
Analysis of Structural Determinants of Peptide MS 9a-1 Essential for Potentiating of TRPA1 Channel

 , , , and
, , , and 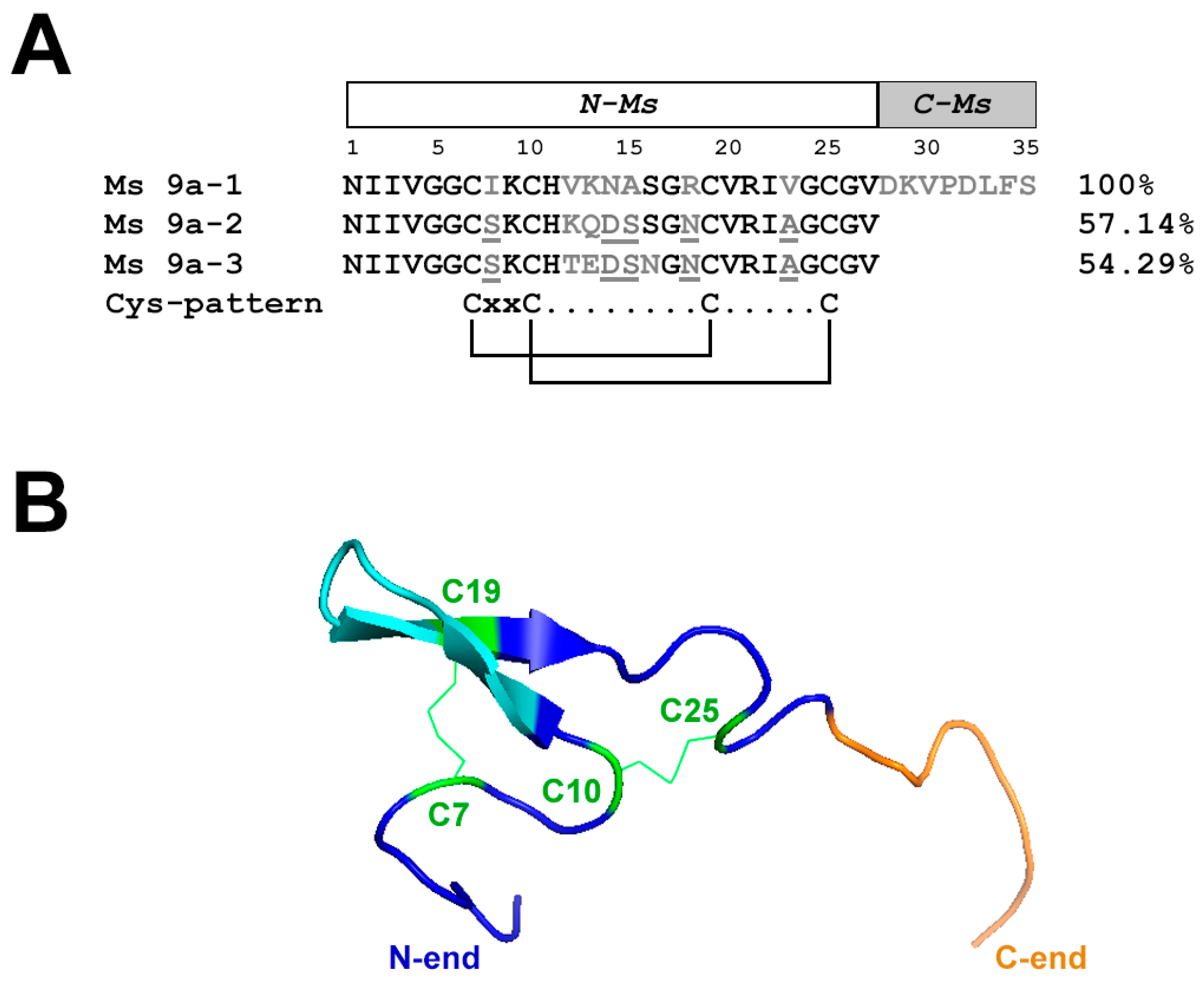{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Alignment of Peptides and Ms 9a-1 Spatial Structure Modeling
2.2. Peptide Production
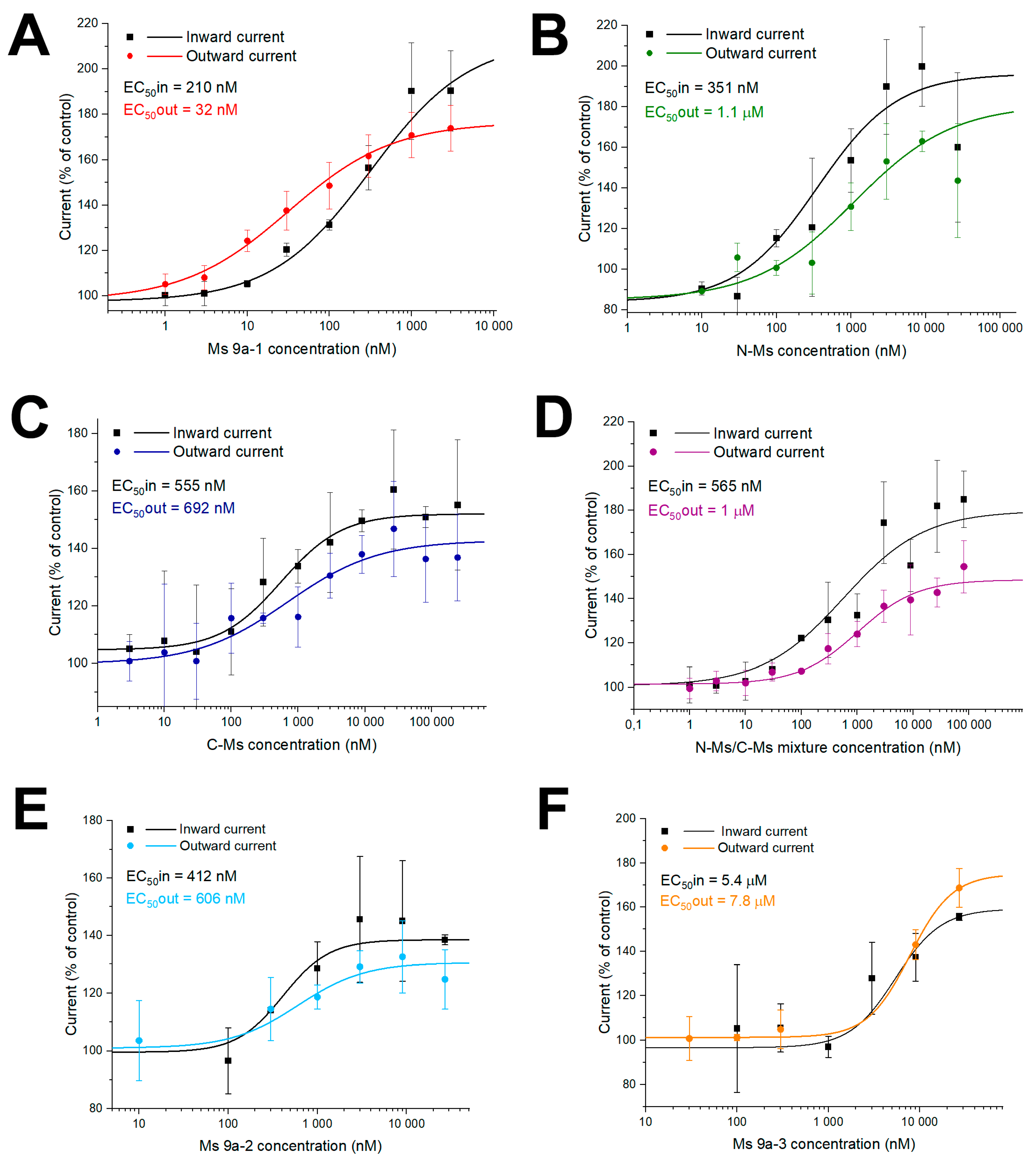
2.3. Electrophysiology
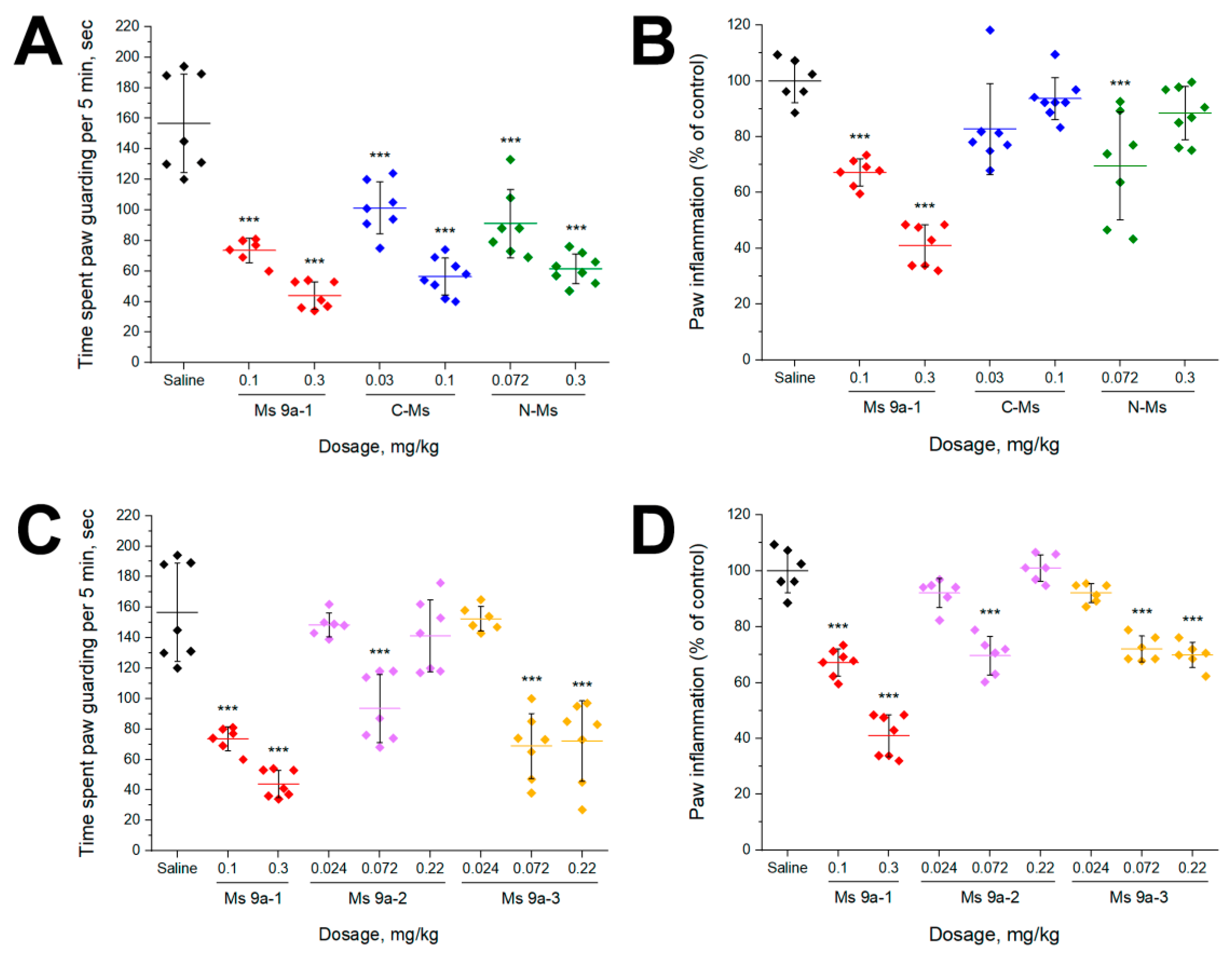
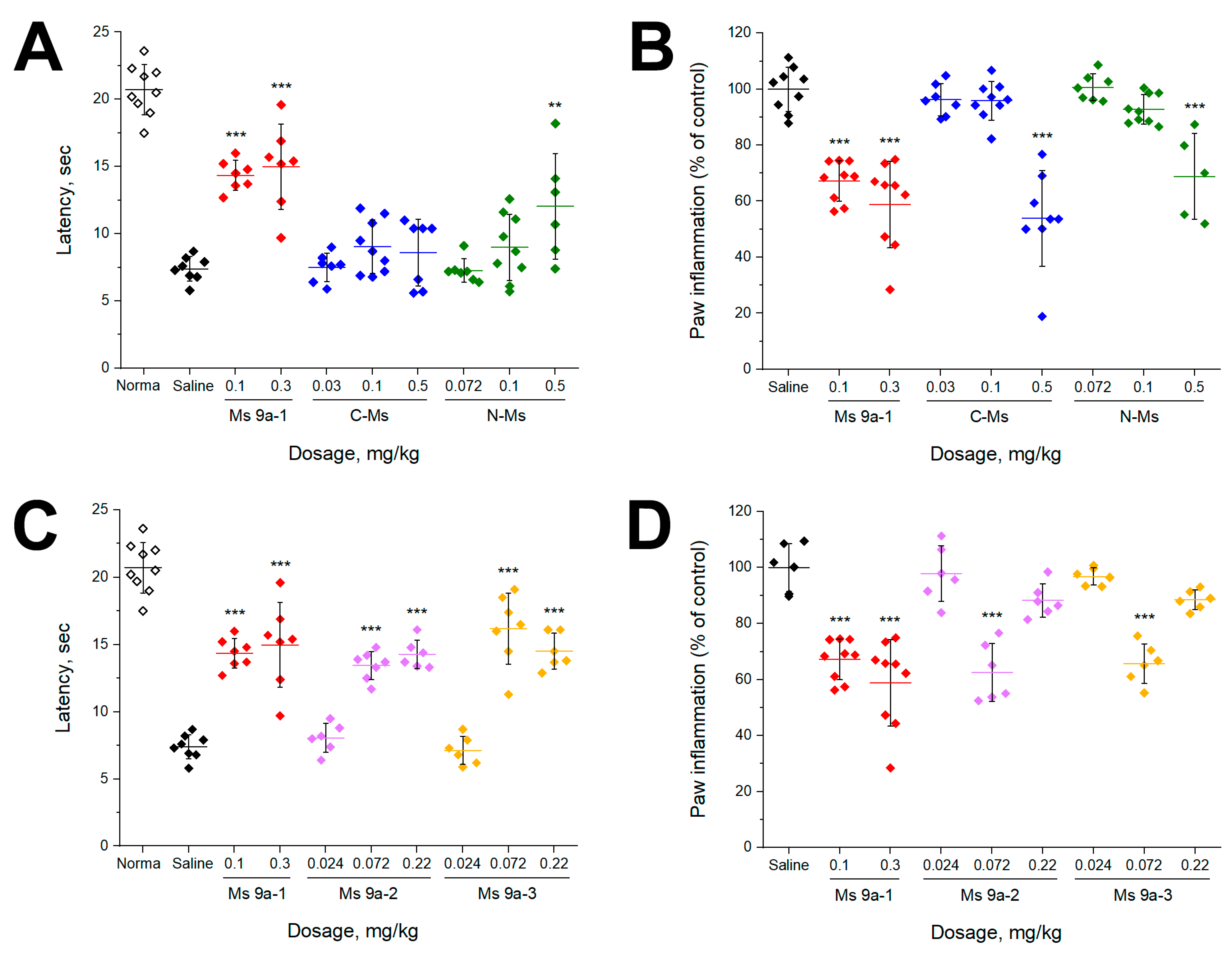
2.4. Experiments In Vivo
3. Discussion
4. Materials and Methods
4.1. Synthesis of C-Ms
4.2. Recombinant Peptide Production
4.3. Electrophysiological Studies on Xenopus Laevis Oocytes
4.4. Computation and Structure Modeling
4.5. In Vivo Experiments
4.5.1. Animal Models
4.5.2. Allyl Isothiocyanate (AITC)-Induced Nocifensive Beaviour
4.5.3. Complete Freund’s Adjuvant (CFA)-Induced Inflammation and Thermal Hyperalgesia
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AITC | allyl isothiocyanate |
| CFA | Complete Freund’s adjuvant |
| C-Ms | short C-terminal domain of Ms 9a-1 |
| Ms 9a-1 (full name—τ-AnmTX Ms 9a-1) | Metridium senile peptide of structural class 9a of sea anemone peptides, member 1 |
| Ms 9a-2 (full name—τ-AnmTX Ms 9a-2) | Metridium senile peptide of structural class 9a of sea anemone peptides, member 2 |
| Ms 9a-3 (full name—τ-AnmTX Ms 9a-3) | Metridium senile peptide of structural class 9a of sea anemone peptides, member 3 |
| N-Ms | Ms 9a-1 without C-terminal domain |
| nAChR | Neuronal nicotinic acetylcholine receptor |
| TRPV6 | Transient Receptor Potential channel, subfamily V (vanilloid), member 6 |
| TRPM8 | Transient Receptor Potential channel, subfamily M (melastatin), member 8 |
| TRPA1 | transient receptor potential cation channel, subfamily A (ankyrin), member 1 |
| Ugr 9a-1 (full name—π-AnmTX Ugr 9a-1) | Urticina grebelnyi peptide of structural class 9a of sea anemone peptides, member 1 |
References
- Pennington, M.W.; Czerwinski, A.; Norton, R.S. Peptide Therapeutics from Venom: Current Status and Potential. Bioorg. Med. Chem. 2018, 26, 2738–2758. [Google Scholar] [CrossRef] [PubMed]
- Utkin, Y.N. Animal Venom Studies: Current Benefits and Future Developments. World J. Biol. Chem. 2015, 6, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y. Why Do We Study Animal Toxins? Zool. Res. 2015, 36, 183–222. [Google Scholar] [CrossRef]
- Logashina, Y.A.; Mosharova, I.V.; Korolkova, Y.V.; Shelukhina, I.V.; Dyachenko, I.A.; Palikov, V.A.; Palikova, Y.A.; Murashev, A.N.; Kozlov, S.A.; Stensvåg, K.; et al. Peptide from Sea Anemone Metridium Senile Affects Transient Receptor Potential Ankyrin-Repeat 1 (TRPA1) Function and Produces Analgesic Effect. J. Biol. Chem. 2017, 292, 2992–3004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logashina, Y.A.; Korolkova, Y.V.; Kozlov, S.A.; Andreev, Y.A. TRPA1 Channel as a Regulator of Neurogenic Inflammation and Pain: Structure, Function, Role in Pathophysiology, and Therapeutic Potential of Ligands. Biochemistry 2019, 84, 101–118. [Google Scholar] [CrossRef]
- Samanta, A.; Hughes, T.E.T.; Moiseenkova-Bell, V.Y. Transient Receptor Potential (TRP) Channels. In Membrane Protein Complexes: Structure and Function; Part of Subcellular Biochemistry Book Series; Springer: Singapore, 2018; Volume 87, pp. 141–165. [Google Scholar] [CrossRef]
- Kremeyer, B.; Lopera, F.; Cox, J.J.; Momin, A.; Rugiero, F.; Marsh, S.; Woods, C.G.; Jones, N.G.; Paterson, K.J.; Fricker, F.R.; et al. A Gain-of-Function Mutation in TRPA1 Causes Familial Episodic Pain Syndrome. Neuron 2010, 66, 671–680. [Google Scholar] [CrossRef] [Green Version]
- Kozlov, S.; Grishin, E. Convenient Nomenclature of Cysteine-Rich Polypeptide Toxins from Sea Anemones. Peptides 2012, 33, 240–244. [Google Scholar] [CrossRef]
- Kini, R.M. Accelerated Evolution of Toxin Genes: Exonization and Intronization in Snake Venom Disintegrin/Metalloprotease Genes. Toxicon 2018, 148, 16–25. [Google Scholar] [CrossRef]
- Osmakov, D.I.; Kozlov, S.A.; Andreev, Y.A.; Koshelev, S.G.; Sanamyan, N.P.; Sanamyan, K.E.; Dyachenko, I.A.; Bondarenko, D.A.; Murashev, A.N.; Mineev, K.S.; et al. Sea Anemone Peptide with Uncommon β-Hairpin Structure Inhibits Acid-Sensing Ion Channel 3 (ASIC3) and Reveals Analgesic Activity. J. Biol. Chem. 2013, 288, 23116–23127. [Google Scholar] [CrossRef] [Green Version]
- Bray, B.L. Large-Scale Manufacture of Peptide Therapeutics by Chemical Synthesis. Nat. Rev. Drug Discov. 2003, 2, 587–593. [Google Scholar] [CrossRef]
- McGaraughty, S.; Chu, K.L.; Perner, R.J.; Didomenico, S.; Kort, M.E.; Kym, P.R. TRPA1 Modulation of Spontaneous and Mechanically Evoked Firing of Spinal Neurons in Uninjured, Osteoarthritic, and Inflamed Rats. Mol. Pain 2010, 6, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwan, K.Y.; Glazer, J.M.; Corey, D.P.; Rice, F.L.; Stucky, C.L. TRPA1 Modulates Mechanotransduction in Cutaneous Sensory Neurons. J. Neurosci. 2009, 29, 4808–4819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrus, M.; Peier, A.M.; Bandell, M.; Hwang, S.W.; Huynh, T.; Olney, N.; Jegla, T.; Patapoutian, A. A Role of TRPA1 in Mechanical Hyperalgesia Is Revealed by Pharmacological Inhibition. Mol. Pain 2007, 3, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knowlton, W.M.; Bifolck-Fisher, A.; Bautista, D.M.; McKemy, D.D. TRPM8, but Not TRPA1, Is Required for Neural and Behavioral Responses to Acute Noxious Cold Temperatures and Cold-Mimetics In Vivo. Pain 2010, 150, 340–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moparthi, L.; Survery, S.; Kreir, M.; Simonsen, C.; Kjellbom, P.; Hogestatt, E.D.; Johanson, U.; Zygmunt, P.M. Human TRPA1 Is Intrinsically Cold- and Chemosensitive with and without Its N-Terminal Ankyrin Repeat Domain. Proc. Natl. Acad. Sci. USA 2014, 111, 16901–16906. [Google Scholar] [CrossRef] [Green Version]
- Hinman, A.; Chuang, H.H.; Bautista, D.M.; Julius, D. TRP Channel Activation by Reversible Covalent Modification. Proc. Natl. Acad. Sci. USA 2006, 103, 19564–19568. [Google Scholar] [CrossRef] [Green Version]
- Nassini, R.; Materazzi, S.; Vriens, J.; Prenen, J.; Benemei, S.; De Siena, G.; la Marca, G.; Andre, E.; Preti, D.; Avonto, C.; et al. The “headache Tree” via Umbellulone and TRPA1 Activates the Trigeminovascular System. Brain 2012, 135, 376–390. [Google Scholar] [CrossRef] [Green Version]
- Bessac, B.F.; Sivula, M.; von Hehn, C.A.; Caceres, A.I.; Escalera, J.; Jordt, S.E. Transient Receptor Potential Ankyrin 1 Antagonists Block the Noxious Effects of Toxic Industrial Isocyanates and Tear Gases. FASEB J. 2009, 23, 1102–1114. [Google Scholar] [CrossRef] [Green Version]
- Leffler, A.; Lattrell, A.; Kronewald, S.; Niedermirtl, F.; Nau, C. Activation of TRPA1 by Membrane Permeable Local Anesthetics. Mol. Pain 2011, 7, 62. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.; Tian, J.; Zhu, Y.; Wang, C.; Xiao, R.; Herz, J.M.; Wood, J.D.; Zhu, M.X. Activation of TRPA1 Channels by Fenamate Nonsteroidal Anti-Inflammatory Drugs. Pflug. Arch. 2010, 459, 579–592. [Google Scholar] [CrossRef] [Green Version]
- Trevisan, G.; Hoffmeister, C.; Rossato, M.F.; Oliveira, S.M.; Silva, M.A.; Silva, C.R.; Fusi, C.; Tonello, R.; Minocci, D.; Guerra, G.P.; et al. TRPA1 Receptor Stimulation by Hydrogen Peroxide Is Critical to Trigger Hyperalgesia and Inflammation in a Model of Acute Gout. Free Radic. Biol. Med. 2014, 72, 200–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersson, D.A.; Gentry, C.; Moss, S.; Bevan, S. Transient Receptor Potential A1 Is a Sensory Receptor for Multiple Products of Oxidative Stress. J. Neurosci. 2008, 28, 2485–2494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motter, A.L.; Ahern, G.P. TRPA1 Is a Polyunsaturated Fatty Acid Sensor in Mammals. PLoS ONE 2012, 7, e38439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersson, D.A.; Gentry, C.; Alenmyr, L.; Killander, D.; Lewis, S.E.; Andersson, A.; Bucher, B.; Galzi, J.L.; Sterner, O.; Bevan, S.; et al. TRPA1 Mediates Spinal Antinociception Induced by Acetaminophen and the Cannabinoid Δ9-Tetrahydrocannabiorcol. Nat. Commun. 2011, 2, 551. [Google Scholar] [CrossRef] [Green Version]
- FLX-787 Significantly Reduces Muscle Cramp/Spasm Frequency and Improves Spasticity in a Phase 2 Study (Flex-201) in Patients with Multiple Sclerosis. ECTRIMS Online Library. Short G. 10 October 2018; 228473. Available online: https://onlinelibrary.ectrims-congress.eu/ectrims/2018/ectrims-2018/228473/glenn.short.flx-787.significantly.reduces.muscle.cramp.spasm.frequency.and.html (accessed on 22 June 2022).
- Eid, S.R.; Crown, E.D.; Moore, E.L.; Liang, H.A.; Choong, K.C.; Dima, S.; Henze, D.A.; Kane, S.A.; Urban, M.O. HC-030031, a TRPA1 Selective Antagonist, Attenuates Inflammatory- and Neuropathy-Induced Mechanical Hypersensitivity. Mol. Pain 2008, 4, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, H.; Hamalainen, M.M.; Saarnilehto, M.; Koivisto, A.; Pertovaara, A. Attenuation of Mechanical Hypersensitivity by an Antagonist of the TRPA1 Ion Channel in Diabetic Animals. Anesthesiology 2009, 111, 147–154. [Google Scholar] [CrossRef] [Green Version]
- Defalco, J.; Steiger, D.; Gustafson, A.; Emerling, D.E.; Kelly, M.G.; Duncton, M.A. Oxime Derivatives Related to AP18: Agonists and Antagonists of the TRPA1 Receptor. Bioorg. Med. Chem. Lett. 2010, 20, 276–279. [Google Scholar] [CrossRef]
- Chen, J.; Joshi, S.K.; Didomenico, S.; Perner, R.J.; Mikusa, J.P.; Gauvin, D.M.; Segreti, J.A.; Han, P.; Zhang, X.F.; Niforatos, W.; et al. Selective Blockade of TRPA1 Channel Attenuates Pathological Pain without Altering Noxious Cold Sensation or Body Temperature Regulation. Pain 2011, 152, 1165–1172. [Google Scholar] [CrossRef]
- Skerratt, S. Recent Progress in the Discovery and Development of TRPA1 Modulators. Prog. Med. Chem. 2017, 56, 81–115. [Google Scholar] [CrossRef]
- Logashina, Y.A.; Solstad, R.G.; Mineev, K.S.; Korolkova, Y.V.; Mosharova, I.V.; Dyachenko, I.A.; Palikov, V.A.; Palikova, Y.A.; Murashev, A.N.; Arseniev, A.S.; et al. New Disulfide-Stabilized Fold Provides Sea Anemone Peptide to Exhibit Both Antimicrobial and TRPA1 Potentiating Properties. Toxins 2017, 9, 154. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, M.L.; Shafee, T.; Papenfuss, A.T.; Norton, R.S. Evolution of Cnidarian Trans-Defensins: Sequence, Structure and Exploration of Chemical Space. Proteins 2019, 87, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J.; Blain, R.B. The Hormesis Database: The Occurrence of Hormetic Dose Responses in the Toxicological Literature. Regul. Toxicol. Pharmacol. 2011, 61, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Cookman, C.J.; Belcher, S.M. Classical Nuclear Hormone Receptor Activity as a Mediator of Complex Concentration Response Relationships for Endocrine Active Compounds. Curr. Opin. Pharmacol. 2014, 19, 112–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreev, Y.A.; Osmakov, D.I.; Koshelev, S.G.; Maleeva, E.E.; Logashina, Y.A.; Palikov, V.A.; Palikova, Y.A.; Dyachenko, I.A.; Kozlov, S.A. Analgesic Activity of Acid-Sensing Ion Channel 3 (ASIC3) Inhibitors: Sea Anemones Peptides Ugr9-1 and APETx2 versus Low Molecular Weight Compounds. Mar. Drugs 2018, 16, 500. [Google Scholar] [CrossRef] [Green Version]
- Andreev, Y.A.; Kozlov, S.A.; Vassilevski, A.A.; Grishin, E. V Cyanogen Bromide Cleavage of Proteins in Salt and Buffer Solutions. Anal. Biochem. 2010, 407, 144–146. [Google Scholar] [CrossRef]
- Mirdita, M.; Schütze, K.; Moriwaki, Y.; Heo, L.; Ovchinnikov, S.; Steinegger, M. ColabFold: Making protein folding accessible to all. Nat. Methods 2022, 19, 679–682. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Logashina, Y.A.; Lubova, K.I.; Maleeva, E.E.; Palikov, V.A.; Palikova, Y.A.; Dyachenko, I.A.; Andreev, Y.A. Analysis of Structural Determinants of Peptide MS 9a-1 Essential for Potentiating of TRPA1 Channel. Mar. Drugs 2022, 20, 465. https://doi.org/10.3390/md20070465
Logashina YA, Lubova KI, Maleeva EE, Palikov VA, Palikova YA, Dyachenko IA, Andreev YA. Analysis of Structural Determinants of Peptide MS 9a-1 Essential for Potentiating of TRPA1 Channel. Marine Drugs. 2022; 20(7):465. https://doi.org/10.3390/md20070465
Chicago/Turabian StyleLogashina, Yulia A., Kseniya I. Lubova, Ekaterina E. Maleeva, Viktor A. Palikov, Yulia A. Palikova, Igor A. Dyachenko, and Yaroslav A. Andreev. 2022. "Analysis of Structural Determinants of Peptide MS 9a-1 Essential for Potentiating of TRPA1 Channel" Marine Drugs 20, no. 7: 465. https://doi.org/10.3390/md20070465
APA StyleLogashina, Y. A., Lubova, K. I., Maleeva, E. E., Palikov, V. A., Palikova, Y. A., Dyachenko, I. A., & Andreev, Y. A. (2022). Analysis of Structural Determinants of Peptide MS 9a-1 Essential for Potentiating of TRPA1 Channel. Marine Drugs, 20(7), 465. https://doi.org/10.3390/md20070465