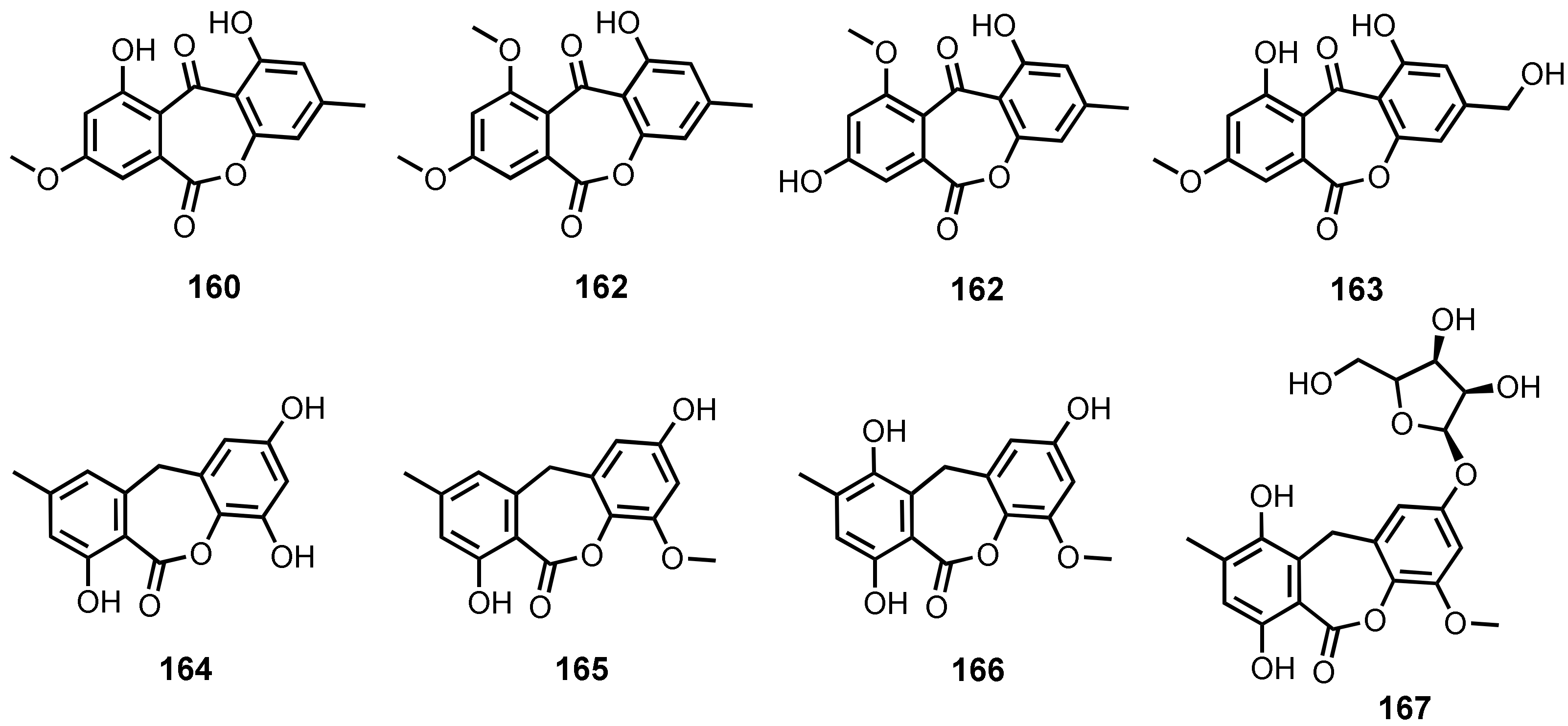
| Wentiquinone A (160) | Aspergillus wentii EN-48 | -Marine alga Sargassum sp. | - | [30] |
| Wentiquinone B (161) | A. wentii EN-48 | -Marine alga Sargassum sp. | - | [30] |
| 1,8-Dihydroxy-10-methoxy-3-methyldibenzo[b,e]oxepin-6,11-dione (162) | A. wentii EN-48 | -Marine alga Sargassum sp. | - | [30] |
| A. europaeus WZXY-SX-4-1 | -Marine sponge Xestospongia testudinaria. | - | [20] | |
| Wentiquinone C (163) | A. europaeus WZXY-SX-4-1 | -Marine sponge X. testudinaria. | - | [20] |
| A. wentii EN-48 | -Marine brown alga Sargassum sp. | - | [23] | |
| 9-Dehydroxyeurotinone (164) | E. rubrum | -Inner tissue of semi-mangrove plant Hibiscus tiliaceus. | -Cytotoxic activity. | [47] |
| 2-O-Methyl-9-dehydroxyeurotinone (165) | E. rubrum | -Inner tissue of semi-mangrove plant H. tiliaceus. | - | [47] |
| Eurotium sp. SCSIO F452 | -Marine sediment. | - | [70] | |
| E. rubrum | -Inner tissue of semi-mangrove plant H. tiliaceus. | -DPPH• radicals scavenging activity. | [106] | |
| 2-O-Methyleurotinone (166) | E. rubrum | -Inner tissue of semi-mangrove plant H. tiliaceus. | -DPPH• radicals scavenging activity. | [106] |
| 2-O-Methyl-4-O-(α-D-ribofuranosyl)-9-dehydroxyeurotinone (167) | Eurotium rubrum | -Inner tissue of semi-mangrove plant H. tiliaceus. | -DPPH• radicals scavenging activity. | [106] |
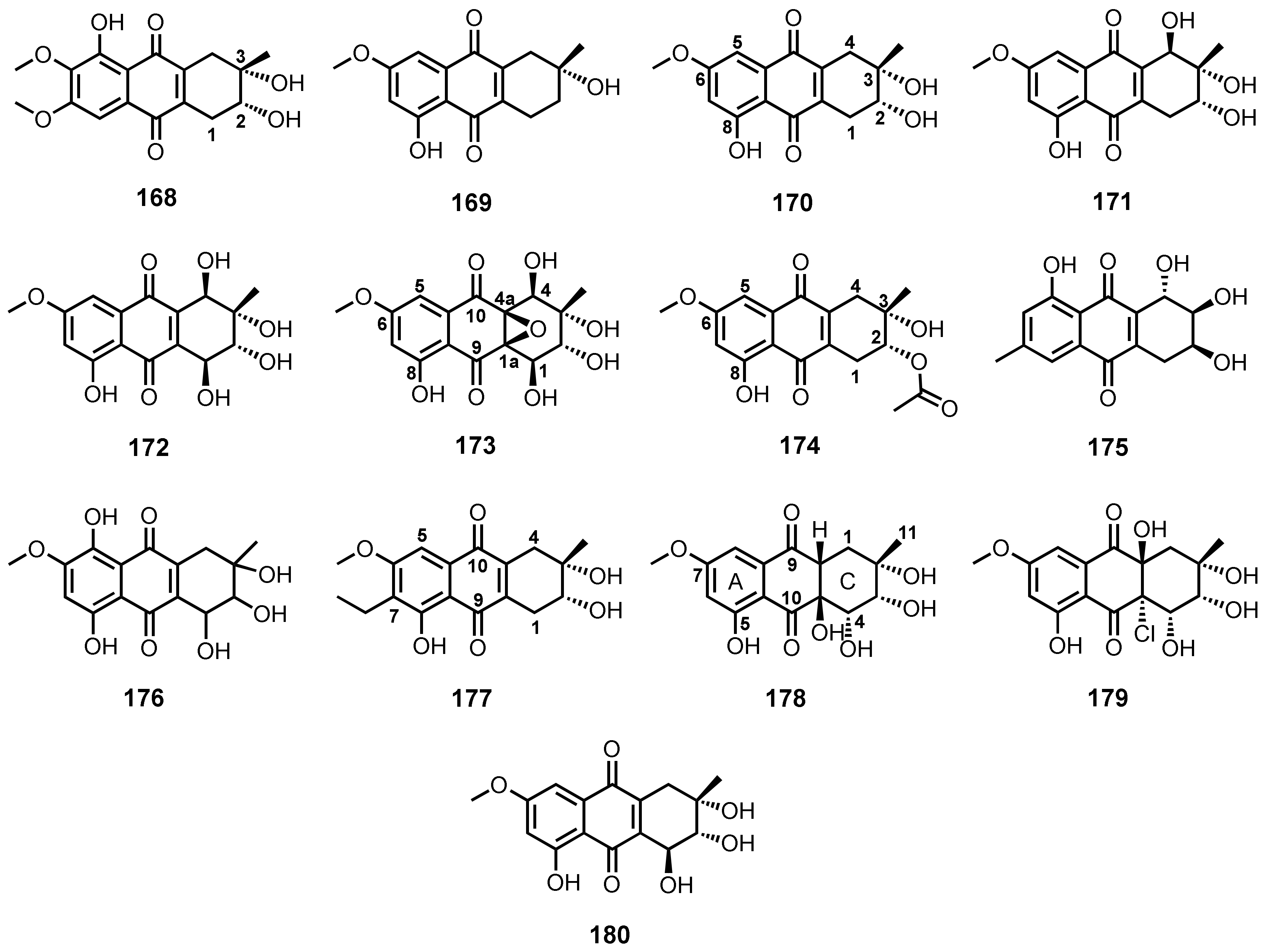
| Aspetritone B (168) | A. tritici SP2-8-1 | -Soft coral Galaxea fascicularis. | -Antibacterial and cytotoxic activities. | [28] |
| (3R)-1-Deoxyaustrocortilutein (169) | Altenaria tenuissima DFFSCS013 | -Marine sediment. | - | [63] |
| Altersolanol B (or dactylarin; 170) | Altenaria tenuissima DFFSCS013 | -Marine sediment. | - | [63] |
| Altenaria sp. ZJ9-6B | -Fruits of a mangrove tree Aegiceras corniculatum. | - | [61] | |
| Altenaria sp. ZJ-2008003 | -Soft coral Sarcophyton sp. | - | [49] | |
| Stemphylium sp. 33231 | -Mangrove tree Bruguiera sexangula var. rhynchopetala. | -Antibacterial activity. | [50] | |
| S. lycopersici | -Inner tissue of gorgonian soft coral Dichotella gammacea. | -Cytotoxic activity. | [51] | |
| Sporendonema casei HDN16-802 | -Marine sediment. | -Anti-Mycobacterium tuberculosis activity. | [109] | |
| Altersolanol C (171) | Altenaria sp. ZJ-2008003 | -Soft coral Sarcophyton sp. | -Antibacterial and cytotoxic activities. | [49] |
| Stemphylium sp. 33231 | -Mangrove tree B. sexangula var. rhynchopetala. | - | [50] | |
| Altersolanol A (172) | Stemphylium sp. 33231 | -Mangrove tree B. sexangula var. rhynchopetala. | -Antibacterial activity. | [50] |
| S. lycopersici | -Inner tissue of gorgonian soft coral Dichotella gammacea. | -Cytotoxic activity. | [51] | |
| Xylaria sp. 2508 | -Mangrove plant. | - | [119] | |
| Auxarthrol C (173) | Stemphylium sp. 33231 | -Mangrove tree Bruguiera sexangula var. rhynchopetala | -Antibacterial activity. | [50] |
| S. lycopersici | -Inner tissue of gorgonian soft coral Dichotella gammacea. | - | [51] | |
| 2-O-Acetylaltersolanol B (174) | Stemphylium sp. 33231 | -Mangrove tree B. sexangula var. rhynchopetala. | -Antibacterial activity. | [50] |
| Lentisone (175) | Trichoderma sp. (H-1) | -Sea cucumber. | -Antibacterial and antiangiogenic activities. | [43] |
| SZ-685C (known as 1403C; 176) | Halorosellinia sp. (no. 1403) | -Mangrove plant. | -Anti-proliferative activity. | [108] |
| - | - | -Cytotoxic activity and induction of cell apoptosis. | [132,133] | |
| (2R,3S)-7-Ethyl-1,2,3,4-tetrahydro-2,3,8-trihydroxy-6-methoxy-3-methyl-9,10-anthracenedione (177) | Phomopsis sp. PSU-MA214 | -Leaves of a mangrove tree Rhizophora apiculata Griff. Ex T. Anderson. | -Antibacterial and cytotoxic activities. | [52] |
| Auxarthrol D (178) | Sporendonema casei HDN16-802 | -Marine sediment. | -Antibacterial and cytotoxic activities. | [109] |
| Auxarthrol G (179) | S. casei HDN16-802 | -Marine sediment. | -Antibacterial and anticoagulant activities. | [109] |
| 4-Dehydroxyaltersolanol A (180) | S. casei HDN16-802 | -Marine sediment. | -Antibacterial and anticoagulant activities. | [109] |
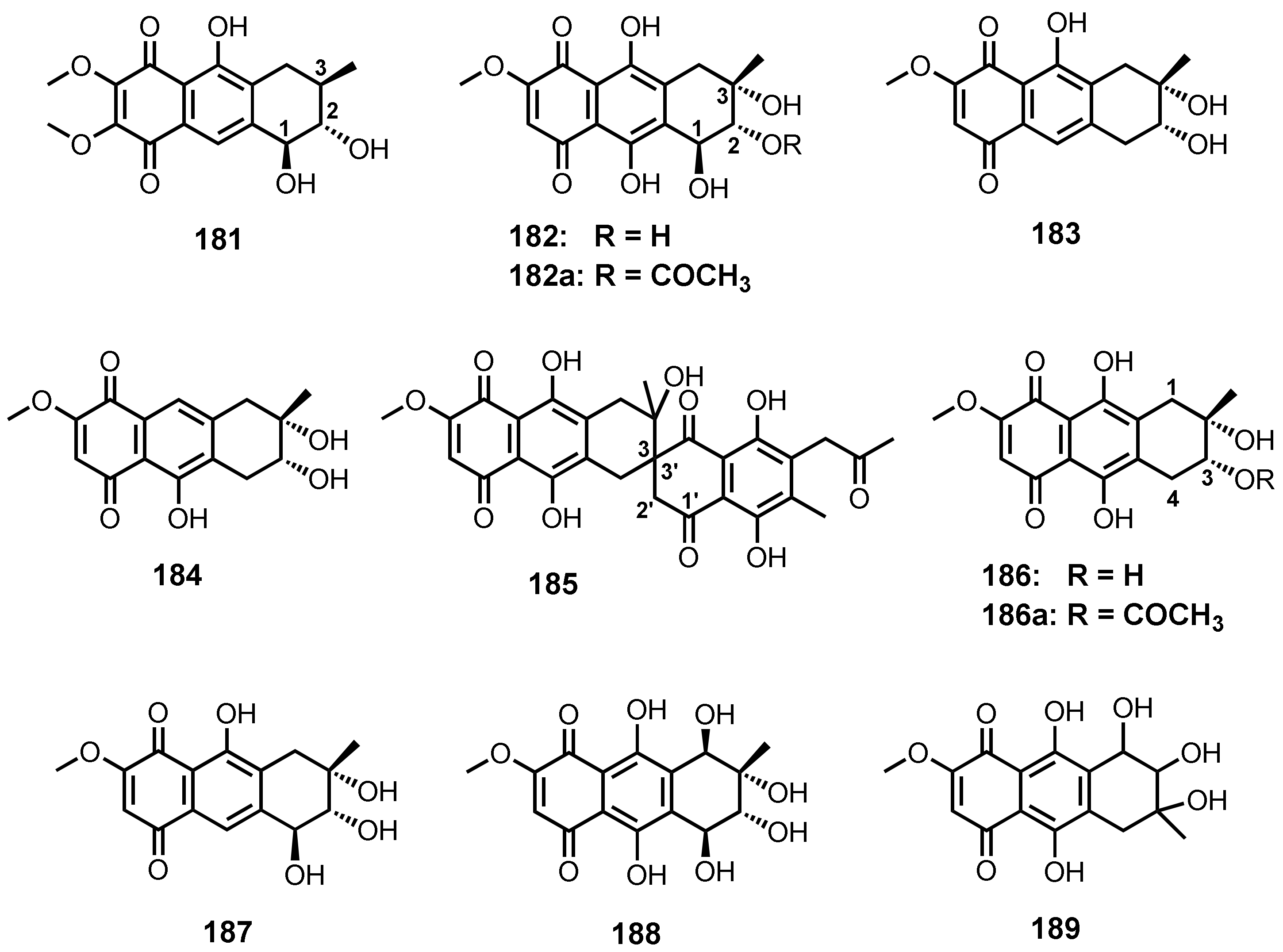
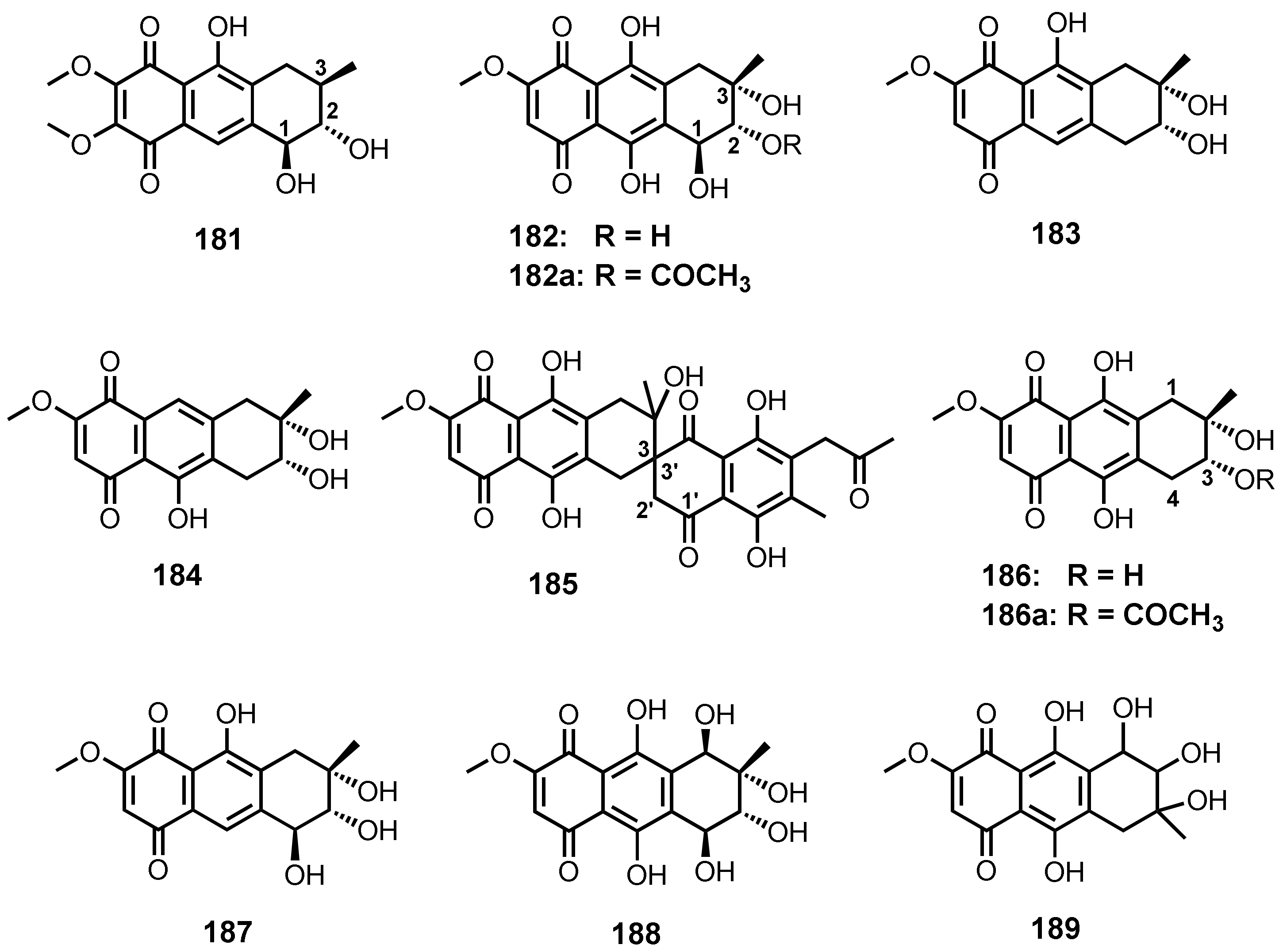
| Aspetritone A (181) | Aspergillus tritici SP2-8-1 | -Soft coral Galaxea fascicularis. | -Antibacterial and cytotoxic activities. | [28] |
| Bostrycin (182) | Aspergillus sp. strain 05F16 | -Unidentified marine alga. | - | [110] |
| Fusarium sp. PSU-F14 and PSU-F135 | -Gorgonian sea fan (Annella sp.) | -Antimalarial and cytotoxic activities. | [59] | |
| Nigrospora sp. (strain no. 1403) | -Decayed wood of Kandelia candel (L.) Druce. | -Antibacterial, antifungal, and cytotoxic activities. | [112] | |
| Nigrospora sp. ZJ-2010006 | -Unidentified sea anemone. | -Antibacterial and cytotoxic activities. | [68] | |
| Xylaria sp. 2508 | -Mangrove plant. | - | [119] | |
| Strain no. 1403 | -Mangrove plant. | -Anti-yeast activity and induction of cell apoptosis. | [111] | |
| - | - | -Cytotoxic activity. | [134] | |
| Nigrosporin A (183) | Fusarium sp. PSU-F14 and PSU-F135 | Gorgonian sea fan (Annella sp.). | - | [59] |
| Nigrosporin B (184) | Fusarium sp. PSU-F14 and PSU-F135 | -Gorgonian sea fan (Annella sp.) | -Antimalarial and anti-Mycobacterium tuberculosis, and cytotoxic activities. | [59,130] |
| Nigrospora sp. ZJ-2010006 | -Unidentified sea anemone. | -Antibacterial and cytotoxic activities. | [68] | |
| Fusarnaphthoquinone C (185) | Fusarium sp. PSU-F14 and PSU-F135 | -Gorgonian sea fan (Annella sp.) | - | [59] |
| 4-Deoxybostrycin (186) | Nigrospora sp. (strain no. 1403) | -Decayed wood of Kandelia candel (L.) Druce. | -Antibacterial, antifungal, and cytotoxic activities. | [112] |
| - | - | -Anti-tumor activity. | [134] | |
| - | - | -Anti-Mycobacterium tuberculosis activity. | [130] | |
| Nigrospora sp. ZJ-2010006 | -Unidentified sea anemone. | -Antibacterial activity. | [68] | |
| Xylaria sp. 2508 | -Mangrove plant. | - | [119] | |
| 10-Deoxybostrycin (187) | Nigrospora sp. ZJ-2010006 | -Unidentified sea anemone. | -Antibacterial and cytotoxic activities. | [68] |
| Nigrospora sp. ZJ-2010006 | -Inner tissue of the zoathid Palythoa haddoni (GX-WZ-20100026). | - | [69] | |
| Hydroxybostrycin (188) | Altenaria sp. (SK11) | -Root of mangrove tree Excoecaria agallocha. | - | [62] |
| 1403P-3 (189) | Halorosellinia sp. (no. 1403) | -Mangrove plant. | -Apoptosis in cancer cells. | [113] |
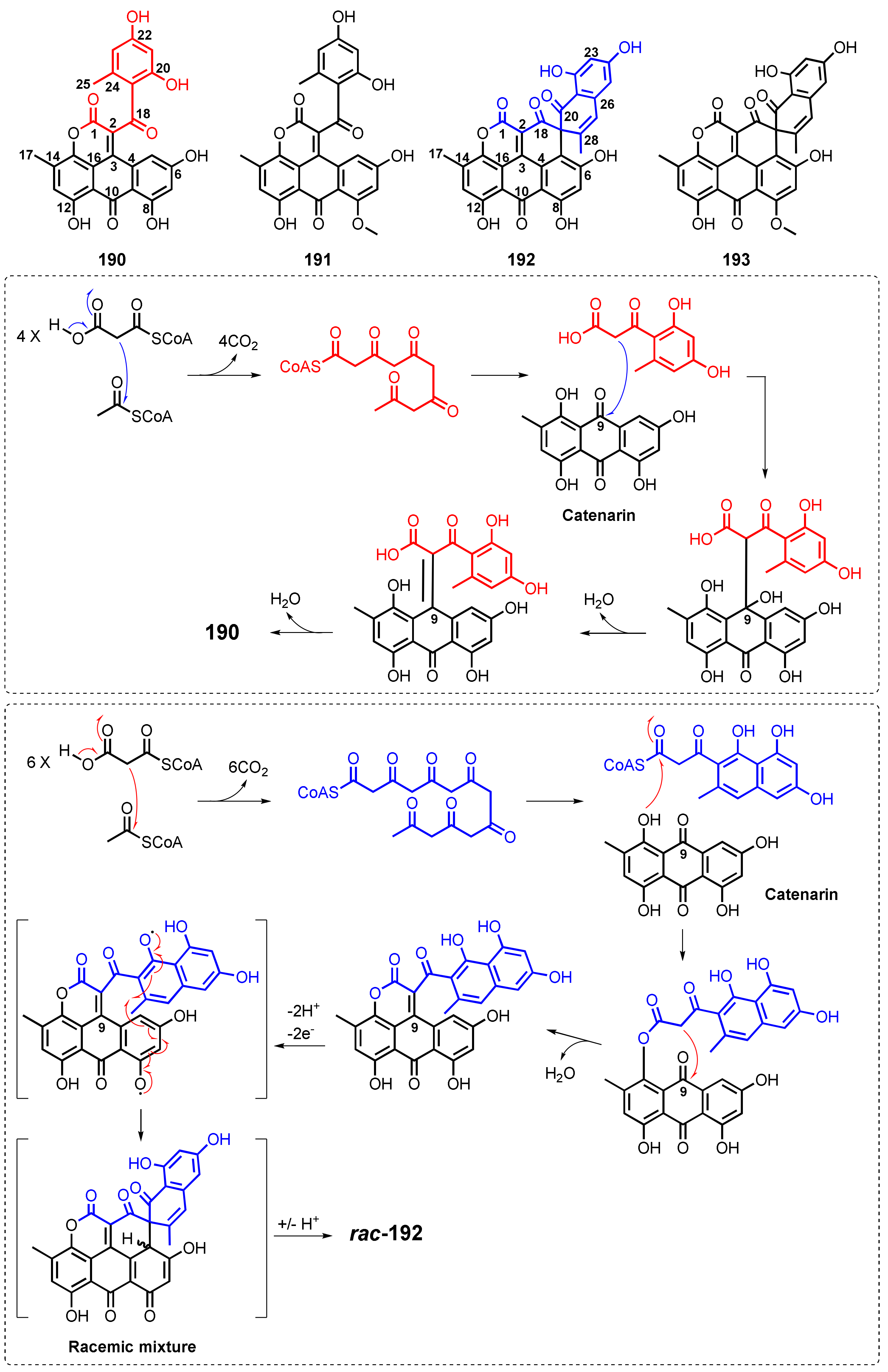
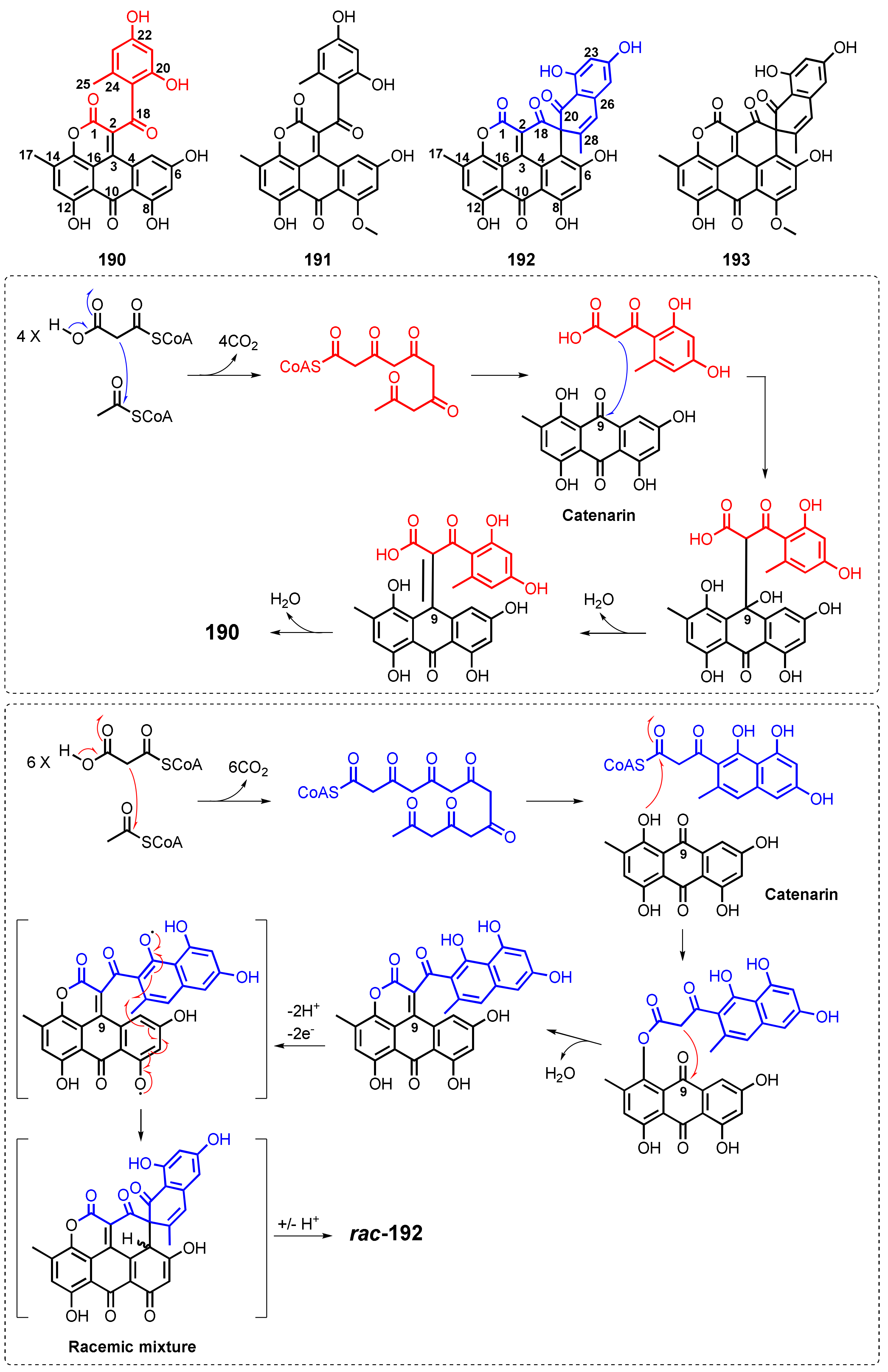
| Aspergiolide A (190) | A. glaucus HB1-19 | -Marine sediment. | -Cytotoxic activity. | [114] |
| Aspergiolide B (191) | A. glaucus HB1-19 | -Marine sediment. | -Cytotoxic activity. | [22] |
| Aspergiolide C (192) | A. glaucus HB1-19 | -Marine sediment. | -Inhibition of receptor tyrosine kinases and anti-parasite activities. | [115] |
| Aspergiolide D (193) | A. glaucus HB1-19 | -Marine sediment. | -Inhibition of receptor tyrosine kinases and anti-parasite activity. | [115] |
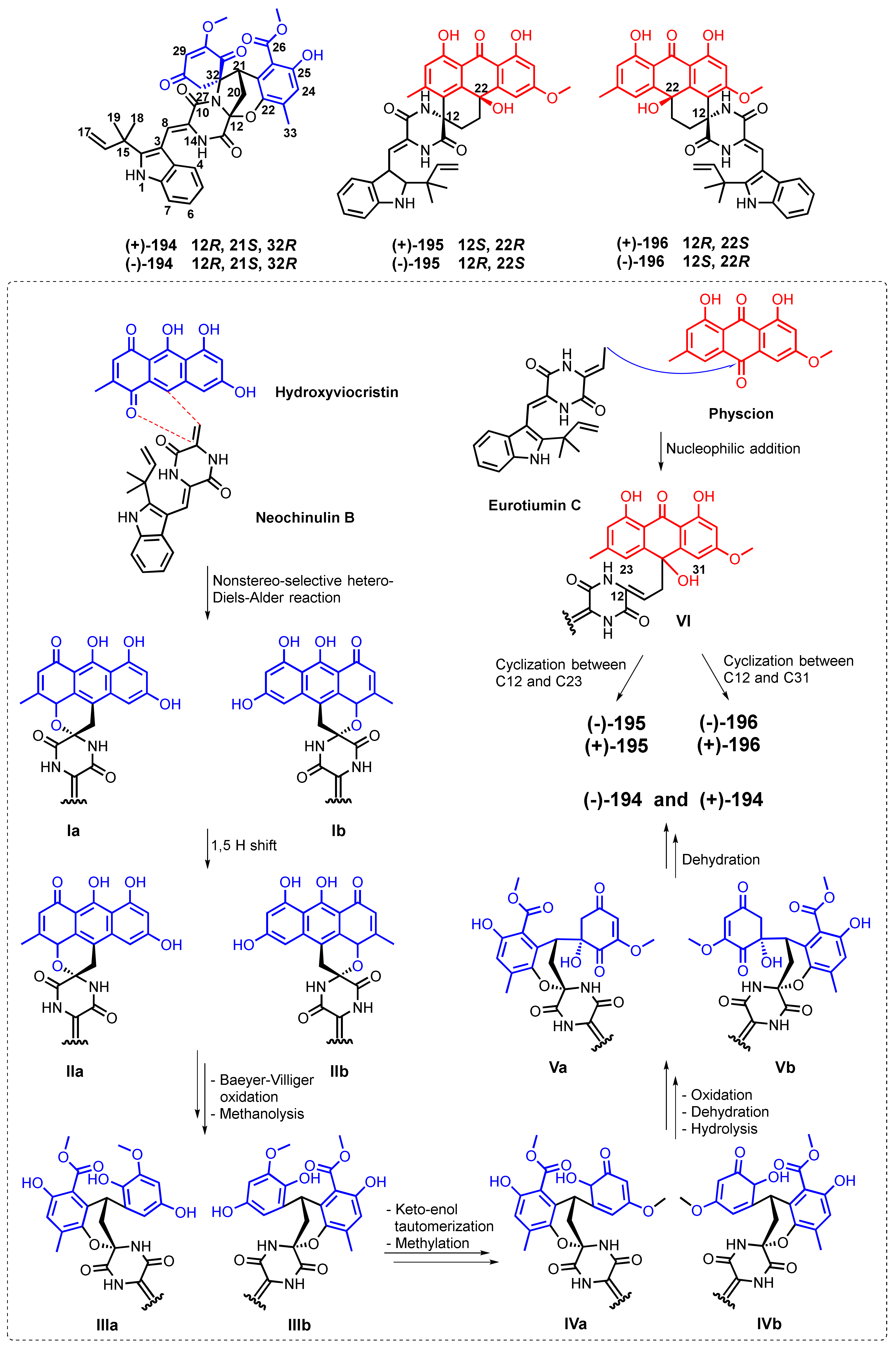
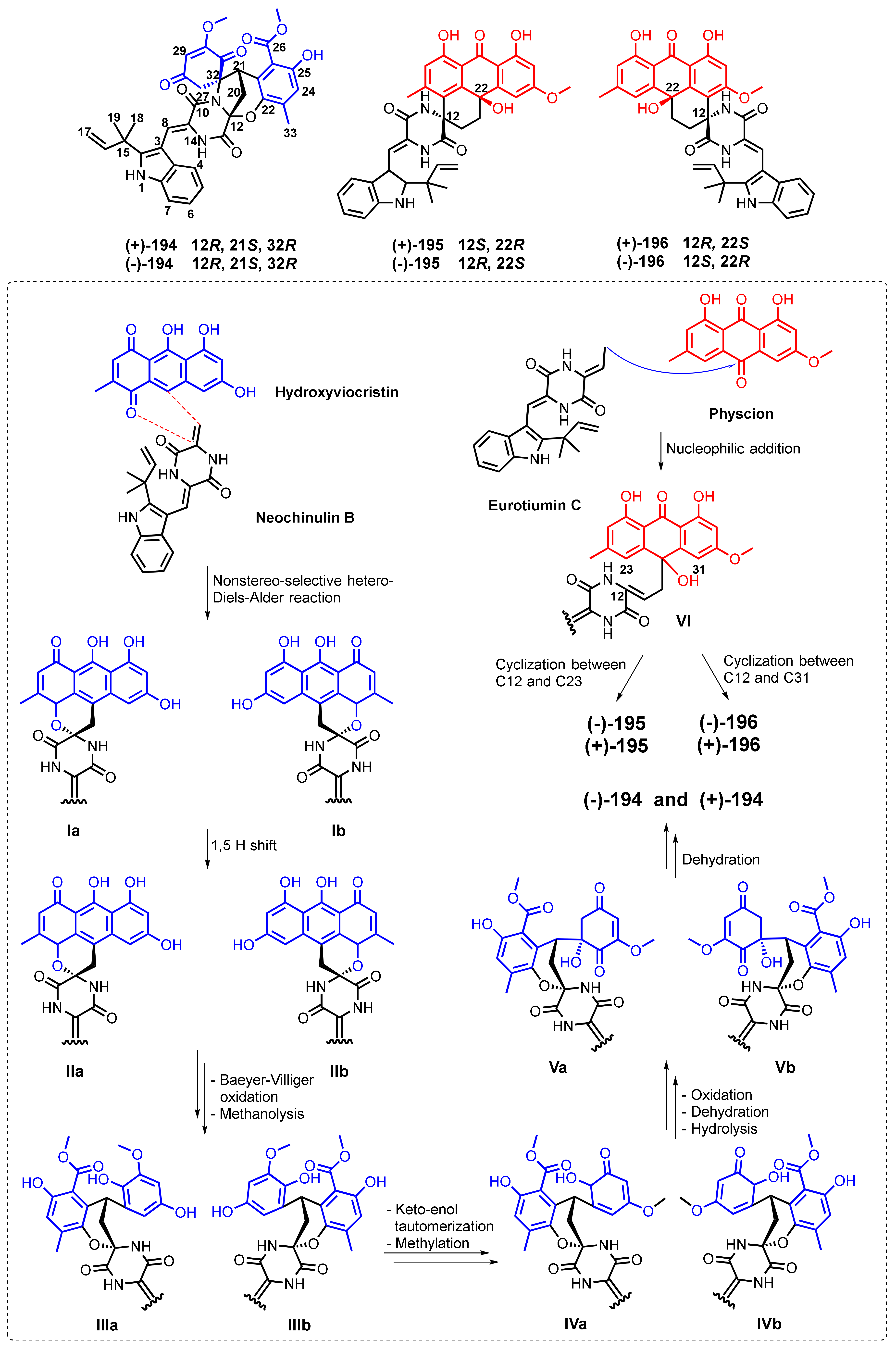
| Variecolortin A (194) | Eurotium sp. SCSIO F452 | -Marine sediment. | -DPPH• radicals scavenging activity. | [116] |
| Variecolortin B (195) | Eurotium sp. SCSIO F452 | -Marine sediment. | -Cytotoxic activity. | [116] |
| Variecolortin C (196) | Eurotium sp. SCSIO F452 | -Marine sediment. | -Cytotoxic activity. | [116] |
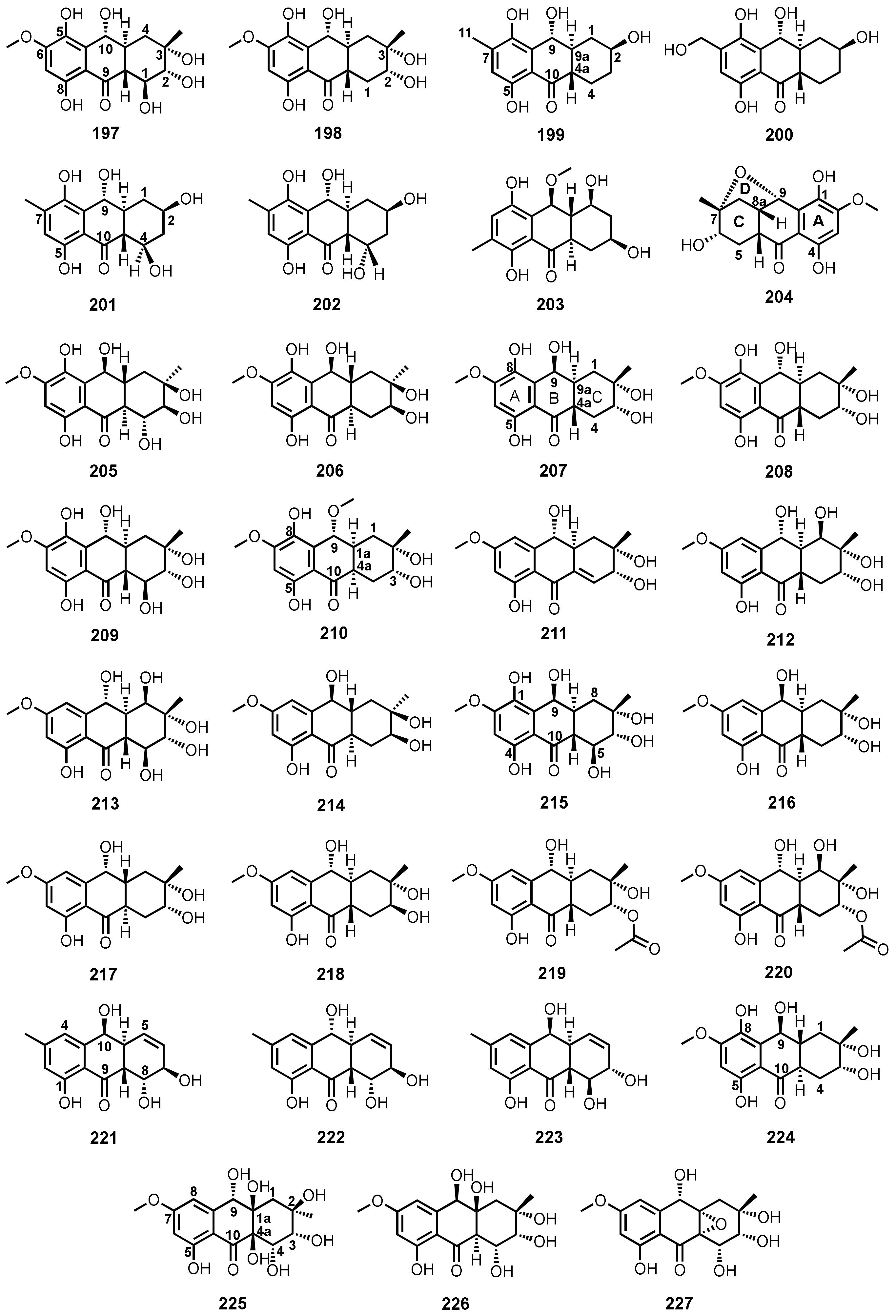
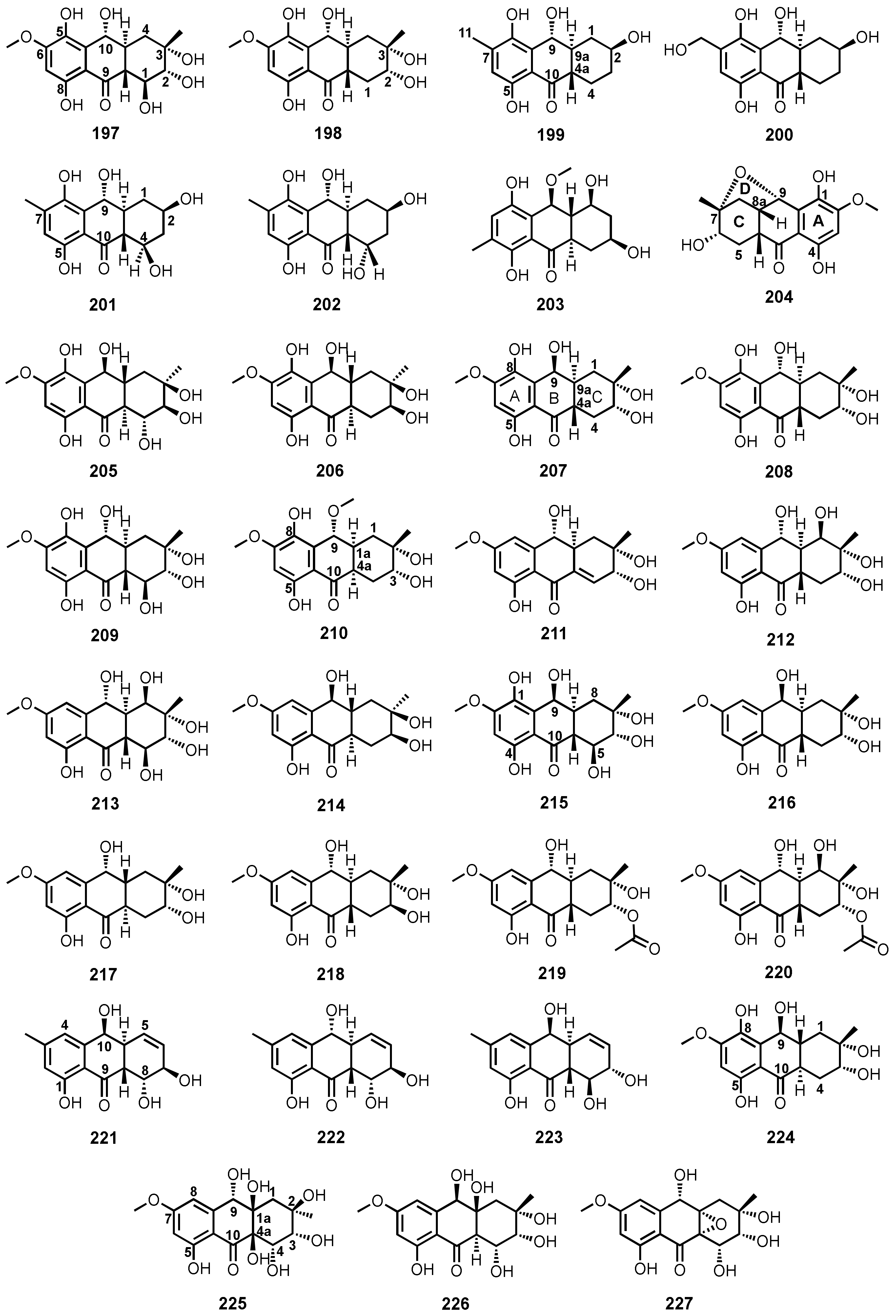
| Tetrahydrobostrycin (197) | Aspergillus sp. strain 05F16 | -Unidentified marine alga. | -Antibacterial activity. | [110] |
| Altenaria sp. (SK11) | -Root of mangrove tree Excoecaria agallocha. | - | [62] | |
| 1-Deoxytetrahydrobostrycin (198) | Aspergillus sp. strain 05F16 | -Unidentified marine alga. | -Antibacterial activity. | [110] |
| 8-Hydroxyconiothyrinone B (199) | Talaromyces islandicus EN-501 | -Inner tissue of marine red alga Laurencia okamurai. | -Antibacterial, DPPH•, and ABTS+• radicals scavenging activities. | [117] |
| 8,11-Dihydroxyconiothyrinone B (200) | T. islandicus EN-501 | -Inner tissue of marine red alga L. okamurai. | -Antibacterial, cytotoxic, DPPH•, and ABTS+• radicals scavenging activities. | [117] |
| 4R,8-Dihydroxyconiothyrinone B (201) | T. islandicus EN-501 | -Inner tissue of marine red alga L. okamurai. | -Antibacterial, DPPH•, and ABTS+• radicals scavenging activities. | [117] |
| 4S,8-Dihydroxyconiothyrinone B (202) | T. islandicus EN-501 | -Inner tissue of marine red alga L. okamurai. | -Antibacterial, DPPH•, and ABTS+• radicals scavenging activities. | [117] |
| 4S,8-Dihydroxy-10-O-methyldendroyl E (203) | T. islandicus EN-501 | -Inner tissue of marine red alga L. okamurai. | -Antibacterial, DPPH•, and ABTS+• radicals scavenging activities. | [117] |
| Fusaquinon A (204) | Fusarium sp. (no. ZH-210) | -Mangrove sediment. | -Cytotoxic activity. | [118] |
| Fusaquinon B (205) | Fusarium sp. (no. ZH-210) | -Mangrove sediment. | -Cytotoxic activity. | [118] |
| Fusaquinon C (206) | Fusarium sp. (no. ZH-210) | -Mangrove sediment. | -Cytotoxic activity. | [118] |
| Fusaranthraquinone (207) | Fusarium sp. PSU-F14 and PSU-F135 | -Gorgonian sea fan (Annella sp.) | - | [59] |
| 9α-Hydroxydihydrodesoxybostrycin (208) | Altenaria sp. (SK11) | -Root of mangrove tree Excoecaria agallocha. | - | [62] |
| Fusarium sp. PSU-F14 and PSU-F135 | -Gorgonian sea fan (Annella sp.) | -Antibacterial and cytotoxic activities. | [59] | |
| Nigrospora sp. ZJ-2010006 | -Unidentified sea anemone. | -Antibacterial activity. | [68] | |
| 9α-Hydroxyhalorosellinia A (209) | Fusarium sp. PSU-F14 and PSU-F135 | -Gorgonian sea fan (Annella sp.) | -Anti-leismanial, antimalarial, anti-Mycobacterium tuberculosis, and cytotoxic activities. | [59] |
| Nigrospora sp. ZJ-2010006 | -Unidentified sea anemone. | -Antibacterial and cytotoxic activities. | [68] | |
| 4a-epi-9α-Methoxydihydrodeoxybostrycin (210) | Nigrospora sp. ZJ-2010006 | -Unidentified sea anemone. | - | [68] |
| Nigrospora sp. ZJ-2010006 | -Inner tissue of the zoathid Palythoa haddoni (GX-WZ-20100026). | - | [69] | |
| Dihydroaltersolanol A (211) | Altenaria tenuissima DFFSCS013 | -Marine sediment. | - | [63] |
| Altenaria sp. ZJ-2008003 | -Soft coral Sarcophyton sp. | - | [49] | |
| Stemphylium sp. 33231 | -Mangrove tree Bruguiera sexangula var. rhynchopetala. | - | [50] | |
| Altersolanol L (212) | A. tenuissima DFFSCS013 | -Marine sediment. | - | [63] |
| Altenaria sp. ZJ-2008003 | -Soft coral Sarcophyton sp. | - | [49] | |
| Stemphylium sp. 33231 | -Mangrove tree B. sexangula var. rhynchopetala. | - | [50] | |
| Phoma sp. L28 | -Mangrove plant Myoporum bontioides A. Gray. | -Anti-fungal activity. | [67] | |
| Ampelanol (213) | Altenaria tenuissima DFFSCS013 | -Marine sediment. | - | [63] |
| Altenaria sp. ZJ-2008003 | -Soft coral Sarcophyton sp. | - | [49] | |
| Stemphylium sp. 33231 | -Mangrove tree Bruguiera sexangula var. rhynchopetala. | - | [50] | |
| S. lycopersici | -Inner tissue of gorgonian soft coral Dichotella gammacea. | - | [51] | |
| Phoma sp. L28 | -Mangrove plant Myoporum bontioides A. Gray. | - | [67] | |
| Phomopsis sp. PSU-MA214 | -Leaves of a mangrove tree Rhizophora apiculata Griff. Ex T. Anderson. | - | [52] | |
| Tetrahydroaltersolanol B (214) | Altenaria sp. ZJ9-6B | -Fruits of a mangrove tree Aegiceras corniculatum. | - | [61] |
| Altenaria sp. ZJ-2008003 | -Soft coral Sarcophyton sp. | - | [49] | |
| Stemphylium sp. 33231 | -Mangrove tree Bruguiera sexangula var. rhynchopetala. | -Antibacterial activity. | [50] | |
| Phoma sp. L28 | -Mangrove plant Myoporum bontioides A. Gray. | -Antifungal activity. | [67] | |
| Phomopsis sp. PSU-MA214 | -Leaves of a mangrove tree R. apiculata Griff. Ex T. Anderson. | - | [52] | |
| Halorosellinia A (215) | Altenaria sp. (SK11) | -Root of mangrove tree Excoecaria agallocha. | - | [62] |
| Halorosellinia sp. (no. 1403) | -Mangrove plant. | - | [58] | |
| Tetrahydroaltersolanol C (216) | Altenaria sp. ZJ-2008003 | -Soft coral Sarcophyton sp. | - | [49] |
| - | - | -Antiviral activity. | [107] | |
| Phomopsis sp. PSU-MA214 | -Leaves of a mangrove tree Rhizophora apiculata Griff. Ex T. Anderson. | - | [52] | |
| Tetrahydroaltersolanol D (217) | Altenaria sp. ZJ-2008003 | -Soft coral Sarcophyton sp. | - | [49] |
| Tetrahydroaltersolanol E (218) | Altenaria sp. ZJ-2008003 | -Soft coral Sarcophyton sp. | - | [49] |
| Tetrahydroaltersolanol F (219) | Altenaria sp. ZJ-2008003 | -Soft coral Sarcophyton sp. | - | [49] |
| 2-O-acetylaltersolanol L (220) | Stemphylium sp. 33231 | -Mangrove tree Bruguiera sexangula var. rhynchopetala. | - | [50] |
| Harzianumnone A (221) | Trichoderma harzianum (XS-20090075) | -Soft coral | - | [55] |
| Harzianumnone B (222) | T. harzianum (XS-20090075) | -Soft coral. | - | [55] |
| Coniothyrinone A (223) | Trichoderma sp. (H-1) | -Sea cucumber. | -Antibacterial and antiangiogenic activities. | [43] |
| Xlyanthraquinone (224) | Xylaria sp. 2508 | -Mangrove plant. | - | [119] |
| Auxarthrol E (225) | Sporendonema casei HDN16-802 | -Marine sediment. | -Antibacterial activity. | [109] |
| Auxarthrol F (226) | S. casei HDN16-802 | -Marine sediment. | -Antibacterial and cytotoxic activities. | [109] |
| Auxarthrol H (227) | Sporendonema casei HDN16-802 | -Marine sediment. | -Antibacterial activity. | [109] |
| Asperflavin (228) | A. glaucus HB1-19 | -Marine sediment. | - | [22] |
| Eurotium repens | -Marine sponge Suberites domuncula. | -Cytotoxicity against sex cells. | [24] | |
| E. rubrum | -Inner tissue of mangrove plant Hibiscus tiliaceus. | -DPPH• radicals scavenging activity. | [106] | |
| Eurotium cristatum EN-220 | -Marine brown alga Sargassum thunbergii. | -Antibacterial activity. | [95] | |
| Isoasperflavin (229) | A. glaucus HB1-19 | -Marine sediment. | - | [22] |
| 3,4-Dihydro-3,9-dihydroxy-6,8-dimethoxy-3-methylanthracen-1(2H)-one (230) | A. wentii EN-48 | -Brown alga Sargassum sp. | - | [30] |
| Eurorubrin (231) | Eurotium rubrum | -Inner tissue of mangrove plant H. tiliaceus. | -DPPH• radicals scavenging activity. | [106] |
| E. cristatum EN-220 | -Marine brown alga Sargassum thunbergii. | -Antibacterial activity and cytotoxicity against Brine Shrimp. | [95] | |
| Asperflavin ribofuranoside (232) | E. cristatum EN-220 | -Marine brown alga Sargassum thunbergii. | - | [95] |
| Anthrininone A (233) | Altenaria tenuissima DFFSCS013 | -Marine sediment. | -Induction of intracellular calcium flux in HEK293 cells and inhibition of indoleamine 2,3-dioxygenase activity. | [63] |
| Scorpinone (234) | Amorosia littoralis | -Inertial sediment. | - | [120] |
| Bispora-like tropical fungus | -Inertial sediment. | - | [121] | |
| Bostrycoidin (235) | - | - | - | [120] |
| 8-O-Methylbostrycoidin (236) | A. terreus (no. GX7-3B) | -Mangrove Bruguiera gymnoihiza (Linn.) Savigny. | -Anti-acetylcholinesterase activity. | [123] |
| 6,6′-Oxybis(1,3,8-trihydroxy-2-((S)-1-methoxyhexyl)anthracene-9,10-dione) (237) | A. versicolor | -The inner tissue of an unidentified marine clam. | -Antibacterial activity. | [88] |
| 6,6′-Oxybis(1,3,8-trihydroxy-2-((S)-1-hydroxyhexyl)anthracene-9,10-dione) (238) | A. versicolor | -The inner tissue of an unidentified marine clam. | -Antibacterial activity. | [88] |
| 2,2′-bis-(7-methyl-1,4,5-trihydroxy-anthracene-9,10-dione) (239) | Talaromyces stipitatus KUFA0207 | -Marine sponge Stylissa flabelliformis. | - | [33] |
| Alterporriol K (240) | Altenaria sp. ZJ9-6B | -Mangrove tree Aegiceras corniculatum fruits. | -Cytotoxic activity. | [61] |
| Alterporriol L (241) | Altenaria sp. ZJ9-6B | -Mangrove tree A. corniculatum fruits. | -Cytotoxic activity. | [61,135] |
| Alterporriol M (242) | Altenaria sp. ZJ9-6B | -Mangrove tree A. corniculatum fruits. | - | [61] |
| Alterporriol S (243) | Altenaria sp. (SK11) | -Root of mangrove tree Excoecaria agallocha. | -Anti-Mycobacterium tuberculosis activity. | [62] |
| (+)-aS-Alterporriol C (244) | Altenaria sp. (SK11) | -Root of mangrove tree E. agallocha. | -Anti-Mycobacterium tuberculosis activity. | [62] |
| Alterporriol C (245) | Altenaria sp. ZJ-2008003 | -Soft coral reef Sarcophyton sp. | -Antibacterial and cytotoxic activities. | [49] |
| Stemphylium sp. 33231 | -Mangrove tree Bruguiera sexangula var. rhynchopetala. | -Antibacterial activity. | [50] | |
| Alterporriol N (246) | Altenaria sp. ZJ-2008003 | -Soft coral Sarcophyton sp. | - | [49] |
| Stemphylium sp. 33231 | -Mangrove tree B. sexangula var. rhynchopetala. | - | [50] | |
| Stemphulium sp. FJJ006 | -Unidentified sponge. | -Anti-inflammatory activity. | [124] | |
| Alterporriol O (247) | Altenaria sp. ZJ-2008003 | -Soft coral Sarcophyton sp. | - | [49] |
| Alterporriol P (248) | Altenaria sp. ZJ-2008003 | -Soft coral Sarcophyton sp. | -Cytotoxic activity. | [49] |
| Alterporriol Q (249) | Altenaria sp. ZJ-2008003 | -Soft coral Sarcophyton sp. | - | [49] |
| Stemphylium sp. 33231 | -Mangrove tree B. sexangula var. rhynchopetala. | - | [50] | |
| Alterporriol R (250) | Altenaria sp. ZJ-2008003 | -Soft coral Sarcophyton sp. | - | [49] |
| Stemphylium sp. 33231 | -Mangrove tree B. sexangula var. rhynchopetala. | - | [50] | |
| Nigrodiquinone A (251) | Nigrospora sp. ZJ-2010006 | -Inner tissue of the zoathid Palythoa haddoni (GX-WZ-20100026). | - | [69] |
| Cytoskyrin A (252) | Curvularia lunata | -Marine sponge Niphates olemda. | -Antibacterial activity. | [64] |
| Alterporriol A (253) | Stemphylium sp. 33231 | -Mangrove tree B. sexangula var. rhynchopetala. | - | [50] |
| Alterporriol B (254) | Stemphylium sp. 33231 | -Mangrove tree B. sexangula var. rhynchopetala. | -Antibacterial activity. | [50] |
| Alterporriol D (255) | Stemphylium sp. 33231 | -Mangrove tree B. sexangula var. rhynchopetala. | -Antibacterial activity. | [50] |
| Alterporriol E (256) | Stemphylium sp. 33231 | -Mangrove tree B. sexangula var. rhynchopetala. | -Antibacterial activity. | [50] |
| Alterporriol T (257) | Stemphylium sp. 33231 | -Mangrove tree B. sexangula var. rhynchopetala. | - | [50] |
| Alterporriol U (258) | Stemphylium sp. 33231 | -Mangrove tree B. sexangula var. rhynchopetala. | -Antibacterial activity. | [50] |
| Alterporriol V (259) | Stemphylium sp. 33231 | -Mangrove tree B. sexangula var. rhynchopetala. | -Antibacterial activity. | [50] |
| Alterporriol W (260) | Stemphylium sp. 33231 | -Mangrove tree B. sexangula var. rhynchopetala. | - | [50] |
| Alterporriol Y (261) | S. lycopersici | -Inner tissue of gorgonian soft coral Dichotella gammacea. | - | [51] |
| Alterporriol F (262) | Stemphulium sp. FJJ006 | -Unidentified sponge. | -Anti-inflammatory activity. | [124] |
| Alterporriol G (263) | Stemphulium sp. FJJ006 | -Unidentified sponge. | -Anti-inflammatory activity. | [124] |
| Alterporriol Z1 (264) | Stemphulium sp. FJJ006 | -Unidentified sponge. | -Anti-inflammatory activity. | [124] |
| Alterporriol Z2 (265) | Stemphulium sp. FJJ006 | -Unidentified sponge. | -Anti-inflammatory activity. | [124] |
| Alterporriol Z3 (266) | Stemphulium sp. FJJ006 | -Unidentified sponge. | - | [124] |
| Rubellin A (267) | Strain F-F-3C | -Unidentified marine red alga. | -Antibacterial activity. | [44] |
| 14-Acetoxyrubellin A (268) | Strain F-F-3C | -Unidentified marine red alga. | -Antibacterial and antifungal activities. | [44] |
| 14-Acetoxyrubellin C (269) | Strain F-F-3C | -Unidentified marine red alga. | -Antibacterial activity. | [44] |
| Physcion-10,10′-bianthrone (270) | A. glaucus HB1-19 | -Deep-sea sediment. | - | [22] |
| A. wentii EN-48 | -Brown alga Sargassum sp. | - | [30] | |
| trans-Emodin-physcion bianthrone (271) | A. glaucus HB1-19 | -Deep-sea sediment. | -Cytotoxic activity. | [22] |
| cis-Emodin-physcion bianthrone (272) | A. glaucus HB1-19 | -Deep-sea sediment. | -Cytotoxic activity. | [22] |
| Atropisomer of 8,8′-dihydroxy-1,1′,3,3′-tetramethoxy-6,6′-dimethyl-10,10-bianthrone (273) | A. wentii EN-48 | -Brown alga Sargassum sp. | - | [30] |
| Atropisomer of 8,8′-dihydroxy-1,1′,3,3′-tetramethoxy-6,6′-dimethyl-10,10-bianthrone (274) | A. wentii EN-48 | -Brown alga Sargassum sp. | - | [30] |
| Allianthrone A (275) | A. alliaceus | -Marine algae. | -Cytotoxic activity. | [77] |
| Allianthrone B (276) | A. alliaceus | -Marine algae. | -Cytotoxic activity. | [77] |
| Allianthrone C (277) | A. alliaceus | -Marine algae. | -Cytotoxic activity. | [77] |
| (±)-Eurotone A (278) | Eurotium sp. SCSIO F452 | -Marine sediment. | - | [70] |
| JBIR-97/98 (279) | Engyodontium album | -Marine sponge Cacospinga scalaris. | -Antibacterial, antifungal, and cytotoxic activities. | [125] |
| JBIR-99 (280) | E. album | -Marine sponge C. scalaris. | -Antibacterial, antifungal, and cytotoxic activities. | [125] |
| Engyodontochone A (281) | E. album | -Marine sponge C. scalaris. | -Antibacterial, antifungal, and cytotoxic activity. | [125] |
| Engyodontochone B (282) | E. album | -Marine sponge C. scalaris. | -Antibacterial, antifungal, and cytotoxic activities. | [125] |
| Engyodontochone C (283) | E. album | -Marine sponge C. scalaris. | -Antibacterial and cytotoxic activities. | [125] |
| Engyodontochone D (284) | E. album | -Marine sponge C. scalaris. | - | [125] |
| Engyodontochone E (285) | E. album | -Marine sponge C. scalaris. | -Antibacterial activity. | [125] |
| Engyodontochone F (286) | Engyodontium album | -Marine sponge C. scalaris. | -Antibacterial activity. | [125] |
| Acremonidin A (287) | Acremonium camptosporum | -Marine sponge Aplysina fulva. | -Antibacterial and cytotoxic activities. | [126] |
| Unidentified fungus of the order Hypocreales (MSX 17022) | - | -20S proteasome inhibitory activity. | [127] | |
| Acremonidin B (288) | A. camptosporum | -Marine sponge A. fulva. | -Antibacterial and cytotoxic activities. | [126] |
| Acremonidin C (289) | A. camptosporum | -Marine sponge A. fulva. | -Antibacterial and cytotoxic activities. | [126] |
| Unidentified fungus of the order Hypocreales (MSX 17022) | - | -20S proteasome inhibitory activity. | [127] | |
| Acremonidin G (290) | A.camptosporum | -Marine sponge A. fulva. | -Antibacterial and cytotoxic activities. | [126] |
| Acremoxanthone A (291) | A. camptosporum | -Marine sponge A. fulva. | -Antibacterial and cytotoxic activities. | [126] |
| Acremoxanthone B (292) | A. camptosporum | -Marine sponge A. fulva. | -Antibacterial and cytotoxic activities. | [126] |
| Acremoxanthone D (293) | A. camptosporum | -Marine sponge A. fulva. | -Antibacterial and cytotoxic activities. | [126] |
| Unidentified fungus of the order Hypocreales (MSX 17022) | - | -20S proteasome inhibitory activity. | [127] | |
| Acremoxanthone F (294) | A. camptosporum | -Marine sponge A. fulva. | -Antibacterial and cytotoxic activities. | [126] |
| Acremoxanthone G (295) | A. camptosporum | -Marine sponge A. fulva. | -Antibacterial and cytotoxic activities. | [126] |
| Acremoxanthone C (296) | Unidentified fungus of the order Hypocreales (MSX 17022) | - | -20S proteasome inhibitory activity. | [127] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hafez Ghoran, S.; Taktaz, F.; Ayatollahi, S.A.; Kijjoa, A. Anthraquinones and Their Analogues from Marine-Derived Fungi: Chemistry and Biological Activities. Mar. Drugs 2022, 20, 474. https://doi.org/10.3390/md20080474
Hafez Ghoran S, Taktaz F, Ayatollahi SA, Kijjoa A. Anthraquinones and Their Analogues from Marine-Derived Fungi: Chemistry and Biological Activities. Marine Drugs. 2022; 20(8):474. https://doi.org/10.3390/md20080474
Chicago/Turabian StyleHafez Ghoran, Salar, Fatemeh Taktaz, Seyed Abdulmajid Ayatollahi, and Anake Kijjoa. 2022. "Anthraquinones and Their Analogues from Marine-Derived Fungi: Chemistry and Biological Activities" Marine Drugs 20, no. 8: 474. https://doi.org/10.3390/md20080474
APA StyleHafez Ghoran, S., Taktaz, F., Ayatollahi, S. A., & Kijjoa, A. (2022). Anthraquinones and Their Analogues from Marine-Derived Fungi: Chemistry and Biological Activities. Marine Drugs, 20(8), 474. https://doi.org/10.3390/md20080474