
Current Status and Perspective on the Use of Viral-Based Vectors in Eukaryotic Microalgae




Abstract
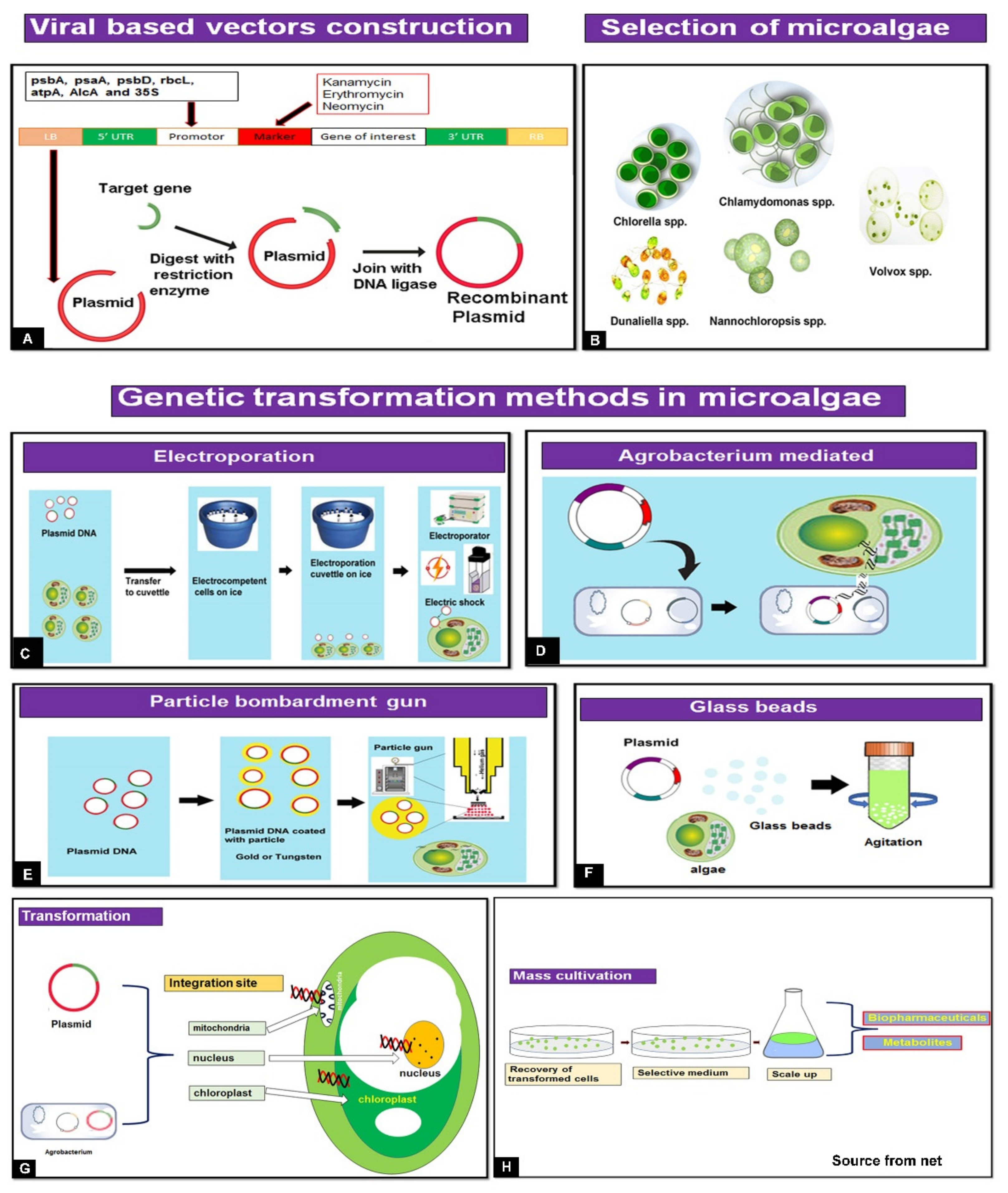
:1. Introduction
2. Genetic Engineering Transformation Methods for Biopharmaceutical Production of Microalgae
3. Microalgae Nuclear and Chloroplast-Based Expression
4. Algal Biotechnology in Pharmaceutical Applications
5. Viral-Based Expression Vectors for Recombinant Protein, Vaccine, and Biopharmaceutical Production
6. Design of a Viral-Based Vector for Microalgae Use
7. Challenges and Perspectives
8. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Morais, M.G.; Vaz, B.D.S.; de Morais, E.G.; Costa, J.A.V. Biologically active metabolites synthesized by microalgae. Biomed. Res. Int. 2015, 2015, 835761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grama, S.B.; Liu, Z.; Li, J. Emerging trends in genetic engineering of microalgae for commercial applications. Mar. Drugs 2022, 20, 285. [Google Scholar] [CrossRef] [PubMed]
- Camacho, F.; Macedo, A.; Malcata, F. Potential industrial applications and commercialization of microalgae in the functional food and feed industries: A short review. Mar. Drugs 2019, 17, 312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmussen, A.S.B.; Hammou, A.; Poulsen, T.F.; Laursen, M.C.; Hansen, S.F. Definition, categorization, and environ-mental risk assessment of biopharmaceuticals. Sci. Total Environ. 2021, 789, 147884. [Google Scholar] [CrossRef]
- Mizukami, A.; Caron, A.L.; Picanço-Castro, V.; Swiech, K. Platforms for recombinant therapeutic glycoprotein production. Methods Mol. Biol. 2018, 1674, 1–14. [Google Scholar] [CrossRef]
- Baeshen, M.N.; Al-Hejin, A.M.; Bora, R.S.; Ahmed, M.M.; Ramadan, H.A.; Saini, K.S.; Baeshen, N.A.; Redwan, E.M. Production of biopharmaceuticals in E. coli: Current scenario and future perspectives. J. Microbiol. Biotechnol. 2015, 25, 953–962. [Google Scholar] [CrossRef]
- Madhavan, A.; Arun, K.B.; Sindhu, R.; Krishnamoorthy, J.; Reshmy, R.; Sirohi, R.; Pugazhendi, A.; Awasthi, M.K.; Szakacs, G.; Binod, P. Customized yeast cell factories for biopharmaceuticals: From cell engineering to process scale up. Microb. Cell Fact. 2021, 20, 124. [Google Scholar] [CrossRef]
- Hanisch, F.G. Recombinant norovirus capsid protein VP1 (GII.4) expressed in H5 insect cells exhibits post-translational modifications with potential impact on lectin activity and vaccine design. Glycobiology 2022, 32, 496–505. [Google Scholar] [CrossRef]
- Rosales-Mendoza, S.; Solís-Andrade, K.I.; Márquez-Escobar, V.A.; González-Ortega, O.; Bañuelos-Hernandez, B. Current advances in the algae-made biopharmaceuticals field. Expert Opin. Biol. Ther. 2020, 20, 751–766. [Google Scholar] [CrossRef]
- Khavari, F.; Saidijam, M.; Taheri, M.; Nouri, F. Microalgae: Therapeutic potentials and applications. Mol. Biol. Rep. 2021, 48, 4757–4765. [Google Scholar] [CrossRef]
- Jareonsin, S.; Pumas, C. Advantages of heterotrophic microalgae as a host for phytochemicals production. Front. Bioeng. Biotechnol. 2021, 9, 628597. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, S.; Lauersen, K.J. Gene delivery technologies with applications in microalgal genetic engineering. Biology 2021, 10, 265. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Hu, H.; Gao, Y.; Xu, X.; Gao, H. Microalgae as platforms for production of recombinant proteins and valuable compounds: Progress and prospects. J. Ind. Microbiol. Biotechnol. 2011, 38, 1879–1890. [Google Scholar] [CrossRef] [PubMed]
- Bañuelos-Hernández, B.; Beltrán-López, J.I.; Rosales-Mendoza, S. Production of biopharmaceuticals in microalgae. In Handbook of Marine Microalgae; Academic Press: Cambridge, MA, USA, 2015; pp. 281–298. [Google Scholar]
- Doron, L.; Segal, N.; Shapira, M. Transgene expression in microalgae-from tools to applications. Front. Plant Sci. 2016, 7, 505. [Google Scholar] [CrossRef]
- Sproles, A.E.; Fields, F.J.; Smalley, T.N.; Le, C.H.; Badary, A.; Mayfield, S.P. Recent advancements in the genetic engineering of microalgae. Algal Res. 2021, 53, 102158. [Google Scholar] [CrossRef]
- Kselíková, V.; Singh, A.; Bialevich, V.; Čížková, M.; Bišová, K. Improving microalgae for biotechnology-From genetics to synthetic biology-Moving forward but not there yet. Biotechnol. Adv. 2022, 58, 107885. [Google Scholar] [CrossRef]
- Kindle, K.L. High-frequency nuclear transformation of Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 1990, 87, 1228–1232. [Google Scholar] [CrossRef] [Green Version]
- Kindle, K.L.; Schnell, R.A.; Fernandez, E.; Lefebvre, P.A. Stable nuclear transformation of Chlamydomonas using the Chlamydomonas gene for nitrate reductase. J. Cell Biol. 1989, 109, 2589–2601. [Google Scholar] [CrossRef]
- Shimogawara, K.; Fujiwara, S.; Grossman, A.; Usuda, H. High-efficiency transformation of Chlamydomonas reinhardtii by electroporation. Genetics 1998, 4, 1821–1828. [Google Scholar] [CrossRef]
- Hwang, H.H.; Yu, M.; Lai, E.M. Agrobacterium-mediated plant transformation: Biology and applications. In Arabidopsis Book; American Society of Plant Biologists: Rockville, MD, USA, 2017; Volume 15, p. e0186. [Google Scholar]
- Kathiresan, S.; Chandrashekar, A.; Ravishankar, G.A.; Sarada, R. Agrobacterium-mediated transformation in the green alga Haematococcus pluvialis (Chlorophyceae, Volvocales). J. Phycol. 2009, 45, 642–649. [Google Scholar] [CrossRef]
- Kumar, S.V.; Misquitta, R.W.; Reddy, V.S.; Rao, B.J.; Rajam, M.V. Genetic transformation of the green alga-Chlamydomonas reinhardtii by Agrobacterium tumefaciens. Plant Sci. 2004, 166, 731–738. [Google Scholar] [CrossRef]
- Kumar, S.V.; Rajam, M.V. Induction of Agrobacterium tumefaciens vir genes by the green alga, Chlamydomonas reinhardtii. Curr. Sci. 2007, 92, 1727–1729. [Google Scholar]
- Wang, P.; Wang, G.; Teng, Y.; Li, X.; Ji, J.; Xu, X.; Li, Y. Effects of cefotaxime and kanamycin on thallus proliferation and differentiation in Porphyra yezoensis and their inhibition on Agrobacterium tumefaciens. Mar. Biol. Res. 2010, 6, 100–105. [Google Scholar] [CrossRef]
- Sodeinde, O.A.; Kindle, K.L. Homologous recombination in the nuclear genome of Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 1993, 90, 9199–9203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sreenikethanam, A.; Raj, S.; Banu, R.J.; Gugulothu, P.; Bajhaiya, A.K. Genetic engineering of microalgae for secondary metabolite production: Recent developments, challenges, and future prospects. Front. Bioeng. Biotechnol. 2022, 10, 836056. [Google Scholar] [CrossRef]
- Kindle, K.L.; Richards, K.L.; Stern, D.B. Engineering the chloroplast genome: Techniques and capabilities for chloroplast transformation in Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 1991, 88, 1721–1725. [Google Scholar] [CrossRef] [Green Version]
- Economou, C.; Wannathong, T.; Szaub, J.; Purton, S. A simple low-cost method for chloroplast transformation of the green alga Chlamydomonas reinhardtii. In Methods in Molecular Biology; Springer International Publishing: Cham, Switzerland, 2014; Volume 1132, pp. 401–411. [Google Scholar] [CrossRef]
- Rochaix, J.D.; Surzycki, R.; Ramundo, S. Tools for regulated gene expression in the chloroplast of Chlamydomonas. In Methods in Molecular Biology; Springer International Publishing: Cham, Switzerland, 2014; Volume 1132, pp. 413–424. [Google Scholar] [CrossRef]
- Jarvis, E.E.; Brown, L.M. Transient expression of firefly luciferase in protoplasts of the green alga Chlorella ellipsoidea. Curr. Genet. 1991, 19, 317–321. [Google Scholar] [CrossRef]
- Wannathong, T.; Waterhouse, J.C.; Young, R.E.; Economou, C.K.; Purton, S. New tools for chloroplast genetic engineering allow the synthesis of human growth hormone in the green alga Chlamydomonas reinhardtii. Appl. Microbiol. Biotechnol. 2016, 100, 5467–5477. [Google Scholar] [CrossRef] [Green Version]
- Rosenberg, J.N.; Oyler, G.A.; Wilkinson, L.; Betenbaugh, M.J. A green light for engineered algae: Redirecting metabolism to fuel a biotechnology revolution. Curr. Opin. Biotechnol. 2008, 19, 430–436. [Google Scholar] [CrossRef]
- Barrera, D.J.; Mayfield, S.P. High-value recombinant protein production in microalgae. In Handbook of Microalgal Culture Applied Phycology and Biotechnology, 2nd ed.; Richmond, A., Hu, Q., Eds.; John Wiley & Sons: London, UK, 2013; pp. 532–544. [Google Scholar] [CrossRef]
- Malla, A.; Rosales-Mendoza, S.; Phoolcharoen, W.; Vimolmangkang, S. Efficient transient expression of recombinant proteins using DNA viral vectors in freshwater microalgal species. Front. Plant Sci. 2021, 12, 513. [Google Scholar] [CrossRef]
- Feng, S.; Xue, L.; Liu, H.; Lu, P. Improvement of efficiency of genetic transformation for Dunaliella salina by glass beads method. Mol. Biol. Rep. 2009, 36, 1433–1439. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Wang, J.; Jiang, P.; Bian, S.; Qin, S. Transformation of Platymonas (Tetraselmis) subcordiformis (Prasinophyceae, Chlorophyta) by agitation with glass beads. World J. Microbiol. Biotechnol. 2010, 26, 1653–1657. [Google Scholar] [CrossRef]
- Wang, L.; Yang, L.; Wen, X.; Chen, Z.; Liang, Q.; Li, J.; Wang, W. Rapid and high efficiency transformation of Chlamydomonas reinhardtii by square-wave electroporation. Biosci. Rep. 2019, 39, BSR20181210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Im, D.J.; Jeong, S.N.; Yoo, B.S.; Kim, B.; Kim, D.P.; Jeong, W.J.; Kang, I.S. Digital microfluidic approach for efficient electroporation with high productivity: Transgene expression of microalgae without cell wall removal. Anal. Chem. 2015, 87, 6592–6599. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Hu, H. High efficiency transformation by electroporation of the freshwater alga Nannochloropsis limnetica. World J. Microbiol. Biotechnol. 2019, 35, 1–10. [Google Scholar] [CrossRef]
- Geng, D.; Wang, Y.; Wang, P.; Li, W.; Sun, Y. Stable expression of hepatitis B surface antigen gene in Dunaliella salina (Chlorophyta). J. Appl. Phycol. 2003, 15, 451–456. [Google Scholar] [CrossRef]
- Guo, S.L.; Zhao, X.Q.; Tang, Y.; Wan, C.; Alam, M.A.; Ho, S.H.; Bai, F.W.; Chang, J.S. Establishment of an efficient genetic transformation system in Scenedesmus obliquus. J. Biotechnol. 2013, 163, 61–68. [Google Scholar] [CrossRef]
- Jaeger, D.; Hübner, W.; Huser, T.; Mussgnug, J.H.; Kruse, O. Nuclear transformation and functional gene expression in the oleaginous microalga Monoraphidium neglectum. J. Biotechnol. 2017, 249, 10–15. [Google Scholar] [CrossRef]
- Run, C.; Fang, L.; Fan, J.; Fan, C.; Luo, Y.; Hu, Z.; Li, Y. Stable nuclear transformation of the industrial alga Chlorella pyrenoidosa. Algal Res. 2016, 17, 196–201. [Google Scholar] [CrossRef]
- Chen, H.L.; Li, S.S.; Huang, R.; Tsai, H.J. Conditional production of a functional fish growth hormone in the transgenic line of nannochloropsis oculata (Eustigmatophyceae) 1. J. Phycol. 2008, 44, 768–776. [Google Scholar] [CrossRef]
- Anila, N.; Chandrashekar, A.; Ravishankar, G.A.; Sarada, R. Establishment of Agrobacterium tumefaciens-mediated genetic transformation in Dunaliella bardawil. Eur. J. Phycol. 2011, 46, 36–44. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, R.; Gothandam, K.M. Synergistic action of D-Glucose and acetosyringone on Agrobacterium strains for efficient Dunaliella transformation. PLoS ONE 2016, 11, e0158322. [Google Scholar] [CrossRef] [PubMed]
- Schiedlmeier, B.; Schmitt, R.; Müller, W.; Kirk, M.M.; Gruber, H.; Mages, W.; Kirk, D.L. Nuclear transformation of Volvox carteri. Proc. Natl. Acad. Sci. USA 1994, 91, 5080–5084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boynton, J.E.; Gillham, N.W.; Harris, E.H.; Hosler, J.P.; Johnson, A.M.; Jones, A.R.; Randolph-Anderson, B.L.; Robertson, D.; Klein, T.M.; Shark, K.B.; et al. Chloroplast transformation in Chlamydomonas with high velocity microprojectiles. Science 1988, 240, 1534–1538. [Google Scholar] [CrossRef]
- Mini, P.; Demurtas, O.C.; Valentini, S.; Pallara, P.; Aprea, G.; Ferrante, P.; Giuliano, G. Agrobacterium-mediated and electroporation mediated transformation of Chlamydomonas reinhardtii: A comparative study. BMC Biotechnol. 2018, 18, 11. [Google Scholar] [CrossRef]
- Gregory, J.A.; Topol, A.B.; Doerner, D.Z.; Mayfield, S. Alga-produced cholera toxin-Pfs25 fusion proteins as oral vaccines. Appl. Environ. Microbiol. 2013, 79, 3917–3925. [Google Scholar] [CrossRef] [Green Version]
- Hempel, F.; Maier, U.G. An engineered diatom acting like a plasma cell secreting human IgG antibodies with high efficiency. Microb. Cell Fact. 2012, 11, 126. [Google Scholar] [CrossRef] [Green Version]
- Dunahay, T.G.; Jarvis, E.E.; Roessler, P.G. Genetic transformation of the diatoms Cyclotella cryptica and Navicula saprophila. J. Phycol. 1995, 31, 1004–1012. [Google Scholar] [CrossRef]
- Fischer, H.; Robl, I.; Sumper, M.; Kröger, N. Targeting and covalent modification of cell wall and membrane proteins heterologously expressed in the diatom Cylindrotheca fusiformis (Bacillariophyceae). J. Phycol. 1999, 35, 113–120. [Google Scholar] [CrossRef]
- Dunahay, T.G. Transformation of Chlamydomonas reinhardtii with silicon carbide whiskers. Biotechniques 1993, 15, 452–455. [Google Scholar]
- Wang, K.; Drayton, P.; Frame, B.; Dunwell, J.; Thompson, J. Whisker mediated plant transformation an alternative technology. In Vitro Cell Dev. Biol. 1995, 31, 101–104. [Google Scholar] [CrossRef]
- Chai, X.J.; Chen, H.X.; Xu, W.Q.; Xu, Y.W. Expression of soybean Kunitz trypsin inhibitor gene SKTI in Dunaliella salina. J. Appl. Phycol. 2013, 25, 139–144. [Google Scholar] [CrossRef]
- Pratheesh, P.T.; Vineetha, M.; Kurup, G.M. An efficient protocol for the Agrobacterium-mediated genetic transformation of microalga Chlamydomonas reinhardtii. Mol. Biotechnol. 2014, 56, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Steinbrenner, J.; Sandmann, G. Transformation of the green alga Haematococcus pluvialis with a phytoene desaturase for accelerated astaxanthin biosynthesis. Appl. Environ. Microbiol. 2006, 72, 7477–7484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cha, T.S.; Yee, W.; Aziz, A. Assessment of factors affecting Agrobacterium-mediated genetic transformation of the unicellular green alga, Chlorella vulgaris. World J. Microbiol. Biotechnol. 2012, 28, 1771–1779. [Google Scholar] [CrossRef]
- Rathod, J.P.; Prakash, G.; Pandit, R.; Lali, A.M. Agrobacterium-mediated transformation of promising oil-bearing marine algae Parachlorella kessleri. Photosynth. Res. 2013, 118, 141–146. [Google Scholar] [CrossRef]
- Simon, D.P.; Anila, N.; Gayathri, K.; Sarada, R. Heterologous expression of β-carotene hydroxylase in Dunaliella salina by Agrobacterium-mediated genetic transformation. Algal Res. 2016, 18, 257–265. [Google Scholar] [CrossRef]
- Khatiwada, B.; Kautto, L.; Sunna, A.; Sun, A.; Nevalainen, H. Nuclear transformation of the versatile microalga Euglena gracilis. Algal Res. 2019, 37, 178–185. [Google Scholar] [CrossRef]
- Dautor, Y.; Úbeda-Mínguez, P.; Chileh, T.; García-Maroto, F.; Alonso, D.L. Development of genetic transformation methodologies for an industrially-promising microalga: Scenedesmus almeriensis. Biotechnol. Lett. 2014, 36, 2551–2558. [Google Scholar] [CrossRef]
- Bashir, K.M.I.; Kim, M.-S.; Stahl, U.; Cho, M.-G. Agrobacterium mediated genetic transformation of Dictyosphaerium pulchellum for the expression of erythropoietin. J. Appl. Phycol. 2018, 30, 3503–3518. [Google Scholar] [CrossRef]
- Rathod, J.P.; Gade, R.M.; Rathod, D.R.; Dudhare, M. A review on molecular tools of microalgal genetic transformation and their application for overexpression of different genes. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 3191–3207. [Google Scholar] [CrossRef]
- Brown, L.E.; Sprecher, S.L.; Keller, L.R. Introduction of exogenous DNA into Chlamydomonas reinhardtii by electroporation. Mol. Cell Biol. 1991, 11, 2328–2332. [Google Scholar] [CrossRef] [PubMed]
- Weaver, J.C. Plant Cell Electroporation and Electrofusion Protocols. In Methods in Molecular Biology; Nickoloff, J.A., Ed.; Springer: Totowa, NJ, USA, 1995; Volume 55. [Google Scholar] [CrossRef]
- Niu, Y.F.; Zhang, M.H.; Xie, W.H.; Li, J.N.; Gao, Y.F.; Yang, W.D.; Liu, J.S.; Li, H.Y. A new inducible expression system in a transformed green alga, Chlorella vulgaris. Genet. Mol. Res. 2011, 10, 3427–3434. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Sun, Z.; Gerken, H.; Huang, J.; Jiang, Y.; Chen, F. Genetic engineering of the green alga Chlorella zofingiensis: A modified norflurazon-resistant phytoene desaturase gene as a dominant selectable marker. Appl. Microbiol. Biotechnol. 2014, 98, 5069–5079. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wang, Y.; Zhang, Y.; Chen, X.; Zhang, P.; Ma, S. Development of a new method for genetic transformation of the green alga Chlorella ellipsoidea. Mol. Biotechnol. 2013, 54, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Yamano, T.; Fukuzawa, H. Transformation of the model microalga Chlamydomonas reinhardtii without cell-wall removal. In Electroporation Protocols; Li, S., Cutrera, J., Heller, R., Teissie, J., Eds.; Humana: New York, NY, USA, 2020. [Google Scholar] [CrossRef]
- Day, A.; Debuchy, R.; van Dillewijn, J.; Purton, S.; Rochaix, J.-D. Studies on the maintenance and expression of cloned DNA fragments in the nuclear genome of the green alga Chlamydomonas reinhardtii. Physiol. Plant. 1990, 78, 254–260. [Google Scholar] [CrossRef]
- Remacle, C.; Cardol, P.; Coosemans, N.; Gaisne, M.; Bonnefoy, N. High-efficiency biolistic transformation of Chlamydomonas mitochondria can be used to insert mutations in complex I genes. Proc. Natl. Acad. Sci. USA 2006, 103, 4771–4776. [Google Scholar] [CrossRef] [Green Version]
- Jones, C.S.; Luong, T.; Hannon, M.; Tran, M.; Gregory, J.A.; Shen, Z.; Briggs, S.P.; Mayfield, S.P. Heterologous expression of the C terminal antigenic domain of the malaria vaccine candidate Pfs48/45 in the green algae Chlamydmonas reinhardtii. Appl. Microbiol. Biotechnol. 2013, 97, 1987–1995. [Google Scholar] [CrossRef]
- Dreesen, I.A.; Charpin-El Hamri, G.; Fussenegger, M. Heat-stable oral alga-based vaccine protects mice from Staphylococcus aureus infection. J. Biotechnol. 2010, 145, 273–280. [Google Scholar] [CrossRef]
- Tan, C.; Qin, S.; Zhang, Q.; Jiang, P.; Zhao, F. Establishment of a micro-particle bombardment transformation system for Dunaliella salina. J. Microbiol. 2005, 43, 361–365. [Google Scholar]
- Bayne, A.C.V.; Boltz, D.; Owen, C.; Betz, Y.; Maia, G.; Azadi, P.; Archer-Hartmann, S.; Zirkle, R.; Lippmeier, J.C. Vaccination against influenza with recombinant hemagglutinin expressed by Schizochytrium sp. confers protective immunity. PLoS ONE 2013, 8, e61790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bañuelos-Hernández, B.; Monreal-Escalante, E.; González-Ortega, O.; Angulo, C.; Rosales-Mendoza, S. Algevir: An expression system for microalgae based on viral vectors. Front. Microbiol. 2017, 8, 1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baek, K.; Kim, D.H.; Jeong, J.; Sim, S.J.; Melis, A.; Kim, J.S.; Jin, E.; Bae, S. DNA-free two-gene knockout in Chlamydomonas reinhardtii via CRISPR-Cas9 ribonucleoproteins. Sci. Rep. 2016, 6, 30620. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.E.; Lim, J.M.; Koh, H.G.; Kim, E.K.; Kang, N.K.; Jeon, S.; Kwon, S.; Shin, W.S.; Lee, B.; Hwangbo, K.; et al. CRISPR/Cas9-induced knockout and knock-inmutations in Chlamydomonas reinhardtii. Sci. Rep. 2016, 6, 27810. [Google Scholar] [CrossRef]
- Serif, M.; Dubois, G.; Finoux, A.-L.; Teste, M.-A.; Jallet, D.; Daboussi, F. One-step generation of multiple gene knock-outs in the diatom Phaeodactylum tricornutum by DNA-free genome editing. Nat. Commun. 2018, 9, 3924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, K.S.; Kim, J.; Park, H.; Hong, S.-J.; Lee, C.-G.; Jin, E. Enhanced lipid productivity in AGP knockout marine microalga Tetraselmis sp. using a DNA-free CRISPR-Cas9 RNP method. Bioresour. Technol. 2020, 303, 122932. [Google Scholar] [CrossRef]
- Hawkins, R.L.; Nakamura, M. Expression of human growth hormone by the eukaryotic alga, Chlorella. Curr. Microbiol. 1999, 38, 335–341. [Google Scholar] [CrossRef]
- Endo, H.; Yoshida, M.; Uji, T.; Saga, N.; Inoue, K.; Nagasawa, H. Stable nuclear transformation system for the coccolithophorid alga Pleurochrysis carterae. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef]
- Karas, B.J.; Diner, R.E.; Lefebvre, S.C.; McQuaid, J.; Phillips, A.P.; Noddings, C.M.; Brunson, J.K.; Valas, R.E.; Deerinck, T.J.; Jablanovic, J.; et al. Designer diatom episomes delivered by bacterial conjugation. Nat. Commun. 2015, 6, 6925. [Google Scholar] [CrossRef]
- Muñoz, C.F.; Sturme, M.H.J.; D’Adamo, S.; Weusthuis, R.A.; Wijffels, R.H. Stable transformation of the green algae Acutodesmus obliquus and Neochloris oleoabundans based on E coli conjugation. Algal Res. 2019, 39, 101453. [Google Scholar] [CrossRef]
- Zhou, X.; Zhang, X.; Boualavong, J.; Durney, A.R.; Wang, T.; Kirschner, S.; Wentz, M.; Mukaibo, H. Electrokinetically controlled fluid injection into unicellular microalgae. Electrophoresis 2017, 38, 2587–2591. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.C.; Long, J.R.; Yaghi, O.M. Introduction to metal-organic frameworks. Chem. Rev. 2012, 112, 673–674. [Google Scholar] [CrossRef]
- Silva, S.; Almeida, A.J.; Vale, N. Combination of cell-penetrating peptides with nanoparticles for therapeutic application: A review. Biomolecules 2019, 9, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanigan, T.M.; Kopera, H.C.; Saunders, T.L. Principles of Genetic Engineering. Genes 2020, 11, 291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radakovits, R.; Jinkerson, R.E.; Fuerstenberg, S.I.; Tae, H.; Settlage, R.E.; Boore, J.L.; Posewitz, M.C. Draft genome sequence and genetic transformation of the oleaginous alga Nannochloropis gaditana. Nat. Commun. 2012, 3, 686. [Google Scholar] [CrossRef] [Green Version]
- León-Bañares, R.; González-Ballester, D.; Galván, A.; Fernández, E. Transgenic microalgae as green cell-factories. Trends Biotechnol. 2004, 22, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Potvin, G.; Zhang, Z. Strategies for high-level recombinant protein expression in transgenic microalgae: A review. Biotechnol. Adv. 2010, 28, 910–918. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K.; Schwender, J.; Disch, A.; Rohmer, M. Biosynthesis of isoprenoids in higher plant chloroplasts proceeds via a mevalonate-independent pathway. FEBS Lett. 1997, 400, 271–274. [Google Scholar] [CrossRef] [Green Version]
- Bowsher, C.G.; Tobin, A.K. Compartmentation of metabolism within mitochondria and plastids. J. Exp. Bot. 2001, 52, 513–527. [Google Scholar] [CrossRef]
- Gallaher, S.D.; Fitz-Gibbon, S.T.; Strenkert, D.; Purvine, S.O.; Pellegrini, M.; Merchant, S.S. High-throughput sequencing of the chloroplast and mitochondrion of Chlamydomonas reinhardtii to generate improved de novo assemblies, analyze expression patterns and transcript speciation, and evaluate diversity among laboratory strains and wild isolates. Plant. J. 2018, 93, 545–565. [Google Scholar] [CrossRef] [Green Version]
- Kwon, Y.M.; Kim, K.W.; Choi, T.-Y.; Kim, S.Y.; Kim, J.Y.H. Manipulation of the microalgal chloroplast by genetic engineering for biotechnological utilization as a green biofactory. World J. Microbiol. Biotechnol. 2018, 34, 183. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Qian, K.; Su, N.; Chang, H.; Liu, J.; Shen, G. Foot-and-mouth disease virus VP1 protein fused with cholera toxin B subunit expressed in Chlamydmonas reinhardtii chloroplast. Biotechnol. Lett. 2003, 25, 1087–1092. [Google Scholar] [CrossRef] [PubMed]
- Bertalan, I.; Munder, M.C.; Weiß, C.; Kopf, J.; Fischer, D.; Johanningmeier, U. A rapid, modular and marker-free chloroplast expression system for the green alga Chlamydomonas reinhardtii. J. Biotechnol. 2015, 195, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.-H.; Zhu, C.-C.; Zhang, N.-S.; Li, D.-W.; Yang, W.-D.; Liu, J.-S.; Sathishkumar, R.; Li, H.Y. Construction of novel chloroplast expression vector and development of an efficient transformation system for the diatom Phaeodactylum tricornutum. Mar. Biotechnol. 2014, 16, 538–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiehn, O. Metabolomics—The link between genotypes and phenotypes. Plant Mol. Biol. 2002, 48, 155–171. [Google Scholar] [CrossRef] [PubMed]
- Senousy, H.H.; Abd Ellatif, S.; Ali, S. Assessment of the antioxidant and anticancer potential of different isolated strains of cyanobacteria and microalgae from soil and agriculture drain water. Environ. Sci. Pollut. Res. Int. 2020, 27, 18463–18474. [Google Scholar] [CrossRef] [PubMed]
- Shanab, S.M.; Mostafa, S.S.; Shalaby, E.A.; Mahmoud, G.I. Aqueous extracts of microalgae exhibit antioxidant and anticancer activities. Asian Pac. J. Trop. Biomed. 2012, 2, 608–615. [Google Scholar] [CrossRef] [Green Version]
- Suh, S.S.; Kim, S.M.; Kim, J.E.; Hong, J.M.; Lee, S.G.; Youn, U.J.; Han, S.J.; Kim, I.C.; Kim, S. Anticancer activities of ethanol extract from the Antarctic freshwater microalga, Botryidiopsidaceae sp. BMC Complement. Altern. Med. 2017, 17, 509. [Google Scholar] [CrossRef] [Green Version]
- Zaharieva, M.M.; Zheleva-Dimitrova, D.; Rusinova-Videva, S.; Ilieva, Y.; Brachkova, A.; Balabanova, V.; Gevrenova, R.; Kim, T.C.; Kaleva, M.; Georgieva, A.; et al. Antimicrobial and antioxidant potential of Scenedesmus obliquus microalgae in the context of integral biorefinery concept. Molecules 2022, 27, 519. [Google Scholar] [CrossRef]
- Hwang, J.; Yadav, D.; Lee, P.C.; Jin, J.O. Immunomodulatory effects of polysaccharides from marine algae for treating cancer, infectious disease, and inflammation. Phytother. Res. 2022, 36, 761–777. [Google Scholar] [CrossRef]
- Mehariya, S.; Goswami, R.K.; Karthikeysan, O.P.; Verma, P. Microalgae for high-value products: A way towards green nutraceutical and pharmaceutical compounds. Chemosphere 2021, 280, 130553. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Manna, M.S.; Bhowmick, T.K.; Gayen, K. Priority-based multiple products from microalgae: Review on techniques and strategies. Crit. Rev. Biotechnol. 2020, 40, 590–607. [Google Scholar] [CrossRef] [PubMed]
- Castro, J.S.; Calijuri, M.L.; Ferreira, J.; Assemany, P.P.; Ribeiro, V.J. Microalgae based biofertilizer: A life cycle approach. Sci. Total Environ. 2020, 724, 138138. [Google Scholar] [CrossRef]
- Lakatos, G.E.; Ranglová, K.; Manoel, J.C.; Grivalský, T.; Kopecký, J.; Masojídek, J. Bioethanol production from microalgae polysaccharides. Folia Microbiol. 2019, 64, 627–644. [Google Scholar] [CrossRef] [PubMed]
- Plöhn, M.; Spain, O.; Sirin, S.; Silva, M.; Escudero-Oñate, C.; Ferrando-Climent, L.; Allahverdiyeva, Y.; Funk, C. Wastewater treatment by microalgae. Physiol. Plant. 2021, 173, 568–578. [Google Scholar] [CrossRef] [PubMed]
- Hom-Diaz, A.; Llorca, M.; Rodríguez-Mozaz, S.; Vicent, T.; Barceló, D.; Blánquez, P. Microalgae cultivation on wastewater digestate: β-estradiol and 17α-ethynylestradiol degradation and transformation products identification. J. Environ. Manag. 2015, 15, 106–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heimann, K.; Huerlimann, R. Microalgal classification: Major classes and genera of commercial microalgal species. In Handbook of Marine Microalgae; Kim, S.-K., Ed.; Academic Press: Busan, South Korea, 2015; pp. 25–41. [Google Scholar]
- Márquez-Escobar, V.A.; Bañuelos-Hernández, B.; Rosales-Mendoza, S. Expression of a Zika virus antigen in microalgae: Towards mucosal vaccine development. J. Biotechnol. 2018, 282, 86–91. [Google Scholar] [CrossRef]
- Castellanos-Huerta, I.; Bañuelos-Hernández, B.; Téllez, G.; Rosales-Mendoza, S.; Brieba, L.G.; Esquivel-Ramos, E.; Beltrán-López, J.I.; Velazquez, G.; Fernandez-Siurob, I. Recombinant Hemagglutinin of Avian Influenza Virus H5 expressed in the chloroplast of Chlamydomonas reinhardtii and evaluation of its immunogenicity in chickens. Avian. Dis. 2016, 60, 784–791. [Google Scholar] [CrossRef]
- Demurtas, O.C.; Massa, S.; Ferrante, P.; Venuti, A.; Franconi, R.; Giuliano, G. A Chlamydomonas-derived Human Papillomavirus 16 E7 vaccine induces specific tumor protection. PLoS ONE 2013, 8, e61473. [Google Scholar] [CrossRef] [Green Version]
- Barahimipour, R.; Neupert, J.; Bock, R. Efficient expression of nuclear transgenes in the green alga Chlamydomonas: Synthesis of an HIV antigen and development of a new selectable marker. Plant Mol. Biol. 2016, 90, 403–418. [Google Scholar] [CrossRef] [Green Version]
- Davis, A.; Crum, L.T.; Corbeil, L.B.; Hildebrand, M. Expression of Histophilus somni IbpA DR2 protective antigen in the diatom Thalassiosira pseudonana. Appl. Microbiol. Biotechnol. 2017, 101, 5313–5324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dauvillée, D.; Delhaye, S.; Gruyer, S.; Slomianny, C.; Moretz, S.E.; d’Hulst, C.; Long, C.A.; Ball, S.G.; Tomavo, S. Engineering the chloroplast targeted malarial vaccine antigens in Chlamydomonas starch granules. PLoS ONE 2010, 5, e15424. [Google Scholar] [CrossRef] [PubMed]
- Shamriz, S.; Ofoghi, H. Engineering the chloroplast of Chlamydomonas reinhardtii to express the recombinant PfCelTOS-Il2 antigen-adjuvant fusion protein. J. Biotechnol. 2018, 266, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Hempel, F.; Maurer, M.; Brockmann, B.; Mayer, C.; Biedenkopf, N.; Kelterbaum, A.; Becker, S.; Maier, U.G. From hybridomas to a robust microalgal-based production platform: Molecular design of a diatom secreting monoclonal antibodies directed against the Marburg virus nucleoprotein. Microb. Cell Fact. 2017, 16, 131. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Kim, Y.T.; Cho, J.J.; Bae, J.H.; Hur, S.B.; Hwang, I.; Choi, T.J. Stable integration and functional expression of flounder growth hormone gene in transformed microalga, Chlorella ellipsoidea. Mar. Biotechnol. 2002, 4, 63–73. [Google Scholar] [CrossRef]
- Tran, M.; Van, C.; Barrera, D.J.; Pettersson, P.L.; Peinado, C.D.; Bui, J.; Mayfield, S.P. Production of unique immunotoxin cancer therapeutics in algal chloroplasts. Proc. Natl. Acad. Sci. USA 2013, 110, E15–E22. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Brandsma, M.; Tremblay, R.; Maxwell, D.; Jevnikar, A.M.; Huner, N.; Ma, S. A novel expression platform for the production of diabetes-associated autoantigen human glutamic acid decarboxylase (hGAD65). BMC Biotechnol. 2008, 8, 87. [Google Scholar] [CrossRef] [Green Version]
- Kwon, K.C.; Lamb, A.; Fox, D.; Jegathese, S.J.P. An evaluation of microalgae as a recombinant protein oral delivery platform for fish using green fluorescent protein (GFP). Fish Shellfish Immunol. 2019, 87, 414–420. [Google Scholar] [CrossRef]
- Castellanos-Huerta, I.; Gómez-Verduzco, G.; Tellez-Isaias, G.; Ayora-Talavera, G.; Bañuelos-Hernández, B.; Petrone-García, V.M.; Velázquez-Juárez, G.; Fernández-Siurob, I. Transformation of Dunaliella salina by Agrobacterium tumefaciens for the expression of the Hemagglutinin of Avian Influenza Virus H5. Microorganisms 2022, 10, 361. [Google Scholar] [CrossRef]
- Feng, S.; Feng, W.; Zhao, L.; Gu, H.; Li, Q.; Shi, K.; Guo, S.; Zhang, N. Preparation of transgenic Dunaliella salina for immunization against white spot syndrome virus in crayfish. Arch. Virol. 2014, 159, 519–525. [Google Scholar] [CrossRef]
- Yang, Z.; Chen, F.; Li, D.; Zhang, Z.; Liu, Y.; Zheng, D.; Wang, Y.; Shen, G. Expression of human soluble TRAIL in Chlamydomonas reinhardtii chloroplast. Chin. Sci. Bull. 2006, 51, 1703–1709. [Google Scholar] [CrossRef]
- Soria-Guerra, R.E.; Ramírez-Alonso, J.I.; Ibáñez-Salazar, A.; Govea-Alonso, D.O.; Paz-Maldonado, L.M.; Bañuelos-Hernández, B.; Korban, S.S.; Rosales-Mendoza, S. Expression of an HBcAg-based antigen carrying angiotensin II in Chlamydomonas reinhardtii as a candidate hypertension vaccine. Plant Cell Tissue Organ Cult. 2014, 116, 133–139. [Google Scholar] [CrossRef]
- El-Ayouty, Y.; El-Manawy, I.; Nasih, S.; Hamdy, E.; Kebeish, R. Engineering Chlamydomonas reinhardtii for expression of functionally active human interferon-α? Mol. Biotechnol. 2019, 61, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Kiataramgul, A.; Maneenin, S.; Purton, S.; Areechon, N.; Hirono, I.; Brocklehurst, T.W.; Unajak, S. An oral delivery system for controlling white spot syndrome virus infection in shrimp using transgenic microalgae. Aquaculture 2020, 521, 735022. [Google Scholar] [CrossRef]
- Charoonnart, P.; Worakajit, N.; Zedler, J.; Meetam, M.; Robinson, C.; Saksmerprome, V. Generation of microalga Chlamydomonas reinhardtii expressing shrimp antiviral dsRNA without supplementation of antibiotics. Sci. Rep. 2019, 9, 3164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gregory, J.A.; Li, F.; Tomosada, L.M.; Cox, C.J.; Topol, A.B.; Vinetz, J.M.; Mayfield, S.P. Algae-produced Pfs25 elicits antibodies that inhibit malaria transmission. PLoS ONE 2012, 7, e37179. [Google Scholar] [CrossRef] [Green Version]
- Jarquín-Cordero, M.; Chávez, M.N.; Centeno-Cerdas, C.; Bohne, A.V.; Hopfner, U.; Machens, H.G.; Egaña, J.T.; Nickelsen, J. Towards a biotechnological platform for the production of human pro-angiogenic growth factors in the green alga Chlamydomonas reinhardtii. Appl. Microbiol. Biotechnol. 2020, 104, 725–739. [Google Scholar] [CrossRef]
- Baier, T.; Kros, D.; Feiner, R.C.; Lauersen, K.J.; Mu?ller, K.M.; Kruse, O. Engineered fusion proteins for efficient protein secretion and purification of a human growth factor from the green microalga Chlamydomonas reinhardtii. ACS Synth. Biol. 2018, 7, 2547–2557. [Google Scholar] [CrossRef]
- Stoffels, L.; Taunt, H.N.; Charalambous, B.; Purton, S. Synthesis of bacteriophage lytic proteins against Streptococcus pneumoniae in the chloroplast of Chlamydomonas reinhardtii. Plant Biotechnol. J. 2017, 15, 1130–1140. [Google Scholar] [CrossRef] [Green Version]
- Li, A.; Huang, R.; Wang, C.; Hu, Q.; Li, H.; Li, X. Expression of anti-lipopolysaccharide factor isoform 3 in Chlamydomonas reinhardtii showing high antimicrobial activity. Mar. Drugs 2021, 19, 239. [Google Scholar] [CrossRef]
- Lima, S.; Webb, C.L.; Deery, E.; Robinson, C.; Zedler, J.A. Human intrinsic factor expression for bioavailable vitamin B12 enrichment in microalgae. Biology 2018, 7, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hempel, F.; Lau, J.; Klingl, A.; Maier, U.G. Algae as protein factories: Expression of a human antibody and the respective antigen in the diatom Phaeodactylum tricornutum. PLoS ONE 2011, 6, e28424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayfield, S.P.; Franklin, S.E. Expression of human antibodies in eukaryotic micro-algae. Vaccine 2005, 23, 1828–1832. [Google Scholar] [CrossRef] [PubMed]
- Mayfield, S.P.; Franklin, S.E.; Lerner, R.A. Expression and assembly of a fully active antibody in algae. Proc. Natl. Acad. Sci. USA 2003, 100, 438–442. [Google Scholar] [CrossRef] [Green Version]
- Manuell, A.L.; Beligni, M.V.; Elder, J.H.; Siefker, D.T.; Tran, M.; Weber, A.; McDonald, T.L.; Mayfield, S.P. Robust expression of a bioactive mammalian protein in Chlamydomonas chloroplast. Plant Biotechnol. J. 2007, 5, 402–412. [Google Scholar] [CrossRef]
- Su, Z.L.; Qian, K.X.; Tan, C.P.; Meng, C.X.; Qin, S. Recombination and heterologous expression of allophycocyanin gene in the chloroplast of Chlamydomonas reinhardtii. Acta Biochim. Biophy. Sin. 2005, 37, 709–712. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.K.; Shen, G.F.; Ru, B.G. Survival of human metallothionein–2 transplastomic Chlamydomonas reinhardtii to ultraviolet B exposure. Acta Biochim. Biophy. Sin. 2006, 38, 187–193. [Google Scholar] [CrossRef] [Green Version]
- Beltrán-López, J.I.; Romero-Maldonado, A.; Monreal-Escalante, E.; Bañuelos-Hernández, B.; Paz-Maldonado, L.M.; Rosales-Mendoza, S. Chlamydomonas reinhardtii chloroplasts express an orally immunogenic protein targeting the p210 epitope implicated in atherosclerosis immunotherapies. Plant Cell Rep. 2016, 35, 1133–1141. [Google Scholar] [CrossRef]
- Gregory, J.A.; Shepley-McTaggart, A.; Umpierrez, M.; Hurlburt, B.K.; Maleki, S.J.; Sampson, H.A.; Mayfield, S.P.; Berin, M.C. Immunotherapy using algal-produced Ara h 1 core domain suppresses peanut allergy in mice. Plant Biotechnol. J. 2016, 14, 1541–1550. [Google Scholar] [CrossRef] [Green Version]
- Hirschl, S.; Ralser, C.; Asam, C.; Gangitano, A.; Huber, S.; Ebner, C.; Bohle, B.; Wolf, M.; Briza, P.; Ferreira, F.; et al. Expression and characterization of functional recombinant Bet v 1.0101 in the chloroplast of Chlamydomonas reinhardtii. Int. Arch. Allergy Immunol. 2017, 173, 44–50. [Google Scholar] [CrossRef]
- Rasala, B.A.; Muto, M.; Lee, P.A.; Jager, M.; Cardoso, R.M.; Behnke, C.A.; Kirk, P.; Hokanson, C.A.; Crea, R.; Mendez, M.; et al. Production of therapeutic proteins in algae, analysis of expression of seven human proteins in the chloroplast of Chlamydomonas reinhardtii. Plant Biotechnol. J. 2010, 8, 719–733. [Google Scholar] [CrossRef] [Green Version]
- Faè, M.; Accossato, S.; Cella, R.; Fontana, F.; Goldschmidt-Clermont, M.; Leelavathi, S.; Reddy, V.S.; Longoni, P. Comparison of transplastomic Chlamydomonas reinhardtii and Nicotiana tabacum expression system for the production of a bacterial endoglucanase. Appl. Microbiol. Biotechnol. 2017, 101, 4085–4092. [Google Scholar] [CrossRef]
- Kumar, A.; Falcao, V.R.; Sayre, R.T. Evaluating nuclear transgene expression systems in Chlamydomonas reinhardtii. Algal Res. 2013, 2, 321–332. [Google Scholar] [CrossRef]
- Dong, B.; Cheng, R.Q.; Liu, Q.Y.; Wang, J.; Fan, Z.C. Multimer of the antimicrobial peptide Mytichitin-A expressed in Chlamydomonas reinhardtii exerts a broader antibacterial spectrum and increased potency. J. Biosci. Bioeng. 2018, 125, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Xue, B.; Dong, C.M.; Hu, H.H.; Dong, B.; Fan, Z.C. Chlamydomonas reinhardtii-expressed multimer of ToAMP4 inhibits the growth of bacteria of both Gram-positive and Gram-negative. Process Biochem. 2020, 91, 311–318. [Google Scholar] [CrossRef]
- Pang, X.; Tong, Y.; Xue, W.; Yang, Y.F.; Chen, X.; Liu, J.; Chen, D. Expression and characterization of recombinant human lactoferrin in edible alga Chlamydomonas reinhardtii. Biosci. Biotechnol. Biochem. 2019, 83, 851–859. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Ramírez, J.; Wong-Arce, A.; González-Ortega, O.; Rosales-Mendoza, S. Expression in algae of a chimeric protein carrying several epitopes from tumor associated antigens. Int. J. Biol. Macromol. 2020, 147, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Berlanga, B.; Bañuelos-Hernández, B.; Rosales-Mendoza, S. Efficient expression of an Alzheimer’s disease vaccine candidate in the microalga Schizochytrium sp. using the Algevir system. Mol. Biotechnol. 2018, 60, 362–368. [Google Scholar] [CrossRef]
- He, Y.; Peng, H.; Liu, J.; Chen, F.; Zhou, Y.; Ma, X.; Chen, H.; Wang, K. Chlorella sp. transgenic with Scy-hepc enhancing the survival of Sparus macrocephalus and hybrid grouper challenged with Aeromonas hydrophila. Fish Shellfish Immunol. 2018, 73, 22–29. [Google Scholar] [CrossRef]
- Georgianna, D.R.; Hannon, M.J.; Marcuschi, M.; Wu, S.; Botsch, K.; Lewis, A.J.; Hyun, J.; Mendez, M.; Mayfield, S.P. Production of recombinant enzymes in the marine alga Dunaliella tertiolecta. Algal Res. 2013, 2, 2–9. [Google Scholar] [CrossRef]
- Wang, K.; Cui, Y.; Wang, Y.; Gao, Z.; Liu, T.; Meng, C.; Qin, S. Chloroplast genetic engineering of a unicellular green alga Haematococcus pluvialis with expression of an antimicrobial peptide. Mar. Biotechnol. 2020, 22, 572–580. [Google Scholar] [CrossRef] [PubMed]
- Mardanova, E.S.; Blokhina, E.A.; Tsybalova, L.M.; Peyret, H.; Lomonossoff, G.P.; Ravin, N.V. Efficient transient expression of recombinant proteins in plants by the novel pEff vector based on the genome of Potato Virus X. Front. Plant Sci. 2017, 28, 247. [Google Scholar] [CrossRef] [PubMed]
- Vidyasagar, P.; Sridevi, V.N.; Rajan, S.; Praveen, A.; Srikanth, A.; Abhinay, G.; Siva Kumar, V.; Verma, R.R.; Rajendra, L. Generation and characterization of neutralizing monoclonal antibodies against baculo-expressed HPV 16 VLPs. Eur. J. Microbiol. Immunol. 2014, 4, 56–64. [Google Scholar] [CrossRef] [Green Version]
- Van Doremalen, N.; Lambe, T.; Spencer, A.; Belij-Rammerstorfer, S.; Purushotham, J.N.; Port, J.R.; Avanzato, V.A.; Bushmaker, T.; Flaxman, A.; Ulaszewska, M.; et al. ChAdOx1 nCoV-19 vaccine prevents SARS-CoV-2 pneumonia in rhesus macaques. Nature 2020, 586, 578–582. [Google Scholar] [CrossRef]
- D’Adamo, S.; Kormelink, R.; Martens, D.; Barbosa, M.J.; Wijffels, R.H. Prospects for viruses infecting eukaryotic microalgae in biotechnology. Biotechnol. Adv. 2022, 54, 107790. [Google Scholar] [CrossRef]
- Ryabov, E.V.; Robinson, D.J.; Taliansky, M.E. A plant virus-encoded protein facilitates long-distance movement of heterologous viral RNA. Proc. Natl. Acad. Sci. USA 1999, 96, 1212–1217. [Google Scholar] [CrossRef] [Green Version]
- McCormick, A.A.; Reinl, S.J.; Cameron, T.I.; Vojdani, F.; Fronefield, M.; Levy, R.; Tuse, D. Individualized human scFv vaccines produced in plants: Humoral anti-idiotype responses in vaccinated mice confirm relevance to the tumor Ig. J. Immunol. Methods 2003, 278, 5–104. [Google Scholar] [CrossRef]
- Kumagai, M.H.; Donson, J.; della-Cioppa, G.; Grill, L.K. Rapid, high-level expression of glycosylated rice alpha-amylase in transfected plants by an RNA viral vector. Gene 2000, 245, 169–174. [Google Scholar] [CrossRef]
- Gleba, Y.; Klimyuk, V.; Marillonnet, S. Magnifection-a new platform for expressing recombinant vaccines in plants. Vaccine 2005, 23, 2042–2048. [Google Scholar] [CrossRef]
- Salazar-González, J.A.; Rosales-Mendoza, S.; Bañuelos-Hernández, B. Viral vector-based expression strategies. In Genetically Engineered Plants as a Source of Vaccines against Wide Spread Diseases; Rosales-Mendoza, S., Ed.; Springer: New York, NY, USA, 2014; pp. 43–60. [Google Scholar] [CrossRef]
- Naseri, Z.; Ghaffar, K.; Seyed Javad, D.; Hamideh, O. Virus-based vectors: A new approach for the production of recombinant proteins. J. Appl. Biotechnol. Rep. 2019, 6, 6–14. [Google Scholar] [CrossRef] [Green Version]
- Hanley-Bowdoin, L.; Bejarano, E.R.; Robertson, D.; Mansoor, S. Geminiviruses: Masters at redirecting and reprogramming plant processes. Nat. Rev. Microbiol. 2013, 11, 777–788. [Google Scholar] [CrossRef]
- Rojas, M.R.; Macedo, M.A.; Maliano, M.R.; Soto-Aguilar, M.; Souza, J.O.; Briddon, R.W.; Kenyon, L.; Rivera Bustamante, R.F.; Zerbini, F.M.; Adkins, S.; et al. World management of geminiviruses. Annu. Rev. Phytopathol. 2018, 56, 637–677. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Phoolcharoen, W.; Lai, H.; Piensook, K.; Cardineau, G.; Zeitlin, L.; Whaley, K.J.; Arntzen, C.J.; Mason, H.S.; Chen, Q. High-level rapid production of full-size monoclonal antibodies in plants by a single-vector DNA replicon system. Biotechnol. Bioeng. 2010, 106, 9–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phakham, T.; Bulaon, C.J.I.; Khorattanakulchai, N.; Shanmugaraj, B.; Buranapraditkun, S.; Boonkrai, C.; Sooksai, S.; Hirankarn, N.; Abe, Y.; Strasser, R.; et al. Functional characterization of pembrolizumab produced in Nicotiana benthamiana using a rapid transient expression system. Front. Plant Sci. 2021, 12, 736299. [Google Scholar] [CrossRef] [PubMed]
- Moon, K.B.; Jeon, J.H.; Choi, H.; Park, J.S.; Park, S.J.; Lee, H.J.; Park, J.M.; Cho, H.S.; Moon, J.S.; Oh, H.; et al. Construction of SARS-CoV-2 virus-like particles in plant. Sci. Rep. 2022, 12, 1005. [Google Scholar] [CrossRef] [PubMed]
- Berndt, A.J.; Smalley, T.N.; Ren, B.; Simkovsky, R.; Badary, A.; Sproles, A.E.; Fields, F.J.; Torres-Tiji, Y.; Heredia, V.; Mayfield, S.P. Recombinant production of a functional SARS-CoV-2 spike receptor binding domain in the green algae Chlamydomonas reinhardtii. PLoS ONE 2021, 16, e0257089. [Google Scholar] [CrossRef]
- Gimpel, J.A.; Hyun, J.S.; Schoepp, N.G.; Mayfield, S.P. Production of recombinant proteins in microalgae at pilot greenhouse scale. Biotechnol. Bioeng. 2015, 112, 339–345. [Google Scholar] [CrossRef]
- Zhao, Z.; Anselmo, A.C.; Mitragotri, S. Viral vector-based gene therapies in the clinic. Bioeng. Transl. Med. 2021, 7, e10258. [Google Scholar] [CrossRef]
- Short, S.M.; Staniewski, M.A.; Chaban, Y.V.; Long, A.M.; Wang, D. Diversity of viruses infecting eukaryotic algae. Curr. Issues Mol. Biol. 2020, 39, 29–62. [Google Scholar] [CrossRef] [Green Version]
- Coy, S.R.; Gann, E.R.; Pound, H.L.; Short, S.M.; Wilhelm, S.W. Viruses of eukaryotic algae: Diversity, methods for detection, and future directions. Viruses 2018, 10, 487. [Google Scholar] [CrossRef] [Green Version]
- Sandaa, R.A.; Saltvedt, M.R.; Dahle, H.; Wang, H.; Våge, S.; Blanc-Mathieu, R.; Steen, I.H.; Grimsley, N.; Edvardsen, B.; Ogata, H.; et al. Adaptive evolution of viruses infecting marine microalgae (haptophytes), from acute infections to stable coexistence. Biol. Rev. Camb. Philos. Soc. 2022, 97, 179–194. [Google Scholar] [CrossRef] [PubMed]
- Kadono, T.; Miyagawa-Yamaguchi, A.; Kira, N.; Tomaru, Y.; Okami, T.; Yoshimatsu, T.; Hou, L.; Ohama, T.; Fukunaga, K.; Okauchi, M.; et al. Characterization of marine diatom-infecting virus promoters in the model diatom Phaeodactylum tricornutum. Sci. Rep. 2015, 5, 18708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanc, G.; Duncan, G.; Agarkova, I.; Borodovsky, M.; Gurnon, J.; Kuo, A.; Lindquist, E.; Lucas, S.; Pangilinan, J.; Polle, J.; et al. The Chlorella variabilis NC64A genome reveals adaptation to photosymbiosis, coevolution with viruses, and cryptic sex. Plant Cell. 2010, 22, 2943–2955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeanniard, A.; Dunigan, D.D.; Gurnon, J.R.; Agarkova, I.V.; Kang, M.; Vitek, J.; Duncan, G.; McClung, O.W.; Larsen, M.; Claverie, J.M.; et al. Towards defining the chloroviruses: A genomic journey through a genus of large DNA viruses. BMC Genom. 2013, 14, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.S.; Zheng, Y.; Labavitch, J.M.; VanderGheynst, J.S. Virus infection of Chlorella variabilis and enzymatic saccharification of algal biomass for bioethanol production. Bioresour. Technol. 2013, 137, 326–331. [Google Scholar] [CrossRef]
- Agarkova, I.V.; Lane, L.C.; Dunigan, D.D.; Quispe, C.F.; Duncan, G.A.; Milrot, E.; Minsky, A.; Esmael, A.; Ghosh, J.S.; Van Etten, J.L. Identification of a Chlorovirus PBCV-1 protein involved in degrading the host cell wall during virus infection. Viruses 2021, 13, 782. [Google Scholar] [CrossRef]
- Quispe, C.F.; Esmael, A.; Sonderman, O.; McQuinn, M.; Agarkova, I.; Battah, M.; Duncan, G.A.; Dunigan, D.D.; Smith, T.; De Castro, C.; et al. Characterization of a new chlorovirus type with permissive and non-permissive features on phylogenetically related algal strains. Virology 2017, 500, 103–113. [Google Scholar] [CrossRef]
- Fitzgerald, L.A.; Graves, M.V.; Li, X.; Hartigan, J.; Pfitzner, A.J.; Hoffart, E.; Van Etten, J.L. Sequence and annotation of the 288-kb ATCV-1 virus that infects an endosymbiotic chlorella strain of the heliozoon Acanthocystis turfacea. Virology 2007, 362, 350–361. [Google Scholar] [CrossRef] [Green Version]
- Rahalkar, H.; Sheppard, A.; Lopez-Morales, C.A.; Lobo, L.; Salek, S. Challenges faced by the biopharmaceutical industry in the development and marketing authorization of biosimilar medicines in BRICS-TM countries: An exploratory study. Pharmaceut. Med. 2021, 35, 235–251. [Google Scholar] [CrossRef]


{kind=link}
| Method | Advantage | Disadvantage | Integration Site | Transformation Efficiency | Microalgae Species | Reference |
|---|---|---|---|---|---|---|
| Glass beads | Simple, controlability, high cell-survival rate, affordable, and minimal physical damage to cells | Cell wall removal and low transformation efficiency | Nucleus | ~103 µg DNA−1 | Chlamydomonas reinhardtii | [18] |
| NR | Dunaliella salina | [36] | ||||
| NR | Platymonas subcordiformis | [37] | ||||
| Electroporation | Simple, affordable equipment, and high transformation efficiency | Transformation frequency affected by higher pulse strength and length, medium composition, temperature and membrane characteristics | Nucleus | 6 × 103 per µg of DNA | Chlamydomonas reinhardtii | [38] |
| 2.5 × 104 per µg of DNA | [39] | |||||
| 1.1 × 107 per µg of DNA | Nannochloropsis limnetica | [40] | ||||
| NR | Chlamydomonas reinhardtii | [20] | ||||
| NR | Dunaliella salina | [41] | ||||
| NR | Scenedesmus obliquus | [42] | ||||
| NR | Monoraphidium neglectum | [43] | ||||
| NR | Chlorella pyrenoidosa | [44] | ||||
| NR | Nannochloropsis oculata | [45] | ||||
| Agrobacterium-mediated | Transformation of large DNA fragments, simple, stable, and efficient | Variation of transformation efficiencies and transformation frequency affected by physical and chemical factors, silenced transformants, lower number of multiple insertions | Nucleus | 311–355 × 10−6 | Chlamydomonas reinhardtii | [23] |
| NR | Haematococcus pluvialis | [22] | ||||
| 41.0 ± 4 CFU per 106 cells | Dunaliella bardawil | [46,47] | ||||
| Biobalistic | Most effective method for the transformation of chloroplasts/nuclear, multiple copies insertion. More DNA integration and copy number | Cost effective, required specialized equipment, and recovery low | Nuclear/Chloroplast Genome | ~2.5 × 10−5 DNA | Volvox carteri | [48] |
| 1.9 × 10−6 to 4.2 × 10−6 per µg of DNA 108 | Chlamydomonas reinhardtii | [49,50] | ||||
| NR | [51] | |||||
| NR | Phaeodactylum tricornutum | [52] | ||||
| NR | Cyclotella cryptica and Navicula saprophila | [53] | ||||
| NR | Cylindrotheca fusiformis | [54] | ||||
| Silicon carbide whiskers | Similar protocol | Low transformation frequency | Nucleus | NR | Chlamydomonas reinhardtii | [55,56] |
| Lithium acetate/polyethylene | Simple operation, low cost, less damage to the host cells and high transformation efficiency | Growth rate transformation temperature and plasmid concentration | Nucleus | 113 colonies μg−1 DNA | Dunaliella salina | [57] |
| Microalgae Strain | Transformation Method | Integration Site | Protein Expressed | Yields Obtained | Application | Reference |
|---|---|---|---|---|---|---|
| Dunaliella salina | Lithium acetate/PEG | Nucleus | SKTI | 0.68% TSP | Antivirus and anticancer | [57] |
| Agrobacterium-mediated transformation | H5HA | 225 µg TSP/2g | Avian influenza | [127] | ||
| Electroporation | HBsAg | 3.11 ± 0.50 ng/mg | Hepatitis B | [41] | ||
| Glass beads | VP28 | 3.04 ± 0.26 ng/mg and 78 µg/100 mL culture | White spot syndrome in crayfish | [128] | ||
| Biolistics | Chloroplast | sTRAIL | 0.67% TSP | Tumor cells and virus-infected cells | [129] | |
| Chlamydomonas reinhardtii | Agrobacterium-mediated transformation | Nucleus | HBcAgII | 0.05% TSP | Hypertension | [130] |
| IFN-α2a | NA | Immunity | [131] | |||
| RBD | 1.61 µg/g FWB | COVID-19 | [35] | |||
| bFGF | 1.025 ng/g FWB | Growth factor | ||||
| Glass beads | Chloroplast | HPV16 E7 mutated form r E7GGG-His6, E7GGG and E7GGG-FLAG | E7GGG-His6 (0.02%), E7GGG (0.1%) and E7GGG-FLAG (0.12%) TSP | Cancer | [117] | |
| WSSV VP28 | NA | White spot disease in shrimp | [132] | |||
| hGH | 0.5 mg hGH/L | Growth Hormone | [32] | |||
| dsRNA | NA | Yellow head virus infection in shrimp | [133] | |||
| Biolistics | ctxB-pfs25 | 0.09% TSP and 20 µg/mL | Malaria | [51] | ||
| pfs25 and pfs28 | Pfs25 (0.5%) and Pfs28 (0.2%) TSP | [134] | ||||
| c.r.pfs48/45 | NR | [75] | ||||
| Glass beads | Nucleus | AMA1/MSP1-GBSS | 0.2 to 1.0 mg of protein/mg | [120] | ||
| hVEGF-165, hPDGF-B, and hSDF-1 | 0.06% TSP, 0.003% TSP, 0.0006% TSP | Tissue hypoxia, wound healing | [135] | |||
| P24, CpP24, CrP24, P24w | 0.25% TSP | AISD | [118] | |||
| hEGF | 0.2%–0.25% TSP (40 mg/L) | hEGF deficiency | [136] | |||
| Endolysin (Cpl-1 and Pal) | ~1.3 mg/g ADW | Streptococcus pneumoniae infection | [137] | |||
| ALFPm3 | 0.35% TSP | Anti-bacteria, anticancer, and antiviral activity | [138] | |||
| IF | NA | Autoimmune disease pernicious anemia | [139] | |||
| Biolistics | Chloroplast | αCD22 | 0.7% TSP | Cancer | [124] | |
| 83K7C | 100 mg/1 g of DAB | Anthrax | [140] | |||
| HSV8 scfv | 0.5% TSP | Herpes simplex virus | [141] | |||
| HSV8-lsc | >1% TSP | Herpes simplex virus | [142] | |||
| M-SAA | 0.25% TSP | Protection against intestinal bacterial and viral infections in newborns | [143] | |||
| apcA and apcB | 2–3% TSP | Inhibit the S-180 carcinoma in mice | [144] | |||
| hMT-2 | NA | UV-B effects | [145] | |||
| CTB:p210 | 60 µg/g of FWB | Atherosclerosis | [146] | |||
| Ara h 1 and Ara h 2 | NA | Peanut allergy | [147] | |||
| Bet v 1.0101 | 0.01 and 0.04% TSP | Allergy | [148] | |||
| IL-2 and PfCelTOS | 1.5% TSP | Malaria | [121] | |||
| IFN-β1 | NA | Multiple sclerosis | [149] | |||
| VEGF | 0.1% TSP | Depression and pulmonary arteries | [149] | |||
| HMGB1 | 1% TSP | Response of the brain to neural injury and wound healing | [149] | |||
| CelK1 | 0.003% TSP | Bioethanol and biogas production | [150] | |||
| Biolistics | Nucleus | huBuChE | 0.4% TSP | Pesticide poisonings | [151] | |
| Electroporation | Nucleus | Mytichitin-A | 0.28% TSP | Growth inhibition of fungi, viruses, parasites, and bacteria | [152] | |
| ToAMP4 | 0.32% TSP | Antimicrobial | [153] | |||
| hLF | 1.82% TSP | Antibacterial | [154] | |||
| Schizochytrium sp | Agrobacterium-mediated transformation | Nucleus | HER-2, MUC1, MAM-A, and WT1 | 637 µg/g FWB | Breast cancer | [155] |
| ZK1, ZK2, ZK3, and LTB | 365 µg/g FWB | Zika disease | [115] | |||
| LTB:RAGE | 380 μg/g FWB | Alzheimer disease | [156] | |||
| GP1 and LTB | 1.25 mg/g FWB (6 mg/L of culture) | Ebola | [79] | |||
| Schizochytrium sp. ATCC 20888 | Biobalistic | Nucleus | HA | 5–20 mg/l | Influenza | [78] |
| Chlorella vulgaris | Agrobacterium-mediated transformation | Nucleus | RBD | 1.14 µg/g FWB | COVID-19 | [35] |
| bFGF | 1.61 ng/g FWB | Growth factor | ||||
| Chlorella sp | Electroporation | Nucleus | Scygonadin and hepcidin | NA | Antibacterial | [157] |
| Chlorella sorokiniana ATCC-22521) or Chlorella vulgaris C-27 | PEG | Nucleus | hGH | 200–600 ng/mL | Cell regeneration/hGH deficiency | [84] |
| Chlorella ellipsoidea | Biobalistic | Chloroplast | fGH | 420 µg fGH protein/L | Growth hormone | [123] |
| Dunaliella tertiolecta and C. reinhardtii | Biobalistic | Plastids | Xylanase, α-galactosidas, Phytase, phosphate anhydrolase, and β-mannanase | NA | Animal feeds and biofuel production | [158] |
| Haematococcus pluvialis | Biobalistic | Chloroplast | Piscidi-4 | NA | Antimicrobial | [159] |
| Microalgae Host | Type of Transformation | Name | Viral Elements | Protein Expressed | References |
|---|---|---|---|---|---|
| Schizochytrium sp. | Transient nuclear/Inducible expression | Algevir | Cauliflower mosaic virus: 35S promoter 35S terminator Ageratum enation virus: Replication protein “Rep” Origin of replication “Ori” | The GP1 from Zaire ebolavirus and LTB RAGE (23–54 amino acids) The ZK1, ZK2, ZK3 from the E protein from Zika virus fused to LTB The multiepitope protein BCB comprised epitopes from HER-2, MUC1, WT1, MAM-A fused to LTB | [79,115,155,156] |
| Chlamydomonas reinhardtii | Transient nuclear | pBYR2e | Cauliflower mosaic virus: 35S promoter Tomato bushy stunt virus: RNA silencing suppressor P19 Bean Yellow Dwarf Virus: Short intergenic region SIR Long intergenic region LIR C1/C2 Replication protein and replication protein A Tobacco mosaic virus Ω: 5’untranslated region | The RBD from SARS-CoV-2 The bFGF | [35] |
| Chlorella vulgaris |
| Viral Genomic Element | Name | Viral Source | Size (bp) | Type of Expression | Transformation Method | Protein Expressed | Heterologous Host | Reference |
|---|---|---|---|---|---|---|---|---|
| Promoters | C1P1 | ClorDNAV | 502 | Stable | Biobalistic | eGFP | Pennales diatom Phaeodactylum tricornutum | [181] |
| Electroporation | Sh ble | Chlamydomonas reinhardtii | ||||||
| ClP2 | 474 | Stable | Biobalistic | eGFP | Pennales diatom Phaeodactylum tricornutum | |||
| CdP1 | CdebDNAV | 477 | ||||||
| TnP1 | TnitDNAV | 424 | ||||||
| TnP2 | 424 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bolaños-Martínez, O.C.; Mahendran, G.; Rosales-Mendoza, S.; Vimolmangkang, S. Current Status and Perspective on the Use of Viral-Based Vectors in Eukaryotic Microalgae. Mar. Drugs 2022, 20, 434. https://doi.org/10.3390/md20070434
Bolaños-Martínez OC, Mahendran G, Rosales-Mendoza S, Vimolmangkang S. Current Status and Perspective on the Use of Viral-Based Vectors in Eukaryotic Microalgae. Marine Drugs. 2022; 20(7):434. https://doi.org/10.3390/md20070434
Chicago/Turabian StyleBolaños-Martínez, Omayra C., Ganesan Mahendran, Sergio Rosales-Mendoza, and Sornkanok Vimolmangkang. 2022. "Current Status and Perspective on the Use of Viral-Based Vectors in Eukaryotic Microalgae" Marine Drugs 20, no. 7: 434. https://doi.org/10.3390/md20070434
APA StyleBolaños-Martínez, O. C., Mahendran, G., Rosales-Mendoza, S., & Vimolmangkang, S. (2022). Current Status and Perspective on the Use of Viral-Based Vectors in Eukaryotic Microalgae. Marine Drugs, 20(7), 434. https://doi.org/10.3390/md20070434