
Therapeutic Potential of Marine Peptides in Prostate Cancer: Mechanistic Insights

 ,
,  ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
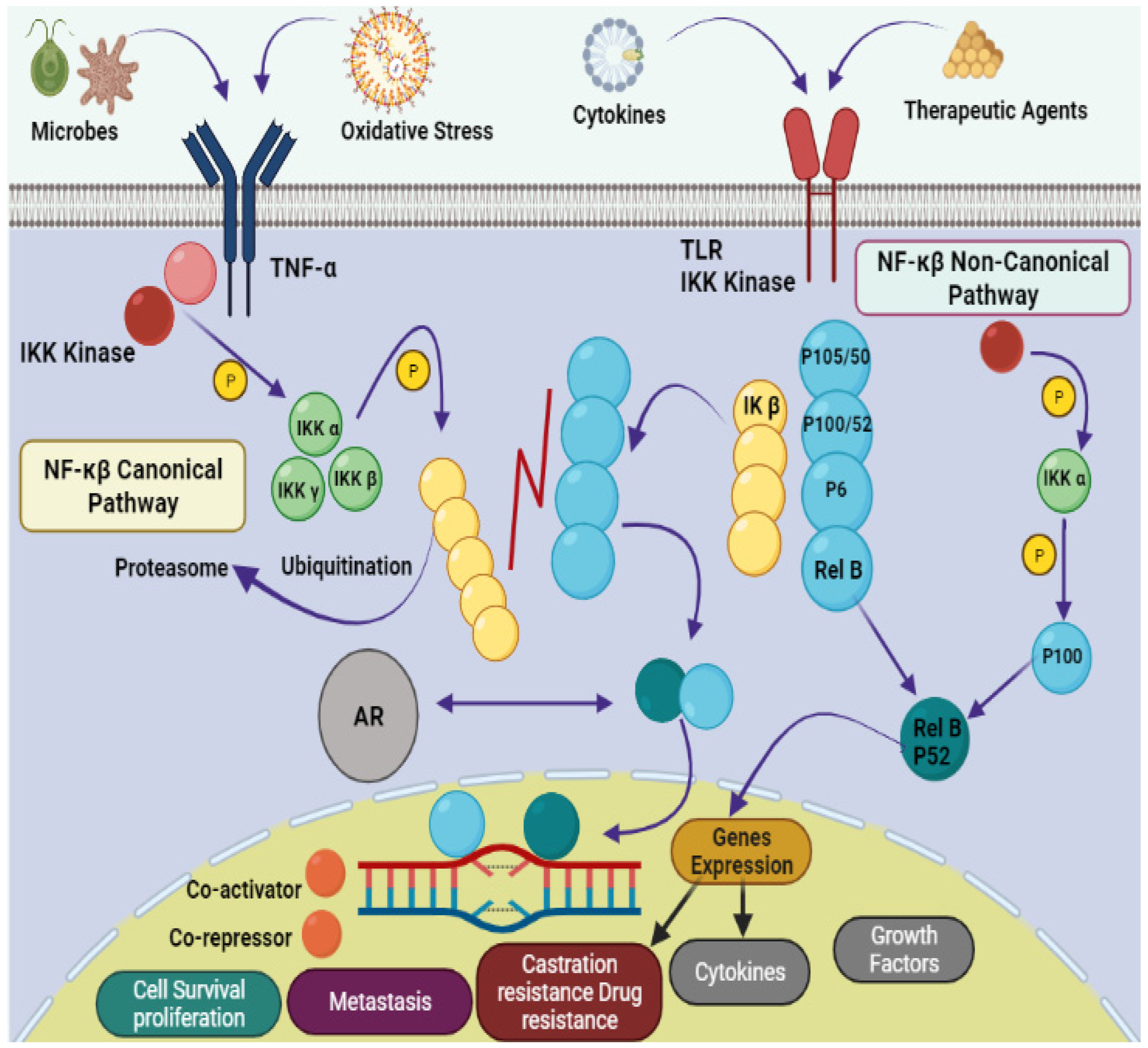
2. Mechanistic Insights
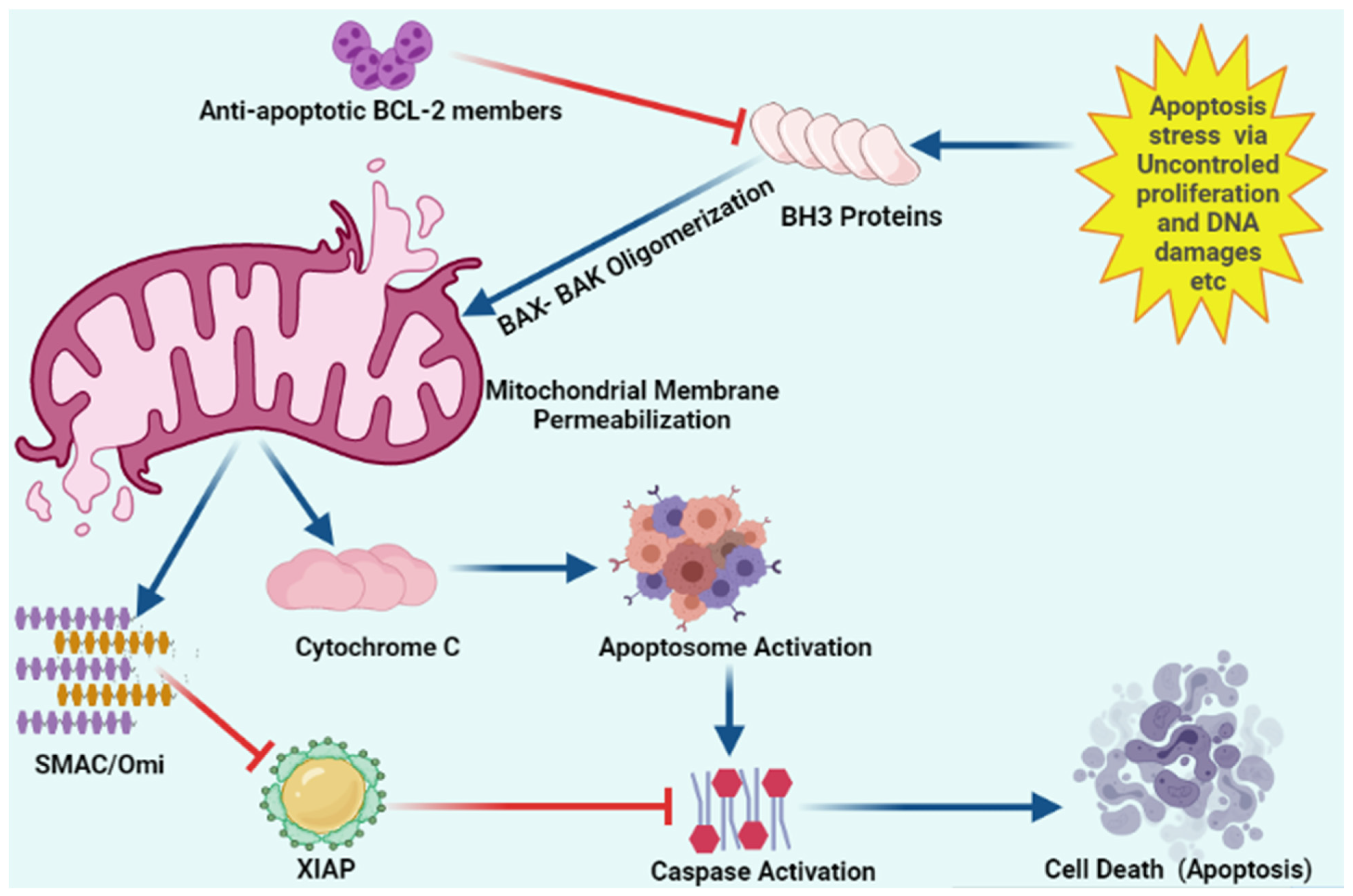
2.1. Apoptosis
2.2. Antimitotic Effect
2.3. Antimetastatic Activity
2.4. Antiangiogenic Effect
2.5. Cell Cycle Arrest
2.6. p53 Upregulation
2.7. Stimulation of Histone Hyperacetylation
2.8. Mitochondrial Dysfunctions and Oxidative Damage
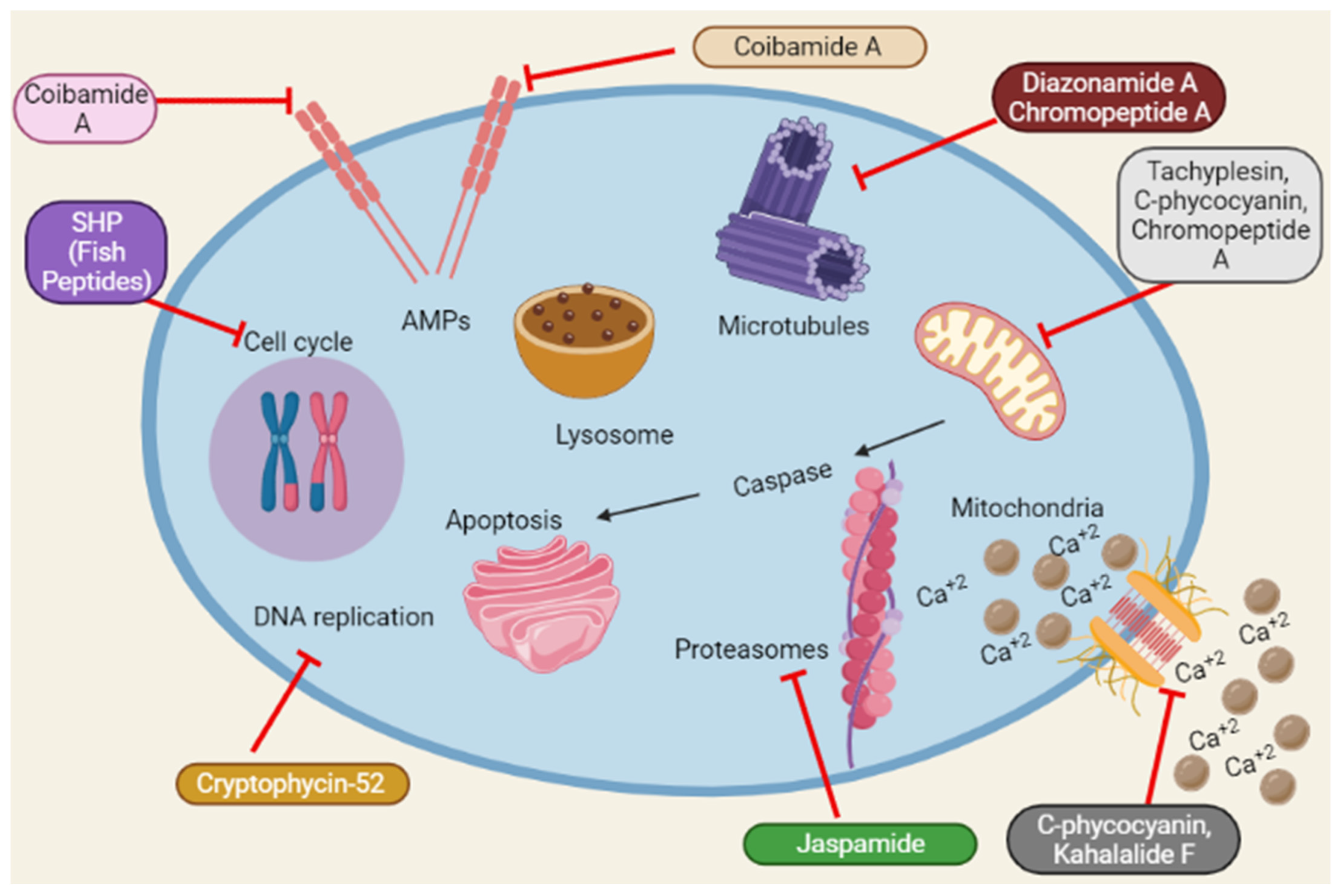
2.9. Unidentified Mechanisms for Anticancer Activity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptides | Marine Sources (Species Name) | Active Derivative | Anticancer Mechanisms | References |
|---|---|---|---|---|
| Aurilide B | Cyanobacteria (Lyngbya majuscula) | Cyclic depsipeptide | Microtubule stabilization | [22] |
| Lagunamide C | ↓ cell viability | [23] | ||
| Cryptophycin-52 (LY355703) | Cyanobacteria (Nostoc sp.) | DNA fragmentation; Bcl2 ↓; Bax ↑; Bcl-xL ↓; caspase 3, 7 ↑; PARP ↑; p53 ↑; G2/M phase arrest; Microtubule depolymerization | [24] | |
| Coibamide A | Cyanobacteria (Leptolyngbya sp.) | ↓ cell viability | [25] | |
| Laxaphycin B | Cyanobacteria (Lyngbya majuscula) | Cyclic dodecapeptide | [39] | |
| C-phycocyanin | Cyanobacteria (Limnothrix sp.) | Peptide | Caspases 3, 9 ↑; cyt c release ↑; DNA fragmentation | [48] |
| Bisebromoamide | Cyanobacteria (Lyngbya sp.) | ↓ cancer cell growth | [106] | |
| Jaspamide | Sponge (Jaspis johnstoni ) | Cyclic depsipeptide | Actin filament disruption | [28] |
| [29] | ||||
| Homophymines A–E | Sponge (Homophymia sp.) | ↓ cell viability | [27] | |
| Neamphamides B–D | Sponge (Neamphius huxleyi) | [30] | ||
| Geodiamolides D–F | Sponge (Pipestela candelabra) | [26] | ||
| Milnamides A–G | N-methylated linear peptide | |||
| Rolloamide A | Sponge (Eurypon laughlini) | Cyclic heptapeptide | [36] | |
| HTI-286 | Sponge (Hemiasterella minor) | Tripeptide | Microtubule depolymerization | [14] |
| [107] | ||||
| Kahalalide F | Mollusk (Elysia rufescens) | Cyclic depsipeptide | PI3K-AKT inhibition; ErbB3 depletion | [31] |
| ↓ cancer cell growth | [108] | |||
| Elisidepsin | PI3K-AKT inhibition; ErbB3 depletion | [56] | ||
| Dolastatin 10 | Mollusk (Dolabella auricularia) | Pentapeptide | Microtubule depolymerization | [19] |
| MCH | Mollusk (Mytilus coruscus) | Peptide | Bcl2 ↓; Bax ↑; caspase 3, 9 ↑ | [43] |
| KLH | Mollusk (Megathura crenulata) | ↓ cancer cell growth | [109] | |
| Tamandarin A | Ascidia (Trididemnum solidum) | Cyclic depsipeptide | ↓ cell viability | [32] |
| Tamandarin B | [33] | |||
| Patellamide B | Ascidia (Lissoclinum patella) | Cyclic octapeptide | [38] | |
| Patellamide F | ||||
| Ulithiacyclamide | Cyclic peptide | |||
| Trunkamide A | Ascidia (Lissoclinum sp.) | Cyclic heptapeptide | [12,37] | |
| Diazonamide A | Ascidia (Diazona angulata) | Macrocyclic peptide | Microtubule depolymerization | [64] |
| Chromopeptide A | Bacteria (Chromobacterium sp. HS-13-94) | Depsipeptide | caspase 3 ↑; PARP cleavage; HDAC inhibition; G2/M phase arrest; p53 ↑ | [51] |
| Sansalvamide A | Fungus (Fusarium sp.) | Cyclic depsipeptide | ↓ cell viability | [34] |
| Microsporin A | Fungus (Microsporum cf. gypseum) | Cyclic tetrapeptide | [17] | |
| Zygosporamide | Fungus (Zygosporium masonii) | Cyclic pentadepsipeptide | ↓ cancer cell growth | [35] |
| SHP | Fish (Sepia esculenta) | Tripeptide | Bcl2 ↓; Bax ↑; caspase 3 ↑; p53 ↑ | [15] |
| SIO | Bcl2 ↓; Bax ↑; caspase 3 ↑; p53 ↑; VEGF ↓ | [44,45] | ||
| S and G2/M phase cell cycle arrest | [16] | |||
| TFD | Fish (Gadus sp.) | Peptide | VEGFR1 ↓; MUC1 ↓ | [94] |
| YALPAH | Fish (Setipinna taty) | ↓ cancer cell growth | [105] | |
| YALRAH | ||||
| YALPAR | ||||
| YALPAG | ||||
| ILYMP | Clam (Cyclina sinensis) | Pentapeptide | Bcl2 ↓; Bax ↑; caspase 3, 9 ↑; cyt c release ↑ | [20] |
| SCH-P9 and SCH-P10 | Clam (Sinonovacula constricta) | Tetrapeptide | [18] | |
| AAP-H | Sea anemone (Anthopleura anjunae) | Oligopeptide | [21] | |
| Tachyplesin | Horseshoe crab (Tachypleus tridentatus) | Cyclic peptide | caspase 3, 6, 7, 8, 9 ↑; cyt c release ↑ | [50] |
3. Clinical Trial Status
| Cell lines/ (Peptides) | Phase | Clinical Trials.Gov Identifier | References |
|---|---|---|---|
| DU145 (Soblidotin) | Phase I | NCT00072228 | [22,110,111] |
| Dolastatin 10 | Phase II | NCT00003626 | [23,110] |
| Tasidotin/synthadotin (ILX651) | Phase II | NCT00082134 | [24,113] |
| Dehydrodidemnine B | Phase III | NCT00780975 | [10,25,32] |
| Kahalalide F | Phase I | NCT00106418 | [39,115] |
| Elisidepsin (Irvalec®) | Phase II | NCT00884845 | [48,117] |
4. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Rebello, R.J.; Oing, C.; Knudsen, K.E.; Loeb, S.; Johnson, D.C.; Reiter, R.E.; Gillessen, S.; Van der Kwast, T.; Bristow, R.G. Prostate cancer. Nat. Rev. Dis. Primers 2021, 7, 9. [Google Scholar] [CrossRef] [PubMed]
- Rawla, P. Epidemiology of Prostate Cancer. World J. Oncol. 2019, 10, 63–89. [Google Scholar] [CrossRef] [PubMed]
- Mokbel, K.; Wazir, U.; Mokbel, K. Chemoprevention of Prostate Cancer by Natural Agents: Evidence from Molecular and Epidemiological Studies. Anticancer. Res. 2019, 39, 5231. [Google Scholar] [CrossRef]
- Fontana, F.; Raimondi, M.; Marzagalli, M.; Di Domizio, A.; Limonta, P. Natural Compounds in Prostate Cancer Prevention and Treatment: Mechanisms of Action and Molecular Targets. Cells 2020, 9, 460. [Google Scholar] [CrossRef]
- Alves, C.; Diederich, M. Marine Natural Products as Anticancer Agents. Mar. Drugs 2021, 19, 447. [Google Scholar] [CrossRef]
- Barreca, M.; Spanò, V.; Montalbano, A.; Cueto, M.; Marrero, A.R.D.; Deniz, I.; Erdoğan, A.; Bilela, L.L.; Moulin, C.; Taffin-De-Givenchy, E.; et al. Marine Anticancer Agents: An Overview with a Particular Focus on Their Chemical Classes. Mar. Drugs 2020, 18, 619. [Google Scholar] [CrossRef]
- Ahmed, S.; Mirzaei, H.; Aschner, M.; Khan, A.; Al-Harrasi, A.; Khan, H. Marine peptides in breast cancer: Therapeutic and mechanistic understanding. Biomed. Pharmacother. 2021, 142, 112038. [Google Scholar] [CrossRef]
- Ahmed, S.; Hasan, M.M.; Aschner, M.; Mirzaei, H.; Alam, W.; Shah, S.M.M.; Khan, H. Therapeutic potential of marine peptides in glioblastoma: Mechanistic insights. Cell. Signal. 2021, 87, 110142. [Google Scholar] [CrossRef]
- Kang, H.K.; Choi, M.-C.; Seo, C.H.; Park, Y. Therapeutic Properties and Biological Benefits of Marine-Derived Anticancer Peptides. Int. J. Mol. Sci. 2018, 19, 919. [Google Scholar] [CrossRef]
- Chiangjong, W.; Chutipongtanate, S.; Hongeng, S. Anticancer peptide: Physicochemical property, functional aspect and trend in clinical application (Review). Int. J. Oncol. 2020, 57, 678–696. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.-T.; Liu, Z.-D.; Wang, Z.; Wang, T.; Wang, N.; Wang, N.; Zhang, B.; Zhao, Y.-F. Recent Advances in Small Peptides of Marine Origin in Cancer Therapy. Mar. Drugs 2021, 19, 115. [Google Scholar] [CrossRef] [PubMed]
- Ucak, I.; Afreen, M.; Montesano, D.; Carrillo, C.; Tomasevic, I.; Simal-Gandara, J.; Barba, F. Functional and Bioactive Properties of Peptides Derived from Marine Side Streams. Mar. Drugs 2021, 19, 71. [Google Scholar] [CrossRef]
- Hadaschik, B.; Ettinger, S.; Sowery, R.D.; Zoubeidi, A.; Andersen, R.J.; Roberge, M.; Gleave, M.E. Targeting prostate cancer with HTI-286, a synthetic analog of the marine sponge product hemiasterlin. Int. J. Cancer 2008, 122, 2368–2376. [Google Scholar] [CrossRef]
- Huang, F.; Jing, Y.; Ding, G.; Yang, Z. Isolation and purification of novel peptides derived from Sepia ink: Effects on apoptosis of prostate cancer cell PC-3. Mol. Med. Rep. 2017, 16, 4222–4228. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Yang, Z.; Yu, D.; Wang, J.; Li, R.; Ding, G. Sepia ink oligopeptide induces apoptosis in prostate cancer cell lines via caspase-3 activation and elevation of Bax/Bcl-2 ratio. Mar. Drugs 2012, 10, 2153–2165. [Google Scholar] [CrossRef] [PubMed]
- Gu, W.; Cueto, M.; Jensen, P.R.; Fenical, W.; Silverman, R.B. Microsporins A and B: New histone deacetylase inhibitors from the marine-derived fungus Microsporum cf. gypseum and the solid-phase synthesis of microsporin A. Tetrahedron 2007, 63, 6535–6541. [Google Scholar] [CrossRef]
- Huang, F.; Ding, G.; Yang, Z.; Yu, F. Two novel peptides derived from Sinonovacula constricta inhibit the proliferation and induce apoptosis of human prostate cancer cells. Mol. Med. Rep. 2017, 16, 6697–6707. [Google Scholar] [CrossRef]
- Turner, T.; Jackson, W.H.; Pettit, G.R.; Wells, A.; Kraft, A.S. Treatment of human prostate cancer cells with dolastatin 10, a peptide isolated from a marine shell-less mollusc. Prostate 1998, 34, 175–181. [Google Scholar] [CrossRef]
- Yu, F.; Zhang, Y.; Ye, L.; Tang, Y.; Ding, G.; Zhang, X.; Yang, Z. A novel anti-proliferative pentapeptide (ILYMP) isolated from Cyclina sinensis protein hydrolysate induces apoptosis of DU-145 prostate cancer cells. Mol. Med. Rep. 2018, 18, 771–778. [Google Scholar]
- Wu, Z.-Z.; Ding, G.F.; Huang, F.F.; Yang, Z.S.; Yu, F.M.; Tang, Y.P.; Jia, Y.L.; Zheng, Y.Y.; Chen, R. Anticancer Activity of Anthopleura anjunae Oligopeptides in Prostate Cancer DU-145 Cells. Mar. Drugs 2018, 16, 125. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Gross, H.; Goeger, D.E.; Mooberry, S.L.; Gerwick, W.H. Aurilides B and C, Cancer Cell Toxins from a Papua New Guinea Collection of the Marine Cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2006, 69, 572–575. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.; Puddick, J.; Prinsep, M.R.; Rottmann, M.; Chan, K.P.; Chen, D.Y.K.; Tan, L.T. Lagunamide C, a cytotoxic cyclodepsipeptide from the marine cyanobacterium Lyngbya majuscula. Phytochemistry 2011, 72, 2369–2375. [Google Scholar] [CrossRef]
- Drew, L.; Fine, R.L.; Do, T.N.; Douglas, G.P.; Petrylak, D.P. The Novel Antimicrotubule Agent Cryptophycin 52 (LY355703) Induces Apoptosis via Multiple Pathways in Human Prostate Cancer Cells. Clin. Cancer Res. 2002, 8, 3922. [Google Scholar] [PubMed]
- Medina, R.A.; Goeger, D.E.; Hills, P.; Mooberry, S.L.; Huang, N.; Romero, L.I.; Ortega-Barría, E.; Gerwick, W.H.; McPhail, K.L. Coibamide A, a potent antiproliferative cyclic depsipeptide from the Panamanian marine cyanobacterium Leptolyngbya sp. J. Am. Chem. Soc. 2008, 130, 6324–6325. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.D.; Pham, N.B.; Fechner, G.A.; Hooper, J.N.A.; Quinn, R.J. Potent cytotoxic peptides from the Australian marine sponge Pipestela candelabra. Mar. Drugs 2014, 12, 3399–3415. [Google Scholar] [CrossRef] [PubMed]
- Zampella, A.; Sepe, V.; Bellotta, F.; Luciano, P.; D’Auria, M.V.; Cresteil, T.; Debitus, C.; Petek, S.; Poupat, C.; Ahond, A. Homophymines B–E and A1–E1, a family of bioactive cyclodepsipeptides from the sponge Homophymia sp. Org. Biomol. Chem. 2009, 7, 4037–4044. [Google Scholar] [CrossRef]
- Takeuchi, H.; Ara, G.; Sausville, E.A.; Teicher, B. Jasplakinolide: Interaction with radiation and hyperthermia in human prostate carcinoma and Lewis lung carcinoma. Cancer Chemother. Pharmacol. 1998, 42, 491–496. [Google Scholar] [CrossRef]
- Senderowicz, A.M.J.; Pham, N.B.; Fechner, G.; Zencak, D.; Vu, H.T.; Hooper, J.N.; Quinn, R.J. Jasplakinolide’s Inhibition of the Growth of Prostate Carcinoma Cells In Vitro with Disruption of the Actin Cytoskeleton. J. Natl. Cancer Inst. 1995, 87, 46–51. [Google Scholar] [CrossRef]
- Tran, T.D.; Pham, N.B.; Fechner, G.; Zencak, D.; Vu, H.T.; Hooper, J.N.; Quinn, R.J. Cytotoxic Cyclic Depsipeptides from the Australian Marine Sponge Neamphius huxleyi. J. Nat. Prod. 2012, 75, 2200–2208. [Google Scholar] [CrossRef]
- Suárez, Y.; González, L.; Cuadrado, A.; Berciano, M.; Lafarga, M.; Muñoz, A. Kahalalide F, a new marine-derived compound, induces oncosis in human prostate and breast cancer cells. Mol. Cancer Ther. 2003, 2, 863. [Google Scholar] [PubMed]
- Vervoort, H.; Fenical, W.; Epifanio, R.D.A. Tamandarins A and B: New Cytotoxic Depsipeptides from a Brazilian Ascidian of the Family Didemnidae. J. Org. Chem. 2000, 65, 782–792. [Google Scholar] [CrossRef] [PubMed]
- Liang, B.; Richard, D.J.; Portonovo, P.S.; Joullie, M.M. Total Syntheses and Biological Investigations of Tamandarins A and B and TamandarinA Analogs. J. Am. Chem. Soc. 2001, 123, 4469–4474. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Gu, W.; Lo, D.; Ding, X.-Z.; Ujiki, M.; Adrian, T.E.; Soff, G.A.; Silverman, R.B. N-Methylsansalvamide A Peptide Analogues. Potent New Antitumor Agents. J. Med. Chem. 2005, 48, 3630–3638. [Google Scholar] [CrossRef]
- Oh, D.-C.; Jensen, P.R.; Fenical, W. Zygosporamide, a cytotoxic cyclic depsipeptide from the marine-derived fungus Zygosporium masonii. Tetrahedron Lett. 2006, 47, 8625–8628. [Google Scholar] [CrossRef]
- Williams, D.E.; Yu, K.; Behrisch, H.W.; Van Soest, R.; Andersen, R.J. Rolloamides A and B, Cytotoxic Cyclic Heptapeptides Isolated from the Caribbean Marine Sponge Eurypon laughlini. J. Nat. Prod. 2009, 72, 1253–1257. [Google Scholar] [CrossRef]
- McKeever, B.; Pattenden, G. Total synthesis of trunkamide A, a novel thiazoline-based prenylated cyclopeptide metabolite from Lissoclinum sp. Tetrahedron 2003, 59, 2713–2727. [Google Scholar] [CrossRef]
- Rashid, M.A.; Gustafson, K.R.; Ii, J.H.C.; Boyd, M.R. Patellamide F, a New Cytotoxic Cyclic Peptide from the Colonial Ascidian Lissoclinum patella. J. Nat. Prod. 1995, 58, 594–597. [Google Scholar] [CrossRef]
- Bonnard, I.; Rolland, M.; Salmon, J.-M.; Debiton, E.; Barthomeuf, C.; Banaigs, B. Total Structure and Inhibition of Tumor Cell Proliferation of Laxaphycins. J. Med. Chem. 2007, 50, 1266–1279. [Google Scholar] [CrossRef]
- Rabiei, S.; Rezaei, M.; Asgharzade, S.; Nikoo, M.; Rafieia-Kopai, M. Antioxidant and cytotoxic properties of protein hydrolysates obtained from enzymatic hydrolysis of Klunzinger’s mullet (Liza klunzingeri) muscle. Braz. J. Pharm. Sci. 2019, 55, e18304. [Google Scholar] [CrossRef]
- Da Rocha, M.; Alemán, A.; Baccan, G.C.; López-Caballero, M.E.; Gómez-Guillén, C.; Montero, P.; Prentice, C. Anti-Inflammatory, Antioxidant, and Antimicrobial Effects of Underutilized Fish Protein Hydrolysate. J. Aquat. Food Prod. Technol. 2018, 27, 592–608. [Google Scholar] [CrossRef]
- Shaik, M.I.; Sarbon, N.M. A review on purification and characterization of anti-proliferative peptides derived from fish protein hydrolysate. Food Rev. Int. 2020, 38, 1389–1409. [Google Scholar] [CrossRef]
- Eun-Kyung, K.; Joung, H.J.; Kim, Y.S.; Hwang, J.W.; Ahn, C.B.; Jeon, Y.J.; Moon, S.H.; Park, P.J. Purification of a Novel Anticancer Peptide from Enzymatic Hydrolysate of Mytilus coruscus. J. Microbiol. Biotechnol. 2012, 22, 1381–1387. [Google Scholar]
- Ding, G.-F.; Huang, F.F.; Yang, Z.-S.; Yu, D.; Yang, Y.F. Anticancer Activity of an Oligopeptide Isolated from Hydrolysates of Sepia Ink. Chin. J. Nat. Med. 2011, 9, 151–155. [Google Scholar]
- Jing, Y.; Yang, Z.; Huang, F. Mechanism of Sepia ink polypeptide-induced apoptosis in DU-145 prostate cancer cells. Mod. Food Sci. Technol. 2014, 30, 1–6. [Google Scholar]
- Khalifa, S.A.M.; Elias, N.; Farag, M.A.; Chen, L.; Saeed, A.; Hegazy, M.-E.F.; Moustafa, M.S.; El-Wahed, A.A.; Al-Mousawi, S.M.; Musharraf, S.G.; et al. Marine natural products: A source of novel anticancer drugs. Mar. Drugs 2019, 17, 491. [Google Scholar] [CrossRef]
- Pfeffer, C.M.; Singh, A.T. Apoptosis: A target for anticancer therapy. Int. J. Mol. Sci. 2018, 19, 448. [Google Scholar] [CrossRef]
- Gantar, M.; Dhandayuthapani, S.; Rathinavelu, A. Phycocyanin Induces Apoptosis and Enhances the Effect of Topotecan on Prostate Cell Line LNCaP. J. Med. Food 2012, 15, 1091–1095. [Google Scholar] [CrossRef]
- Parrish, A.B.; Freel, C.D.; Kornbluth, S. Cellular mechanisms controlling caspase activation and function. Cold Spring Harb. Perspect. Biol. 2013, 5, a008672. [Google Scholar] [CrossRef]
- Chen, Y.; Xu, X.; Hong, S.; Chen, J.; Liu, N.; Underhill, C.B.; Creswell, K.; Zhang, L. RGD-Tachyplesin Inhibits Tumor Growth. Cancer Res. 2001, 61, 2434. [Google Scholar]
- Sun, J.-Y.; Wang, J.-D.; Wang, X.; Liu, H.-C.; Zhang, M.-M.; Liu, Y.-C.; Zhang, C.-H.; Su, Y.; Shen, Y.-Y.; Guo, Y.-W.; et al. Marine-derived chromopeptide A, a novel class I HDAC inhibitor, suppresses human prostate cancer cell proliferation and migration. Acta Pharmacol. Sin. 2017, 38, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Campbell, K.J.; Tait, S.W. Targeting BCL-2 regulated apoptosis in cancer. Open Biol. 2018, 8, 180002. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.-J.; Dasgupta, A.; Jung, K.-J.; Um, J.-H.; Burke, A.; Park, H.U.; Brady, J.N. PI3K/AKT inhibition induces caspase-dependent apoptosis in HTLV-1-transformed cells. Virology 2008, 370, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Chin, H.; Kim, K.; Lee, D. ERBB3 knockdown induces cell cycle arrest and activation of Bak and Bax-dependent apoptosis in colon cancer cells. Oncotarget 2014, 5, 5138–5152. [Google Scholar] [CrossRef] [PubMed]
- Janmaat, M.L.; Rodriguez, J.A.; Jimeno, J.; Kruyt, F.A.E.; Giaccone, G. Kahalalide F Induces Necrosis-Like Cell Death that Involves Depletion of ErbB3 and Inhibition of Akt Signaling. Mol. Pharmacol. 2005, 68, 502. [Google Scholar] [CrossRef]
- Serova, M.; de Gramont, A.; Bieche, I.; Riveiro, M.E.; Galmarini, C.M.; Aracil, M.; Jimeno, J.; Faivre, S.; Raymond, E. Predictive factors of sensitivity to elisidepsin, a novel Kahalalide F-derived marine compound. Mar. Drugs 2013, 11, 944–959. [Google Scholar] [CrossRef]
- Wang, E.; Sorolla, M.A.; Krishnan, P.D.G.; Sorolla, A. From Seabed to Bedside: A Review on Promising Marine Anticancer Compounds. Biomolecules 2020, 10, 248. [Google Scholar] [CrossRef]
- Molina-Guijarro, J.M.; Macías, Á.; García, C.; Muñoz, E.; García-Fernandez, L.F.; David, M.; Núñez, L.; Martínez-Leal, J.F.; Moneo, V.; Cuevas, C.; et al. Irvalec inserts into the plasma membrane causing rapid loss of integrity and necrotic cell death in tumor cells. PLoS ONE 2011, 6, e19042. [Google Scholar] [CrossRef]
- Yue, J.; López, J.M. Understanding MAPK Signaling Pathways in Apoptosis. Int. J. Mol. Sci. 2020, 21, 2346. [Google Scholar] [CrossRef]
- Canovas, B.; Nebreda, A.R. Diversity and versatility of p38 kinase signalling in health and disease. Nat. Rev. Mol. Cell Biol. 2021, 22, 346–366. [Google Scholar] [CrossRef]
- van Vuuren, R.J.; Visagie, M.H.; Theron, A.E.; Joubert, A.M. Antimitotic drugs in the treatment of cancer. Cancer Chemother. Pharmacol. 2015, 76, 1101–1112. [Google Scholar] [CrossRef] [PubMed]
- Fanale, D.; Bronte, G.; Passiglia, F.; Calò, V.; Castiglia, C.; Di Piazza, F.; Barraco, N.; Cangemi, A.; Catarella, M.T.; Insalaco, L.; et al. Stabilizing versus destabilizing the microtubules: A double-edge sword for an effective cancer treatment option? Anal. Cell. Pathol. 2015, 2015, 690916. [Google Scholar] [CrossRef] [PubMed]
- Alves, C.; Silva, J.; Pinteus, S.; Gaspar, H.; Alpoim, M.C.; Botana, L.M.; Pedrosa, R. From Marine Origin to Therapeutics: The Antitumor Potential of Marine Algae-Derived Compounds. Front. Pharmacol. 2018, 9, 777. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Monserrate, Z.; Vervoort, H.C.; Bai, R.; Newman, D.J.; Howell, S.B.; Los, G.; Mullaney, J.T.; Williams, M.D.; Pettit, G.R.; Fenical, W.; et al. Diazonamide A and a Synthetic Structural Analog: Disruptive Effects on Mitosis and Cellular Microtubules and Analysis of Their Interactions with Tubulin. Mol. Pharmacol. 2003, 63, 1273. [Google Scholar] [CrossRef]
- Welch, D.R.; Hurst, D.R. Defining the Hallmarks of Metastasis. Cancer Res. 2019, 79, 3011. [Google Scholar] [CrossRef]
- Lambert, A.W.; Pattabiraman, D.R.; Weinberg, R.A. Emerging Biological Principles of Metastasis. Cell 2017, 168, 670–691. [Google Scholar] [CrossRef]
- Hockla, A.; Miller, E.; Salameh, M.D.A.; Copland, J.A.; Radisky, D.C.; Radisky, E.S. PRSS3/mesotrypsin is a therapeutic target for metastatic prostate cancer. Mol. Cancer Res. 2012, 10, 1555–1566. [Google Scholar] [CrossRef]
- Wei, X.; Zeng, W.; Xie, K.; Diao, P.; Tang, P. Potential use of chymotrypsin-like proteasomal activity as a biomarker for prostate cancer. Oncol. Lett. 2018, 15, 5149–5154. [Google Scholar] [CrossRef]
- Lerman, I. Neutrophil Elastase and SERPINB1 Are Critical Regulators of Prostate Cancer Progression; University of Rochester: Ann Arbor, MI, USA, 2018; p. 159. [Google Scholar]
- Linington, R.G.; Edwards, D.J.; Shuman, C.F.; McPhail, K.L.; Matainaho, T.; Gerwick, W.H. Symplocamide A, a potent cytotoxin and chymotrypsin inhibitor from the marine Cyanobacterium Symploca sp. J. Nat. Prod. 2008, 71, 22–27. [Google Scholar] [CrossRef]
- Taori, K.; Paul, V.J.; Luesch, H. Kempopeptins A and B, Serine Protease Inhibitors with Different Selectivity Profiles from a Marine Cyanobacterium, Lyngbya sp. J. Nat. Prod. 2008, 71, 1625–1629. [Google Scholar] [CrossRef]
- Rubio, B.K.; Parrish, S.M.; Yoshida, W.; Schupp, P.J.; Schils, T.; Williams, P.G. Depsipeptides from a Guamanian marine cyanobacterium, Lyngbyabouillonii, with selective inhibition of serine proteases. Tetrahedron Lett. 2010, 51, 6718–6721. [Google Scholar] [CrossRef] [PubMed]
- Gunasekera, S.P.; Miller, M.W.; Kwan, J.C.; Luesch, H.; Paul, V.J. Molassamide, a Depsipeptide Serine Protease Inhibitor from the Marine Cyanobacterium Dichothrixutahensis. J. Nat. Prod. 2010, 73, 459–462. [Google Scholar] [CrossRef] [PubMed]
- Matthew, S.; Paul, V.J.; Luesch, H. Largamides A-C, tiglic acid-containing cyclodepsipeptides with elastase-inhibitory activity from the marine cyanobacterium Lyngbya confervoides. Planta Med. 2009, 75, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Plaza, A.; Bewley, C.A. Largamides A−H, Unusual Cyclic Peptides from the Marine Cyanobacterium Oscillatoria sp. J. Org. Chem. 2006, 71, 6898–6907. [Google Scholar] [CrossRef]
- Matthew, S.; Ross, C.; Rocca, J.R.; Paul, V.J.; Luesch, H. Pompanopeptins A and B, new cyclic peptides from the marine cyanobacterium Lyngbyaconfervoides. Tetrahedron 2008, 64, 4081–4089. [Google Scholar] [CrossRef]
- Matthew, S.; Ross, C.; Rocca, J.R.; Paul, V.J.; Luesch, H. Lyngbyastatin 4, a Dolastatin 13 Analogue with Elastase and Chymotrypsin Inhibitory Activity from the Marine Cyanobacterium Lyngbyaconfervoides. J. Nat. Prod. 2007, 70, 124–127. [Google Scholar] [CrossRef]
- Taori, K.; Matthew, S.; Rocca, J.R.; Paul, V.J.; Luesch, H. Lyngbyastatins 5–7, Potent Elastase Inhibitors from Floridian Marine Cyanobacteria, Lyngbya sp. J. Nat. Prod. 2007, 70, 1593–1600. [Google Scholar] [CrossRef]
- Kwan, J.C.; Taori, K.; Paul, V.J.; Luesch, H. Lyngbyastatins 8-10, elastase inhibitors with cyclic depsipeptide scaffolds isolated from the marine cyanobacterium Lyngbyasemiplena. Mar. Drugs 2009, 7, 528–538. [Google Scholar] [CrossRef]
- Matthew, S.; Paul, V.J.; Luesch, H. Tiglicamides A–C, cyclodepsipeptides from the marine cyanobacterium Lyngbyaconfervoides. Phytochemistry 2009, 70, 2058–2063. [Google Scholar] [CrossRef]
- Luesch, H.; Pangilinan, R.; Yoshida, W.Y.; Moore, R.E.; Paul, V.J. Pitipeptolides A and B, New Cyclodepsipeptides from the Marine Cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2001, 64, 304–307. [Google Scholar] [CrossRef]
- Pruitt, F.L.; He, Y.; Franco, O.E.; Jiang, M.; Cates, J.; Hayward, S.W. Cathepsin D acts as an essential mediator to promote malignancy of benign prostatic epithelium. Prostate 2013, 73, 476–488. [Google Scholar] [CrossRef]
- Eatemadi, A.; Aiyelabegan, H.T.; Negahdari, B.; Mazlomi, M.A.; Daraee, H.; Daraee, N.; Eatemadi, R.; Sadroddiny, E. Role of protease and protease inhibitors in cancer pathogenesis and treatment. Biomed. Pharmacother. 2017, 86, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Kwan, J.C.; Eksioglu, E.A.; Liu, C.; Paul, V.J.; Luesch, H. Grassystatins A−C from Marine Cyanobacteria, Potent Cathepsin E Inhibitors That Reduce Antigen Presentation. J. Med. Chem. 2009, 52, 5732–5747. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Qin, Y.; Kun, L.; Zhou, Y. The Significant Role of the Microfilament System in Tumors. Front. Oncol. 2021, 11, 620390. [Google Scholar] [CrossRef]
- Yildirim, S.; Altun, S.; Gumushan, H.; Patel, A.; Djamgoz, M.B. Voltage-gated sodium channel activity promotes prostate cancer metastasis in vivo. Cancer Lett. 2012, 323, 58–61. [Google Scholar] [CrossRef] [PubMed]
- Angus, M.; Ruben, P. Voltage gated sodium channels in cancer and their potential mechanisms of action. Channels 2019, 13, 400–409. [Google Scholar] [CrossRef] [PubMed]
- Mao, W.; Zhang, J.; Körner, H.; Jiang, Y.; Ying, S. The Emerging Role of Voltage-Gated Sodium Channels in Tumor Biology. Front. Oncol. 2019, 9, 124. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, E.O.; Graf, K.M.; Patel, M.K.; Baheti, A.; Kong, H.-S.; MacArthur, L.H.; Dakshanamurthy, S.; Wang, K.; Brown, M.L.; Paige, M. Synthesis and evaluation of hermitamides A and B as human voltage-gated sodium channel blockers. Bioorganic Med. Chem. 2011, 19, 4322–4329. [Google Scholar] [CrossRef]
- Taniguchi, M.; Nunnery, J.K.; Engene, N.; Esquenazi, E.; Byrum, T.; Dorrestein, P.C.; Gerwick, W.H. Palmyramide A, a Cyclic Depsipeptide from a Palmyra Atoll Collection of the Marine Cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2010, 73, 393–398. [Google Scholar] [CrossRef]
- Shibuya, M. Vascular Endothelial Growth Factor (VEGF) and Its Receptor (VEGFR) Signaling in Angiogenesis: A Crucial Target for Anti- and Pro-Angiogenic Therapies. Genes Cancer 2011, 2, 1097–1105. [Google Scholar] [CrossRef]
- Ceci, C.; Atzori, M.G.; Lacal, P.M.; Graziani, G. Role of VEGFs/VEGFR-1 Signaling and its Inhibition in Modulating Tumor Invasion: Experimental Evidence in Different Metastatic Cancer Models. Int. J. Mol. Sci. 2020, 21, 1388. [Google Scholar] [CrossRef] [PubMed]
- Khodabakhsh, F.; Merikhian, P.; Eisavand, M.R.; Farahmand, L. Crosstalk between MUC1 and VEGF in angiogenesis and metastasis: A review highlighting roles of the MUC1 with an emphasis on metastatic and angiogenic signaling. Cancer Cell Int. 2021, 21, 200. [Google Scholar] [CrossRef]
- Guha, P.; Kaptan, E.; Bandyopadhyaya, G.; Kaczanowska, S.; Davila, E.; Thompson, K.; Martin, S.S.; Kalvakolanu, D.V.; Vasta, G.R.; Ahmed, H. Cod glycopeptide with picomolar affinity to galectin-3 suppresses T-cell apoptosis and prostate cancer metastasis. Proc. Natl. Acad. Sci. 2013, 110, 5052–5057. [Google Scholar] [CrossRef] [PubMed]
- Visconti, R.; Della Monica, R.; Grieco, D. Cell cycle checkpoint in cancer: A therapeutically targetable double-edged sword. J. Exp. Clin. Cancer Res. CR 2016, 35, 153. [Google Scholar] [CrossRef]
- Liu, K.; Zheng, M.; Lu, R.; Du, J.; Zhao, Q.; Li, Z.; Li, Y.; Zhang, S. The role of CDC25C in cell cycle regulation and clinical cancer therapy: A systematic review. Cancer Cell Int. 2020, 20, 213. [Google Scholar] [CrossRef] [PubMed]
- Ozen, M.; Ittmann, M. Increased Expression and Activity of CDC25C Phosphatase and an Alternatively Spliced Variant in Prostate Cancer. Clin. Cancer Res. 2005, 11, 4701. [Google Scholar] [CrossRef]
- Wan, J.; Zhang, J.; Zhang, J. Expression of p53 and its mechanism in prostate cancer. Oncol. Lett. 2018, 16, 378–382. [Google Scholar] [CrossRef]
- Pietsch, E.C.; Sykes, S.M.; McMahon, S.B.; Murphy, M.E. The p53 family and programmed cell death. Oncogene 2008, 27, 6507–6521. [Google Scholar] [CrossRef]
- Chen, J. The Cell-Cycle Arrest and Apoptotic Functions of p53 in Tumor Initiation and Progression. Cold Spring Harb. Perspect. Med. 2016, 6, a026104. [Google Scholar] [CrossRef]
- Li, G.; Tian, Y.; Zhu, W.-G. The Roles of Histone Deacetylases and Their Inhibitors in Cancer Therapy. Front. Cell Dev. Biol. 2020, 8, 576946. [Google Scholar] [CrossRef]
- Aggarwal, V.; Tuli, H.S.; Varol, A.; Thakral, F.; Yerer, M.B.; Sak, K.; Varol, M.; Jain, A.; Khan, M.A.; Sethi, G. Role of Reactive Oxygen Species in Cancer Progression: Molecular Mechanisms and Recent Advancements. Biomolecules 2019, 9, 735. [Google Scholar] [CrossRef] [PubMed]
- Poetsch, A.R. The genomics of oxidative DNA damage, repair, and resulting mutagenesis. Comput. Struct. Biotechnol. J. 2020, 18, 207–219. [Google Scholar] [CrossRef]
- Ueoka, R.; Ise, Y.; Ohtsuka, S.; Okada, S.; Yamori, T.; Matsunaga, S. Yaku’amides A and B, Cytotoxic Linear Peptides Rich in Dehydroamino Acids from the Marine Sponge Ceratopsions. J. Am. Chem. Soc. 2010, 132, 17692–17694. [Google Scholar] [CrossRef] [PubMed]
- Song, R.; Wei, R.B.; Luo, H.Y.; Yang, Z.S. Isolation and identification of an antiproliferative peptide derived from heated products of peptic hydrolysates of half-fin anchovy (Setipinnataty). J. Funct. Foods 2014, 10, 104–111. [Google Scholar] [CrossRef]
- Teruya, T.; Sasaki, H.; Fukazawa, H.; Suenaga, K. Bisebromoamide, a Potent Cytotoxic Peptide from the Marine Cyanobacterium Lyngbya sp.: Isolation, Stereostructure, and Biological Activity. Org. Lett. 2009, 11, 5062–5065. [Google Scholar] [CrossRef] [PubMed]
- Loganzo, F.; Discafani, C.M.; Annable, T.; Beyer, C.; Musto, S.; Hari, M.; Tan, X.; Hardy, C.; Hernandez, R.; Baxter, M.; et al. HTI-286, a synthetic analogue of the tripeptide hemiasterlin, is a potent antimicrotubule agent that circumvents P-glycoprotein-mediated resistance in vitro and in vivo. Cancer Res. 2003, 63, 1838–1845. [Google Scholar]
- Faircloth, G.; Marchante, M.d.C.C. Kahalalide F and ES285: Potent anticancer agents from marine molluscs. In Molluscs; Springer: Berlin/Heidelberg, Germany, 2006; pp. 363–379. [Google Scholar]
- Riggs, D.R.; Jackson, B.; Vona-Davis, L.; McFadden, D. In vitro anticancer effects of a novel immunostimulant: Keyhole limpet hemocyanin. J. Surg. Res. 2002, 108, 279–284. [Google Scholar] [CrossRef]
- Vaishampayan, U.; Glode, M.; Du, W.; Kraft, A.; Hudes, G.; Wright, J.; Hussain, M. Phase II study of dolastatin-10 in patients with hormone-refractory metastatic prostate adenocarcinoma. Clin. Cancer Res. 2000, 6, 4205–4208. [Google Scholar]
- Kobayashi, M.; Natsume, T.; Watanabe, J.-I.; Fujio, N.; Mikami, T.; Miyasaka, K.; Tsukagoshi, S. Activity of a novel antitumor agent, TZT-1027. Nihon yakurigakuzasshi. Folia Pharmacol. Jpn. 1999, 114, 230–235. [Google Scholar] [CrossRef][Green Version]
- Greystoke, A.; Blagden, S.; Thomas, A.L.; Scott, E.; Attard, G.; Molife, R.; Vidal, L.; Pacey, S.C.; Sarkar, D.; Jenner, A.; et al. A phase I study of intravenous TZT-1027 administered on day 1 and day 8 of a three-weekly cycle in combination with carboplatin given on day 1 alone in patients with advanced solid tumours. Ann. Oncol. 2006, 17, 1313–1319. [Google Scholar] [CrossRef]
- Gogineni, V.; Hamann, M.T. Marine natural product peptides with therapeutic potential: Chemistry, biosynthesis, and pharmacology. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 81–196. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Currano, J.N.; Carroll, P.J.; Joullié, M.M. Didemnins, tamandarins and related natural products. Nat. Prod. Rep. 2012, 29, 404–424. [Google Scholar] [CrossRef] [PubMed]
- Rademaker-Lakhai, J.M.; Horenblas, S.; Meinhardt, W.; Stokvis, E.; de Reijke, T.M.; Jimeno, J.M.; Lopez-Lazaro, L.; Martin, J.A.L.; Beijnen, J.H.; Schellens, J.H. Phase I clinical and pharmacokinetic study of kahalalide F in patients with advanced androgen refractory prostate cancer. Clin. Cancer Res. 2005, 11, 1854–1862. [Google Scholar] [CrossRef] [PubMed]
- Ghareeb, M.A.; Tammam, M.A.; El-Demerdash, A.; Atanasov, A.G. Insights about clinically approved and Preclinically investigated marine natural products. Curr. Res. Biotechnol. 2020, 2, 88–102. [Google Scholar] [CrossRef]
- Ruiz-Torres, V.; Encinar, J.A.; Herranz-López, M.; Pérez-Sánchez, A.; Galiano, V.; Barrajón-Catalán, E.; Micol, V. An Updated Review on Marine Anticancer Compounds: The Use of Virtual Screening for the Discovery of Small-Molecule Cancer Drugs. Molecules 2017, 22, 1037. [Google Scholar] [CrossRef]
- Kalimuthu, S.; Venkatesan, J.; Kim, S.-K. Marine derived bioactive compounds for breast and prostate cancer treatment: A review. Curr. Bioact. Compd. 2014, 10, 62–74. [Google Scholar] [CrossRef]
- Craik, D.J.; Kan, M.-W. How can we improve peptide drug discovery? Learning from the past. Expert Opin. Drug Discov. 2021, 16, 1399–1402. [Google Scholar] [CrossRef]
- Muttenthaler, M.; King, G.F.; Adams, D.J.; Alewood, P.F. Trends in peptide drug discovery. Nat. Rev. Drug Discov. 2021, 20, 309–325. [Google Scholar] [CrossRef]
- Lee, A.C.-L.; Harris, J.L.; Khanna, K.K.; Hong, J.-H. A Comprehensive Review on Current Advances in Peptide Drug Development and Design. Int. J. Mol. Sci. 2019, 20, 2383. [Google Scholar] [CrossRef]




| In Vitro | In Vivo | References | ||
|---|---|---|---|---|
| Human Prostate Cancer Cell Lines | IC50 | Experimental Model | Dose | |
| DU145 and PC-3 Aurilide B | <10 nM | ----- | ----- | [22] |
| PC-3 Lagunamide C | 2.6 nM | ----- | ----- | [23] |
| DU145 and LNCaP Cryptophycin-52 | 1–10 pM | ----- | ----- | [24] |
| DU145 and PC-3 Coibamide A | 300 ng mL−1 | ----- | ----- | [10,25,32] |
| PC-3 Laxaphycin B | 0.58 µM | ----- | ----- | [39] |
| LNCaP C-phycocyanin | 500 µg mL−1 | ----- | ----- | [48] |
| DU145 and PC-3 Bisebromoamide | GI50:40 nM | ----- | ----- | [106] |
| DU145 Jaspamide | 0.8 µM | DU-145 xenograft | 10 mg/kg s.c. | [28] |
| LNCaP Jaspamide | 0.07 µM | ----- | ----- | |
| PC-3 Jaspamide | 0.3 µM | ----- | ----- | |
| TSU-Pr1 Jaspamide | 170 nM | ----- | ----- | [29] |
| PC-3 Homophymines A–E | A:4.2, B:6.2, C:3.0, D:6.3, E:3.9 nM | ----- | ----- | [27] |
| LNCaP Neamphamides B-D | B:230, C:190, D:110 nM | ----- | ----- | [30] |
| PC-3 Geodiamolides D-F | B: 170, C:110, D:130 nM | ----- | ----- | |
| D:33.1, E:118, F:155 nM | ----- | ----- | [26] | |
| DU145 Rolloamide A | 0.85 µM | ----- | ----- | [36] |
| LNCaP Rolloamide A | 0.8 µM | ----- | ----- | |
| PC-3 Rolloamide A | 1.4 µM | ----- | ----- | |
| PC3MM2 Rolloamide A | 4.7 µM | ----- | ----- | |
| LNCaP, C4-2, PC-3, PC-3dR HTI-286 | 0.65–4.6 nM | PC-3 and PC-3dR xenografts | 1.5 mg/kg i.v. | [14] |
| ----- | ----- | PC3-MM2 xenograft | 1.0 mg/kg i.v. | [107] |
| PC-3 Kahalalide F | 0.07 µM | ----- | ----- | [31] |
| DU145 and LNCaP | 0.28 µM | ----- | ----- | |
| ----- | ----- | PC-3 and DU145 xenografts | 123 μg/kg i.v. | [108] |
| PC-3 Elisidepsin | 1.80 µM | ----- | ----- | [56] |
| DU145 | 1.26 µM | ----- | ----- | |
| DU145 Dolastatin 10 | 0.5 nM | DU145 xenograft | 5 µg q4d i.p. | [19] |
| PC-3 MCH | LC50:0.94 mg mL−1 | ----- | ----- | [43] |
| DU145 Tamandarin A | GI50:12.5 μg | ----- | ----- | [109] |
| 1.36 ng mL−1 | ----- | ----- | [32] | |
| PC-3 Tamandarin B | 1.4 µM | ----- | ----- | [33] |
| DU145 and PC-3 Patellamide B Patellamide F | LC50: 48 µM | ----- | ----- | [38] |
| LC50: 13 µM | ----- | ----- | ||
| LC50: 3 µM | ----- | ----- | ||
| DU145 Trunkamide A | 7.08 nM | ----- | ----- | [12,37] |
| PC-3 Diazonamide A | 2.3 nM | ----- | ----- | [64] |
| 2.43 nmol L−1 | PC-3 xenograft | 1.6 mg/kg i.v. | [51] | |
| DU145 Chromopeptide A | 2.08 nmol L−1 | ----- | ----- | |
| LNCaP Chromopeptide A | 1.75 nmol L−1 | ----- | ----- | |
| PC-3 Sansalvamide A | 27.4 μg mL−1 | ----- | ----- | [34] |
| DU145 and PC-3 Microsporin A Zygosporamide | 2.7 µM | ----- | ----- | [17] |
| GI50:9.1 µM | ----- | ----- | [35] | |
| PC-3 SHP | 15 mg mL−1 | ----- | ----- | [15] |
| DU145 | 1 mg mL−1 | ----- | ----- | [44,45] |
| DU145 and PC-3 SIO | 15 mg mL−1 | ----- | ----- | [16] |
| PC-3 TFD, YALRAH, YALPAH, YALPAG, YALPAR | 3.5 nM | ----- | ----- | [94] |
| GI50:16.9 μM | ----- | ----- | [105] | |
| GI50:11.1 μM | ----- | ----- | ||
| GI50:19.0 μM | ----- | ----- | ||
| GI50:71.2 μM | ----- | ----- | ||
| DU145, ILYMP, SCH-P9 and SCH-P10 | 11.25 mM | ----- | ----- | [20] |
| SCH-P9:1.21, SCH-P10: 1.41 mg mL−1 | ----- | ----- | [18] | |
| PC-3 | SCH-P9:1.09, SCH-P10: 0.91 mg mL−1 | ----- | ----- | |
| DU145, AAP-H | 2.298 mM | ----- | ----- | [21] |
| TSU, Tachyplesin | 75 μg mL−1 | ----- | ----- | [50] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, S.; Alam, W.; Jeandet, P.; Aschner, M.; Alsharif, K.F.; Saso, L.; Khan, H. Therapeutic Potential of Marine Peptides in Prostate Cancer: Mechanistic Insights. Mar. Drugs 2022, 20, 466. https://doi.org/10.3390/md20080466
Ahmed S, Alam W, Jeandet P, Aschner M, Alsharif KF, Saso L, Khan H. Therapeutic Potential of Marine Peptides in Prostate Cancer: Mechanistic Insights. Marine Drugs. 2022; 20(8):466. https://doi.org/10.3390/md20080466
Chicago/Turabian StyleAhmed, Salman, Waqas Alam, Philippe Jeandet, Michael Aschner, Khalaf F. Alsharif, Luciano Saso, and Haroon Khan. 2022. "Therapeutic Potential of Marine Peptides in Prostate Cancer: Mechanistic Insights" Marine Drugs 20, no. 8: 466. https://doi.org/10.3390/md20080466
APA StyleAhmed, S., Alam, W., Jeandet, P., Aschner, M., Alsharif, K. F., Saso, L., & Khan, H. (2022). Therapeutic Potential of Marine Peptides in Prostate Cancer: Mechanistic Insights. Marine Drugs, 20(8), 466. https://doi.org/10.3390/md20080466