
Fungal Bergamotane Sesquiterpenoids—Potential Metabolites: Sources, Bioactivities, and Biosynthesis

 ,
,  ,
,  and
and 
Abstract

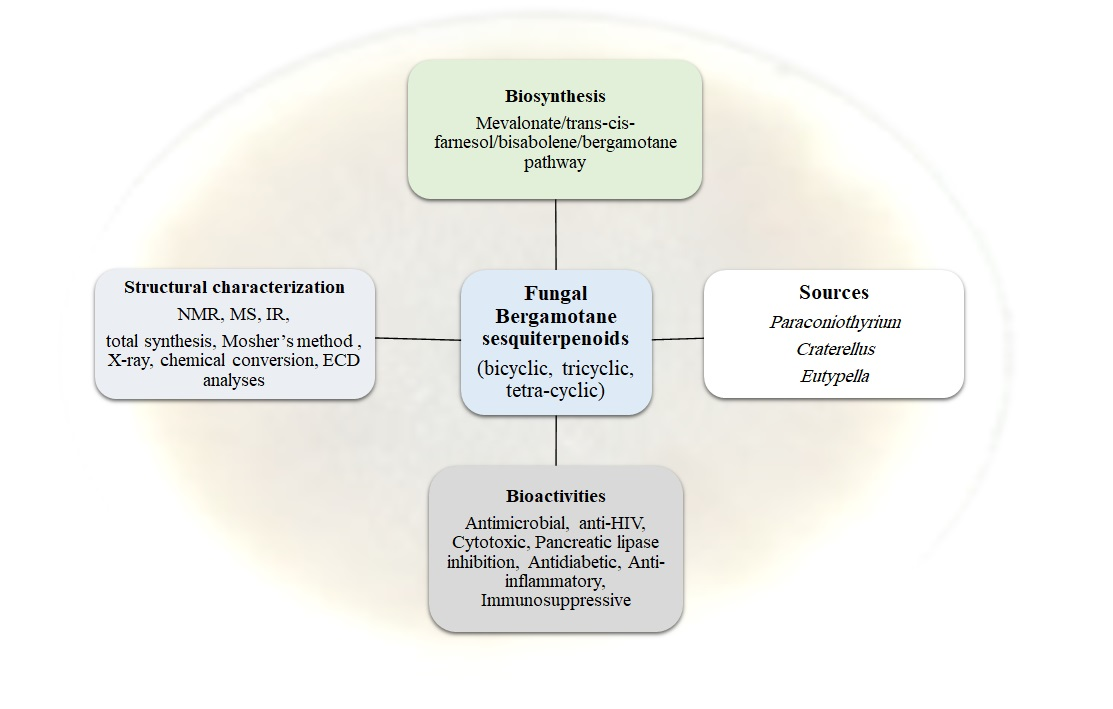
1. Introduction
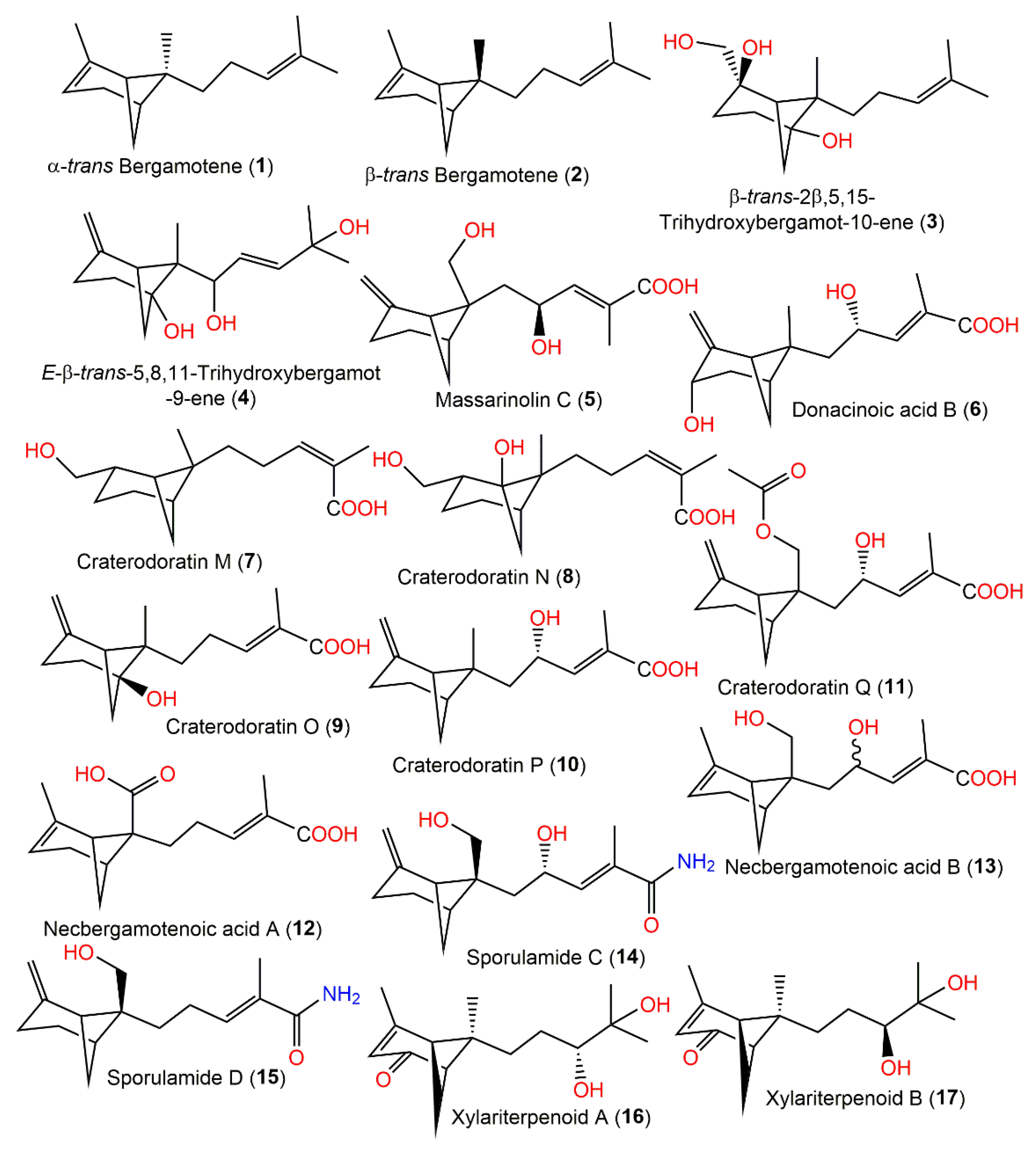
2. Structural Assignment and Stereochemistry Determination
3. Biological Activities of Bergamotane Sesquiterpenoids
3.1. Anti-Inflammatory Activity
3.2. Phytotoxic Activity
3.3. Anti-HIV Activity
3.4. Immunosuppressive Activity
3.5. Antimicrobial Activity
3.6. Pancreatic Lipase Inhibition
3.7. Antidiabetic Activity
3.8. Plant Growth Regulation
3.9. Cytotoxic Activity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 5-Aza | 5-azacytidine |
| A2780 | Human ovarian cancer cell line |
| 4-MUO | 4-methylumbelliferyl oleate |
| A549 | Lung adenocarcinoma epithelial cell line |
| Bel-7402 | Human hepatoma cell line |
| BGC-823 | Human stomach cancer cell line |
| BuOH | n-Butanol |
| C8166 | Human T-cell leukaemia |
| CC | Column chromatography |
| CC50 | The 50% cytotoxic concentration |
| CCK-8 | Cell Counting Kit-8 |
| CHCl3 | Chloroform |
| CH2Cl2 | Dichloromethane |
| CNE1-LMP1 | Stable oncoprotein LMP1 integrated nasopharyngeal carcinoma cell line |
| DU145 | Human prostate carcinoma cell line |
| EC109 | Human esophageal cancer cell line |
| ED50 | Half-maximal effective concentration |
| H2SO4 | Sulfuric acid |
| Hep3B-2 | Human hepatoma carcinoma cell line |
| HCT-116 | Human colon cancer cell line |
| HIV | Human immunodeficiency virus |
| HPLC | High-performance liquid chromatography |
| Huh7 | Human hepatoma adenocarcinoma cell line |
| IR | Infrared |
| HL-60 | Human myeloid leukemia cell line |
| LPS | Lipopolysaccharide |
| M14 | Human melanoma cell line |
| MCF-7 | Human breast cancer cell line |
| MGC | Human gastric cancer cell line |
| MTS | (3-(4,5-Dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazoliuminner salt) |
| MTT | 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| NMR | Nuclear magnetic resonance |
| NO | Nitric oxide |
| RP-18 | Reversed phase-18 |
| SBHA | Histonedeacetylase inhibitor, suberohydroxamic acid |
| SiO2 | Silica gel |
| SMMC-7721 | Hepatocellular carcinoma cell line |
| SW480 | Colon cancer cell line |
| SW1990 | Human pancreatic adenocarcinoma cell line |
| PANC-1 | Human pancreatic carcinoma cell line |
| PC-3 | Human prostate cancer cell line |
| TLC | Thin-layer chromatography |
| U937 | Human leukemic monocyte lymphoma cell line |
References
- Shen, B. A New Golden Age of Natural Products Drug Discovery. Cell 2015, 163, 1297–1300. [Google Scholar] [CrossRef] [PubMed]
- Krabberød, A.K.; Deutschmann, I.M.; Bjorbækmo, M.F.; Balagué, V.; Giner, C.R.; Ferrera, I.; Garcés, E.; Massana, R.; Gasol, J.M.; Logares, R. Long-Term Patterns of an Interconnected Core Marine Microbiota. Environ. Microbiome 2022, 17, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Imhoff, J.F. Natural Products from Marine fungi—Still an Underrepresented Resource. Mar. Drugs 2016, 14, 19. [Google Scholar] [CrossRef] [PubMed]
- Alves, A.; Sousa, E.; Kijjoa, A.; Pinto, M. Marine-Derived Compounds with Potential use as Cosmeceuticals and Nutricosmetics. Molecules 2020, 25, 2536. [Google Scholar] [CrossRef]
- Ibrahim, S.R.; Fadil, S.A.; Fadil, H.A.; Hareeri, R.H.; Alolayan, S.O.; Abdallah, H.M.; Mohamed, G.A. Dactylospongia Elegans—A Promising Drug Source: Metabolites, Bioactivities, Biosynthesis, Synthesis, and Structural-Activity Relationship. Mar. Drugs 2022, 20, 221. [Google Scholar] [CrossRef]
- Ibrahim, S.R.; Fadil, S.A.; Fadil, H.A.; Hareeri, R.H.; Abdallah, H.M.; Mohamed, G.A. Genus Smenospongia: Untapped Treasure of Biometabolites—Biosynthesis, Synthesis, and Bioactivities. Molecules 2022, 27, 5969. [Google Scholar] [CrossRef]
- Mohamed, G.A.; Ibrahim, S.R. Untapped Potential of Marine-Associated Cladosporium Species: An Overview on Secondary Metabolites, Biotechnological Relevance, and Biological Activities. Mar. Drugs 2021, 19, 645. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Mohamed, G.A.; Al Haidari, R.A.; El-Kholy, A.A.; Zayed, M.F.; Khayat, M.T. Biologically Active Fungal Depsidones: Chemistry, Biosynthesis, Structural Characterization, and Bioactivities. Fitoterapia 2018, 129, 317–365. [Google Scholar] [CrossRef]
- Omar, A.M.; Mohamed, G.A.; Ibrahim, S.R.M. Chaetomugilins and Chaetoviridins-Promising Natural Metabolites: Structures, Separation, Characterization, Biosynthesis, Bioactivities, Molecular Docking, and Molecular Dynamics. J. Fungi 2022, 8, 127. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Sirwi, A.; Eid, B.G.; Mohamed, S.G.A.; Mohamed, G.A. Fungal Depsides-Naturally Inspiring Molecules: Biosynthesis, Structural Characterization, and Biological Activities. Metabolites 2021, 11, 683. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Fadil, S.A.; Fadil, H.A.; Eshmawi, B.A.; Mohamed, S.G.A.; Mohamed, G.A. Fungal Naphthalenones; Promising Metabolites for Drug Discovery: Structures, Biosynthesis, Sources, and Pharmacological Potential. Toxins 2022, 14, 154. [Google Scholar] [CrossRef] [PubMed]
- Ameen, F.; AlNadhari, S.; Al-Homaidan, A.A. Marine Microorganisms as an Untapped Source of Bioactive Compounds. Saudi J. Biol. Sci. 2021, 28, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Hasan, S.; Ansari, M.I.; Ahmad, A.; Mishra, M. Major Bioactive Metabolites from Marine Fungi: A Review. Bioinformation 2015, 11, 176. [Google Scholar] [CrossRef] [PubMed]
- Wibowo, J.T.; Ahmadi, P.; Rahmawati, S.I.; Bayu, A.; Putra, M.Y.; Kijjoa, A. Marine-Derived Indole Alkaloids and their Biological and Pharmacological Activities. Mar. Drugs 2021, 20, 3. [Google Scholar] [CrossRef]
- Giddings, L.; Newman, D.J. Extremophilic Fungi from Marine Environments: Underexplored Sources of Antitumor, Anti-Infective and Other Biologically Active Agents. Mar. Drugs 2022, 20, 62. [Google Scholar] [CrossRef]
- Hafez Ghoran, S.; Taktaz, F.; Ayatollahi, S.A.; Kijjoa, A. Anthraquinones and their Analogues from Marine-Derived Fungi: Chemistry and Biological Activities. Mar. Drugs 2022, 20, 474. [Google Scholar] [CrossRef]
- Kramer, R.; Abraham, W. Volatile Sesquiterpenes from Fungi: What are they Good for? Phytochem. Rev. 2012, 11, 15–37. [Google Scholar] [CrossRef]
- Gui, P.; Fan, J.; Zhu, T.; Fu, P.; Hong, K.; Zhu, W. Sesquiterpenoids from the Mangrove-Derived Aspergillus Ustus 094102. Mar. Drugs 2022, 20, 408. [Google Scholar] [CrossRef]
- Dai, Q.; Zhang, F.; Feng, T. Sesquiterpenoids Specially Produced by Fungi: Structures, Biological Activities, Chemical and Biosynthesis (2015–2020). J. Fungi 2021, 7, 1026. [Google Scholar] [CrossRef]
- Minami, A.; Ozaki, T.; Liu, C.; Oikawa, H. Cyclopentane-Forming Di/Sesterterpene Synthases: Widely Distributed Enzymes in Bacteria, Fungi, and Plants. Nat. Prod. Rep. 2018, 35, 1330–1346. [Google Scholar] [CrossRef]
- Fraga, B.M. Natural Sesquiterpenoids. Nat. Prod. Rep. 2013, 30, 1226–1264. [Google Scholar] [CrossRef] [PubMed]
- Cane, D.E. Enzymic Formation of Sesquiterpenes. Chem. Rev. 1990, 90, 1089–1103. [Google Scholar] [CrossRef]
- Oh, H.; Gloer, J.B.; Shearer, C.A. Massarinolins A−C: New Bioactive Sesquiterpenoids from the Aquatic Fungus Massarina Tunicata. J. Nat. Prod. 1999, 62, 497–501. [Google Scholar] [CrossRef] [PubMed]
- Che, Y.; Gloer, J.B.; Koster, B.; Malloch, D. Decipinin A and Decipienolides A and B: New Bioactive Metabolites from the Coprophilous Fungus Podospora Decipiens. J. Nat. Prod. 2002, 65, 916–919. [Google Scholar] [CrossRef]
- Massias, M.; Rebuffat, S.; Molho, L.; Chiaroni, A.; Riche, C.; Bodo, B. Expansolides A and B: Tetracyclic Sesquiterpene Lactones from Penicillium Expansum. J. Am. Chem. Soc. 1990, 112, 8112–8115. [Google Scholar] [CrossRef]
- Macías, F.A.; Varela, R.M.; Simonet, A.M.; Cutler, H.G.; Cutler, S.J.; Hill, R.A. Absolute Configuration of Bioactive Expansolides A and B from Aspergillus Fumigatus Fresenius. Tetrahedron Lett. 2003, 44, 941–943. [Google Scholar] [CrossRef]
- Wen, Y.; Chen, T.; Jiang, L.; Li, L.; Guo, M.; Peng, Y.; Chen, J.; Pei, F.; Yang, J.; Wang, R. Unusual (2R, 6R)-Bicyclo [3.1. 1] Heptane Ring Construction in Fungal A-Trans-Bergamotene Biosynthesis. Iscience 2022, 25, 104030. [Google Scholar] [CrossRef]
- Nozoe, S.; Kobayashi, H.; Morisaki, N. Isolation of Beta Trans Bergamotene from Aspergillus Fumigatus, a Fumagillin Producing Fungi. Tetrahedron Lett. 1976, 50, 4625–4626. [Google Scholar] [CrossRef]
- Wang, Y.; Li, D.; Li, Z.; Sun, Y.; Hua, H.; Liu, T.; Bai, J. Terpenoids from the Marine-Derived Fungus Aspergillus Fumigatus YK-7. Molecules 2016, 21, 31. [Google Scholar] [CrossRef]
- Dai, Q.; Zhang, F.; Li, Z.; He, J.; Feng, T. Immunosuppressive Sesquiterpenoids from the Edible Mushroom Craterellus Odoratus. J. Fungi 2021, 7, 1052. [Google Scholar] [CrossRef]
- Pei, Y.; Zhang, L.; Wu, X.; Wu, H.; Wang, H.; Wang, Y.; Chen, G. Polyhydroxylated Bergamotane-Type Sesquiterpenoids from Cultures of Paraconiothyrium Sporulosum YK-03 and their Absolute Configurations. Phytochemistry 2022, 194, 113000. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Zhao, K.; Chen, H.; Bai, X.; Zhang, L.; Liu, J. Terpenoids from the Mushroom-Associated Fungus Montagnula Donacina. Phytochemistry 2018, 147, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Wu, Y.; Chen, G.; Hu, D.; Li, X.; Sun, X.; Guo, L.; Li, Y.; Yao, X.; Gao, H. Xylariterpenoids A–D, Four New Sesquiterpenoids from the Xylariaceae Fungus. RSC Adv. 2014, 4, 54144–54148. [Google Scholar] [CrossRef]
- Niu, S.; Xie, C.; Zhong, T.; Xu, W.; Luo, Z.; Shao, Z.; Yang, X. Sesquiterpenes from a Deep-Sea-Derived Fungus Graphostroma Sp. MCCC 3A00421. Tetrahedron 2017, 73, 7267–7273. [Google Scholar] [CrossRef]
- Niu, S.; Liu, D.; Shao, Z.; Liu, J.; Fan, A.; Lin, W. Chemical Epigenetic Manipulation Triggers the Production of Sesquiterpenes from the Deep-Sea Derived Eutypella Fungus. Phytochemistry 2021, 192, 112978. [Google Scholar] [CrossRef]
- Pena-Rodriguez, L.M.; Armingeon, N.A.; Chilton, W.S. Toxins from Weed Pathogens, I. Phytotoxins from a Bipolaris Pathogen of Johnson Grass. J. Nat. Prod. 1988, 51, 821–828. [Google Scholar] [CrossRef]
- Pena-Rodriguez, L.M.; Chilton, W.S. Victoxinine and Prehelminthosporolactone, Two Minor Phytotoxic Metabolites Produced by Bipolaris Sp., a Pathogen of Johnson Grass. J. Nat. Prod. 1989, 52, 899–901. [Google Scholar] [CrossRef]
- Pringle, R.B.; Braun, A.C. Constitution of the Toxin of Helminthosporium Victoriae. Nature 1958, 181, 1205–1206. [Google Scholar] [CrossRef]
- Dorn, F.; Arigoni, D. Structure of Victoxinine. J. Chem. Soc. Chem. Commun. 1972, 24, 1342–1343. [Google Scholar] [CrossRef]
- Pringle, R. Comparative Biochemistry of the Phytopathogenic Fungus Helminthosporium. XVI. the Production of Victoxinine by H. Sativum and H. Victoriae. Can. J. Biochem. 1976, 54, 783–787. [Google Scholar] [CrossRef]
- Kono, Y.; Takeuchi, S.; Daly, J.M. Isolation and Structure of a New Victoxinine Derivative Produced by Helminthosporium Victoriae. Agric. Biol. Chem. 1983, 47, 2701–2702. [Google Scholar] [CrossRef]
- Kimura, Y.; Nakajima, H.; Hamasaki, T.; Sugawara, F.; Parkanyi, L.; Clardy, J. Pinthunamide, a New Tricyclic Sesquiterpene Amide Produced by a Fungus, Ampullifernia Sp. Tetrahedron Lett. 1989, 30, 1267–1270. [Google Scholar] [CrossRef]
- Liu, L.; Gao, H.; Chen, X.; Cai, X.; Yang, L.; Guo, L.; Yao, X.; Che, Y. Brasilamides A–D: Sesquiterpenoids from the Plant Endophytic Fungus Paraconiothyrium Brasiliense. Eur. J. Org. Chem. 2010, 3302–3306. [Google Scholar] [CrossRef]
- Guo, Z.; Ren, F.; Che, Y.; Liu, G.; Liu, L. New Bergamotane Sesquiterpenoids from the Plant Endophytic Fungus Paraconiothyrium Brasiliense. Molecules 2015, 20, 14611–14620. [Google Scholar] [CrossRef]
- Wang, W.; Shi, Y.; Liu, Y.; Zhang, Y.; Wu, J.; Zhang, G.; Che, Q.; Zhu, T.; Li, M.; Li, D. Brasilterpenes A-E, Bergamotane Sesquiterpenoid Derivatives with Hypoglycemic Activity from the Deep Sea-Derived Fungus Paraconiothyrium Brasiliense HDN15-135. Mar Drugs 2022, 20, 514. [Google Scholar] [CrossRef]
- Ying, Y.; Fang, C.; Yao, F.; Yu, Y.; Shen, Y.; Hou, Z.; Wang, Z.; Zhang, W.; Shan, W.; Zhan, Z. Bergamotane Sesquiterpenes with Alpha-glucosidase Inhibitory Activity from the Plant Pathogenic Fungus Penicillium Expansum. Chem. Biodivers. 2017, 14, e1600184. [Google Scholar] [CrossRef]
- Wang, Y.; Xia, G.; Wang, L.; Ge, G.; Zhang, H.; Zhang, J.; Wu, Y.; Lin, S. Purpurolide A, 5/5/5 Spirocyclic Sesquiterpene Lactone in Nature from the Endophytic Fungus Penicillium Purpurogenum. Org. Lett. 2018, 20, 7341–7344. [Google Scholar] [CrossRef]
- Xia, G.; Wang, L.; Zhang, J.; Wu, Y.; Ge, G.; Wang, Y.; Lin, P.; Lin, S. Three New Polyoxygenated Bergamotanes from the Endophytic Fungus Penicillium Purpurogenum IMM 003 and their Inhibitory Activity Against Pancreatic Lipase. Chin. J. Nat. Med. 2020, 18, 75–80. [Google Scholar] [CrossRef]
- Zhang, L.; Feng, B.; Chen, G.; Li, S.; Sun, Y.; Wu, H.; Bai, J.; Hua, H.; Wang, H.; Pei, Y. Sporulaminals A and B: A Pair of Unusual Epimeric Spiroaminal Derivatives from a Marine-Derived Fungus Paraconiothyrium Sporulosum YK-03. RSC Adv. 2016, 6, 42361–42366. [Google Scholar] [CrossRef]
- Kimura, Y.; Nakajima, H.; Hamasaki, T.; Matsumoto, T.; Matsuda, Y.; Tsuneda, A. Ampullicin and Isoampullicin, New Metabolites from an Ampulliferina-Like Fungus Sp. no. 27. Agric. Biol. Chem. 1990, 54, 813–814. [Google Scholar] [CrossRef]
- Kimura, Y.; Matsumoto, T.; Nakajima, H.; Hamasaki, T.; Matsuda, Y. Dihydroampullicin, a New Plant Growth Regulators Produced by the Ampulliferina-Like Fungus Sp. no. 27. Biosci. Biotechnol. Biochem. 1993, 57, 687–688. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, H.; Xu, W.; Hu, B.; Guild, A.; Zhang, J.; Lu, X.; Liu, X.; Jiao, B. Eutypellacytosporins A–D, Meroterpenoids from the Arctic Fungus Eutypella Sp. D-1. J. Nat. Prod. 2019, 82, 3089–3095. [Google Scholar] [CrossRef] [PubMed]
- Bermejo, F.A.; Mateos, A.F.; Escribano, A.M.; Lago, R.M.; Burón, L.M.; López, M.R.; González, R.R. Ti (III)-Promoted Cyclizations. Application to the Synthesis of (E)-Endo-Bergamoten-12-Oic Acids. Moth Oviposition Stimulants Isolated from Lycopersicon Hirsutum. Tetrahedron 2006, 62, 8933–8942. [Google Scholar] [CrossRef]
- López, M.R.; Bermejo, F.A. Total Synthesis of ( )-Massarinolin B and ( )-4-Epi-Massarinolin B, Fungal Metabolites from Massarina Tunicata. Tetrahedron 2006, 62, 8095–8102. [Google Scholar] [CrossRef]
- Zhao, J.; Feng, J.; Tan, Z.; Liu, J.; Zhang, M.; Chen, R.; Xie, K.; Chen, D.; Li, Y.; Chen, X. Bistachybotrysins A-C, Three Phenylspirodrimane Dimers with Cytotoxicity from Stachybotrys Chartarum. Bioorg. Med. Chem. Lett. 2018, 28, 355–359. [Google Scholar] [CrossRef]
- Zamora, R.; Vodovotz, Y.; Billiar, T.R. Inducible Nitric Oxide Synthase and Inflammatory Diseases. Mol. Med. 2000, 6, 347–373. [Google Scholar] [CrossRef]
- Kovarik, J.M.; Burtin, P. Immunosuppressants in Advanced Clinical Development for Organ Transplantation and Selected Autoimmune Diseases. Expert Opin. Emerg. Drugs 2003, 8, 47–62. [Google Scholar] [CrossRef]
- Klinker, M.W.; Lundy, S.K. Multiple Mechanisms of Immune Suppression by B Lymphocytes. Mol. Med. 2012, 18, 123–137. [Google Scholar] [CrossRef]
- Bermejo, F.A.; Rico-Ferreira, R.; Bamidele-Sanni, S.; García-Granda, S. Total Synthesis of (+)-Ampullicin and (+)-Isoampullicin: Two Fungal Metabolites with Growth Regulatory Activity Isolated from Ampulliferina Sp. 27. J. Org. Chem. 2001, 66, 8. [Google Scholar] [CrossRef]



















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
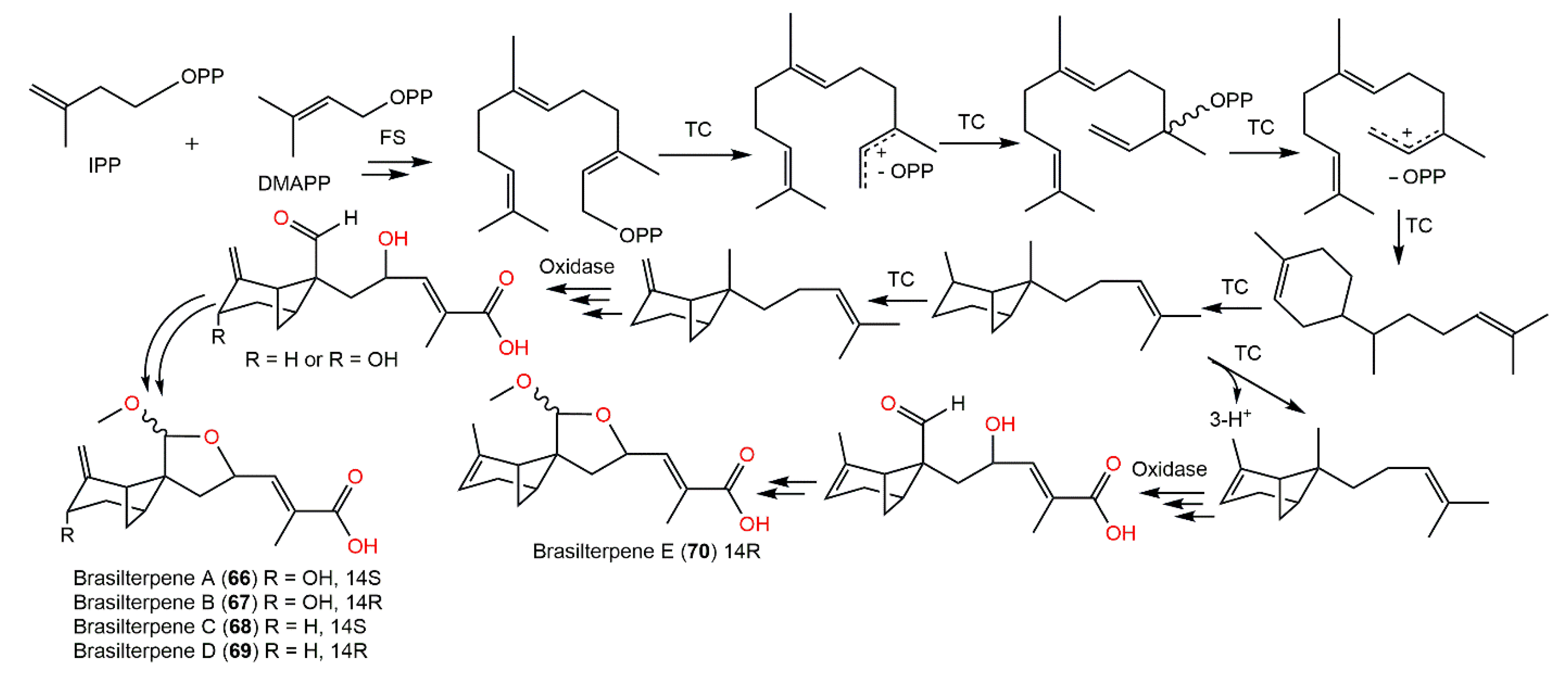{kind=link}
{kind=link}
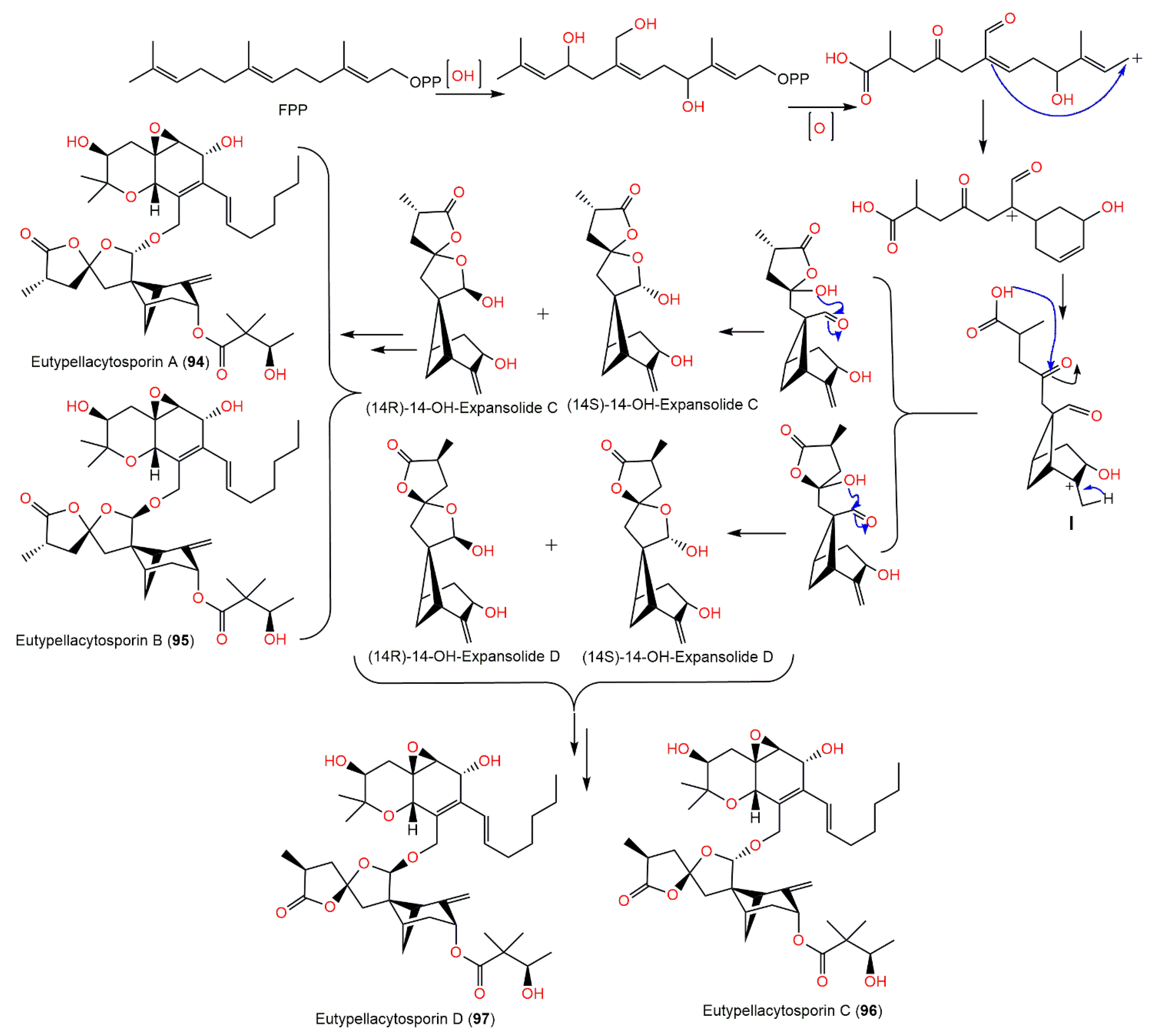{kind=link}
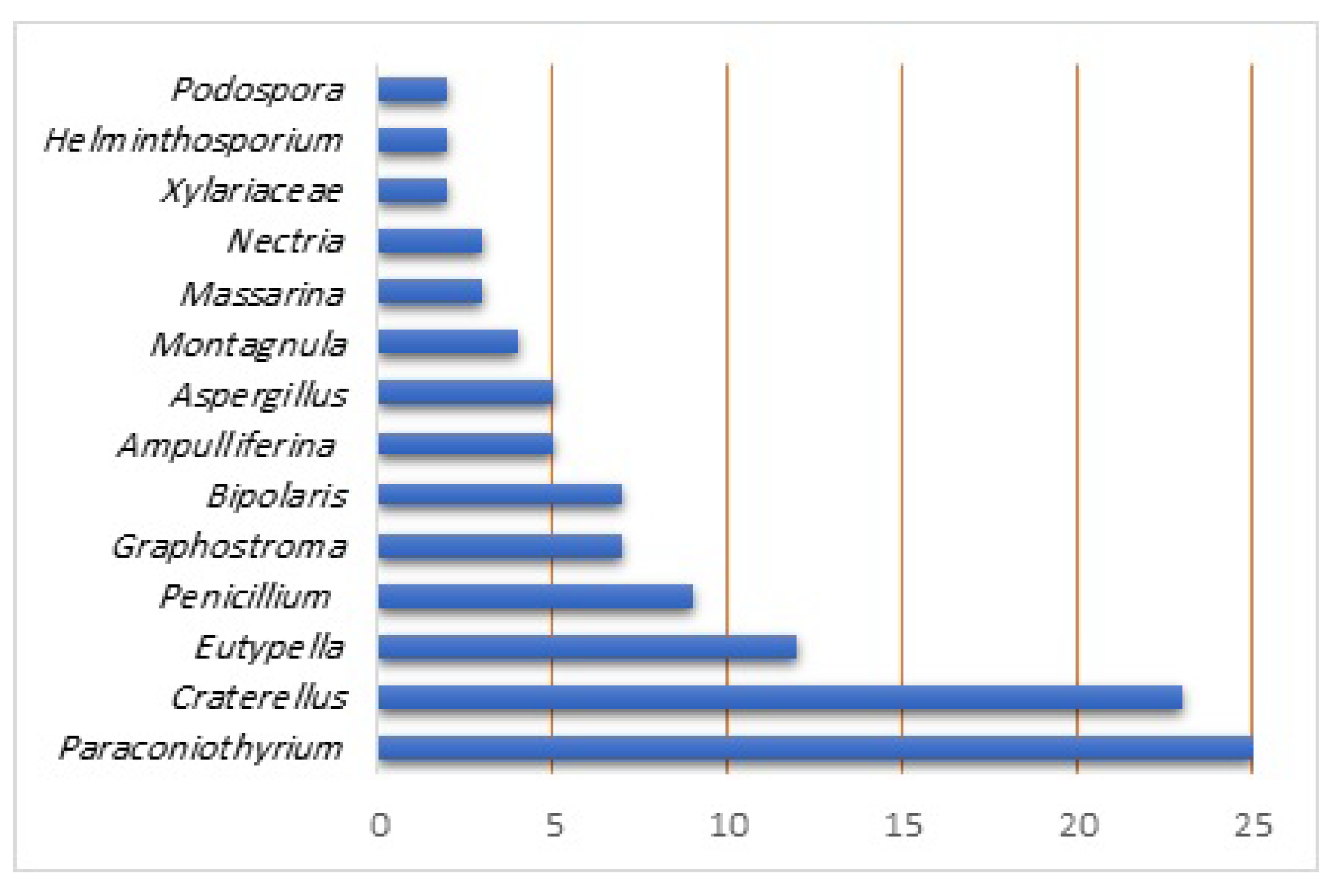
| Compound Name | Fungal Source/Host | Extract/Fraction | Mol. Wt. | Mol. Formula | Location | Ref. |
|---|---|---|---|---|---|---|
| Bicyclic Bergamotane Sesquiterpenoids | ||||||
| α-trans Bergamotene (1) | Nectria sp. HLS206 (Nectriaceae)/Gelliodas carnosa (marine sponge Geodiidae) | EtOAc extract | 204 | C15H24 | China | [27] |
| β-trans Bergamotene (2) | Aspergillus fumigatus (Trichocomaceae)/Cultured | Acetone extract | 204 | C15H24 | Japan | [28] |
| β-trans-2β,5,15-Trihydroxybergamot-10-ene (3) | Aspergillus fumigatus YK-7 (Trichocomaceae)/Sea mud | EtOAc extract | 254 | C15H26O3 | Intertidal zone sea mud, Yingkou, China | [29] |
| E-β-trans-5,8,11-Trihydroxybergamot-9-ene (4) | Aspergillus fumigatus YK-7 (Trichocomaceae)/Sea mud | EtOAc extract | 252 | C15H24O3 | Intertidal zone sea mud, Yingkou, China | [29] |
| Massarinolin C (5) | Massarina tunicata (Lophiostomataceae)/Submerged twig | EtOAc extract | 266 | C15H22O4 | Lemonweir River in Adams County, Wisconsin, USA | [23] |
| Craterellus odoratus (Cantharellaceae) | EtOAc extract | - | - | Southern part of the GaoligongMountains, Yunnan, China | [30] | |
| Paraconiothyrium sporulosum YK-03 Verkley (Leptosphaeriaceae)/Sea mud | EtOAc extract | - | - | Intertidal zone of Bohai Bay river in Liaoning, China | [31] | |
| Donacinoic acid B (6) | Montagnula donacina (Montagnulaceae)/Craterellus odoratus (fruiting bodies, Cantharellaceae) | EtOAc extract | 266 | C15H22O4 | Southern part of the Gaoligong Mountains in Yunnan, China | [32] |
| Craterodoratin M (7) | Craterellus odoratus (Cantharellaceae) | EtOAc extract | 252 | C15H24O3 | Southern part of the GaoligongMountains, Yunnan, China | [30] |
| Craterodoratin N (8) | Craterellus odoratus (Cantharellaceae) | EtOAc extract | 268 | C15H24O4 | Southern part of the GaoligongMountains, Yunnan, China | [30] |
| Craterodoratin O (9) | Craterellus odoratus (Cantharellaceae) | EtOAc extract | 250 | C15H22O3 | Southern part of the GaoligongMountains, Yunnan, China | [30] |
| Craterodoratin P (10) | Craterellus odoratus (Cantharellaceae) | EtOAc extract | 250 | C15H22O3 | Southern part of the GaoligongMountains, Yunnan, China | [30] |
| Craterodoratin Q (11) | Craterellus odoratus (Cantharellaceae) | EtOAc extract | 308 | C17H24O5 | Southern part of the GaoligongMountains, Yunnan, China | [30] |
| Necbergamotenoic acid A (12) | Nectria sp. HLS206 (Nectriaceae)/Gelliodas carnosa (marine sponge, Geodiidae) | EtOAc extract | 264 | C15H20O4 | China | [27] |
| Necbergamotenoic acid B (13) | Nectria sp. HLS206 (Nectriaceae)/Gelliodas carnosa (marine sponge, Geodiidae) | EtOAc extract | 266 | C15H22O4 | China | [27] |
| Sporulamide C (14) | Paraconiothyrium sporulosum YK-03 Verkley (Leptosphaeriaceae)/Sea mud | EtOAc extract | 265 | C15H23NO3 | Intertidal zone of Bohai Bay river in Liaoning, China | [31] |
| Sporulamide D (15) | Paraconiothyrium sporulosum YK-03 Verkley (Leptosphaeriaceae)/Sea mud | EtOAc extract | 249 | C15H23NO2 | Intertidal zone of Bohai Bay river in Liaoning, China | [31] |
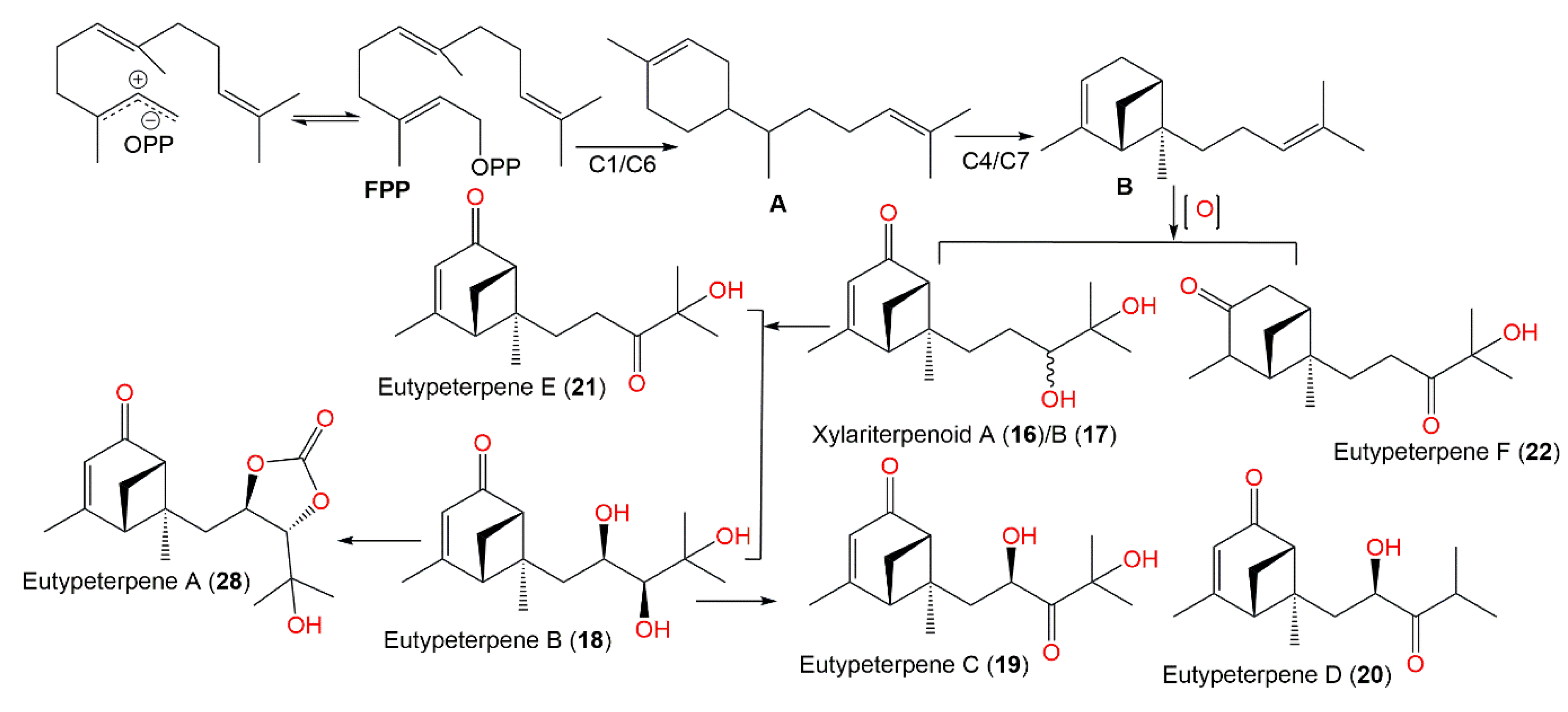
| Xylariterpenoid A (16) | Xylariaceae fungus (No. 63-19-7-3)/Everniastrum cirrhatum (Fr.) Haleex Sipman (lichen, Parmeliaceae) | EtOAc extract | 252 | C15H24O3 | Zixi Mountain, Yunnan, China | [33] |
| Graphostroma sp. MCCC 3A00421/Deep-sea hydrothermal sulfide deposit | EtOAc extract | - | - | Atlantic Ocean, China | [34] | |
| Eutypella sp. MCCC 3A00281 (Diatrypaceae)/Deep-sea sediment | EtOAc extract | - | - | South Atlantic Ocean, China | [35] | |
| Xylariterpenoid B (17) | Xylariaceae fungus (No. 63-19-7-3)/Everniastrum cirrhatum (Fr.) Haleex Sipman (lichen, Parmeliaceae) | EtOAc extract | 252 | C15H24O3 | Zixi Mountain, Yunnan, China | [33] |
| Graphostroma sp. MCCC 3A00421/Deep-sea hydrothermal sulfide deposit | EtOAc extract | - | - | Atlantic Ocean, China | [34] | |
| Eutypella sp. MCCC 3A00281 (Diatrypaceae)/Deep-sea sediment | EtOAc extract | - | - | South Atlantic Ocean, China | [35] | |
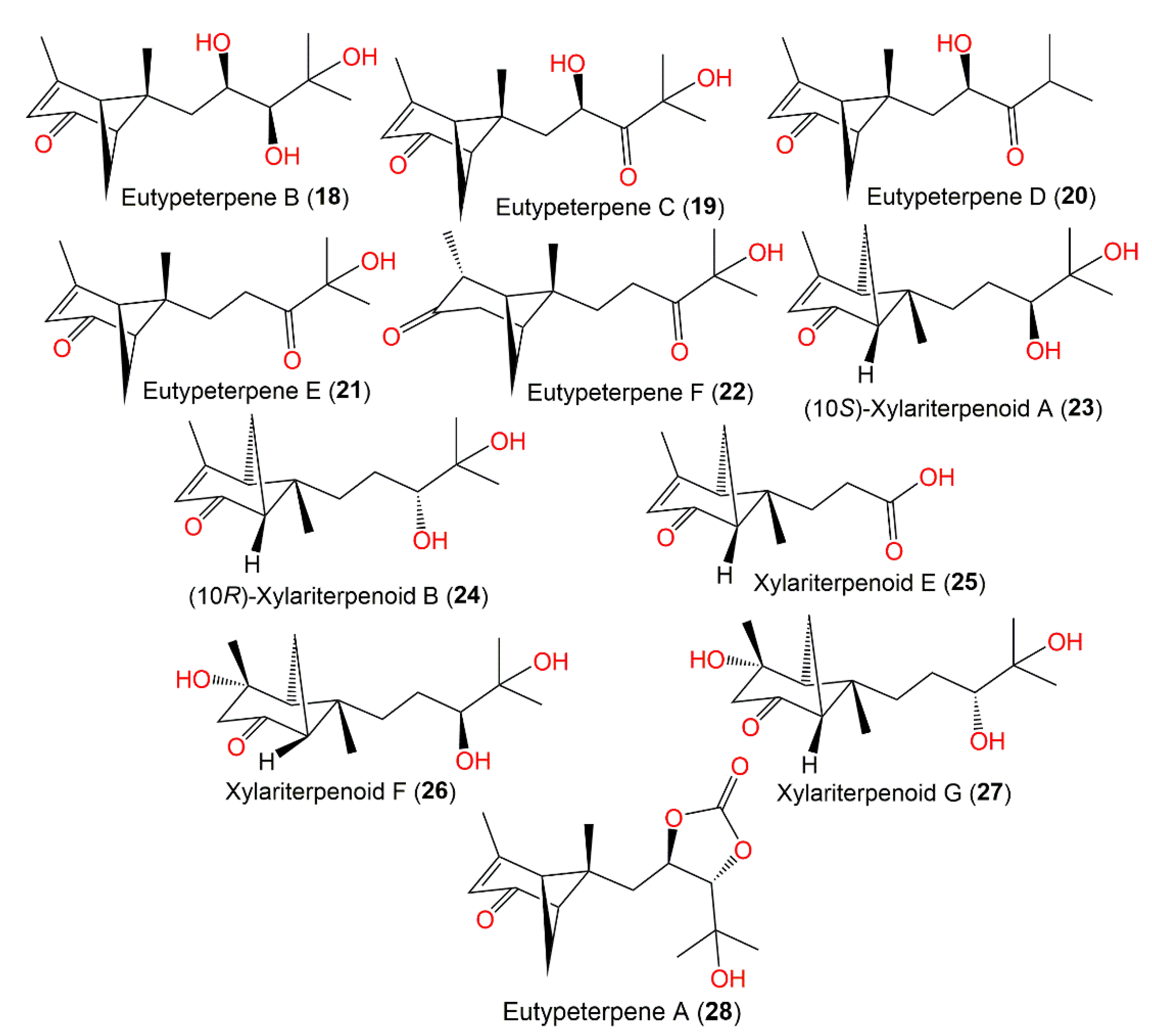
| Eutypeterpene B (18) | Eutypella sp. MCCC 3A00281 (Diatrypaceae)/Deep-sea sediment | EtOAc extract | 268 | C15H24O4 | South Atlantic Ocean, China | [35] |
| Eutypeterpene C (19) | Eutypella sp. MCCC 3A00281 (Diatrypaceae)/Deep-sea sediment | EtOAc extract | 266 | C15H22O4 | South Atlantic Ocean, China | [35] |
| Eutypeterpene D (20) | Eutypella sp. MCCC 3A00281 (Diatrypaceae)/Deep-sea sediment | EtOAc extract | 250 | C15H22O3 | South Atlantic Ocean, China | [35] |
| Eutypeterpene E (21) | Eutypella sp. MCCC 3A00281 (Diatrypaceae)/Deep-sea sediment | EtOAc extract | 250 | C15H22O3 | South Atlantic Ocean, China | [35] |
| Eutypeterpene F (22) | Eutypella sp. MCCC 3A00281 (Diatrypaceae)/Deep-sea sediment | EtOAc extract | 252 | C15H24O3 | South Atlantic Ocean, China | [35] |
| (10S)-Xylariterpenoid A (23) | Graphostroma sp. MCCC 3A00421/Deep-sea hydrothermal sulfide deposit | EtOAc extract | 252 | C15H24O3 | Atlantic Ocean. China | [34] |
| (10R)-Xylariterpenoid B (24) | Graphostroma sp. MCCC 3A00421/Deep-sea hydrothermal sulfide deposit | EtOAc extract | 252 | C15H24O3 | Atlantic Ocean. China | [34] |
| Xylariterpenoid E (25) | Graphostroma sp. MCCC 3A00421/Deep-sea hydrothermal sulfide deposit | EtOAc extract | 208 | C12H16O3 | Atlantic Ocean. China | [34] |
| Xylariterpenoid F (26) | Graphostroma sp. MCCC 3A00421/Deep-sea hydrothermal sulfide deposit | EtOAc extract | 270 | C15H26O4 | Atlantic Ocean. China | [34] |
| Xylariterpenoid G (27) | Graphostroma sp. MCCC 3A00421/Deep-sea hydrothermal sulfide deposit | EtOAc extract | 270 | C15H26O4 | Atlantic Ocean. China | [34] |
| Eutypeterpene A (28) | Eutypella sp. MCCC 3A00281 (Diatrypaceae)/Deep-sea sediment | EtOAc extract | 294 | C16H22O5 | South Atlantic Ocean, China | [35] |
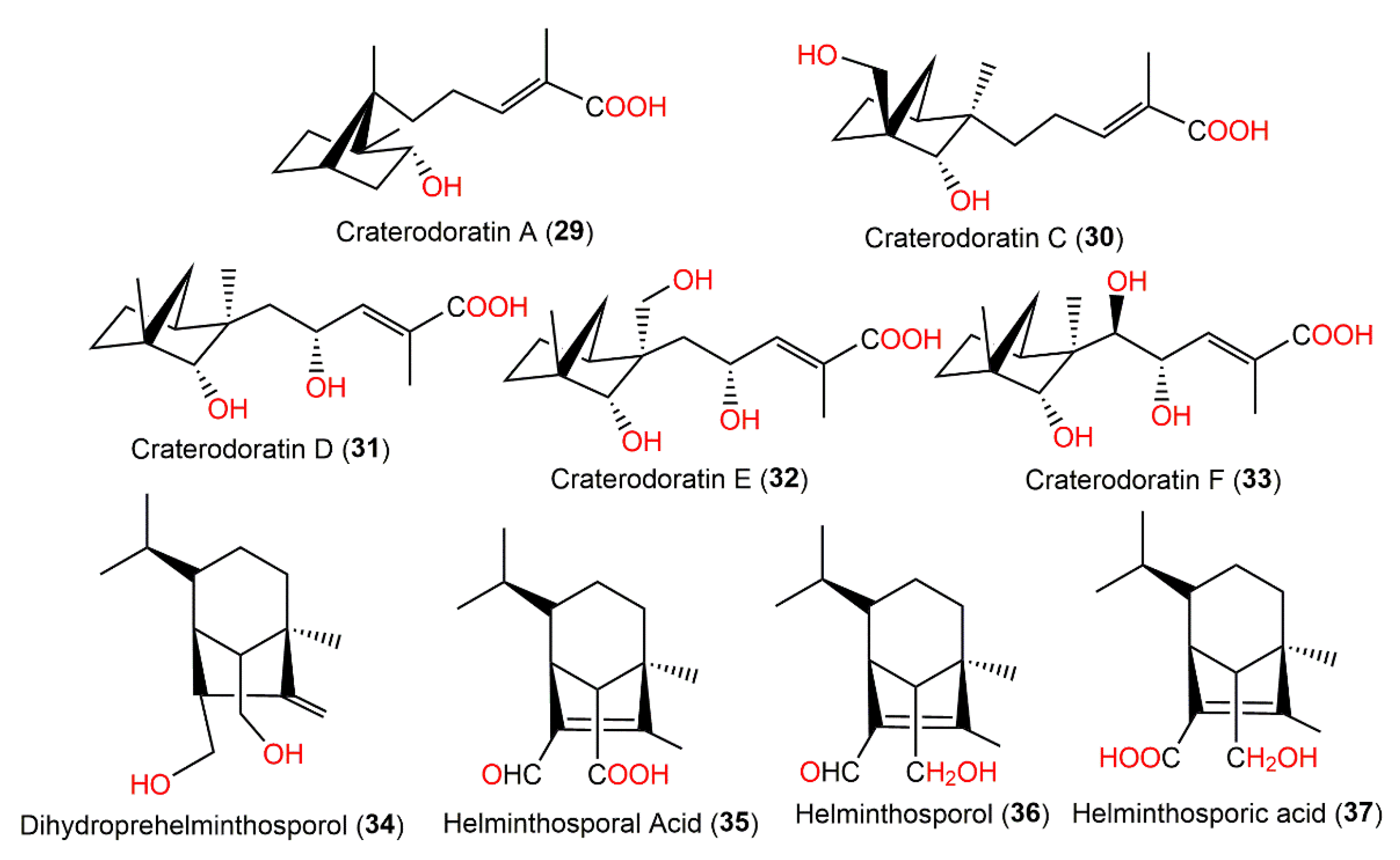
| Craterodoratin A (29) | Craterellus odoratus (Cantharellaceae) | EtOAc extract | 252 | C15H24O3 | Southern part of the GaoligongMountains, Yunnan, China | [30] |
| Craterodoratin C (30) | Craterellus odoratus (Cantharellaceae) | EtOAc extract | 268 | C15H24O4 | Southern part of the GaoligongMountains, Yunnan, China | [30] |
| Craterodoratin D (31) | Craterellus odoratus (Cantharellaceae) | EtOAc extract | 268 | C15H24O4 | Southern part of the GaoligongMountains, Yunnan, China | [30] |
| Craterodoratin E (32) | Craterellus odoratus (Cantharellaceae) | EtOAc extract | 284 | C15H24O5 | Southern part of the GaoligongMountains, Yunnan, China | [30] |
| Craterodoratin F (33) | Craterellus odoratus (Cantharellaceae) | EtOAc extract | 284 | C15H24O5 | Southern part of the GaoligongMountains, Yunnan, China | [30] |
| Dihydroprehelminthosporol (34) | Bipolaris sp. No. 36/Johnson grass leaf | EtOAc extract | 238 | C15H26O2 | Wake County, North Carolina, USA | [36,37] |
| Helminthosporal acid (35) | Bipolaris sp. No. 36/Johnson grass leaf | EtOAc extract | 250 | C15H22O3 | Wake County, North Carolina, USA | [36] |
| Helminthosporol (36) | Bipolaris sp. No. 36/Johnson grass leaf | EtOAc extract | 236 | C15H24O2 | Wake County, North Carolina, USA | [36] |
| Helminthosporic acid (37) | Bipolaris sp. No. 36/Johnson grass leaf | EtOAc extract | 252 | C15H24O3 | Wake County, North Carolina, USA | [36] |
| Tricyclic Bergamotane Sesquiterpenoids | ||||||
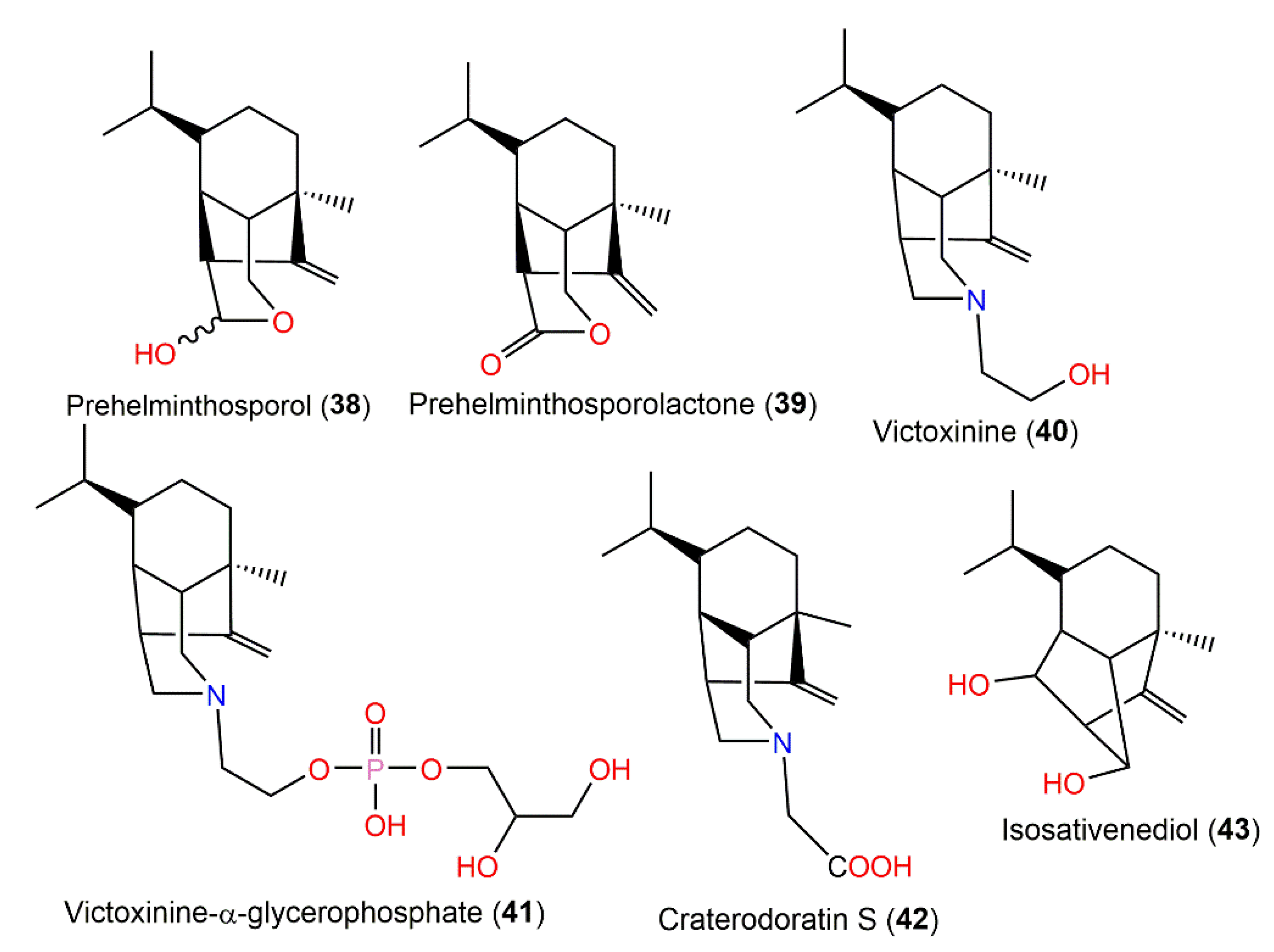
| Prehelminthosporol (38) | Bipolaris sp. No. 36/Johnson grass leaf | EtOAc extract | 236 | C15H24O2 | Wake County, North Carolina, USA | [36,37] |
| Prehelminthosporolactone (39) | Bipolaris sp. No. 36/Johnson grass leaf | EtOAc extract | 234 | C15H22O2 | Wake County, North Carolina, USA | [37] |
| Victoxinine (40) | Helminthosporium victoriae (Totiviridae) | Diethyl ether extract | 263 | C17H29NO | USA | [36,38,39] |
| Helminthosporium sativum (Totiviridae) | Diethyl ether fraction/CHCl3 extract | - | - | Canada | [40] | |
| Victoxinine-α-glycerophosphate (41) | H. sativum (Totiviridae) | n-BuOH extract | 417 | C20H36NO6P | USA | [41] |
| Craterodoratin S (42) | Craterellus odoratus (Cantharellaceae) | EtOAc extract | 277 | C17H27NO2 | Southern part of the GaoligongMountains, Yunnan, China | [30] |
| Isosativenediol (43) | Bipolaris sp. No. 36/Johnson grass leaf | EtOAc extract | 236 | C15H24O2 | Wake County, North Carolina, USA | [36] |
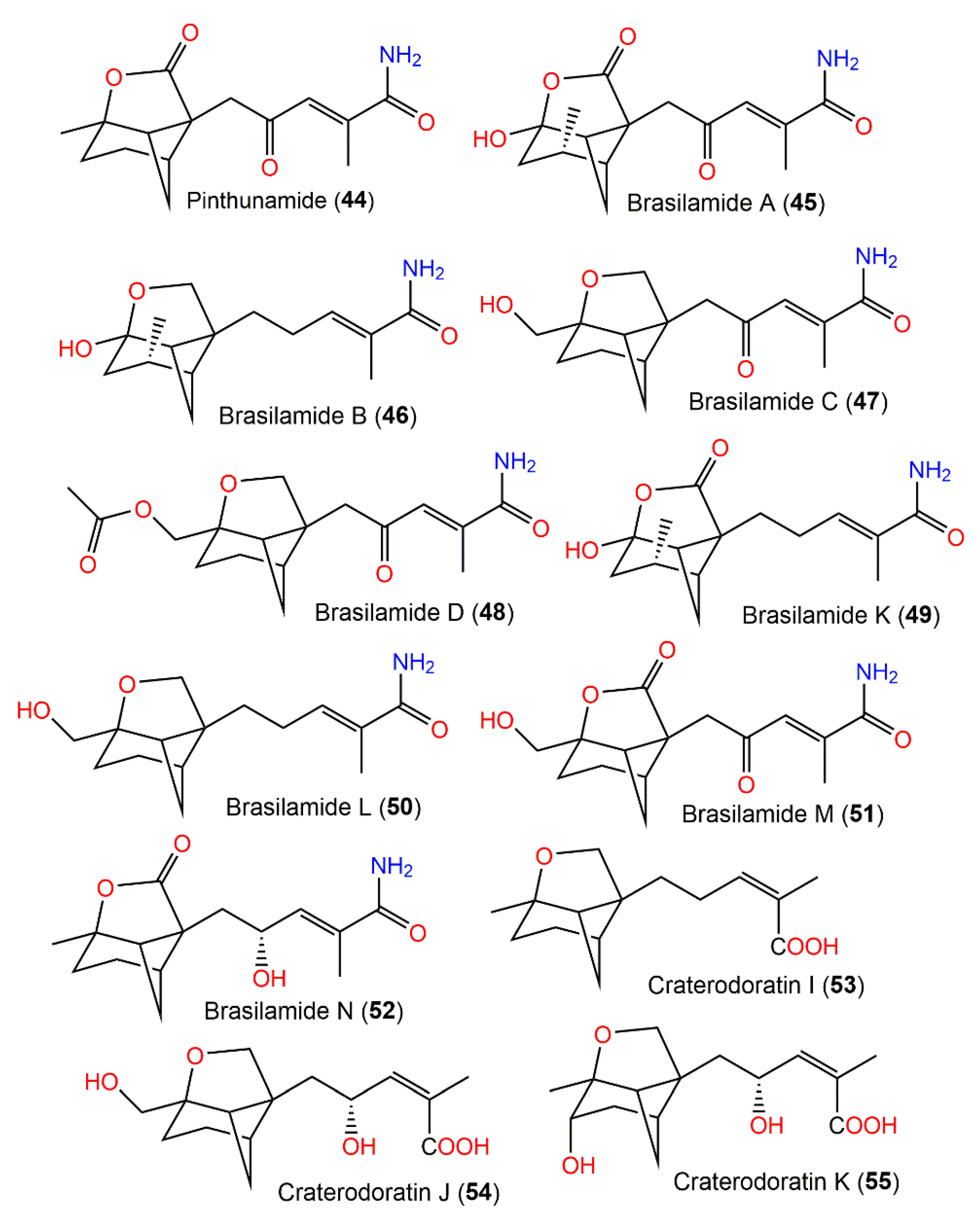
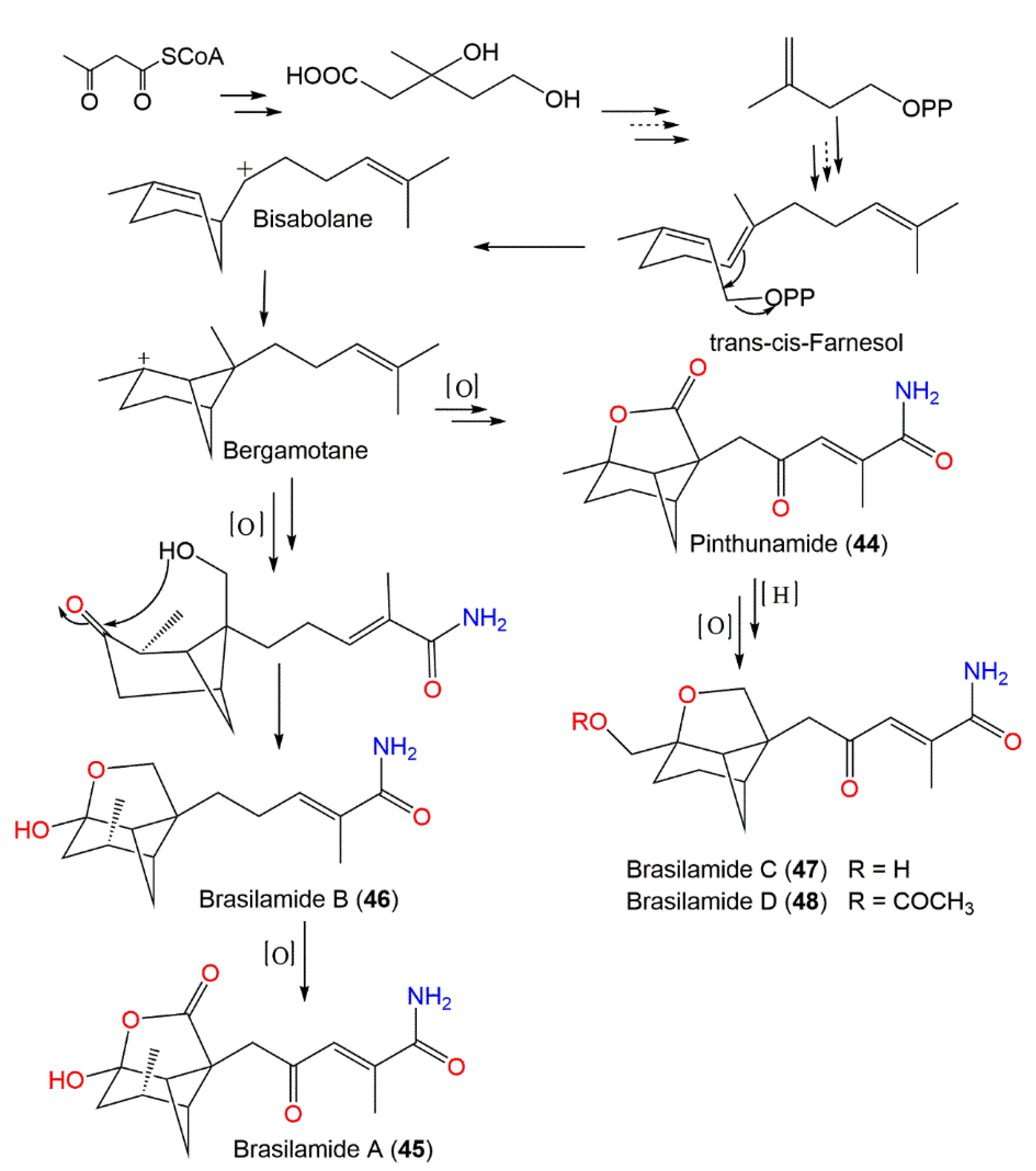
| Pinthunamide (44) | Ampulliferina sp. No. 27 (Ampullicephala)/Pinus thunbergii (dead tree, Pinaceae) | Acetone extract | 277 | C15H19NO4 | Japan | [42] |
| Paraconiothyrium brasiliense Verkley (M3–3341) (Leptosphaeriaceae)/ Acer truncatum Bunge (branches, Sapindaceae) | Acetone extract | - | - | Dongling Mountain, Beijing, China | [43] | |
| Brasilamide A (45) | Paraconiothyrium brasiliense Verkley (M3–3341) (Leptosphaeriaceae)/ Acer truncatum Bunge (branches, Sapindaceae) | Acetone extract | 293 | C15H19NO5 | Dongling Mountain, Beijing, China | [43,44] |
| Brasilamide B (46) | Paraconiothyrium brasiliense Verkley (M3–3341) (Leptosphaeriaceae)/ Acer truncatum Bunge (branches, Sapindaceae) | Acetone extract | 265 | C15H23NO3 | Dongling Mountain, Beijing, China | [43] |
| Brasilamide C (47) | Paraconiothyrium brasiliense Verkley (M3–3341) (Leptosphaeriaceae)/ Acer truncatum Bunge (branches, Sapindaceae) | Acetone extract | 279 | C15H21NO4 | Dongling Mountain, Beijing, China | [43,44] |
| Brasilamide D (48) | Paraconiothyrium brasiliense Verkley (M3–3341) (Leptosphaeriaceae)/ Acer truncatum Bunge (branches, Sapindaceae) | Acetone extract | 321 | C17H23NO5 | Dongling Mountain, Beijing, China | [43] |
| Brasilamide K (49) | Paraconiothyrium brasiliense Verkley (M3–3341) (Leptosphaeriaceae)/ Acer truncatum Bunge (branches, Sapindaceae) | EtOAc extract | 279 | C15H21NO4 | Dongling Mountain, Beijing, China | [44] |
| Brasilamide L (50) | Paraconiothyrium brasiliense Verkley (M3–3341) (Leptosphaeriaceae)/ Acer truncatum Bunge (branches, Sapindaceae) | EtOAc extract | 265 | C15H23NO3 | Dongling Mountain, Beijing, China | [44] |
| Brasilamide M (51) | Paraconiothyrium brasiliense Verkley (M3–3341) (Leptosphaeriaceae)/ Acer truncatum Bunge (branches, Sapindaceae) | EtOAc extract | 293 | C15H19NO5 | Dongling Mountain, Beijing, China, | [44] |
| Brasilamide N (52) | Paraconiothyrium brasiliense Verkley (M3–3341) (Leptosphaeriaceae)/ Acer truncatum Bunge (branches, Sapindaceae) | EtOAc extract | 279 | C15H21NO4 | Dongling Mountain, Beijing, China | [44] |
| Craterodoratin I (53) | Craterellus odoratus (Cantharellaceae) | EtOAc extract | 250 | C15H22O3 | Southern part of the GaoligongMountains, Yunnan, China | [30] |
| Craterodoratin J (54) | Craterellus odoratus (Cantharellaceae) | EtOAc extract | 282 | C15H22O5 | Southern part of the GaoligongMountains, Yunnan, China | [30] |
| Craterodoratin K (55) | Craterellus odoratus (Cantharellaceae) | EtOAc extract | 282 | C15H22O5 | Southern part of the GaoligongMountains, Yunnan, China | [30] |
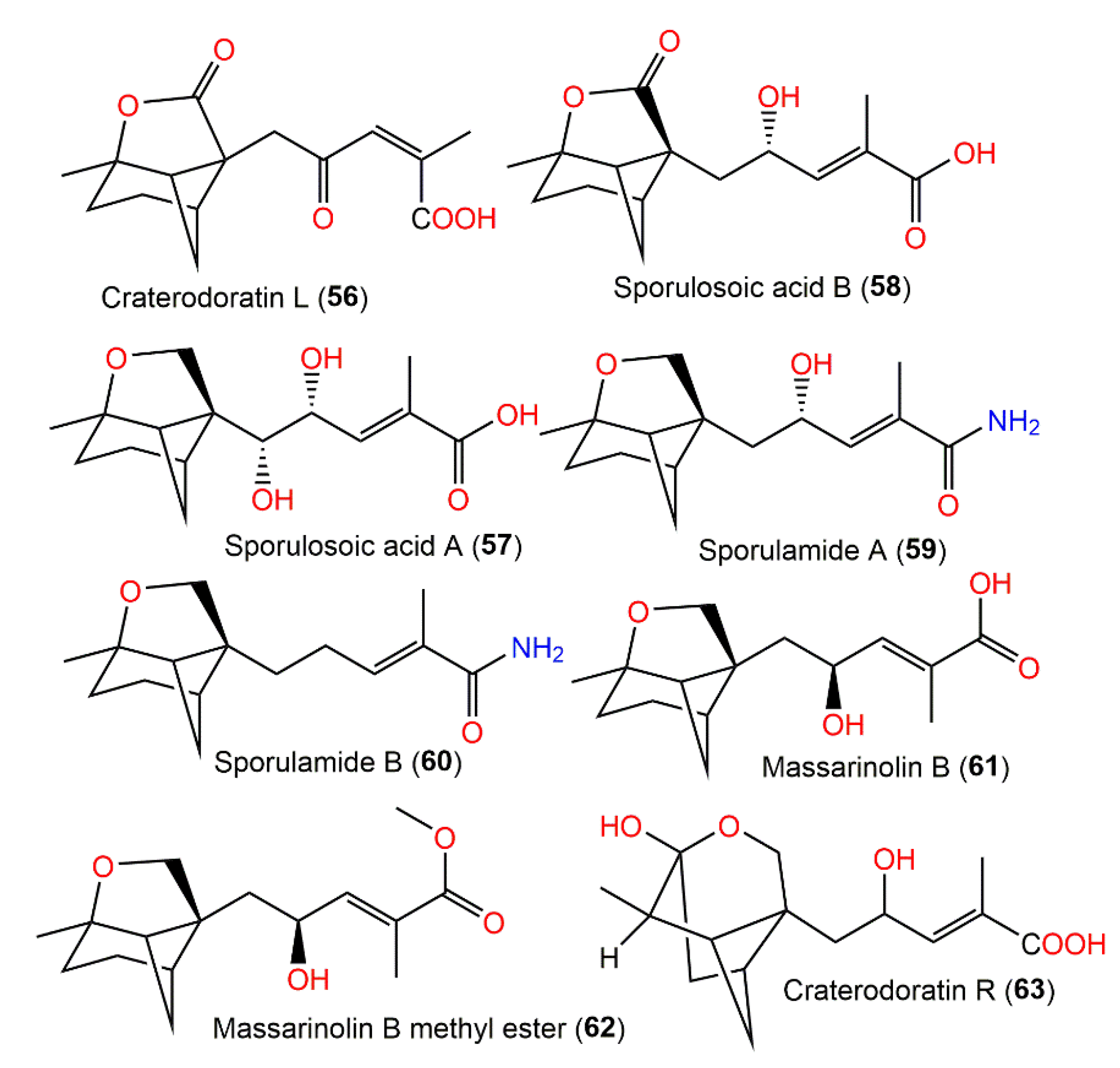
| Craterodoratin L (56) | Craterellus odoratus (Cantharellaceae) | EtOAc extract | 278 | C15H18O5 | Southern part of the GaoligongMountains, Yunnan, China | [30] |
| Sporulosoic acid A (57) | Paraconiothyrium sporulosum YK-03 Verkley (Leptosphaeriaceae)/Sea mud | EtOAc extract | 282 | C15H22O5 | Intertidal zone of Bohai Bay river in Liaoning, China | [31] |
| Sporulosoic acid B (58) | Paraconiothyrium sporulosum YK-03 Verkley (Leptosphaeriaceae)/Sea mud | EtOAc extract | 280 | C15H20O5 | Intertidal zone of Bohai Bay river in Liaoning, China | [31] |
| Sporulamide A (59) | Paraconiothyrium sporulosum YK-03 Verkley (Leptosphaeriaceae)/Sea mud | EtOAc extract | 265 | C15H23NO3 | Intertidal zone of Bohai Bay river in Liaoning, China | [31] |
| Sporulamide B (60) | Paraconiothyrium sporulosum YK-03 Verkley (Leptosphaeriaceae)/Sea mud | EtOAc extract | 249 | C15H23NO2 | Intertidal zone of Bohai Bay river in Liaoning, China | [31] |
| Massarinolin B (61) | Massarina tunicata (Lophiostomataceae)/Submerged twig | EtOAc extract | 266 | C15H22O4 | Lemonweir River in Adams County, Wisconsin, USA | [23] |
| Craterellus odoratus (Cantharellaceae) | EtOAc extract | - | - | Southern part of the GaoligongMountains, Yunnan, China | [30] | |
| Massarinolin B methyl ester (62) | Paraconiothyrium sporulosum YK-03 Verkley (Leptosphaeriaceae)/Sea mud | EtOAc extract | 280 | C16H24O4 | Intertidal zone of Bohai Bay river in Liaoning, China | [31] |
| Craterodoratin R (63) | Craterellus odoratus (Cantharellaceae) | EtOAc extract | 282 | C15H22O5 | Southern part of the GaoligongMountains, Yunnan, China | [30] |
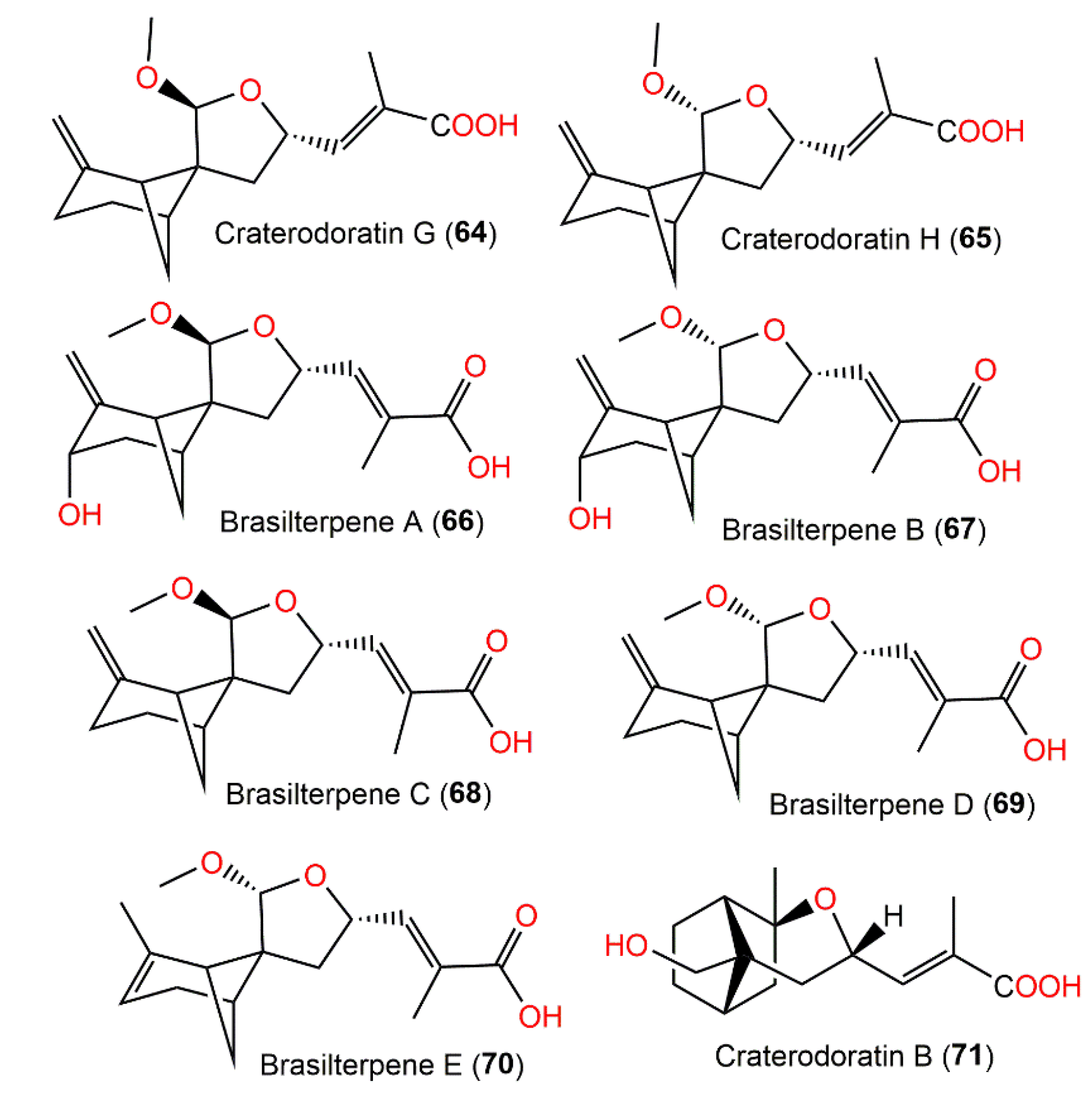
| Craterodoratin G (64) | Craterellus odoratus (Cantharellaceae) | EtOAc extract | 278 | C16H22O4 | Southern part of the GaoligongMountains, Yunnan, China | [30] |
| Craterodoratin H (65) | Craterellus odoratus (Cantharellaceae) | EtOAc extract | 278 | C16H22O4 | Southern part of the GaoligongMountains, Yunnan, China | [30] |
| Brasilterpene A (66) | Paraconiothyrium brasiliense HDN15-135 (Leptosphaeriaceae)/Deep-sea sediment | EtOAc extract | 294 | C16H22O5 | Indian Ocean, China | [45] |
| Brasilterpene B (67) | Paraconiothyrium brasiliense HDN15-135 (Leptosphaeriaceae)/Deep-sea sediment | EtOAc extract | 294 | C16H22O5 | Indian Ocean, China | [45] |
| Brasilterpene C (68) | Paraconiothyrium brasiliense HDN15-135 (Leptosphaeriaceae)/Deep-sea sediment | EtOAc extract | 278 | C16H22O4 | Indian Ocean, China | [45] |
| Brasilterpene D (69) | Paraconiothyrium brasiliense HDN15-135 (Leptosphaeriaceae)/Deep-sea sediment | EtOAc extract | 278 | C16H22O4 | Indian Ocean, China | [45] |
| Brasilterpene E (70) | Paraconiothyrium brasiliense HDN15-135 (Leptosphaeriaceae)/Deep-sea sediment | EtOAc extract | 278 | C16H22O4 | Indian Ocean, China | [45] |
| Craterodoratin B (71) | Craterellus odoratus (Cantharellaceae) | EtOAc extract | 266 | C15H22O4 | Southern part of the GaoligongMountains, Yunnan, China | [30] |
| Tetracyclic Bergamotane Sesquiterpenoids | ||||||
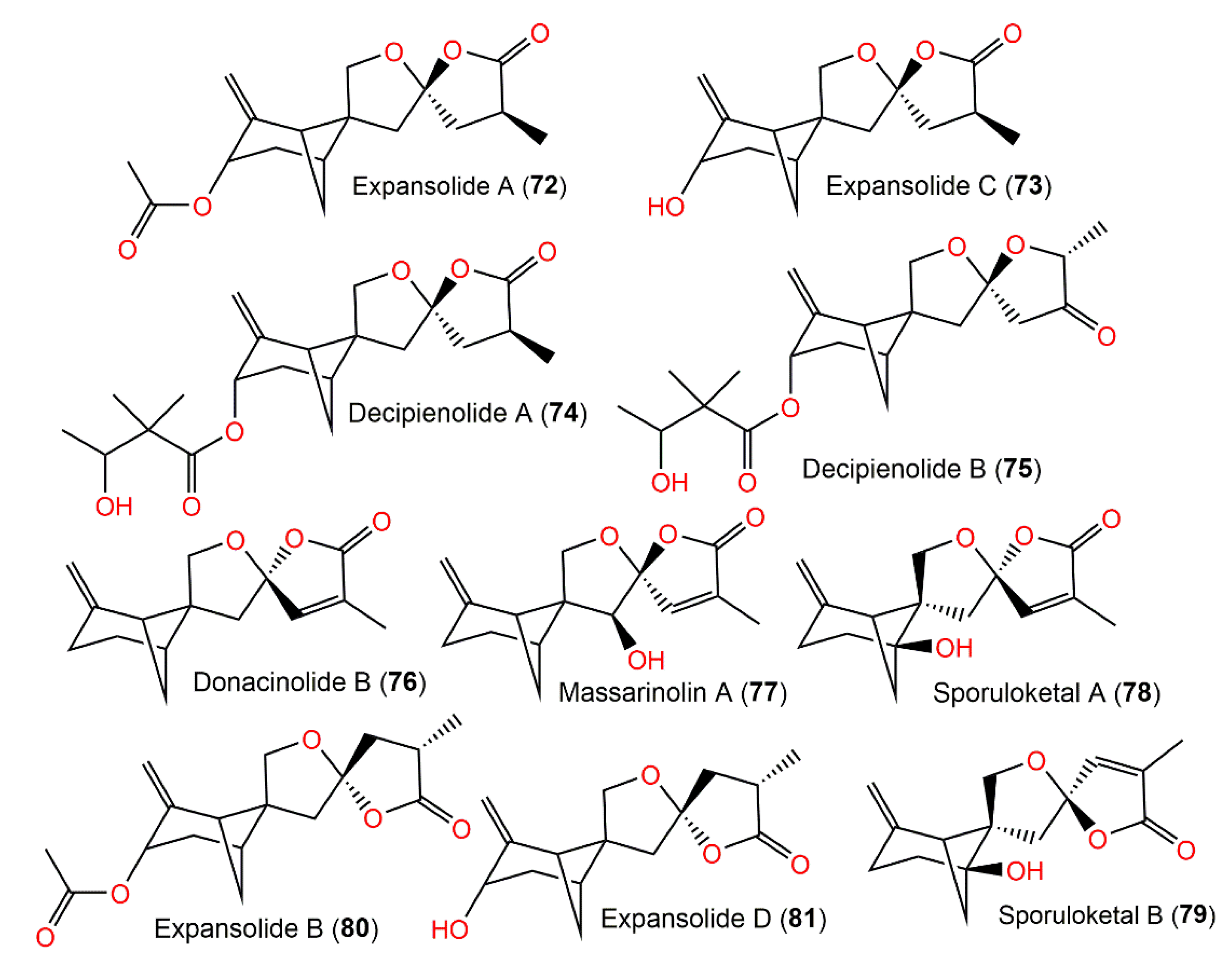
| Expansolide A (72) | Penicillium expansum (Trichocomaceae)/Fruit | EtOAc extract | 306 | C17H22O5 | France | [25] |
| Aspergillus fumigatus Fresenius (Trichocomaceae)/Leaf litter | EtOAc extract | - | - | Waipoua Forest, New Zealand | [26] | |
| Expansolide C (73) | Penicillium expansum ACCC37275/Agricultural Culture | Acetone extract | 264 | C15H20O4 | China | [46] |
| Decipienolide A (74) | Podospora decipiens Niessl (JS 270) (Podosporaceae)/Sheep dung | EtOAc extract | 378 | C21H30O6 | South Australia | [24] |
| Decipienolide B (75) | Podospora decipiens Niessl (JS 270) (Podosporaceae)/Sheep dung | EtOAc extract | 378 | C21H30O6 | South Australia | [24] |
| Donacinolide B (76) | Montagnula donacina (Montagnulaceae)/Craterellus odoratus (fruiting bodies, Cantharellaceae) | EtOAc extract | 246 | C15H18O3 | Southern part of the Gaoligong Mountains in Yunnan, China | [32] |
| Massarinolin A (77) | Massarina tunicata (Lophiostomataceae)/Submerged twig | EtOAc extract | 262 | C15H18O4 | LemonweirRiver in Adams County, Wisconsin, USA | [23] |
| Craterellus odoratus (Cantharellaceae) | EtOAc extract | - | - | Southern part of the GaoligongMountains, Yunnan, China | [30] | |
| Sporuloketal A (78) | Paraconiothyrium sporulosum YK-03 Verkley (Leptosphaeriaceae)/Sea mud | EtOAc extract | 262 | C15H18O4 | Intertidal zone of Bohai Bay river in Liaoning, China | [31] |
| Sporuloketal B (79) | Paraconiothyrium sporulosum YK-03 Verkley (Leptosphaeriaceae)/Sea mud | EtOAc extract | 262 | C15H18O4 | Intertidal zone of Bohai Bay river in Liaoning, China | [31] |
| Expansolide B (80) | Penicillium expansum (Trichocomaceae) | EtOAc extract | 306 | C17H22O5 | France | [25] |
| Aspergillus fumigatus Fresenius (Trichocomaceae)/Leaf litter | EtOAc extract | - | - | Waipoua Forest, New Zealand | [26] | |
| Expansolide D (81) | Penicillium expansum ACCC37275, (Trichocomaceae)/Agricultural Culture | Acetone extract | 264 | C15H20O4 | China | [46] |
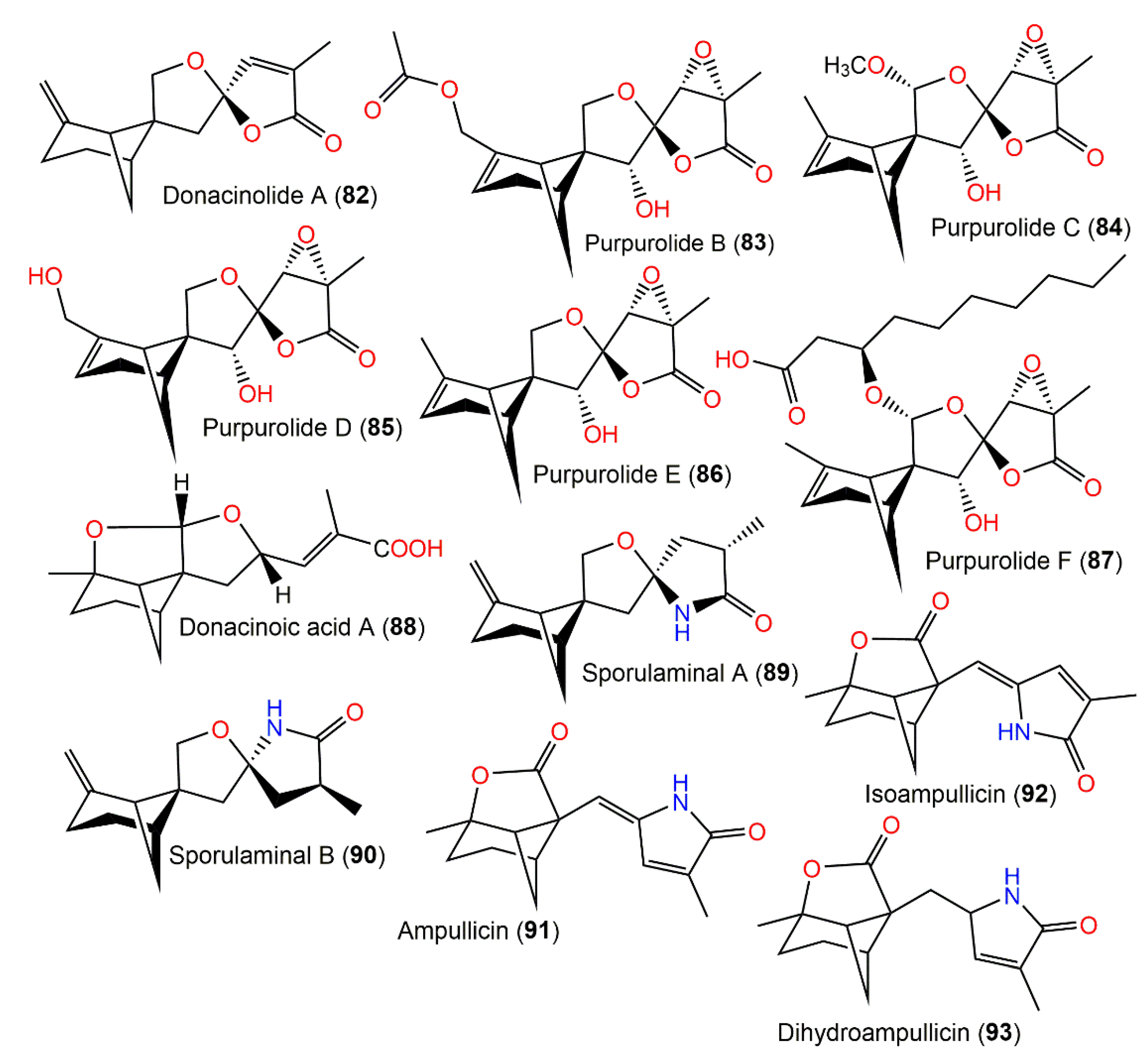
| Donacinolide A (82) | Montagnula donacina (Montagnulaceae)/Craterellus odoratus (fruiting bodies, Cantharellaceae) | EtOAc extract | 246 | C15H18O3 | Southern part of the Gaoligong Mountains in Yunnan, China | [32] |
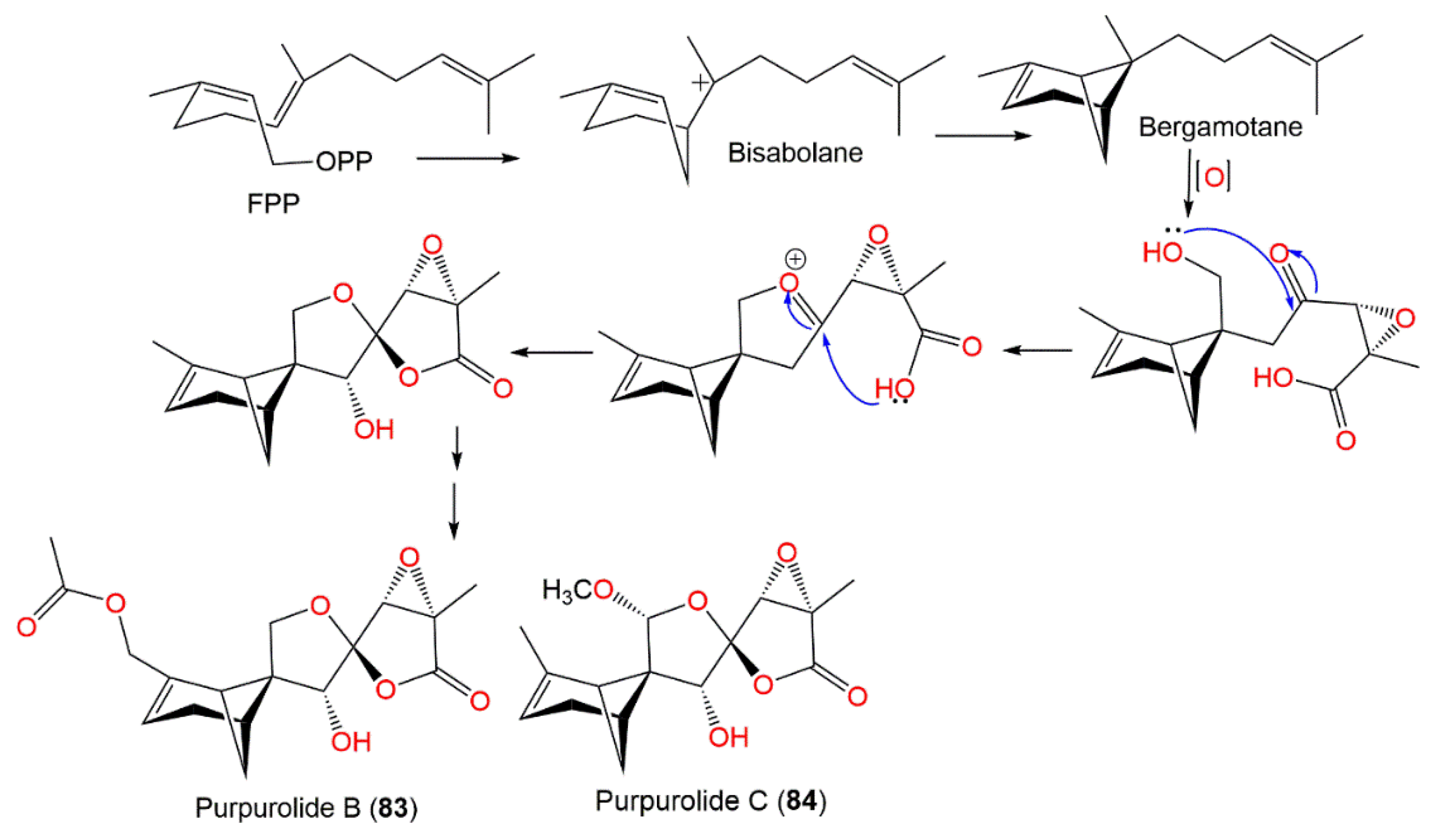
| Purpurolide B (83) | Penicillium purpurogenum IMM003 (Trichocomaceae)/Edgeworthia Chrysantha (leaves, Thymelaeaceae) | EtOAc extract | 336 | C17H20O7 | Hangzhou Bay, Hangzhou, Zhejiang, China | [47] |
| Purpurolide C (84) | Penicillium purpurogenum IMM003 (Trichocomaceae)/Edgeworthia Chrysantha (leaves, Thymelaeaceae) | EtOAc extract | 308 | C16H20O6 | Hangzhou Bay, Hangzhou, Zhejiang, China | [47] |
| Purpurolide D (85) | Penicillium purpurogenum IMM003 (Trichocomaceae)/Edgeworthia Chrysantha (leaves, Thymelaeaceae) | EtOAc extract | 294 | C15H18O6 | Hangzhou Bay, Hangzhou, Zhejiang, China | [48] |
| Purpurolide E (86) | Penicillium purpurogenum IMM003 (Trichocomaceae)/Edgeworthia Chrysantha (leaves, Thymelaeaceae) | EtOAc extract | 278 | C15H18O5 | Hangzhou Bay, Hangzhou, Zhejiang, China | [48] |
| Purpurolide F (87) | Penicillium purpurogenum IMM003 (Trichocomaceae)/Edgeworthia Chrysantha (leaves, Thymelaeaceae) | EtOAc extract | 464 | C25H36O8 | Hangzhou Bay, Hangzhou, Zhejiang, China | [48] |
| Donacinoic acid A (88) | Montagnula donacina (Montagnulaceae)/Craterellus odoratus (fruiting bodies, Cantharellaceae) | EtOAc extract | 264 | C15H20O4 | Southern part of the Gaoligong Mountains in Yunnan, China | [32] |
| Craterellus odoratus (Cantharellaceae) | EtOAc extract | - | - | Southern part of the GaoligongMountains, Yunnan, China | [30] | |
| Sporulaminal A (89) | Paraconiothyrium sporulosum YK-03 (Leptosphaeriaceae)/Sea mud | EtOAc extract | 247 | C15H21NO2 | Intertidal zone of Bohai river in Liaonign, China | [49] |
| Sporulaminal B (90) | Paraconiothyrium sporulosum YK-03 (Leptosphaeriaceae)/Sea mud | EtOAc extract | 247 | C15H21NO2 | Intertidal zone of Bohai river in Liaonign, China | [49] |
| Ampullicin (91) | Ampulliferina-like sp. No. 27 (Ampullicephala)/Pinus thunbergii (dead tree, Pinaceae) | Acetone extract | 259 | C15H17NO3 | Japan | [50,51] |
| Isoampullicin (92) | Ampulliferina-like sp. No. 27 (Ampullicephala)/Pinus thunbergii (dead tree, Pinaceae) | Acetone extract | 259 | C15H17NO3 | Japan | [50] |
| Dihydroampullicin (93) | Ampulliferina-like sp. No. 27 (Ampullicephala)/Pinus thunbergii (dead tree, Pinaceae) | Acetone extract | 261 | C15H19NO3 | Japan | [51] |
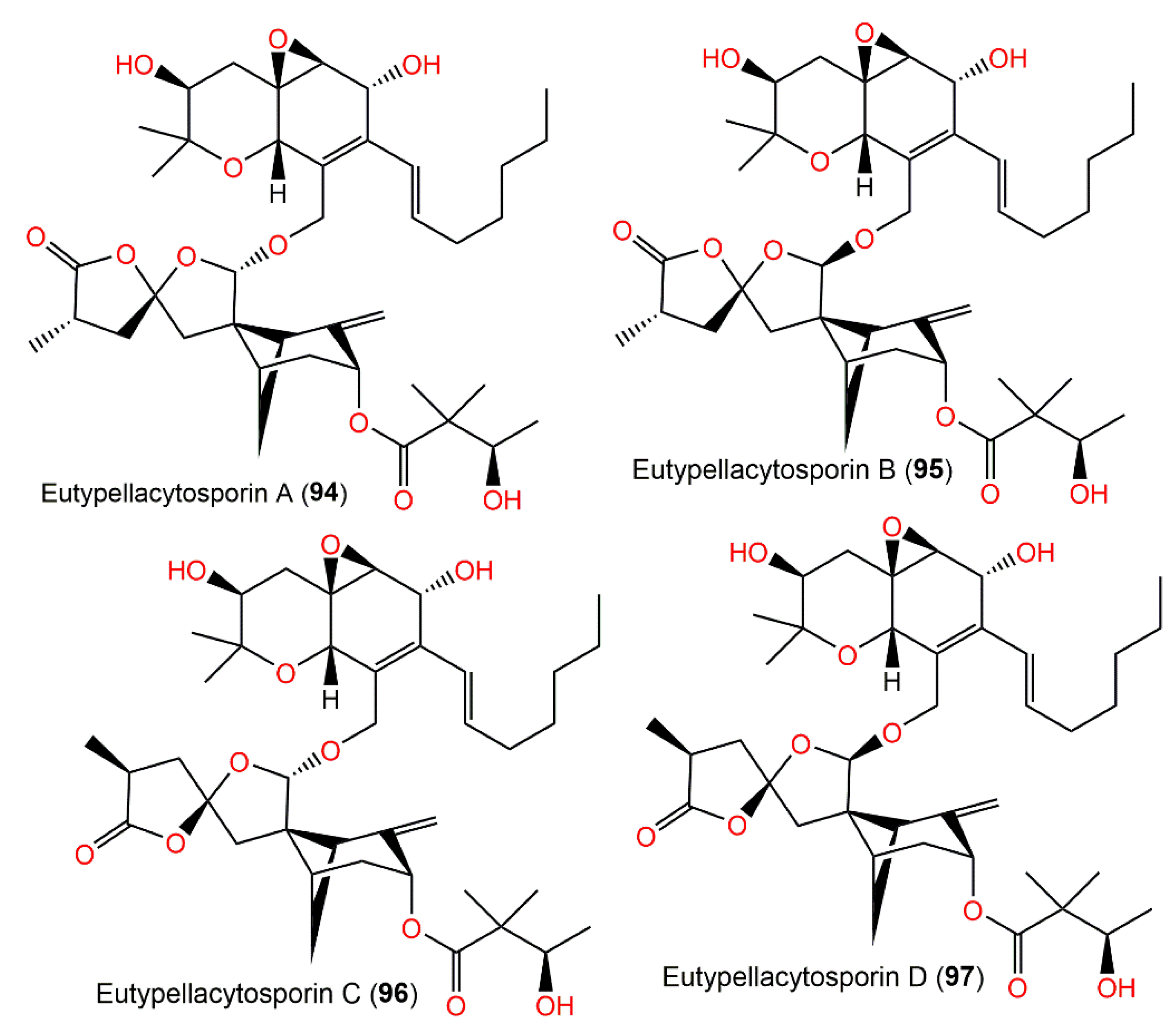
| Eutypellacytosporin A (94) | Eutypella sp. D-1 (Diatrypaceae)/Soil sample | CH2Cl2 fraction of EtOAc extract | 714 | C40H58O11 | London Island of Kongsfjorden of the Ny-Ålesund District, Arctic, Norway | [52] |
| Eutypellacytosporin B (95) | Eutypella sp. D-1 (Diatrypaceae)/Soil sample | CH2Cl2 fraction of EtOAc extract | 714 | C40H58O11 | London Island of Kongsfjorden of the Ny-Ålesund District, Arctic, Norway | [52] |
| Eutypellacytosporin C (96) | Eutypella sp. D-1 (Diatrypaceae)/Soil sample | CH2Cl2 fraction of EtOAc extract | 714 | C40H58O11 | London Island of Kongsfjorden of the Ny-Ålesund District, Arctic, Norway | [52] |
| Eutypellacytosporin D (97) | Eutypella sp. D-1 (Diatrypaceae)/Soil sample | CH2Cl2 fraction of EtOAc extract | 714 | C40H58O11 | London Island of Kongsfjorden of the Ny-Ålesund District, Arctic, Norway | [52] |
| Compound Name | Biological Activity | Assay, Organism, or Cell Line | Biological Results | Ref. | |
|---|---|---|---|---|---|
| Compound | Positive Control | ||||
| E-β-trans-5,8,11-trihydroxybergamot-9-ene (4) | Cytotoxicity | MTT/U937 | 84.9 (IC50) | Doxorubicin 0.021 µM (IC50) | [29] |
| Craterodoratin M (7) | Immunosuppressive | BALB/c mice T and B lymphocyte/LPS | 15.43 μM (IC50) | Cyclosporin A 0.47 μM (IC50) | [30] |
| Craterodoratin N (8) | Immunosuppressive | BALB/c mice T and B lymphocyte/LPS | 13.26 μM (IC50) | Cyclosporin A 0.47 μM (IC50) | [30] |
| Craterodoratin O (9) | Immunosuppressive | BALB/c mice T and B lymphocyte/LPS | 17.12 μM (IC50) | Cyclosporin A 0.47 μM (IC50) | [30] |
| Craterodoratin Q (11) | Immunosuppressive | BALB/c mice T and B lymphocyte/Concanavalin A | 31.50 μM (IC50) | Cyclosporin A 0.04 μM (IC50) | [30] |
| Xylariterpenoid A (16) | Anti-inflammatory | Spectrophotometrically/LPS | 17.5 μM (IC50) | Quercetin 17.0 μM (IC50) NG-monomethyl-L-arginine 9.7 μM (IC50) | [35] |
| Xylariterpenoid B (17) | Anti-inflammatory | Spectrophotometrically/LPS | 21.0 μM (IC50) | Quercetin 17.0 μM (IC50) NG-monomethyl-L-arginine 9.7 μM (IC50) | [35] |
| Eutypeterpene B (18) | Anti-inflammatory | Spectrophotometrically/LPS | 13.4 μM (IC50) | Quercetin 17.0 μM (IC50) NG-monomethyl-L-arginine 9.7 μM (IC50) | [35] |
| Eutypeterpene C (19) | Anti-inflammatory | Spectrophotometrically/LPS | 16.8 μM (IC50) | Quercetin 17.0 μM (IC50) NG-monomethyl-L-arginine 9.7 μM (IC50) | [35] |
| Eutypeterpene D (20) | Anti-inflammatory | Spectrophotometrically/LPS | 21.4 μM (IC50) | Quercetin 17.0 μM (IC50) NG-monomethyl-L-arginine 9.7 μM (IC50) | [35] |
| Eutypeterpene E (21) | Anti-inflammatory | Spectrophotometrically/LPS | 18.7 μM (IC50) | Quercetin 17.0 μM (IC50) NG-monomethyl-L-arginine 9.7 μM (IC50) | [35] |
| Eutypeterpene F (22) | Anti-inflammatory | Spectrophotometrically/LPS | 24.3 μM (IC50) | Quercetin 17.0 μM (IC50) NG-monomethyl-L-arginine 9.7 μM (IC50) | [35] |
| (10S)-Xylariterpenoid A (23) | Anti-inflammatory | Spectrophotometrically/LPS | 86.0 μM (IC50) | Aminoguanidine 23.0 μM (IC50) | [34] |
| (10R)-Xylariterpenoid B (24) | Anti-inflammatory | Spectrophotometrically/LPS | 230.0 μM (IC50) | Aminoguanidine 23.0 μM (IC50) | [34] |
| Xylariterpenoid E (25) | Anti-inflammatory | Spectrophotometrically/LPS | 120.0 μM (IC50) | Aminoguanidine 23.0 μM (IC50) | [34] |
| Xylariterpenoid F (26) | Anti-inflammatory | Spectrophotometrically/LPS | 85.0 μM (IC50) | Aminoguanidine 23.0 μM (IC50) | [34] |
| Xylariterpenoid G (27) | Anti-inflammatory | Spectrophotometrically/LPS | 85.0 μM (IC50) | Aminoguanidine 23.0 μM (IC50) | [34] |
| Eutypeterpene A (28) | Anti-inflammatory | Spectrophotometrically/LPS | 21.0 μM (IC50) | Quercetin 17.0 μM (IC50) NG-monomethyl-L-arginine 9.7 μM (IC50) | [35] |
| Craterodoratin C (30) | Immunosuppressive | BALB/c mice T and B lymphocyte/LPS | 12.62 μM (IC50) | Cyclosporin A 0.47 μM (IC50) | [30] |
| Craterodoratin S (42) | Immunosuppressive | BALB/c mice T and B lymphocyte/LPS | 22.68 μM (IC50) | Cyclosporin A 0.47 μM (IC50) | [30] |
| Craterodoratin J (54) | Immunosuppressive | BALB/c mice T and B lymphocyte/LPS | 19.40 μM (IC50) | Cyclosporin A 0.47 μM (IC50) | [30] |
| Craterodoratin L (56) | Immunosuppressive | BALB/c mice T and B lymphocyte/LPS | 13.71 μM (IC50) | Cyclosporin A 0.47 μM (IC50) | [30] |
| Massarinolin B (61) | Immunosuppressive | BALB/c mice T and B lymphocyte/Concanavalin A | 0.98 μM (IC50) | Cyclosporin A 0.04 μM (IC50) | [30] |
| Brasilterpene A (66) | Hypoglycemic | Spectrophotometrically/Diabetic zebrafish model | 449.3 pmol/larva (IC50) | Rosiglitazone 395.6 pmol/larva (IC50) | [45] |
| Brasilterpene C (68) | Hypoglycemic | Spectrophotometrically/Diabetic zebrafish model | 420.4 pmol/larva (IC50) | Rosiglitazone 395.6 pmol/larva (IC50) | [45] |
| Expansolide C (73) | α-Glucosidase inhibition | Spectrophotometrically/α-glucosidase enzyme | 0.50 mM (IC50) | Acarbose 1.90 mM (IC50) | [46] |
| Expansolide D (81) | α-Glucosidase inhibition | Spectrophotometrically/α-glucosidase enzyme | 0.50 mM (IC50) | acarbose 1.90 mM (IC50) | [46] |
| Purpurolide B (83) | Pancreatic lipase inhibition | Spectrophotometrically/pancreatic lipase enzyme | 5.45 μM (IC50) | Kaempferol 1.50 μM (IC50) | [47] |
| Purpurolide C (84) | Pancreatic lipase inhibition | Spectrophotometrically/pancreatic lipase enzyme | 6.63 μM (IC50) | Kaempferol 1.50 μM (IC50) | [47] |
| Purpurolide D (85) | Pancreatic lipase inhibition | Spectrophotometrically/pancreatic lipase enzyme | 1.22 μM (IC50) | Kaempferol 1.50 μM (IC50) | [48] |
| Purpurolide E (86) | Pancreatic lipase inhibition | Spectrophotometrically/pancreatic lipase enzyme | 6.50 μM (IC50) | Kaempferol 1.50 μM (IC50) | [48] |
| Purpurolide F (87) | Pancreatic lipase inhibition | Spectrophotometrically/pancreatic lipase enzyme | 7.88 μM (IC50) | Kaempferol 1.50 μM (IC50) | [48] |
| Donacinoic acid A (88) | Immunosuppressive | BALB/c mice T and B lymphocyte/LPS | 13.23 μM (IC50) | Cyclosporin A 0.47 μM (IC50) | [30] |
| Eutypellacytosporin A (94) | Cytotoxicity | CCK-8/DU145 | 17.1 μM (IC50) | Cisplatin 2.9 μM (IC50) | [52] |
| CCK-8/SW1990 | 7.3 μM (IC50) | Cisplatin 1.2 μM (IC50) | [52] | ||
| CCK-8/Huh7 | 8.4 μM (IC50) | Cisplatin 2.2 μM (IC50) | [52] | ||
| CCK-8/PANC-1 | 9.7 μM (IC50) | Cisplatin 4.5 μM (IC50) | [52] | ||
| Eutypellacytosporin B (95) | Cytotoxicity | CCK-8/DU145 | 11.0 μM (IC50) | Cisplatin 2.9 μM (IC50) | [52] |
| CCK-8/SW1990 | 4.9 μM (IC50) | Cisplatin 1.2 μM (IC50) | [52] | ||
| CCK-8/Huh7 | 4.9 μM (IC50) | Cisplatin 2.2 μM (IC50) | [52] | ||
| CCK-8/PANC-1 | 7.9 μM (IC50) | Cisplatin 4.5 μM (IC50) | [52] | ||
| Eutypellacytosporin C (96) | Cytotoxicity | CCK-8/DU145 | 13.5 μM (IC50) | Cisplatin 2.9 μM (IC50) | [52] |
| CCK-8/SW1990 | 9.6 μM (IC50) | Cisplatin 1.2 μM (IC50) | [52] | ||
| CCK-8/Huh7 | 11.2 μM (IC50) | Cisplatin 2.2 μM (IC50) | [52] | ||
| CCK-8/PANC-1 | 10.2 μM (IC50) | Cisplatin 4.5 μM (IC50) | [52] | ||
| Eutypellacytosporin D (97) | Cytotoxicity | CCK-8/DU145 | 13.4 μM (IC50) | Cisplatin 2.9 μM (IC50) | [52] |
| CCK-8/SW1990 | 8.2 μM (IC50) | Cisplatin 1.2 μM (IC50) | [52] | ||
| CCK-8/Huh7 | 9.6 μM (IC50) | Cisplatin 2.2 μM (IC50) | [52] | ||
| CCK-8/PANC-1 | 7.5 μM (IC50) | Cisplatin 4.5 μM (IC50) | [52] | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khayat, M.T.; Mohammad, K.A.; Omar, A.M.; Mohamed, G.A.; Ibrahim, S.R.M. Fungal Bergamotane Sesquiterpenoids—Potential Metabolites: Sources, Bioactivities, and Biosynthesis. Mar. Drugs 2022, 20, 771. https://doi.org/10.3390/md20120771
Khayat MT, Mohammad KA, Omar AM, Mohamed GA, Ibrahim SRM. Fungal Bergamotane Sesquiterpenoids—Potential Metabolites: Sources, Bioactivities, and Biosynthesis. Marine Drugs. 2022; 20(12):771. https://doi.org/10.3390/md20120771
Chicago/Turabian StyleKhayat, Maan T., Khadijah A. Mohammad, Abdelsattar M. Omar, Gamal A. Mohamed, and Sabrin R. M. Ibrahim. 2022. "Fungal Bergamotane Sesquiterpenoids—Potential Metabolites: Sources, Bioactivities, and Biosynthesis" Marine Drugs 20, no. 12: 771. https://doi.org/10.3390/md20120771
APA StyleKhayat, M. T., Mohammad, K. A., Omar, A. M., Mohamed, G. A., & Ibrahim, S. R. M. (2022). Fungal Bergamotane Sesquiterpenoids—Potential Metabolites: Sources, Bioactivities, and Biosynthesis. Marine Drugs, 20(12), 771. https://doi.org/10.3390/md20120771