
Exploring the Mangrove Fruit: From the Phytochemicals to Functional Food Development and the Current Progress in the Middle East

 and
and
Abstract
1. Introduction
2. Nutrition Composition and Bioactivity of Mangrove Fruit Extract


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Solvent | Antimicrobial | Ref |
|---|---|---|---|
| Avicennia marina | Ethanol | Aspergillus fumigatus | [58] |
| Candida albicans | |||
| A. officinalis | Methanol | Escherichia coli | [47] |
| Enterobacter aerogenes | |||
| Klebsiella pneumoniae | |||
| Pseudomonas aeruginosa | |||
| Bacillus subtilis | |||
| Lactobacillus delbrueckii | |||
| Staphylococcus aureus | |||
| Streptococcus pyogenes | |||
| B. gymnorrhiza | Methanol | E. coli | [51] |
| P. aeruginosa | |||
| K. pneumoniae | |||
| S. aureus | |||
| Salmonella enteritidis | |||
| Sarcina lutea | |||
| Proteus mirabilis | |||
| Bacillus cereus | |||
| C. albicans | |||
| R. mangle | Ethanol | Enterococcus faecalis | [52] |
| Bacillus thuringiensis | |||
| Bacillus cereus | |||
| Streptococcus lactis | |||
| S. aureus | |||
| S. apetala | Methanol | E. coli | [40] |
| E. faecalis | |||
| Pseudomonas sp. | |||
| Shigella flexneri | |||
| Staphylococcus epidermidis | |||
| S. caseolaris | Ethyl acetate | E. coli | [48] |
| C. albicans | |||
| Ethanol | E. coli | ||
| S. aureus | |||
| C. albicans | |||
| Methanol | S. aureus | [49,62] | |
| E. coli | |||
| C. albicans | |||
| P. aeruginosa | |||
| Acenobacter baumannii | |||
| Methanol:ethanol | E. coli | [63] | |
| Klebsiella sp. | |||
| Shigella boydii | |||
| S. sonnei | |||
| S. aureus | |||
| X. mekongensis | Methanol:ethanol | S. aureus | [63] |
3. Secondary Metabolites Isolated from Mangrove Fruit
4. The Secondary Metabolites from the Seed of Mangrove
5. The Endophytic Fungus in the Mangrove Fruit as a Source of Secondary Metabolites
6. The Mangrove Fruit in the Intermediary Stage of Processed Food
7. The Prospect of Mangrove Fruit in the Functional Food Development
8. The Possible Strategic Development for Mangrove Fruit as Functional Food Material
9. Current Studies of Mangrove Fruit in Middle East
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Chiwona-Karltun, L.; Amuakwa-Mensah, F.; Wamala-Larsson, C.; Amuakwa-Mensah, S.; Abu Hatab, A.; Made, N.; Taremwa, N.K.; Melyoki, L.; Rutashobya, L.K.; Madonsela, T.; et al. COVID-19: From health crises to food security anxiety and policy implications. Ambio 2021, 50, 794–811. [Google Scholar] [CrossRef] [PubMed]
- Savary, S.; Akter, S.; Almekinders, C.; Harris, J.; Korsten, L.; Rötter, R.; Waddington, S.; Watson, D. Mapping disruption and resilience mechanisms in food systems. Food Secur. 2020, 12, 695–717. [Google Scholar] [CrossRef] [PubMed]
- Baer-Nawrocka, A.; Sadowski, A. Food security and food self-sufficiency around the world: A typology of countries. PLoS ONE 2019, 14, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Del Rio Osorio, L.L.; Flórez-López, E.; Grande-Tovar, C.D. The potential of selected agri-food loss and waste to contribute to a circular economy: Applications in the food, cosmetic and pharmaceutical industries. Molecules 2021, 26, 515. [Google Scholar] [CrossRef]
- da Cunha, J.A.; Rolim, P.M.; Damasceno, K.S.F.d.S.C.; de Sousa, F.C.; Nabas, R.C.; Seabra, L.M.A.J. From seed to flour: Sowing sustainability in the use of cantaloupe melon residue (Cucumis melo L. Var. Reticulatus). PLoS ONE 2020, 15, 1–18. [Google Scholar] [CrossRef]
- Mason-D’Croz, D.; Bogard, J.R.; Sulser, T.B.; Cenacchi, N.; Dunston, S.; Herrero, M.; Wiebe, K. Gaps between fruit and vegetable production, demand, and recommended consumption at global and national levels: An integrated modelling study. Lancet Planet Health 2019, 3, e318–e329. [Google Scholar] [CrossRef]
- Dreher, M.L. Whole fruits and fruit fiber emerging health effects. Nutrients 2018, 10, 1833. [Google Scholar] [CrossRef]
- Chakraborti, U.; Mitra, B.; Bhadra, K. Island Based Assemblage Pattern and Foraging Profile of Insect Flower Visitors on Aegialitis rotundifolia—A Near Threatened Mangrove Plant from Indian Sundarban. Neotrop. Entomol. 2022, 51, 32–42. [Google Scholar] [CrossRef]
- Jänes, H.; Macreadie, P.I.; Rizzari, J.; Ierodioconou, D.; Reeves, S.E.; Dwyer, P.G.; Carnell, P.E. The value of estuarine producers to fisheries: A case study of Richmond River Estuary. Ambio 2022, 51, 875–887. [Google Scholar] [CrossRef]
- He, W.; Nabangchang, O.; Erdman, K.; Vanko, A.C.A.; Poudel, P.; Giri, C.; Vincent, J.R. Inferring economic impacts from a program’s physical outcomes: An application to forest protection in Thailand. Environ. Resour. Econ. 2022, in press. [Google Scholar] [CrossRef]
- Bandaranayake, W.M. Traditional and medicinal uses of mangroves. Mangroves Salt Marshes 1998, 2, 133–148. [Google Scholar] [CrossRef]
- Kacem, H.A.; Bouroubi, Y.; Khomalli, Y.; Elyaagoubi, S.; Maanan, M.; Rhinane, H.; Maanan, M. The economic benefit of coastal blue carbon stocks in a Moroccan Lagoon ecosystem: A case study at Moulay Bousselham Lagoon. Wetlands 2022, 42, 1–15. [Google Scholar] [CrossRef]
- Romero-Uribe, H.M.; López-Portillo, J.; Reverchon, F.; Hernández, M.E. Effect of degradation of a black mangrove forest on seasonal greenhouse gas emissions. Environ. Sci. Pollut. Res. 2022, 29, 11951–11965. [Google Scholar] [CrossRef] [PubMed]
- Nazareth, D.R.; Gonsalves, M.-J. Influence of seasonal and environmental variables on the emission of methane from the mangrove sediments of Goa. Environ. Monit. Assess. 2022, 194, 1–17. [Google Scholar] [CrossRef]
- Hussain, S.A.; Badola, R. Valuing mangrove ecosystem services: Linking nutrient retention function of mangrove forests to enhanced agroecosystem production. Wetl. Ecol. Manag. 2008, 16, 441–450. [Google Scholar] [CrossRef]
- Zhu, D.H.; Song, Q.L.; Nie, F.H.; Wei, W.; Chen, M.M.; Zhang, M.; Lin, H.Y.; Kang, D.J.; Chen, Z.B.; Hay, A.G.; et al. Effects of environmental and spatial variables on bacteria in Zhanjiang mangrove sediments. Curr. Microbiol. 2022, 79, 1–11. [Google Scholar] [CrossRef]
- del Campo, J.T.F.; Olvera-Vargas, M.; Silla-Cortés, F.; Figueroa-Rangel, B.L.; Iñiguez-Dávalos, L.I. Composition and structure of vegetation and tide regulate the occurrence of Oryzomys couesi and Hodomys alleni in mangrove forests of Laguna de Cuyutlán, West-Central Mexico. Wetl. Ecol. Manag. 2022, 30, 67–82. [Google Scholar] [CrossRef]
- Yan, J.; Du, J.; Su, F.; Zhao, S.; Zhang, S.; Feng, P. Reclamation and ecological service value evaluation of coastal wetlands using multispectral satellite imagery. Wetlands 2022, 42, 1–15. [Google Scholar] [CrossRef]
- Thakur, N.K.; Singh, R.; Ojha, A. Dynamical study of harmful algal bloom in Sundarban mangrove wetland with spatial interaction and competing effects. Model Earth Syst. Environ. 2022, 8, 555–577. [Google Scholar] [CrossRef]
- Huang, Z.; Huang, Y.; Lai, Q.; Chen, X.; Dong, C.; Huang, X. Oceanobacter mangrovi sp. Nov., a Novel Poly-β-hydroxybutyrate accumulating bacterium isolated from mangrove sediment. Curr. Microbiol. 2022, 79, 1–6. [Google Scholar] [CrossRef]
- Wei, M.Y.; Li, H.; Zhong, Y.H.; Shen, Z.J.; Ma, D.N.; Gao, C.H.; Liu, Y.L.; Wang, W.H.; Zhang, J.Y.; You, Y.P.; et al. Transcriptomic analyses reveal the effect of nitric oxide on the lateral root development and growth of mangrove plant Kandelia obovata. Plant Soil. 2022, in press. [Google Scholar] [CrossRef]
- Devadatha, B.; Jones, E.B.G.; Pang, K.L.; Abdel-Wahab, M.A.; Hyde, K.D.; Sakayaroj, J.; Bahkali, A.H.; Calabon, M.S.; Sarma, V.V.; Sutreong, S.; et al. Occurrence and geographical distribution of mangrove fungi. Fungal Divers. 2021, 106, 137–227. [Google Scholar] [CrossRef]
- Handayani, D.P.; Isnansetyo, A.; Istiqomah, I.; Jumina, J. New report: Genome mining untaps the antibiotics biosynthetic gene cluster of Pseudoalteromonas xiamenensis STKMTI.2 from a mangrove soil sediment. Mar. Biotechnol. 2022, in press. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.J.; Wu, M.-D.; Khamthong, N.; Tseng, M. Polar metabolites from the Actinobacterium Isoptericola chiayiensis isolated from mangrove soil. Chem. Nat. Compd. 2021, 57, 1134–1136. [Google Scholar] [CrossRef]
- Biswas, K.; Bhattarcharya, D.; Saha, M.; Mukherjee, J.; Karmakar, S. Evaluation of antimicrobial activity of the extract of Streptomyces euryhalinus isolated from the Indian Sundarbans. Arch. Microbiol. 2022, 204, 1–9. [Google Scholar] [CrossRef]
- Lee, N.L.Y.; Huang, D.; Quek, Z.B.R.; Lee, J.N.; Wainwright, B.J. Distinct fungal communities associated with different organs of the mangrove Sonneratia alba in the Malay Peninsula. IMA Fungus 2020, 11, 1–9. [Google Scholar] [CrossRef]
- Mesquita, L.M.d.S.; Caria, C.R.e.P.; Santos, P.S.; Ruy, C.C.; Da Silva Lima, N.; Moreira, D.K.T.; da Rocha, C.Q.; Murador, D.C.; de Rosso, V.V.; Gambero, A.; et al. Modulatory effect of polyphenolic compounds from the mangrove tree rhizophora mangle L. on non-alcoholic fatty liver disease and insulin resistance in high-fat diet obese mice. Molecules 2018, 23, 2114. [Google Scholar] [CrossRef]
- Dey, D.; Quispe, C.; Hossain, R.; Jain, D.; Ahmed Khan, R.; Janmeda, P.; Islam, M.T.; Suleria, A.R.H.; Martorell, M.; Daştan, S.D.; et al. Ethnomedicinal use, phytochemistry, and pharmacology of Xylocarpus granatum J. Koenig. Evid.-Based Complement. Altern. Med. 2021, 2021, 8922196. [Google Scholar] [CrossRef] [PubMed]
- Das, S.K.; Samantaray, D.; Mahapatra, A.; Pal, N.; Munda, R.; Thatoi, H. Pharmacological activities of leaf and bark extracts of a medicinal mangrove plant Avicennia officinalis L. Clin. Phytosci. 2018, 4, 1–10. [Google Scholar] [CrossRef]
- Mazlan, N.W.; Clements, C.; Edrada-Ebel, R.A. Targeted isolation of anti-trypanosomal naphthofuran-quinone compounds from the mangrove plant Avicennia lanata. Mar. Drugs 2020, 18, 661. [Google Scholar] [CrossRef]
- Zhou, H.C.; Tam, N.F.Y.; Lin, Y.M.; Ding, Z.H.; Chai, W.M.; Wei, S.D. Relationships between degree of polymerization and antioxidant activities: A study on proanthocyanidins from the leaves of a medicinal mangrove plant Ceriops tagal. PLoS ONE 2014, 9, e107606. [Google Scholar] [CrossRef] [PubMed]
- Bibi, S.N.; Fawzi, M.M.; Gokhan, Z.; Rajesh, J.; Nadeem, N.; Kannan, R.R.R.; Albuquerque, R.D.D.G.; Pandian, S.K. Ethnopharmacology, phytochemistry, and global distribution of mangroves-a comprehensive review. Mar. Drugs 2019, 17, 231. [Google Scholar] [CrossRef] [PubMed]
- Nawar, M.K.; Basyuni, M.; Hanum, C.; Siregar, E.S. Bioprospecting opportunities of mangrove fruits for the coastal community in lubuk kertang and pulau sembilan, north sumatra, indonesia. Asian J. Plant Sci. 2022, 21, 145–153. [Google Scholar]
- Arshad, M.; Eid, E.M.; Hasan, M. Mangrove health along the hyper-arid southern Red Sea coast of Saudi Arabia. Environ. Monit. Assess. 2020, 192, 189. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; El-Askary, H.; Qurban, M.A.; Li, J.; ManiKandan, K.P.; Piechota, T. Using multi-indices approach to quantify mangrove changes over the Western Arabian Gulf along Saudi Arabia coast. Ecol. Indic. 2019, 102, 734–745. [Google Scholar] [CrossRef]
- Almahasheer, H. Spatial coverage of mangrove communities in the Arabian Gulf. Environ. Monit. Assess. 2018, 190, 85. [Google Scholar] [CrossRef]
- Wonggo, D.; Berhimpon, S.; Kurnia, D.; Dotulong, V. Antioxidant activities of mangrove fruit (Sonneratia alba) taken from Wori Village, North Sulawesi, Indonesia. Int. J. ChemTech Res. 2017, 10, 284–290. [Google Scholar]
- Riyadi, P.H.; Tanod, W.A.; Dewanto, D.K.; Herawati, V.E.; Susanto, E.; Aisiah, S. Chemical profiles and antioxidant properties of Bruguiera gymnorrhiza fruit extracts from central sulawesi, indonesia. Food Res. 2021, 5, 37–47. [Google Scholar] [CrossRef]
- Abu Bakar, F.I.; Abu Bakar, M.F.; Hassan, S.H.A.; Sanusi, S.B.; Kormin, F.; Sabran, S.F.; Fuzi, F.Z.M. Comparison of phytochemicals, antioxidant and anti-cholinesterase activity of unripe and ripe fruit of Sonneratia caseolaris. Food Res. 2020, 4, 507–514. [Google Scholar] [CrossRef]
- Hossain, S.J.; Basar, M.H.; Rokeya, B.; Arif, K.M.T.; Sultana, M.S.; Rahman, M.H. Evaluation of antioxidant, antidiabetic and antibacterial activities of the fruit of Sonneratia apetala (Buch.-Ham.). Orient. Pharm. Exp. Med. 2013, 13, 95–102. [Google Scholar] [CrossRef]
- Basyuni, M.; Yusraini, E.; Susilowati, A.; Hayati, R.; Siregar, E.S.; Desrita; Susetya, I.E.; Kajita, T. Bioprospecting of selected mangrove fruits based-nutritional, antioxidant, and element properties to support functional food materials for Pulau Sembilan coastal communities, Indonesia. Int. J. Adv. Sci. Eng. Inf. Technol. 2021, 11, 1661–1667. [Google Scholar] [CrossRef]
- Ray, R.; Banerjee, A.; Mullick, J.; Jana, T.K. Nutritional composition of some selected wild mangrove fruits of Sundarbans. Indian J. Geo-Mar. Sci. 2015, 44, 1059–1066. [Google Scholar]
- Sudirman, S.; Nurjanah; Jacoeb, A.M. Proximate compositions, bioactive compounds and antioxidant activity from large-leafed mangrove (Bruguiera gymnorrhiza) fruit. Int. Food Res. J. 2014, 21, 2387–2391. [Google Scholar]
- Mahmud, S.; Paul, G.K.; Afroze, M.; Islam, S.; Gupt, S.B.R.; Razu, M.H.; Biswas, S.; Zaman, S.; Uddin, M.S.; Khan, M.; et al. Efficacy of phytochemicals derived from Avicennia officinalis for the management of covid-19: A combined in silico and biochemical study. Molecules 2021, 26, 2210. [Google Scholar] [CrossRef] [PubMed]
- Cahyadi, J.; Satriani, G.I.; Gusman, E.; Weliyadi, E. Inhibiting vibrio harveyi infection in Penaeus monodon using enriched Artemia salina with mangrove fruit Sonneratia alba extract. AACL Bioflux 2020, 13, 1674–1681. [Google Scholar]
- Othman, L.; Sleiman, A.; Abdel-Massih, R.M. Antimicrobial activity of polyphenols and alkaloids in middle eastern plants. Front. Microbiol. 2019, 10, 911. [Google Scholar] [CrossRef] [PubMed]
- Sharief, M.N.; Srinivasulu, A.; Veni, P.S.; Rao, V.U.M. Quantification of phytochemicals and antibacterial activity of fruit extract of Avicennia officinalis. Asian J. Pharm. Clin. Res. 2014, 7, 127–130. [Google Scholar]
- Pagarra, H.; Hartati; Rachmawaty; Hala, Y.; Rahman, R.A. Phytochemical screening and antimicrobial activity from Sonneratia caseolaris fruit extract. Mater. Sci. Forum. 2019, 967, 28–33. [Google Scholar]
- Ahmad, I.; Ambarwati, N.S.S.; Lukman, A.; Masruhim, M.A.; Rijai, L.; Mun’Im, A. In vitro antimicrobial activity evaluation of mangrove fruit (Sonneratia caseolaris L.) extract. Pharmacogn. J. 2018, 10, 598–601. [Google Scholar] [CrossRef]
- Sadeer, N.B.; Rocchetti, G.; Senizza, B.; Montesano, D.; Zengin, G.; Uysal, A.; Jeewon, R.; Lucini, L.; Mahomoodally, M.F. Untargeted metabolomic profiling, multivariate analysis and biological evaluation of the true mangrove (Rhizophora mucronata lam.). Antioxidants 2019, 8, 489. [Google Scholar] [CrossRef]
- Sadeer, N.B.; Haddad, J.G.; Ezzat, M.O.; Desprès, P.; Abdallah, H.H.; Zengin, G.; Uysal, A.; El Kalamouni, C.; Gallo, M.; Montesano, D.; et al. Bruguiera gymnorhiza (L.) lam. at the forefront of pharma to confront zika virus and microbial infections—an in vitro and in silico perspective. Molecules 2021, 26, 5768. [Google Scholar] [CrossRef] [PubMed]
- Hicks, M.; Bailey, M.A.; Thiagarajan, T.R.; Troyer, T.L.; Huggins, L.G. Antibacterial and cytotoxic effects of red mangrove (Rhizophora mangle L.Rhizophoraceae) fruit extract. Eur. J. Sci. Res. 2011, 63, 439–446. [Google Scholar]
- Wetwitayaklung, P.; Limmatvapirat, C.; Phaechamud, T. Antioxidant and anticholinesterase activities in various parts of Sonneratia caseolaris (L.). Indian J. Pharm. Sci. 2013, 75, 649–656. [Google Scholar] [PubMed]
- Sulmartiwi, L.; Pujiastuti, D.Y.; Tjahjaningsih, W.; Jariyah. Potential of mangrove Avicennia rumphiana extract as an antioxidant agent using multilevel extraction. In Proceedings of the ASEAN-Fen International Fisheries Symposium, Batu, East Java, Indonesia, 7–9 November 2017. [Google Scholar]
- Deng, Y.; Liao, Y.; Li, J.; Yang, L.; Zhong, H.; Zhou, Q.; Qing, Z. Acaricidal activity against Panonychus citri and active ingredient of the mangrove plant Cerbera manghas. Nat. Prod. Commun. 2014, 9, 1265–1268. [Google Scholar] [CrossRef] [PubMed]
- Parthiban, A.; Sivasankar, R.; Sachithanandam, V.; Khan, S.A.; Jayshree, A.; Murugan, K.; Sridhar, R. An integrative review on bioactive compounds from Indian mangroves for future drug discovery. S. Afr. J. Bot. 2021, in press. [Google Scholar] [CrossRef]
- Masdar, H.; Hamidy, M.Y.; Darmawi; Trihardi, R.; Perwira, A.; Utari, D. Anti-atherosclerotic effects of Sonneratia Alba fruit extract in atherosclerotic-induced rats. Int. J. Appl. Pharm. 2020, 12, 41–43. [Google Scholar] [CrossRef]
- Okla, M.K.; Alatar, A.A.; Al-Amri, S.S.; Soufan, W.H.; Ahmad, A.; Abdel-Maksoud, M.A. Antibacterial and antifungal activity of the extracts of different parts of Avicennia marina (Forssk.) vierh. Plants 2021, 10, 252. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.K.; Tiwari, P.; Srivastava, S.P.; Srivastava, R.; Mishra, A.; Rahuja, N.; Pandeti, S.; Tamrakar, A.K.; Narender, T.; Srivastava, M.N.; et al. Antihyperglycaemic and antidyslipidemic activities in ethyl acetate fraction of fruits of marine mangrove Xylocarpus Moluccensis. Int. J. Pharm. Pharm. Sci. 2014, 6, 809–826. [Google Scholar]
- Liu, J.; Luo, D.; Wu, Y.; Gao, C.; Lin, G.; Chen, J.; Wu, X.; Zhang, Q.; Cai, J.; Su, Z. The protective effect of Sonneratia apetala fruit extract on acetaminophen-induced liver injury in mice. Evid.-Based Complement. Altern. Med. 2019, 2019, 6919834. [Google Scholar] [CrossRef]
- Di Meo, S.; Venditti, P. Evolution of the knowledge of free radicals and other oxidants. Oxid. Med. Cell Longev. 2020, 2020, 9829176. [Google Scholar] [CrossRef]
- Yompakdee, C.; Thunyaharn, S.; Phaechamud, T. Bactericidal activity of methanol extracts of crabapple mangrove tree (Sonneratia caseolaris Linn.) against multi-drug resistant pathogens. Indian J. Pharm. Sci. 2012, 74, 230–236. [Google Scholar] [PubMed]
- Hosen, M.Z.; Biswas, A.; Islam, M.R.; Hossain, S.J. Anti-bacterial, anti-diarrheal, and cytotoxic activities of edible fruits in the Sundarbans mangrove forest of Bangladesh. Prev. Nutr. Food Sci. 2021, 26, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Hamidy, M.Y.; Masdar, H.; Darmawi. Effect of mangrove (Rhizophora sp.) fruit extract on foam cell formation at the initiation stage of atherosclerosis. Biomed. Pharmacol. J. 2020, 13, 423–427. [Google Scholar] [CrossRef]
- Khatana, C.; Saini, N.K.; Chakrabarti, S.; Saini, V.; Sharma, A.; Saini, R.V.; Saini, A.K. Mechanistic insights into the oxidized low-density lipoprotein-induced atherosclerosis. Oxid. Med. Cell Longev. 2020, 2020, 5245308. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Song, D.; Wu, J.; Wang, J. Long non-coding RNAs link oxidized low-density lipoprotein with the inflammatory response of macrophages in atherogenesis. Front. Immunol. 2020, 11, 1–11. [Google Scholar] [CrossRef]
- Hao, W.; Friedman, A. The LDL-HDL profile determines the risk of atherosclerosis: A mathematical model. PLoS ONE 2014, 9, e90497. [Google Scholar] [CrossRef] [PubMed]
- Mulholland, D.A.; Parel, B.; Coombes, P.H. The chemistry of the Meliaceae and Ptaeroxylaceae of Southern and Eastern Africa and Madagascar. Curr. Org. Chem. 2000, 4, 1011–1054. [Google Scholar] [CrossRef]
- Cui, J.; Wu, J.; Deng, Z.; Proksch, P.; Lin, W. Xylocarpins A-I, limonoids from the Chinese mangrove plant Xylocarpus granatum. J. Nat. Prod. 2007, 70, 772–778. [Google Scholar] [CrossRef]
- Sahai, R.; Bhattacharjee, A.; Shukla, V.N.; Yadav, P.; Hasanain, M.; Sarkar, J.; Narender, T.; Mitra, K. Gedunin isolated from the mangrove plant Xylocarpus granatum exerts its anti-proliferative activity in ovarian cancer cells through G2/M-phase arrest and oxidative stress-mediated intrinsic apoptosis. Apoptosis 2020, 25, 481–499. [Google Scholar] [CrossRef]
- Wu, J.; Xiao, Z.; Song, Y.; Zhang, S.; Xiao, Q.; Ma, C.; Ding, H.; Li, Q. Complete assignments of 1H and 13C NMR data for two 3β,8β-epoxymexicanolides from the fruit of a Chinese mangrove Xylocarpus granatum. Magn. Reson. Chem. 2006, 44, 87–89. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, S.; Song, Y.; Xiao, Z.; Xiao, Q.; Li, Q. Two new mexicanolides from the fruit of a Chinese mangrove Xylocarpus granatum. Z. Nat. B, Chem. Sci. 2005, 60b, 1291–1294. [Google Scholar] [CrossRef]
- Cheng, F.; Zhou, Y.; Wu, J.; Zou, K. Xyloccensins X1 and X2, two new mexicanolides from the fruit of a Chinese mangrove Xylocarpus granatum. Z. Nat. B, Chem. Sci. 2005, 61b, 626–628. [Google Scholar]
- Zhou, Y.; Cheng, F.; Wu, J.; Zou, K. Polyhydroxylated phragmalins from the fruit of a Chinese Mangrove, Xylocarpus granatum. J. Nat. Prod. 2006, 69, 1083–1085. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Zhang, S.; Li, M.; Zhou, Y.; Xiao, Q. Xylogranatins A-D, new mexicanolides from the fruit of a Chinese mangrove Xylocarpus granatum. Chem. Pharm. Bull. 2006, 54, 1582–1585. [Google Scholar] [CrossRef]
- Wu, J.; Ding, H.; Li, M.; Zhang, S. Xylogranatin E, a new phragmalin with a rare oxygen bridge between C 1 and C29, from the fruit of a Chinese mangrove Xylocarpus granatum. Z. Nat. Sect. B J. Chem. Sci. 2007, 62b, 569–572. [Google Scholar]
- Zhou, Y.; Wu, J.; Zou, K. Xylogranatinin, a new pyrido[1, 2-a]pyrazine alkaloid from the fruit of a Chinese mangrove Xylocarpus granatum. Chem. Nat. Compd. 2007, 43, 426–428. [Google Scholar] [CrossRef]
- Cui, J.; Deng, Z.; Xu, M.; Proksch, P.; Li, Q.; Lin, W. Protolimonoids and limonoids from the chinese mangrove plant Xylocarpus granatum. Helv. Chim. Acta 2009, 92, 139–150. [Google Scholar] [CrossRef]
- Wu, J.; Li, M.; Xiao, Z.; Zhou, Y. Butyrospermol fatty acid esters from the fruit of a Chinese mangrove Xylocarpus granatum. Z. Nat. Sect. B J. Chem. Sci. 2006, 61, 1447–1449. [Google Scholar] [CrossRef]
- Yan, D.M.; Gao, C.H.; Yi, X.X.; Xie, W.P.; Xu, M.B.; Huang, R.M. Two new secondary metabolites from the fruits of mangrove Avicennia marina. Z. Nat. 2015, 70, 691–694. [Google Scholar] [CrossRef]
- Yi, X.X.; Chen, Y.; Xie, W.P.; Xu, M.B.; Chen, Y.N.; Gao, C.H.; Huang, R.M. Four new jacaranone analogs from the fruits of a Beibu Gulf mangrove Avicennia marina. Mar. Drugs 2014, 12, 2515–2525. [Google Scholar] [CrossRef]
- Gao, C.H.; Yi, X.X.; Xie, W.P.; Chen, Y.N.; Xu, M.B.; Su, Z.W.; Yu, L.; Huang, R.M. New antioxidative secondary metabolites from the fruits of a beibu gulf mangrove, Avicennia marina. Mar. Drugs 2014, 12, 4353–4360. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Ouyang, J.; Deng, Z.; Lin, W. Structure elucidation of an unprecedented alkaloid and a new limonoid from Xylocarpus granatum. Magn. Reson. Chem. 2008, 46, 894–897. [Google Scholar] [CrossRef] [PubMed]
- Pakhathirathien, C.; Karalai, C.; Ponglimanont, C.; Subhadhirasakul, S.; Chantrapromma, K. Dammarane triterpenes from the hypocotyls and fruits of Ceriops tagal. J. Nat. Prod. 2005, 68, 1787–1789. [Google Scholar] [CrossRef] [PubMed]
- Yi, X.; Jiang, S.; Qin, M.; Liu, K.; Cao, P.; Chen, S.; Deng, J.; Gao, C. Compounds from the fruits of mangrove Sonneratia apetala: Isolation, molecular docking and antiaging effects using a Caenorhabditis elegans model. Bioorg. Chem. 2020, 99, 103813. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Wu, Y.; Lv, T.; Wang, Y.; Fu, Y.; Sun, M.; Shi, Q.; Huo, C.; Wang, Q.; Gu, Y. A chemometric-assisted LC–MS/MS method for the simultaneous determination of 17 limonoids from different parts of Xylocarpus granatum fruit. Anal. Bioanal. Chem. 2017, 409, 4669–4679. [Google Scholar] [CrossRef] [PubMed]
- Narender, T.; Khaliq, T.; Shweta; Reddy, K.P.; Sharma, R.K. Occurrence, biosynthesis, biological activity and NMR spectroscopy of D and B, D ring seco-limonoid of Meliaceae family. Natural Product Commun. 2007, 2, 203–221. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, S.; Xiao, Q.; Li, Q.; Huang, J.; Xiao, Z.; Long, L. Xyloccensin M and N, Two New B, D-seco limonoids from Xylocarpus granatum. Z. Nat. Sect. B J. Chem. Sci. 2003, 58b, 1216–1219. [Google Scholar] [CrossRef]
- Alvi, K.A.; Crews, P.; Aalbersberf, B.; Prasa, R. Limonoids from the Fijian medicinal plant dabi (Xylocarpus). Tetrahedron 1991, 41, 8943–8948. [Google Scholar] [CrossRef]
- Li, J.; Li, M.Y.; Feng, G.; Xiao, Q.; Sinkkonen, J.; Satyanandamurty, T.; Wu, J. Limonoids from the seeds of a Godavari mangrove, Xylocarpus moluccensis. Phytochemistry 2010, 71, 1917–1924. [Google Scholar] [CrossRef]
- Wu, J.; Li, M.; Zhang, S.; Xiao, Q.; Li, Q. Two new limonoids with a 3-0-β-tigloyl group from the seeds of the Chinese mangrove Xylocarpus granatum. Z. Nat. Sect. B J. Chem. Sci. 2007, 62, 859–862. [Google Scholar] [CrossRef]
- Ren, J.L.; Zou, X.P.; Li, W.S.; Shen, L.; Wu, J. Limonoids containing a C1-O-C29 Moiety: Isolation, structural modification, and antiviral activity. Mar. Drugs. 2018, 16, 434. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.R.; Yin, B.W.; Zhang, M.L.; Shi, Q.W.; Huo, C.H.; Dong, M.; Cong, B.; Guo, D.; Kiyota, H.; Suzuki, N. Xylomexicanins A and B, new Δ14,15-Mexicanolides from seeds of the Chinese mangrove Xylocarpus granatum. Z. Nat. Sect. C J. Biosci. 2009, 64, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, M.Y.; Feng, G.; Zhang, J.; Karonen, M.; Sinkkonen, J.; Satyanandamurty, T.; Wu, J. Moluccensins R-Y, Limonoids from the Seeds of a Mangrove, Xylocarpus moluccensis. J. Nat. Prod. 2012, 75, 1277–1283. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Satyanandamurty, T.; Shen, L.; Wu, J. Krishnolides A–D: New 2-ketokhayanolides from the Krishna mangrove, Xylocarpus moluccensis. Mar. Drugs 2017, 15, 333. [Google Scholar] [CrossRef]
- Zhang, J.; Li, W.; Dai, Y.; Shen, L.; Wu, J. Twenty-nine new limonoids with skeletal diversity from the mangrove plant, Xylocarpus moluccensis. Mar. Drugs 2018, 16, 38. [Google Scholar] [CrossRef]
- Dai, Y.G.; Li, W.S.; Pedpradab, P.; Liu, J.J.; Wu, J.; Shen, L. Thaixylomolins O-R: Four new limonoids from the Trang mangrove, Xylocarpus moluccensis. RSC Adv. 2016, 6, 85978–85984. [Google Scholar] [CrossRef]
- Liu, R.X.; Liao, Q.; Shen, L.; Wu, J. Krishnagranatins A–I: New limonoids from the mangrove, Xylocarpus granatum, and NF-κB inhibitory activity. Fitoterapia 2018, 131, 96–104. [Google Scholar] [CrossRef]
- Dai, Y.G.; Wu, J.; Padmakumar, K.P.; Shen, L. Sundarbanxylogranins A–E, five new limonoids from the Sundarban Mangrove, Xylocarpus granatum. Fitoterapia 2017, 122, 85–89. [Google Scholar] [CrossRef]
- Yin, S.; Wang, X.-N.; Fan, C.-Q.; Lin, L.-P.; Ding, J.; Yue, J.-M. Limonoids from the seeds of the marine mangrove, Xylocarpus granatum. J. Nat. Prod. 2007, 70, 682–685. [Google Scholar] [CrossRef]
- Liao, M.; Pedpradab, P.; Wu, J. Thaixylogranins A–H: Eight new limonoids from the Thai mangrove, Xylocarpus granatum. Phytochem. Lett. 2017, 19, 126–131. [Google Scholar] [CrossRef]
- Li, W.; Shen, L.; Bruhn, T.; Pedpradab, P.; Wu, J.; Bringmann, G. Trangmolins A–F with an unprecedented structural plasticity of the rings A and B: New insight into limonoid biosynthesis. Chem. A Eur. J. 2016, 22, 11719–11727. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Jiang, Z.; Shen, L.; Pedpradab, P.; Bruhn, T.; Wu, J.; Bringmann, G. Antiviral limonoids including khayanolides from the trang mangrove plant Xylocarpus moluccensis. J. Nat. Prod. 2015, 78, 1570–1578. [Google Scholar] [CrossRef] [PubMed]
- Sarigaputi, C.; Sommit, D.; Teerawatananond, T.; Pudhom, K. Weakly anti-inflammatory limonoids from the seeds of Xylocarpus rumphii. J. Nat. Prod. 2014, 77, 2037–2043. [Google Scholar] [CrossRef] [PubMed]
- Li, M.Y.; Xiao, Q.; Satyanandamurty, T.; Wu, J. Limonoids with an oxygen bridge between C(1) and C(29) from the seeds of a Krishna Mangrove, Xylocarpus granatum. Chem. Biodivers. 2014, 11, 262–275. [Google Scholar] [CrossRef] [PubMed]
- Cai, R.; Wu, Y.; Chen, S.; Cui, H.; Liu, Z.; Li, C.; She, Z. Peniisocoumarins A-J: Isocoumarins from Penicillium commune QQF-3, an Endophytic Fungus of the Mangrove Plant Kandelia candel. J. Nat. Prod. 2018, 81, 1376–1383. [Google Scholar] [CrossRef]
- Liu, Y.; Wu, Y.; Zhai, R.; Liu, Z.; Huang, X.; She, Z. Altenusin derivatives from mangrove endophytic fungus: Alternaria sp. SK6YW3L. RSC Adv. 2016, 6, 72127–72132. [Google Scholar] [CrossRef]
- Yan, Z.; Huang, C.; Guo, H.; Zheng, S.; He, J.; Lin, J.; Long, Y. Isobenzofuranone monomer and dimer derivatives from the mangrove endophytic fungus Epicoccum nigrum SCNU-F0002 possess α-glucosidase inhibitory and antioxidant activity. Bioorg. Chem. 2020, 94, 103407. [Google Scholar] [CrossRef]
- Elsbaey, M.; Tanaka, C.; Miyamoto, T. Allantopyrone E, a rare α-pyrone metabolite from the mangrove derived fungus Aspergillus versicolor. Nat. Prod. Res. 2020, 36, 1–5. [Google Scholar] [CrossRef]
- Ju, Z.; Lin, X.; Lu, X.; Tu, Z.; Wang, J.; Kaliyaperumal, K.; Liu, J.; Tian, Y.; Xu, S.; Liu, Y. Botryoisocoumarin A, a new COX-2 inhibitor from the mangrove Kandelia candel endophytic fungus Botryosphaeria sp. KcF6. J. Antibiot. 2015, 68, 653–656. [Google Scholar] [CrossRef]
- Liu, H.; Yan, C.; Li, C.; You, T.; She, Z. Naphthoquinone derivatives with anti-inflammatory activity from mangrove-derived endophytic fungus Talaromyces sp. SK-S009. Molecules 2020, 25, 576. [Google Scholar] [CrossRef]
- Qiu, P.; Liu, Z.; Chen, Y.; Cai, R.; Chen, G.; She, Z. Secondary metabolites with α-Glucosidase inhibitory activity from the mangrove fungus mycosphaerella sp. SYSU-DZG01. Mar. Drugs 2019, 17, 483. [Google Scholar] [CrossRef] [PubMed]
- Jia, S.; Su, X.; Yan, W.; Wu, M.; Wu, Y.; Lu, J.; He, X.; Ding, X.; Xue, Y. Acorenone C: A New spiro-sesquiterpene from a mangrove-associated fungus, Pseudofusicoccum sp. J003. Front. Chem. 2021, 9, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.H.; Pan, J.H.; Chen, B.; Yu, M.; Huang, H.B.; Zhu, X.; Lu, Y.J.; She, Z.G.; Lin, Y.C. Three bianthraquinone derivatives from the mangrove endophytic fungus Alternaria sp. ZJ9-6B from the South China Sea. Mar. Drugs 2011, 9, 832–843. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.-K.; Seok, S.-J.; Kim, W.-G.; Yun, B.-S. Diaporthin and orthosporin from the fruiting body of Daldinia concentrica. Mycobiology 2006, 34, 38–40. [Google Scholar] [CrossRef] [PubMed]
- Larsen, T.O.; Breinholt, J. Dichlorodiaportin, diaportinol, and diaportinic acid: Three novel isocoumarins from Penicillium nalgiovense. J. Nat. Prod. 1999, 62, 1182–1184. [Google Scholar] [CrossRef]
- Naganuma, M.; Nishida, M.; Kuramochi, K.; Sugawara, F.; Yoshida, H.; Mizushina, Y. 1-Deoxyrubralactone, a novel specific inhibitor of families X and Y of eukaryotic DNA polymerases from a fungal strain derived from sea algae. Bioorganic Med. Chem. 2008, 16, 2939–2944. [Google Scholar] [CrossRef]
- Kimura, Y.; Yoshinari, T.; Koshino, H.; Fujioka, S.; Okada, K.; Shimada, A. Rubralactone, rubralides A, B and C, and rubramin produced by Penicillium rubrum. Biosci. Biotechnol. Biochem. 2007, 71, 1896–1901. [Google Scholar] [CrossRef]
- Koch, K.; Podlech, J.; Pfeiffer, E.; Metzler, M. Total synthesis of alternariol. J. Org. Chem. 2005, 70, 3275–3276. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Krohn, K.; Draeger, S.; Schulz, B. Bioactive isocoumarins isolated from the endophytic fungus Microdochium bolleyi. J. Nat. Prod. 2008, 71, 1078–1081. [Google Scholar] [CrossRef] [PubMed]
- Nurunnabi, T.R.; Sabrin, F.; Sharif, D.I.; Nahar, L.; Sohrab, M.H.; Sarker, S.D.; Rahman, S.M.M.; Billah, M.M. Antimicrobial activity of endophytic fungi isolated from the mangrove plant Sonneratia apetala (Buch.-Ham) from the Sundarbans mangrove forest. Adv. Tradit. Med. 2020, 20, 419–425. [Google Scholar] [CrossRef]
- Ghanate, A.S.; Annapure, U.S. Effect of physicochemical and rheological properties of flour from different local wheat varieties on the quality of varanphal: An Indian traditional product. J. Food Sci. Technol. 2019, 56, 3033–3042. [Google Scholar] [CrossRef] [PubMed]
- Khoza, M.; Kayitesi, E.; Dlamini, B.C. Physicochemical characteristics, microstructure and health promoting properties of green banana flour. Foods 2021, 10, 1894. [Google Scholar] [CrossRef] [PubMed]
- Pangestuti, R.; Getachew, A.T.; Siahaan, E.A.; Chun, B.S. Characteristics of functional materials recovered from Indonesian mangroves (Sonneratia alba and Rhizhophora mucronata) using subcritical water extraction. In Proceedings of the E3S Web of Conferences, the 3rd International Symposium on Marine and Fisheries Research, Yogyakarta, Indonesia, 8–9 July 2020; p. 030313. [Google Scholar]
- Fan, X.; Zhu, J.; Dong, W.; Sun, Y.; Lv, C.; Guo, B.; Xu, R. Comparison of pasting properties measured from the whole grain flour and extracted starch in barley (Hordeum vulgare L.). PLoS ONE 2019, 14, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Torres-Gallo, R.; Durán, R.; García-Camargo, J.; Morales, O.; Acevedo, D.; Tirado, D.F. Pasting and dough rheological properties of ackee (Blighia sapida) aril flour: A contribution to the search for wheat flour substitutes. Int. J. Food Sci. 2021, 2021, 5526912. [Google Scholar] [CrossRef] [PubMed]
- Adefegha, S.A.; Olasehinde, T.A.; Oboh, G. Pasting alters glycemic index, antioxidant activities, and starch-hydrolyzing enzyme inhibitory properties of whole wheat flour. Food Sci. Nutr. 2018, 6, 1591–1600. [Google Scholar] [CrossRef]
- Amin, M.N.G.; Pebruwantoro, D.; Pralebda, S.A.; Hasan, M.N.; Zakariya; Subekti, S.; Pramono, H.; Alamsjah, M.A. Functional and rheological properties of mixed flour from mangrove fruit of Bruguiera gymnorrhiza flour and wheat flour. Food Res. 2021, 5, 167–173. [Google Scholar] [CrossRef]
- Bala, M.; Handa, S.D.M.; Singh, R.K. Physicochemical, functional and rheological properties of grass pea (Lathyrus sativus L.) flour as influenced by particle size. Heliyon 2020, 6, e05471. [Google Scholar] [CrossRef]
- Awolu, O.O.; Sudha, L.M.; Manohar, B. Influence of defatted mango kernel seed flour addition on the rheological characteristics and cookie making quality of wheat flour. Food Sci. Nutr. 2018, 6, 2363–2373. [Google Scholar] [CrossRef]
- Lara-Espinoza, C.; Carvajal-Millán, E.; Balandrán-Quintana, R.; López-Franco, Y.; Rascón-Chu, A. Pectin and pectin-based composite materials: Beyond food texture. Molecules 2018, 23, 945. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhao, C.; Tang, X.; Zhou, J.; Li, H.; Zhang, H.; Liu, J. Physicochemical properties and microstructure of corn flour–cellulose fiber extrudates. Food Sci. Nutr. 2021, 9, 2497–2507. [Google Scholar] [CrossRef]
- Han, K.T.; Kim, H.R.; Moon, T.W.; Choi, S.J. Isothermal and temperature-cycling retrogradation of high-amylose corn starch: Impact of sonication on its structural and retrogradation properties. Ultrason. Sonochem. 2021, 76, 105650. [Google Scholar] [CrossRef] [PubMed]
- Dong, R.; Niu, Q.; Zhang, K.; Hu, X.; Bu, Y. The effect of retrogradation time and ambient relative humidity on the quality of extruded oat noodles. Food Sci. Nutr. 2020, 8, 2940–2949. [Google Scholar] [CrossRef] [PubMed]
- Stribiţcaia, E.; Evans, C.E.L.; Gibbons, C.; Blundell, J.; Sarkar, A. Food texture influences on satiety: Systematic review and meta-analysis. Sci. Rep. 2020, 10, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Jariyah; Widjanarko, S.B.; Yunianta; Estiasih, T.; Sopade, P.A. Pasting properties mixtures of mangrove fruit flour (Sonneratia caseolaris) and starches. Int. Food Res. J. 2014, 21, 2161–2167. [Google Scholar]
- Le Cosquer, G.; Buscail, E.; Gilletta, C.; Deraison, C.; Duffas, J.-P.; Bournet, B.; Tuyeras, G.; Vergnolle, N.; Buscail, L. Incidence and risk factors of cancer in the anal transitional zone and ileal pouch following surgery for ulcerative colitis and familial adenomatous polyposis. Cancers 2022, 14, 530. [Google Scholar] [CrossRef]
- Rabe, H.; Malmquist, M.; Barkman, C.; Östman, S.; Gjertsson, I.; Saalman, R.; Wold, A.E. Distinct patterns of naive, activated and memory T and B cells in blood of patients with ulcerative colitis or Crohn’s disease. Clin. Exp. Immunol. 2019, 197, 111–129. [Google Scholar] [CrossRef]
- Hu, R.; Saw, C.L.L.; Yu, R.; Kong, A.N.T. Regulation of NF-E2-related factor 2 signaling for cancer chemoprevention: Antioxidant coupled with antiinflammatory. Antioxid. Redox Signal. 2010, 13, 1679–1698. [Google Scholar] [CrossRef]
- Guo, F.; Tsao, R.; Li, C.; Wang, X.; Zhang, H.; Jiang, L.; Sun, Y.; Xiong, H. Green pea (Pisum sativum L.) hull polyphenol extracts ameliorate dss-induced colitis through keap1/nrf2 pathway and gut microbiota modulation. Foods 2021, 10, 2765. [Google Scholar] [CrossRef]
- Lin, Y.; Zheng, X.; Chen, J.; Luo, D.; Xie, J.; Su, Z.; Huang, X.; Yi, X.; Wei, L.; Cai, J.; et al. Protective effect of Bruguiera gymnorrhiza (L.) lam. fruit on dextran sulfate sodium-induced ulcerative colitis in mice: Role of Keap1/Nrf2 pathway and gut microbiota. Front. Pharmacol. 2020, 10, 1–17. [Google Scholar] [CrossRef]
- Hardoko; Suprayitno, E.; Puspitasari, Y.E.; Amalia, R. Study of ripe Rhizophora mucronata fruit flour as functional food for antidiabetic. Int. Food Res. J. 2015, 22, 953–959. [Google Scholar]
- Livesey, G.; Taylor, R.; Livesey, H.F.; Buyken, A.E.; Jenkins, D.J.A.; Augustin, L.S.A.; Sievenpiper, J.L.; Barclay, A.W.; Liu, S.; Wolever, T.M.S.; et al. Dietary glycemic index and load and the risk of type 2 diabetes: Assessment of causal relations. Nutrients 2019, 11, 1436. [Google Scholar] [CrossRef] [PubMed]
- Livesey, G.; Taylor, R.; Livesey, H.F.; Buyken, A.E.; Jenkins, D.J.A.; Augustin, L.S.A.; Sievenpiper, J.L.; Barclay, A.W.; Liu, S.; Wolever, T.M.S.; et al. Dietary glycemic index and load and the risk of type 2 diabetes: A systematic review and updated meta-analyses of prospective cohort studies. Nutrients 2019, 11, 1280. [Google Scholar] [CrossRef] [PubMed]
- Jariyah; Susiloningsih, E.K.B.; Nilasari, K. Glycemic index biscuits formulation of pedada flour (Sonneratia caseolaris) with Tubers Starch. In Proceedings of the 2nd International Joint Conference on Science and Technology, Bali, Indonesia, 27–28 September 2017. [Google Scholar]
- Harada, R.; Nomura, T.; Yamada, K.; Mochida, K.; Suzuki, K. Genetic engineering strategies for Euglena gracilis and its industrial contribution to sustainable development goals: A review. Front. Bioeng. Biotechnol. 2020, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Yanxin, L. Avicennia marina Chocolate and Making Process Thereof. CN103141648A, 12 June 2013. [Google Scholar]
- Wen, Q.; Yejun, C. Mangrove Fruit Flavor Food. CN103750209A, 30 April 2014. [Google Scholar]
- Shaozhong, S. Flavoring Sauce with Avicennia marina Fruit and Production Method Thereof. CN104323217A, 9 December 2015. [Google Scholar]
- Liu, N.; Mai, R.; Long, W.; Ma, C. Mangrove Fruit Wine and Its Preparation Method. CN107557227A, 9 January 2018. [Google Scholar]
- Liu, N.; Mai, R.; Long, W.; Ma, C. A Kind of Method for Preparing Mangrove Fruit Tea Bag. CN107593976A, 19 January 2018. [Google Scholar]
- Liu, N.; Mai, R.; Long, W.; Ma, C. A Kind of Method for Preparing Mangrove Fruit Alcohol for Promoting Blood Circulation and Removing Blood Statis. CN107574079A, 12 January 2018. [Google Scholar]
- Su, S. Method for Preparing Synthetic Rice from Mangrove Fruit Starch. CN10516628A, 23 December 2015. [Google Scholar]
- Yanxin, L. Processing Technique of Avicennia marina Fruit. CN103141781A, 25 June 2014. [Google Scholar]
- Huichuan, L. Method for Processing Avicennia marina Fruits. CN104605431A, 18 August 2017. [Google Scholar]
- Zou, T. An Antitumor Mangrove Fruit Particle. CN106107961A, 16 November 2016. [Google Scholar]
- Nan, L.; Rigui, M.; Wanshen, L.; Chunyan, M. Mangrove Fruit Blood-Pressure Decreasing Tea and Preparation Method Thereof. CN107549412A, 9 January 2018. [Google Scholar]
- Fu, H.; Wendong, S.; Fuyong, F.; Weidong, H.; Suhua, O. Method for Making Chocolate with Avicennia Marina. CN101496549A, 5 August 2009. [Google Scholar]
- Weidong, H.; Xiujuan, L. Technique for Processing Fruit of Avicennia Marina. CN101268796A, 24 September 2008. [Google Scholar]
- Erokhin, V.; Gao, T. Impacts of COVID-19 on trade and economic aspects of food security: Evidence from 45 developing countries. Int. J. Environ. Res. Public Health 2020, 17, 5775. [Google Scholar] [CrossRef]
- Minh, N.P. Investigation of mangrove apple (Sonneratia caseolaris) juice production. J. Pharm. Sci. Res. 2019, 11, 809–812. [Google Scholar]
- Hossain, S.J.; Pervin, T.; Suma, S.A. Effects of cooking methods at different time durations on total phenolics and antioxidant activities of fresh and dried-stored fruits of Sonneratia apetala (Buch.-Ham.). Int. Food Res. J. 2016, 23, 556–563. [Google Scholar]
- Basyuni, M.; Siagian, Y.S.; Wati, R.; Putri, L.A.P.; Yusraini, E.; Lesmana, I. Fruit nutrition content, hedonic test, and processed products of pidada (Sonneratia caseolaris). IOP Conf. Ser. Earth Environ. Sci. 2019, 251, 012042. [Google Scholar] [CrossRef]
- Jayatissa, L.P.; Hettiarachi, S.; Dahdouh-Guebas, F. An attempt to recover economic losses from decadal changes in two lagoon systems of Sri Lanka through a newly patented mangrove product. Environ. Dev. Sustain. 2006, 8, 585–595. [Google Scholar] [CrossRef][Green Version]
- Afifah, D.N.; Alamsyah, A.A.M.; Huwaida, A.; Nissa, C.; Wijayanti, H.S.; Purwanti, R.; Hastuti, V.N.; Sugianto, D.N. Cookies made from mangrove (Bruquiera gymnorrhiza) fruit and soybean (Glycine max) flour. Food Res. 2021, 5, 24–36. [Google Scholar] [CrossRef]
- Miranti, D.I.; Ichiura, H.; Ohtani, Y. The bioactive compounds and antioxidant activity of food products of Rhizophora stylosa fruit (coffee and tea mangrove). Int. J. For. Res. 2018, 2018, 2315329. [Google Scholar] [CrossRef]
- Basyuni, M.; Prabuanisa, A.N.; Hamiuddin; Guntur; Kusuma, I.K.T.W.; Oku, H. Distribution of polyprenols and dolichols from fruits and rinds of Barringtonia racemosa, Thespesia populnea, and Xylocarpus granatum. In Proceedings of the 3rd International Seminar on Chemistry: Greeen Chemistry and its Role for Sustainability, Surabaya, Indonesia, 18–19 July 2018; pp. 020075-1–020075-5. [Google Scholar]
- Jariyah, J.; Widjanarko, S.B.; Yunianta; Estiasih, T. Quality Evaluation of Wheat-Pedada Fruit Flour (PFF) Biscuit with Different Emulsifiers. Agric. Agric. Sci. Procedia 2016, 9, 518–524. [Google Scholar] [CrossRef][Green Version]
- Jariyah; Yektiningsih, E.; Sarofa, U. Evaluation of antidiabetic and anticholesterol properties of biscuit product with mangrove fruit flour (MFF) substitution. Carpathian J. Food Sci. Technol. 2019, 11, 141–152. [Google Scholar]
- Fatmah, F.; Utomo, S.W.; Lestari, F. Broccoli-soybean-mangrove food bar as an emergency food for older people during natural disaster. Int. J. Environ. Res. Public Health 2021, 18, 3686. [Google Scholar] [CrossRef] [PubMed]
- Antolak, H.; Piechota, D.; Kucharska, A. Kombucha tea—A double power of bioactive compounds from tea and symbiotic culture of bacteria and yeasts (SCOBY). Antioxidants 2021, 10, 1541. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, S.M.; Hashemi, S.A.; Zarei, M.; Gholami, A.; Lai, C.W.; Chiang, W.H.; Omidifar, N.; Bahrani, S.; Mazraedoost, S. Recent progress in chemical composition, production, and pharmaceutical effects of kombucha beverage: A complementary and alternative medicine. Evid.-Based Complement. Altern. Med. 2020, 2020, 4397543. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.D.; Saimaiti, A.; Luo, M.; Huang, S.Y.; Xiong, R.G.; Shang, A.; Gan, R.Y.; Li, H.B. Fermentation with tea residues enhances antioxidant activities and polyphenol contents in kombucha beverages. Antioxidants 2022, 11, 155. [Google Scholar] [CrossRef] [PubMed]
- Kaewkod, T.; Bovonsombut, S.; Tragoolpua, Y. Efficacy of kombucha obtained from green, oolongand black teas on inhibition of pathogenic bacteria, antioxidation, and toxicity on colorectal cancer cell line. Microorganisms 2019, 7, 700. [Google Scholar] [CrossRef]
- Hardoko; Harisman, E.K.; Puspitasari, Y.E. The kombucha from Rhizophora mucronata Lam. herbal tea: Characteristics and the potential as an antidiabetic beverage. J. Pharm. Pharmacogn. Res. 2020, 8, 410–421. [Google Scholar]
- Yoong, M.H.; Rozaina, T.M.T. Effects of mangrove apple (Sonneratia caseolaris) fruit extract on oxidative stability of palm olein under accelerated storage. Food Res. 2021, 5, 461–470. [Google Scholar] [CrossRef]
- Mondal, S.; Ghosh, D.; Ramakrishna, K. A complete profile on blind-your-eye mangrove Excoecaria agallocha L. (Euphorbiaceae): Ethnobotany, phytochemistry, and pharmacological aspects. Pharmacogn. Rev. 2016, 10, 123–138. [Google Scholar] [CrossRef]
- Mahmud, I.; Islam, M.K.; Saha, S.; Barman, A.K.; Rahman, M.M.; Anisuzzman, M.; Rahman, T.; Al-Nahain, A.; Jahan, R.; Rahmatullah, M. Pharmacological and ethnomedicinal overview of Heritiera fomes: Future prospects. Int. Sch. Res. Not. 2014, 2014, 938543. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.C.; Lin, Y.M.; Feng, D.Q.; Ke, C.H.; Peng, L.; Yan, C.L.; Chen, J.D. A new atisane-type diterpene from the bark of the mangrove plant Excoecaria agallocha. Molecules 2009, 14, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.P.; Luan, Z.L.; Liu, R.X.; Zhang, Q.; Ma, X.C.; Shen, L.; Wu, J. Mangrove tirucallane- and apotirucallane-type triterpenoids: Structure diversity of the C-17 side-chain and natural agonists of human farnesoid/Pregnane–X–Receptor. Mar. Drugs 2018, 16, 488. [Google Scholar] [CrossRef] [PubMed]
- Karthi, S.; Uthirarajan, K.; Manohar, V.; Venkatesan, M.; Chinnaperumal, K.; Vasantha-Srinivasan, P.; Krutmuang, P. Larvicidal enzyme inhibition and repellent activity of red mangrove Rhizophora mucronata (Lam.) leaf extracts and their biomolecules against three medically challenging aarthropod vectors. Molecules 2020, 25, 3844. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.; Baalkhuyur, F.; Valluzzi, L.; Saderne, V.; Cusack, M.; Almahasheer, H.; Krishnakumar, P.K.; Rabaoui, L.; Qurban, M.A.; Arias-Ortiz, A.; et al. Exponential increase of plastic burial in mangrove sediments as a major plastic sink. Sci. Adv. 2020, 6, eaaz5593. [Google Scholar] [CrossRef] [PubMed]
- Analuddin, K.; Sharma, S.; Jamili; Septiana, A.; Sahidin, I.; Rianse, U.; Nadaoka, K. Heavy metal bioaccumulation in mangrove ecosystem at the coral triangle ecoregion, Southeast Sulawesi, Indonesia. Mar. Pollut. Bull. 2017, 125, 472–480. [Google Scholar] [CrossRef]
- Gan, H.; Lin, J.; Liang, K.; Xia, Z. Selected trace metals (As, Cd and Hg) distribution and contamination in the coastal wetland sediment of the northern Beibu Gulf, South China Sea. Mar. Pollut. Bull. 2013, 66, 252–258. [Google Scholar] [CrossRef]
- Chai, M.; Shen, X.; Li, R.; Qiu, G. The risk assessment of heavy metals in Futian mangrove forest sediment in Shenzhen Bay (South China) based on SEM–AVS analysis. Mar. Pollut. Bull. 2015, 97, 431–439. [Google Scholar] [CrossRef]
- Mohebbi-nozar, S.L.; Pauzi, M.; Ruslan, W. Total petroleum hydrocarbons in sediments from the coastline and mangroves of the northern Persian Gulf. Mar. Pollut. Bull. 2015, 95, 407–411. [Google Scholar] [CrossRef]
- Chaudhuri, P.; Nath, B.; Birch, G. Accumulation of trace metals in grey mangrove Avicennia marina fine nutritive roots: The role of rhizosphere processes. Mar. Pollut. Bull. 2014, 79, 284–292. [Google Scholar] [CrossRef]
- Bennett, E.L.; Reynolds, C.J. The value of a mangrove area in Sarawak. Biodivers. Conserv. 1993, 2, 359–375. [Google Scholar] [CrossRef]
- Fourqurean, J.W.; Smith, T.J.; Possley, J.; Collins, T.M.; Lee, D.; Namoff, S. Are mangroves in the tropical Atlantic ripe for invasion? Exotic mangrove trees in the forests of South Florida. Biol. Invasions 2010, 12, 2509–2522. [Google Scholar] [CrossRef]
- Cecep, K. Integrated sustainable mangrove forest management. J. Nat. Resour. Environ. Manag. 2015, 5, 1–6. [Google Scholar] [CrossRef]
- Udoh, J.P. Sustainable nondestructive mangrove-friendly aquaculture in Nigeria II: Models, best practices and policy frame work. AACL Bioflux 2016, 9, 151–173. [Google Scholar]
- Ellison, A.M.; Felson, A.J.; Friess, D.A. Mangrove rehabilitation and restoration as experimental adaptive management. Front. Mar. Sci. 2020, 7, 327. [Google Scholar] [CrossRef]
- Ahmed, N.; Thompson, S.; Glaser, M. Integrated mangrove-shrimp cultivation: Potential for blue carbon sequestration. Ambio 2018, 47, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Chitra, J.; Yacoob, S.A.M.; Kumar, S.S.; Venkataraman, A.; Vijayaraghavan, R.; Nagarajan, Y. HPLC characterization, acute and sub-acute toxicity evaluation of bark extract of Rhizophora mucronata in Swiss Albino mice. Heliyon 2020, 6, e03108. [Google Scholar] [CrossRef]
- Milani, A.S. Mangrove forests of the Persian Gulf and the Gulf of Oman. In Threat to Mangrove Forests., 1st ed.; Makowski, C., Finkl, C.W., Eds.; Springer: Cham, Switzerland, 2018; pp. 53–75. [Google Scholar]
- Rasul, N.M.A.; Stewart, I.C.F.; Nawab, Z.A. Introduction to the Red Sea: Its origin, structure, and environment. In The Red Sea: The Formation, Morphology, Oceanography and Environment of a Young Ocean Basin, 1st ed.; Rasul, N.M.A., Stewart, I.C.F., Eds.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 1–28. [Google Scholar]
- El-Juhany, L.I. Present status and degradation trends of mangrove forests on the Southern Red Sea Coast of Saudi Arabia. Am. J. Agric. Environ. Sci. 2009, 6, 328–340. [Google Scholar]
- Almahasheer, H.; Aljowair, A.; Duarte, C.M.; Irigoien, X. Decadal stability of Red Sea mangroves. Estuar. Coast. Shelf. Sci. 2016, 169, 164–172. [Google Scholar] [CrossRef]
- Chithambaran, S. Restoration of mangrove vegetation at Red Sea coast, Saudi Arabia. Indian J. Geo-Marine Sci. 2019, 48, 1755–1760. [Google Scholar]
- Anton, A.; Almahasheer, H.; Delgado, A.; Garcias-Bonet, N.; Carrillo-de-Albornoz, P.; Marbà, N.; Hendriks, I.E.; Krause-Jensen, D.; Saderne, V.; Baldry, K.; et al. Stunted mangrove trees in the oligotrophic central red sea relate to nitrogen limitation. Front. Mar. Sci. 2020, 7, 597. [Google Scholar] [CrossRef]
- Abohassan, R.A.A.; Okia, C.A.; Agea, J.G.; Kimondo, J.M.; McDonald, M.M. Perennial biomass production in arid mangrove systems on the Red Sea Coast of Saudi Arabia. Environ. Res. J. 2012, 6, 22–31. [Google Scholar]
- Almahasheer, H.; Duarte, C.M.; Irigoien, X. Phenology and growth dynamics of Avicennia marina in the Central Red Sea. Sci. Rep. 2016, 6, 37785. [Google Scholar] [CrossRef] [PubMed]
- Almahasheer, H.; Serrano, O.; Duarte, C.M.; Arias-Ortiz, A.; Masque, P.; Irigoien, X. Low Carbon sink capacity of Red Sea mangroves. Sci. Rep. 2017, 7, 9700. [Google Scholar] [CrossRef] [PubMed]
- Al-Guwaiz, S.M.; Alatar, A.A.; El-Sheikh, M.A.; Al-Gehni, G.A.; Faisal, M.; Qahtan, A.A.; Abdel-Salam, E.M. Role of mangrove rehabilitation and protection plans on carbon storage in Yanbu industrial city, Saudi Arabia: A case study. Sustainability 2021, 13, 13149. [Google Scholar] [CrossRef]
- Abdel-Hamid, A.; Dubovyk, O.; El-Magd, I.A.; Menz, G. Mapping mangroves extents on the Red Sea coastline in Egypt using polarimetric SAR and high resolution optical remote sensing data. Sustainability 2018, 10, 646. [Google Scholar] [CrossRef]
- Gab-Alla, A.A.; Ishrak, K.K.; Waleed, M.M.; Fouda, M.M. Ecology of Avicennia marina mangals along Gulf of Aqaba, South Sinai, Red Sea. Egypt. J. Aquat. Biol. Fish. 2010, 14, 79–93. [Google Scholar] [CrossRef]
- Afefe, A.A.; Khedr, A.H.A.; Abbas, M.S.; Soliman, A.S. Responses and tolerance mechanisms of mangrove trees to the ambient salinity along the Egyptian Red Sea Coast. Limnol. Rev. 2021, 21, 3–13. [Google Scholar] [CrossRef]
- Afefe, A.A.; Abbas, M.S.; Soliman, A.S.; Khedr, A.H.A.; Hatab, E.B.E. Physical and chemical characteristics of mangrove soil under marine influence. A case study on the Mangrove Forests at Egyptian-African Red Sea Coast. Egypt. J. Aquat. Biol. Fish. 2019, 23, 385–399. [Google Scholar] [CrossRef]
- Kumar, A.; Khan, M.A.; Muqtadir, A. Distribution of mangroves along the Red Sea Coast of the Arabian Peninsula: Part-3: Coast of Yemen. Earth Sci. India 2010, 3, 28–42. [Google Scholar]
- Alshawafi, A.; Analla, M.; Aksissou, M.; Triplet, P. Physicochemical properties of water, soil, and morphological characteristics of mangrove forests in the Island of Kamaran, Al Hodaidah, Yemen. J. Ecosyst. Ecography 2016, 06, 4. [Google Scholar] [CrossRef]
- Almahasheer, H. Internodal analysis of Avicennia marina in the Western Arabian Gulf. Front. Mar. Sci. 2021, 8, 698596. [Google Scholar] [CrossRef]
- Al-Khayat, J.; Balakrishnan, P. Avicennia marina around Qatar: Tree, seedling and pneumatophore densities in natural and planted mangroves using remote sensing. Int. J. Sci. 2014, 3, 18–27. [Google Scholar]
- Al Jufaili, S.M.; Jawad, L.A.; Park, J.M.; Al Sariri, T.S.; Al Balushi, B.Y. Fish diversity of mangrove ecosystems in Sultanate of Oman. Cah. Biol. Mar. 2021, 62, 235–249. [Google Scholar]
- Samara, F.; Solovieva, N.; Ghalayini, T.; Nasrallah, Z.A.; Saburova, M. Assessment of the environmental status of the mangrove ecosystem in the United Arab Emirates. Water 2020, 12, 1623. [Google Scholar] [CrossRef]
- Ibrahim, H.A.H.; Abdel-Latif, H.H.; Zaghloul, E.H. Phytochemical composition of Avicennia marina leaf extract, its antioxidant, antimicrobial potentials and inhibitory properties on Pseudomonas fluorescens biofilm. Egypt. J. Aquat. Res. 2022, 48, 29–35. [Google Scholar] [CrossRef]
- Abu-Odeh, A.M.; Talib, W.H. Middle East medicinal plants in the treatment of diabetes: A review. Molecules 2021, 26, 742. [Google Scholar] [CrossRef]
- Tounekti, T.; Mahdhi, M.; Khemira, H. Ethnobotanical study of indigenous medicinal plants of Jazan region, Saudi Arabia. Evid.-Based Complement. Altern. Med. 2019, 2019, 3190670. [Google Scholar] [CrossRef]
- Abou-Elela, G.M.; El-Sersy, M.A.; El-Shenawy, M.A.; Abd-Elnabi, H.; Ibrahim, H.A.H. Bio-control of Vibrio fluvialis in aquaculture by mangrove (Avicennia marina) seeds extracts. Res. J. Microbiol. 2009, 4, 38–48. [Google Scholar]
- Hamed, M.M.; Abdrabo, M.A.A.; Fahmy, N.M.; Abdelfattah, L.S.; Kelany, M.S.; Abd-El Latif, H.H.; Abou-Elela, G.M.; Abd-Elnaby, H.M.; Hassan, S.W.M. Distribution and characterization of actinomycetes in mangrove habitats (Red Sea, Egypt) with special emphasis on Streptomyces mutabilis M3MT483919. J. Pure Appl. Microbiol. 2021, 15, 246–261. [Google Scholar] [CrossRef]
- Hodhod, M.S.E.D.; Gaafar, A.R.Z.; Alshameri, A.; Qahtan, A.A.; Noor, A.; Abdel-Wahab, M. Molecular characterization and bioactive potential of newly identified strains of the extremophilic black yeast Hortaea werneckii isolated from Red Sea mangrove. Biotechnol. Biotechnol. Equip. 2020, 34, 1288–1298. [Google Scholar] [CrossRef]
 , carbohydrate;
, carbohydrate;  , lipid;
, lipid;  , protein; *, carbohydrate is presented by total sugar and lipid is presented by fat. Modified from [41,42,43].
, protein; *, carbohydrate is presented by total sugar and lipid is presented by fat. Modified from [41,42,43].

















| Mangrove species | Solvent | Compound | Bioactivity | Ref |
|---|---|---|---|---|
| Xylocarpus granatum | Water-ethanol | Gedunin (1) | Anticancer | [70] |
| X. granatum | Ethanol | Xyloccensin K (2) | Nt | [71] |
| 6-acetoxycedrodorin (3) | Nt | |||
| Xyloccensin W (4) | Nt | |||
| X. granatum | Ethanol | 3-deacetyl xyloccensin M (5) | Nt | [72] |
| 3-deacetyl xyloccensin N (6) | Nt | |||
| X. granatum | Ethanol | Xyloccensin X1 (7) | Nt | [73] |
| Xyloccensin X2 (8) | Nt | |||
| X. granatum | Ethanol | Xyloccensin Y (9) | - | [74] |
| Xyloccensin Z1 (10) | - | |||
| Xyloccensin Z2 (11) | - | |||
| X. granatum | Ethanol | Xylogranatin A (12) | Nt | [75] |
| Xylogranatin B (13) | Nt | |||
| Xylogranatin C (14) | Nt | |||
| Xylogranatin D (15) | Nt | |||
| X. granatum | Ethanol | Xylogranatin E (16) | Nt | [76] |
| X. granatum | Ethanol | Xylocarpin A (17) | Nt | [69] |
| Xylocarpin B (18) | Nt | |||
| Xylocarpin C (19) | Nt | |||
| Xylocarpin D (20) | Nt | |||
| Xylocarpin E (21) | Nt | |||
| 6-dehydroxyxylocarpin D (22) | Nt | |||
| Xylocarpin F (23) | Nt | |||
| Xylocarpin G (24) | Nt | |||
| Xylocarpin H (25) | Nt | |||
| Xylocarpin I (26) | Nt | |||
| X. granatum | Ethanol | Xylogranatinin (27) | Nt | [77] |
| X. granatum | Ethanol | Protoxylocarpin A (28) | Antitumor | [78] |
| Protoxylocarpin B (29) | Antitumor | |||
| Protoxylocarpin C (30) | Antitumor | |||
| Protoxylocarpin D (31) | Antitumor | |||
| Protoxylocarpin E (32) | Antitumor | |||
| Xylocarpin J (33) | Antitumor | |||
| Xylocarpin K (34) | - | |||
| Xyloccensin M (35) | Antitumor | |||
| X. granatum | Ethanol | Butyrospermol 3β-O-palmitate (36) | - | [79] |
| Butyrospermol 3β-O-oleate (37) | - | |||
| Butyrospermol 3β-O-stearate (38) | - | |||
| Butyrospermol 3β-O-linoleate (39) | - | |||
| A. marina | Ethanol | Maricaffeolylide A (40) | Antioxidant | [80] |
| Maricyclohexene A (41) | Antioxidant | |||
| A. marina | Ethanol-CH2Cl2 | Marinoid F (42) | Antioxidant | [81] |
| Marinoid G (43) | Antioxidant | |||
| Marinoid H (44) | Antioxidant | |||
| Marinoid I (45) | Antioxidant | |||
| A. marina | Ethanol-CH2Cl2 | Marinoid J (46) | Antioxidant | [82] |
| Marinoid K (47) | Antioxidant | |||
| Marinoid L (48) | Antioxidant | |||
| Marinoid M (49) | Antioxidant | |||
| X. granatum | Ethanol | Granatione (50) | Nt | [83] |
| Xylocarpin L (51) | Nt | |||
| C. tagal | Hexane-CH2Cl2 | Cereotagaloperoxide (52) | - | [84] |
| Cereotagalol A (53) | - | |||
| Cereotagalol B (54) | - | |||
| Isofouquierol (55) | - | |||
| Fouquierol (56) | - | |||
| 3β-E-feruloylbetulinic acid (57) | Anticancer | |||
| S. apetala | Ethanol | Sonneradon A (58) | Nematode’s life expansion | [85] |
| Sonneradon B (59) | Nematode’s life expansion | |||
| Sonneradon C (60) | - | |||
| Sonneradon D (61) | Nematode’s life expansion | |||
| Ranuncoside (62) | - | |||
| Apigenin (63) | Nematode’s life expansion | |||
| Luteoline (64) | - | |||
| 6-O-galloyl-d-glucopyranose (65) | - | |||
| O-B-(6-O-galloyl)-glucopyranoside (66) | - | |||
| 2-ethylhexyl phthalate (67) | - | |||
| Methyl gallate (68) | - | |||
| Methyl 4-O-methylgallate (69) | - | |||
| 4-O-methylgallic acid (70) | - | |||
| 4-methoxybenzoic acid (71) | - | |||
| 3,4-dihydrobenzoic acid (72) | - | |||
| Bibutyl malate (73) | - | |||
| Dimethyl malate (74) | Nematode’s life expansion | |||
| Bibutyl malate (75) | - | |||
| Ethylmethyl malate (76) | - | |||
| 2-butenedioic acid (77) | - | |||
| 3-hydroxy-4-oxobutanoic acid (78) | - | |||
| Butylmethyl malate (79) | Nematode’s life expansion | |||
| X. granatum | Acetonitrile | Piscidinol G (80) | Nt | [86] |
| Xylogranation D (81) | Nt | |||
| Spicatin (82) | Nt | |||
| Xylogranatin C (83) | Nt | |||
| Xyloccensin V (84) | Nt | |||
| Proceranolide (85) | Nt | |||
| Xylomexicanin D (86) | Nt | |||
| Sapelin E acetate (87) | Nt | |||
| Xylomexicanin A (88) | Nt | |||
| Grandifoliolenone (89) | Nt | |||
| Odoratone (90) | Nt |
| Mangrove species | Solvent | Compound | Bioactivity | Ref |
|---|---|---|---|---|
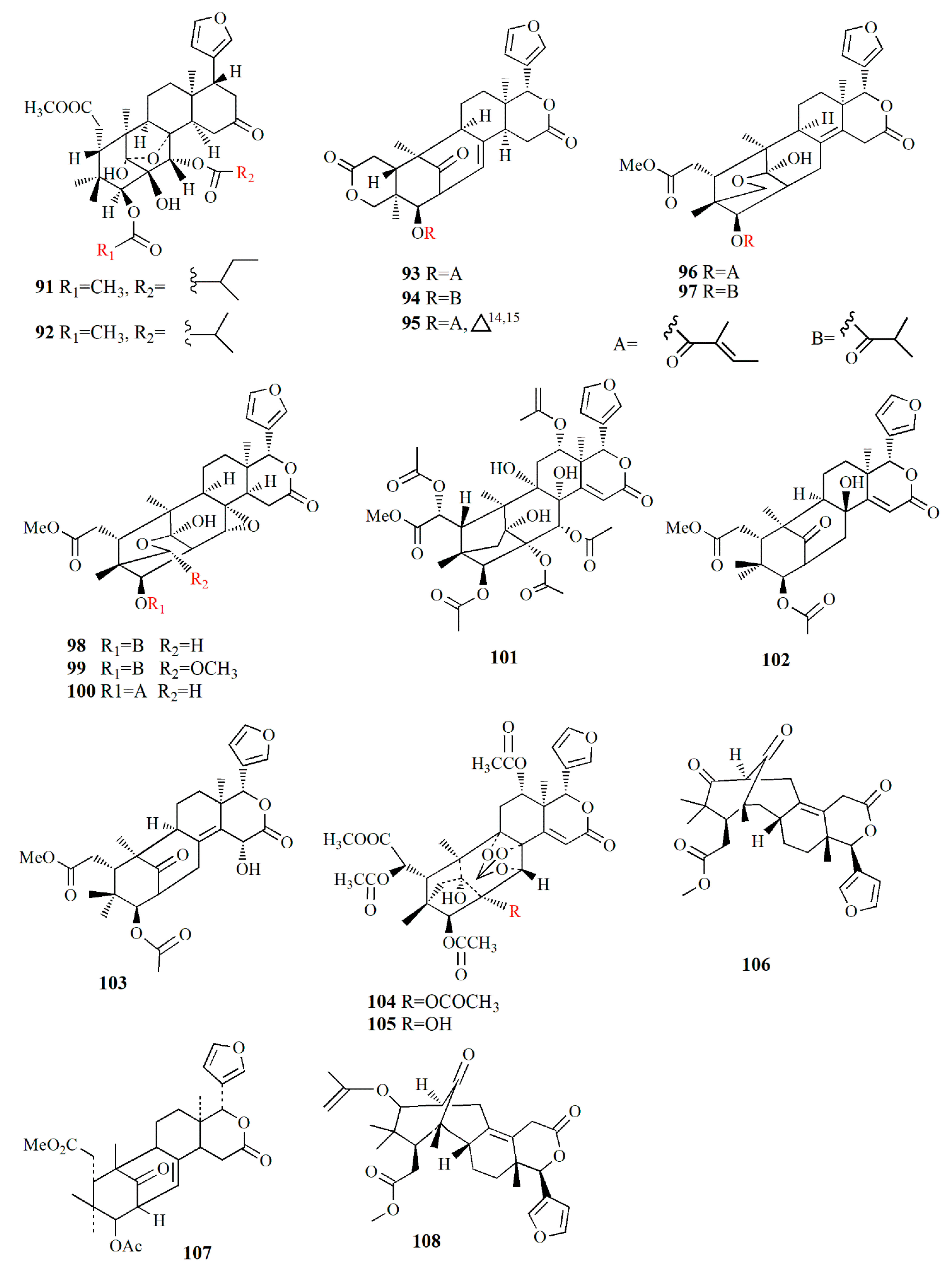
| X. granatum | Light petroleum | Xyloccensin I (91) | - | [89] |
| Xyloccensin J (92) | - | |||
| X. moluccensis | Ethanol | Godavarin A (93) | Insecticidal and antifeedant | [90] |
| Godavarin B (94) | - | |||
| Godavarin C (95) | - | |||
| Godavarin D (96) | Insecticidal and antifeedant | |||
| Godavarin E (97) | - | |||
| Godavarin F (98) | - | |||
| Godavarin G (99) | - | |||
| Xyloccensin L (100) | - | |||
| Godavarin H (101) | - | |||
| Godavarin I (102) | - | |||
| Godavarin J (103) | Insecticidal and antifeedant | |||
| Xyloccensin P (104) | - | |||
| Xyloccensin Q (105) | - | |||
| Angustidienolide (106) | Insecticidal and antifeedant | |||
| 6-deoxy-3detigloyl-swietenine acetate (107) | Insecticidal and antifeedant | |||
| Fissinolide (108) | Insecticidal and antifeedant | |||
| Methyl 3β-acetoxy-1-oxomeliaca-8(9),14-dienoate (109) | Insecticidal and antifeedant | |||
| X. granatum | Ethanol | 30α-hydroxyl xylogranatin A (110) | Nt | [91] |
| Xylogranatin E2 (111) | Nt | |||
| X granatum | Ethanol | Thaigranatin A (112) | - | [92] |
| Thaigranatin B (113) | - | |||
| Thaigranatin C (114) | - | |||
| Thaigranatin D (115) | - | |||
| Thaigranatin E (116) | - | |||
| Granatumin L (117) | Anti-HIV | |||
| X. granatum | Ethanol | Xylomexicanin A (118) | Antitumor | [93] |
| Xylomexicanin B (119) | - | |||
| X. moluccensis | Ethanol | Moluccensin R (120) | Antifeedant | [94] |
| Moluccensin S (121) | - | |||
| 6-hydroxymexicanolide (122) | Antifeedant | |||
| 2-hydroxyfissinoide (123) | Antifeedant | |||
| Moluccensin T (124) | - | |||
| Moluccensin U (125) | - | |||
| Moluccensin V (126) | - | |||
| Moluccensin W (127) | - | |||
| Moluccensin X (128) | - | |||
| Moluccensin Y (129) | - | |||
| X. moluccensis | Ethanol | Krishnolide A (130) | Anti-HIV | [95] |
| Krishnolide B (131) | - | |||
| Krishnolide C (132) | - | |||
| Krishnolide D (133) | - | |||
| X. moluccensis | Ethanol | Xylomolin A1 (134) | - | [96] |
| Xylomolin A2 (135) | - | |||
| Xylomolin A3 (136) | - | |||
| Xylomolin A4 (137) | - | |||
| Xylomolin A5 (138) | - | |||
| Xylomolin A6 (139) | - | |||
| Xylomolin A7 (140) | - | |||
| Xylomolin B1 (141) | - | |||
| Xylomolin B2 (142) | - | |||
| Xylomolin C1 (143) | - | |||
| Xylomolin C2 (144) | - | |||
| Xylomolin D (145) | - | |||
| Xylomolin E (146) | - | |||
| Xylomolin F (147) | - | |||
| Xylomolin G1 (148) | - | |||
| Xylomolin G2 (149) | - | |||
| Xylomolin G3 (150) | - | |||
| Xylomolin G4 (151) | - | |||
| Xylomolin G5 (152) | - | |||
| Xylomolin H (153) | - | |||
| Xylomolin I (154) | - | |||
| Xylomolin J1 (155) | - | |||
| Xylomolin J2 (156) | Anticancer | |||
| Xylomolin K1 (157) | - | |||
| Xylomolin K2 (158) | - | |||
| Xylomolin L1 (159) | - | |||
| Xylomolin L2 (160) | - | |||
| Xylomolin M (161) | - | |||
| Xylomolin N (162) | - | |||
| X. moluccensis | Ethanol | Thaixylomolin O (163) | - | [97] |
| Thaixylomolin P (164) | - | |||
| Thaixylomolin Q (165) | - | |||
| Thaixylomolin R (166) | - | |||
| X. granatum | Ethanol | Granatumin M (167) | - | [98] |
| Granatumin N (168) | - | |||
| Granatumin O (169) | - | |||
| Granatumin P (170) | - | |||
| Granatumin Q (171) | - | |||
| Granatumin R (172) | - | |||
| Granatumin S (173) | - | |||
| Granatumin T (174) | - | |||
| Granatumin U (175) | - | |||
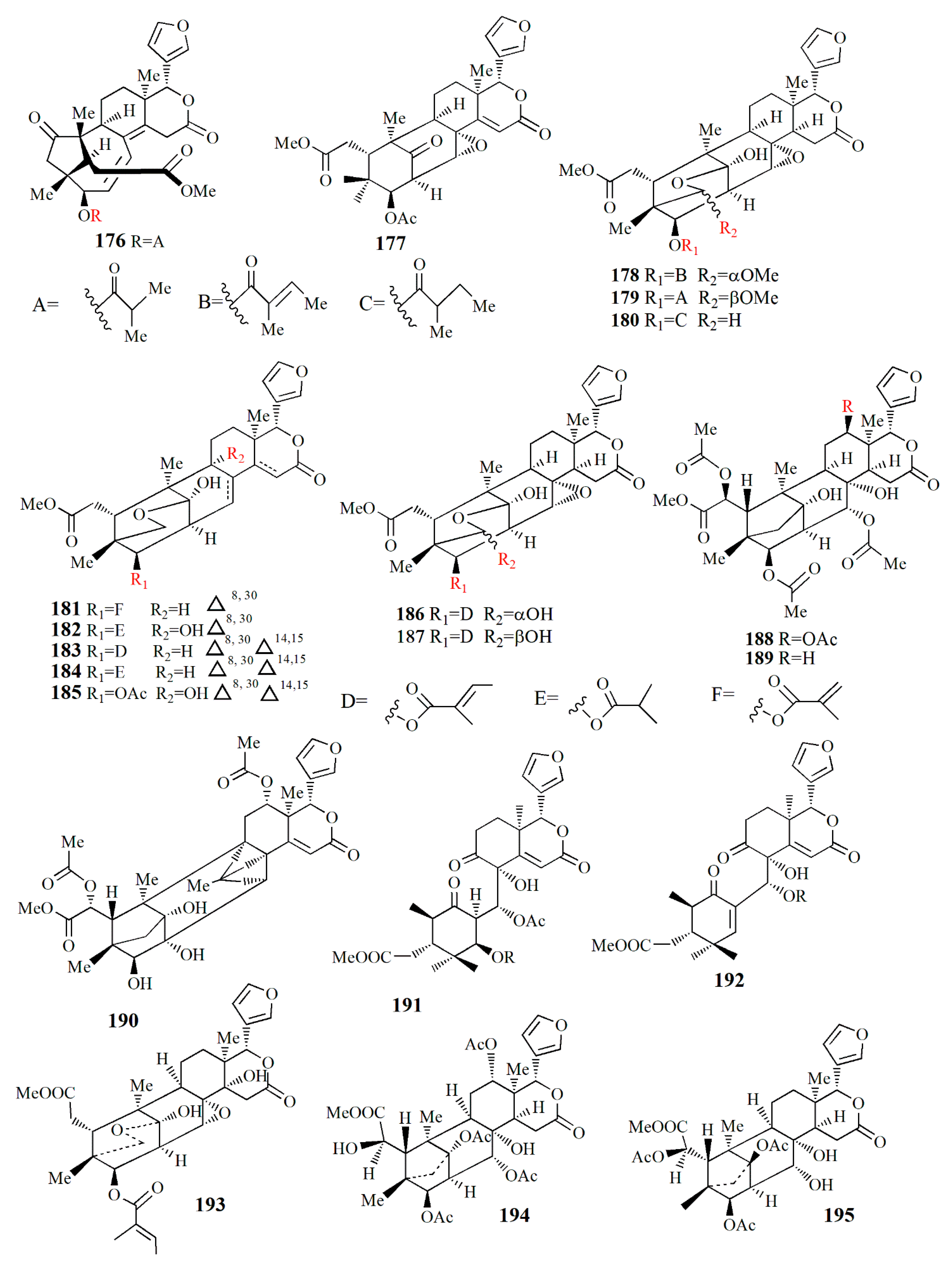
| X. granatum | Methanol | Sundarbanxylogranin A (176) | - | [99] |
| Sundarbanxylogranin B (177) | Anti-HIV | |||
| Sundarbanxylogranin C (178) | - | |||
| Sundarbanxylogranin D (179) | - | |||
| Sundarbanxylogranin E (180) | - | |||
| X granatum | Ethanol | Granatumin X (181) | - | [98] |
| Krishnagranatinin A (182) | - | |||
| Krishnagranatinin B (183) | - | |||
| Krishnagranatinin C (184) | - | |||
| Krishnagranatinin D (185) | - | |||
| Krishnagranatinin E (186) | - | |||
| Krishnagranatinin F (187) | - | |||
| Krishnagranatinin G (188) | Inhibit NF-κB | |||
| Krishnagranatinin H (189) | Inhibit NF-κB | |||
| Krishnagranatinin I (190) | Inhibit NF-κB | |||
| X. granatum | Ethanol | Granaxylocarpin A (191) | Anticancer | [100] |
| Granaxylocarpin B (192) | Anticancer | |||
| Granaxylocarpin C (193) | - | |||
| Granaxylocarpin D (194) | - | |||
| Granaxylocarpin E (195) | - | |||
| X. granatum | Ethanol | Thaixylogranin A (196) | Anticancer | [101] |
| Thaixylogranin B (197) | Anticancer | |||
| Thaixylogranin C (198) | Anticancer | |||
| Thaixylogranin D (199) | Anticancer | |||
| Thaixylogranin E (200) | Anticancer | |||
| Thaixylogranin F (201) | Anticancer | |||
| Thaixylogranin G (202) | Anticancer | |||
| Thaixylogranin H (203) | Anticancer | |||
| X. moluccensis | Ethanol | Trangmolin A (204) | - | [102] |
| Trangmolin B (205) | - | |||
| Trangmolin C (206) | - | |||
| Trangmolin D (207) | - | |||
| Trangmolin E (208) | - | |||
| Trangmolin F (209) | - | |||
| X. moluccensis | Ethanol | Thaixylomolin G (210) | - | [103] |
| Thaixylomolin H (211) | - | |||
| Thaixylomolin I (212) | Anti-H1N1 | |||
| Thaixylomolin J (213) | - | |||
| Thaixylomolin K (214) | Anti-H1N1 | |||
| Thaixylomolin L (215) | - | |||
| Thaixylomolin M (216) | Anti-H1N1 | |||
| Thaixylomolin N (217) | - | |||
| 12-deacetylxyloccensin U (218) | - | |||
| 2-O-acetyl-2-dehydroxy-12-deacetylxyloccensin (219) | - | |||
| 6-O-acetyl-2a-hydroxymexicanolide (220) | - | |||
| 6-O-acetyl-6-dehydroxymoluccensin T (221) | - | |||
| X. rumphii | Methanol | Xylorumphiin E (222) | - | [104] |
| Xylorumphiin F (223) | - | |||
| 2-hydroxyxylorumphiin F (224) | Antiinflammatory | |||
| Xylorumphiin G (225) | Antiinflammatory | |||
| Xylorumphiin H (226) | - | |||
| Xylorumphiin I (227) | - | |||
| Xylorumphiin J (228) | - |
| Mangrove species | Fungus species | Cultivation media | Compound | Bioactivity | Ref |
|---|---|---|---|---|---|
| K. candel | Penicillium commune | Rice substrate | Peniisocoumarin A (229) | - | [106] |
| Peniisocoumarin B (230) | - | ||||
| Peniisocoumarin C (231) | α-glucosidase inhibition | ||||
| Peniisocoumarin D (232) | - | ||||
| Peniisocoumarin E (233) | α-glucosidase inhibition | ||||
| Peniisocoumarin F (234) | α-glucosidase inhibition | ||||
| Peniisocoumarin G (235) | α-glucosidase inhibition | ||||
| Peniisocoumarin H (236) | - | ||||
| Peniisocoumarin I (237) | α-glucosidase inhibition | ||||
| 3-[-(R)-3,3-dichloro-2-hydroxypropyl]-8-hydroxy-6-methoxy-1H-isochromen-1-on1 (238) | α-glucosidase inhibition | ||||
| Peniisocoumarin J (239) | α-glucosidase inhibition | ||||
| (+)-6-methyl-citreoisocoumarin (240) | - | ||||
| (+)-diaporthin (241) | - | ||||
| S. caseolaris | Alternaria sp. | Rice substrate | Altenusin derivative 1 (242) | - | [107] |
| Altenusin derivative 2 (243) | α-glucosidase inhibition | ||||
| Altenusin derivative 3 (244) | α-glucosidase inhibition | ||||
| Altenusin derivative 4 (245) | α-glucosidase inhibition | ||||
| Altenusin derivative 5 (246) | - | ||||
| Talaroflavone (247) | α-glucosidase inhibition | ||||
| Deoxyrubralactone (248) | - | ||||
| Rubralactone (249) | α-glucosidase inhibition | ||||
| 2-OH-AOH (250) | α-glucosidase inhibition | ||||
| Alternariol (251) | α-glucosidase inhibition | ||||
| Alternariol methyl ether (252) | - | ||||
| Acanthus ilicifolius | Epicoccum nigrum | Wheat solid substrate | Racemix (±)-epicoccone C (253) | α-glucosidase inhibition | [108] |
| Epicoccone D (254) | α-glucosidase inhibition | ||||
| Epicoccone E (255) | α-glucosidase inhibition | ||||
| Epicolactone A (256) | α-glucosidase inhibition | ||||
| Epicolactone (257) | - | ||||
| Flavimycins A (258) | α-glucosidase inhibition | ||||
| Epicocconigrone A (259) | α-glucosidase inhibition | ||||
| Epicoccolide B (260) | α-glucosidase inhibition | ||||
| A. marina | Aspergillus versicolor | White bean | Allantopyrone E (261) | Anticancer | [109] |
| K. candel | Botryosphaeria sp. | Rice substrate | Botryoisocoumarin A (262) | COX-2 inhibition | [110] |
| Monocerin (263) | - | ||||
| 3-methyl-6,8-dihydroxyisocoumarin (264) | - | ||||
| 8-methoxymellein (265) | - | ||||
| Trans-4-hydroxymellein (266) | - | ||||
| 5-hydroxy-7-methoxy-4,6-dimethyl phthalide (267) | - | ||||
| K. obovata | Talaromyces amestolkiae | Rice substrate | Talanaphthoquinone A (268) | Antioxidant | [111] |
| Talanaphthoquinone B (269) | Antioxidant | ||||
| Anhydrojavanicin (270) | Antioxidant | ||||
| 2,3-dihydro-5-hydroxy-4-hydroxymethyl-8-methoxy-2-methylnaphtho[1,2-b]furan-6,9-dione (271) | Antioxidant | ||||
| Anhydrofusarubin (272) | Antioxidant | ||||
| 2-acetonyl-3-methyl-5-hydroxy-7-methoxy-naphthazarin (273) | Antioxidant | ||||
| 6-ethyl-2,7-dimethoxyjuglone (274) | Antioxidant | ||||
| 6-[1-(acetyloxy)ethyl]-5-hydroxy-2,7-dimethoxy1,4-naphthalenedione (275) | Antioxidant | ||||
| 5-hydroxy-6-(1-hydroxyethyl)-2,7-dimethoxy-1,4-naphthalenedione (276) | Antioxidant | ||||
| Solaniol (277) | Antioxidant | ||||
| Javanicin (278) | Antioxidant | ||||
| Bruguiera sp. | Mycosphaerella sp. | Rice substrate | Asperchalasine I (279) | α-glucosidase inhibitor, antioxidant | [112] |
| Dibefurin B (280) | - | ||||
| (R)-9-((R)-10-hydroxyethyl)-7,9-dihydroisobenzofuran-1-ol (281) | - | ||||
| 2-methoxycarbonyl-4,5,6-trihydroxy-3-methyl-benzaldehyde (282) | Antioxidant | ||||
| Epicoccone B (283) | - | ||||
| 1,3-dihydro-5-methoxy-7-methylisobenzofuran (284) | Antioxidant | ||||
| Paeciloside A (285) | - | ||||
| Asperchalasine A (286) | α-glucosidase inhibitor, antioxidant | ||||
| Aspochalasin I (287) | - | ||||
| S. apetala | Pseudofusicoccum sp. | Rice substrate | Acorenone C (288) | AChE inhibition | [113] |
| Uracil (289) | - | ||||
| Cyclo-(L-Pro-L-Tyr) (290) | - | ||||
| Bis-(2-ethylhexyl) terephthalate (291) | - | ||||
| 4-hydroxybenzaldehyde (292) | - | ||||
| 2-phenylethanol (293) | - | ||||
| 4-hydroxyphenethyl alcohol (294) | - | ||||
| Estigmast-4-en-6β-ol-3-ona (295) | - | ||||
| Ergosterol (296) | NO production inhibition; anticancer | ||||
| Ergosterol peroxide (297) | - | ||||
| Cerevisterol (298) | - | ||||
| Aegiceras corniculatum | Alternaria sp. | Potato dextrose broth | Alterporriol K (299) | Anticancer | [114] |
| Alterporriol L (300) | Anticancer | ||||
| Alterporriol M (301) | - | ||||
| Physcion (302) | - | ||||
| Marcrospin (303) | - | ||||
| Dactylariol (304) | - | ||||
| Tetrahydroaltersolanol B (305) | - |
| Innovation | Patent No | Ref |
|---|---|---|
| Processing the mangrove fruit | CN10314178B | [154] |
| The fruit is soaked in saline water and then cleaned after the peel is softened. The fruit is sprayed with white wine and then soaked in hot water at 70–90 °C. The peel is removed afterward. The fruit is soaked in warm water at 30–40 °C then dried, sterilized, and packaged. | ||
| Processing the mangrove fruit | CN10460543A | [155] |
| The fruit is cleaned and peeled then ground using an ultra-micro grinder to make a fine powder. The powder is mixed with water, homogenized, and enzymolized by protease. The filtrate is removed and vacuumed to decolorize from dark green to white. The slurry is dry, sterilized, and packaged. | ||
| Synthetic rice from mangrove fruit starch | CN105166628 | [153] |
| Brief description: mangrove fruit flour, glutinous rice flour, cornstarch, converted starch, and konjac flour are mixed and then pre-gelatinized. The pre-gelatinized dough is granulated, steamed, dried, then polished to make synthetic rice. | ||
| Antitumor from mangrove fruit particle | CN106107961 | [156] |
| Brief description: the fruit is cleaned and then mixed with water to make liquor. White sugar is added and centrifuged. The slurry is mixed with methylcellulose, glyceryl monostearate, and banana juice. The mixture is pelleted, dried, and sterilized. | ||
| Wine from mangrove fruit | CN107557227 | [150] |
| Brief description: mangrove fruit is soaked with limewash for 12–24 h at 50–80 °C, then cleaned. The yeast is added to the paste and fermented at 34–36 °C and the filtrate is collected afterward. The filtrate is fermented for 10–20 days to produce wine. | ||
| Alcohol from mangrove fruit for removing blood stasis | CN107574079 | [152] |
| Brief description: mangrove fruit is soaked in limewash, cleaned, then mixed with glutinous rice to make a paste. The yeast is added and fermented to produce alcohol. | ||
| Teabag from mangrove fruit | CN107593976 | [151] |
| Brief description: the mangrove fruit is soaked with limewash then cleaned and dried. The biomass is mixed with the fresh flower and then dried together as tea. | ||
| Tea to decrease the blood-pressure from mangrove fruit | CN107549412A | [157] |
| Brief description: mangrove fruit is soaked in limewash then cleaned and dried. Thorn apple is soaked in an alcoholic solution and then dried. The dried thorn apple and mangrove fruit are ground and mixed. The powder is ready as tea. | ||
| Chocolate from mangrove fruit Avicennia marina | CN103141648A | [147] |
| Brief description: the mangrove fruit is cleaned and mixed with liquor. The pulp is mixed with cocoa powder, whole milk powder, and skimmed milk powder to form chocolate. | ||
| Chocolate from mangrove fruit | CN101496549A | [158] |
| The fruit is soaked in boiling water followed by cold water, then the peel is removed. The fruit is soaked in boiling water several times. The fruit is dried, crushed, and ground to make powder. The powder is mixed with milk powder, cocoa butter, sugar, and cocoa powder. The mixture is mixed, molded, and packed. | ||
| Flavoring food from mangrove fruit | CN103750209A | [148] |
| Brief description: the mangrove fruit is mixed with vegetable protein hydrolase. The enzymolysis pulp is mixed with xanthan, acesulfame, and sugar, and then sterilized. The mixture is ready for flavoring food. | ||
| Flavoring sauce from mangrove fruit | CN104323217A | [149] |
| Brief description: the mangrove fruit is boiled and then mixed with pepper powder, Chinese cassia, and cardamom. The mixture is then boiled and packaged. | ||
| Syrup from mangrove fruit | CN101268796A | [159] |
| Brief description: the mangrove fruit is sterilized at 95–100 °C for 5–6 minutes then cooled. The fruit is soaked with CaCl2 and then cleaned. The honey containing sugar (1:30) is added followed by citric acid. The fruit is boiled and then packed into a can. |
| Location | Sample type | Experiment | Activity | Ref |
|---|---|---|---|---|
| Jazan, Red Sea coast of Saudi Arabia | Fruit of A. marina | Ethanol extract | Antibacterial against P. aeruginosa, B. subtilis, S. aureus, E.coli Antifungal against A. fumigatus, C. albicans | [58] |
| Seed of A. marina | Ethanol extract | Antifungal activities against A. fumigatus | ||
| Root of A. marina | Chloroform extract | Antibacterial against S. aureus, E. coli Antifungal against A. fumigatus | ||
| Leaves of A. marina | Ethyl acetate extract | Antibacterial against S. aureus, E. coli | ||
| Safaga, Red Sea coast of Egypt | Seed of A. marina | Chloroform and ethanol extract | Antibacterial against P. aeruginosa, V. fluvialis, V. vulnificus, S. fecalis, E. coli, B. subtilis, S. aureus | [218] |
| Leaves, stems, and roots of A. marina | Chloroform extract | Antibacterial against P. aeruginosa, V. fluvialis, V. vulnificus, S. fecalis, E. coli, B. subtilis, S. aureus | ||
| Leaves, stems, and roots of A. marina | Ethanol extract | Antibacterial against V. fluvialis, V. vulnificus | ||
| Red Sea coast of Egypt | Sediment sample from mangrove forest | Actinomycetes isolation and extraction | Antimicrobial against B. subtilis, E. coli, S. aureus, P. aeruginosa, C. albicans. | [219] |
| Red Sea coast of Saudi Arabia | Decayed leaves of A. marina | Black yeast Hortaea werneckii | Antimicrobial against pathogen S. aureus, Campylobacter jejuni, and S. typhimurium. | [220] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Budiyanto, F.; Alhomaidi, E.A.; Mohammed, A.E.; Ghandourah, M.A.; Alorfi, H.S.; Bawakid, N.O.; Alarif, W.M. Exploring the Mangrove Fruit: From the Phytochemicals to Functional Food Development and the Current Progress in the Middle East. Mar. Drugs 2022, 20, 303. https://doi.org/10.3390/md20050303
Budiyanto F, Alhomaidi EA, Mohammed AE, Ghandourah MA, Alorfi HS, Bawakid NO, Alarif WM. Exploring the Mangrove Fruit: From the Phytochemicals to Functional Food Development and the Current Progress in the Middle East. Marine Drugs. 2022; 20(5):303. https://doi.org/10.3390/md20050303
Chicago/Turabian StyleBudiyanto, Fitri, Eman A. Alhomaidi, Afrah E. Mohammed, Mohamed A. Ghandourah, Hajer S. Alorfi, Nahed O. Bawakid, and Wailed M. Alarif. 2022. "Exploring the Mangrove Fruit: From the Phytochemicals to Functional Food Development and the Current Progress in the Middle East" Marine Drugs 20, no. 5: 303. https://doi.org/10.3390/md20050303
APA StyleBudiyanto, F., Alhomaidi, E. A., Mohammed, A. E., Ghandourah, M. A., Alorfi, H. S., Bawakid, N. O., & Alarif, W. M. (2022). Exploring the Mangrove Fruit: From the Phytochemicals to Functional Food Development and the Current Progress in the Middle East. Marine Drugs, 20(5), 303. https://doi.org/10.3390/md20050303