
Actinoporin-like Proteins Are Widely Distributed in the Phylum Porifera


Abstract
:
1. Introduction
2. Results
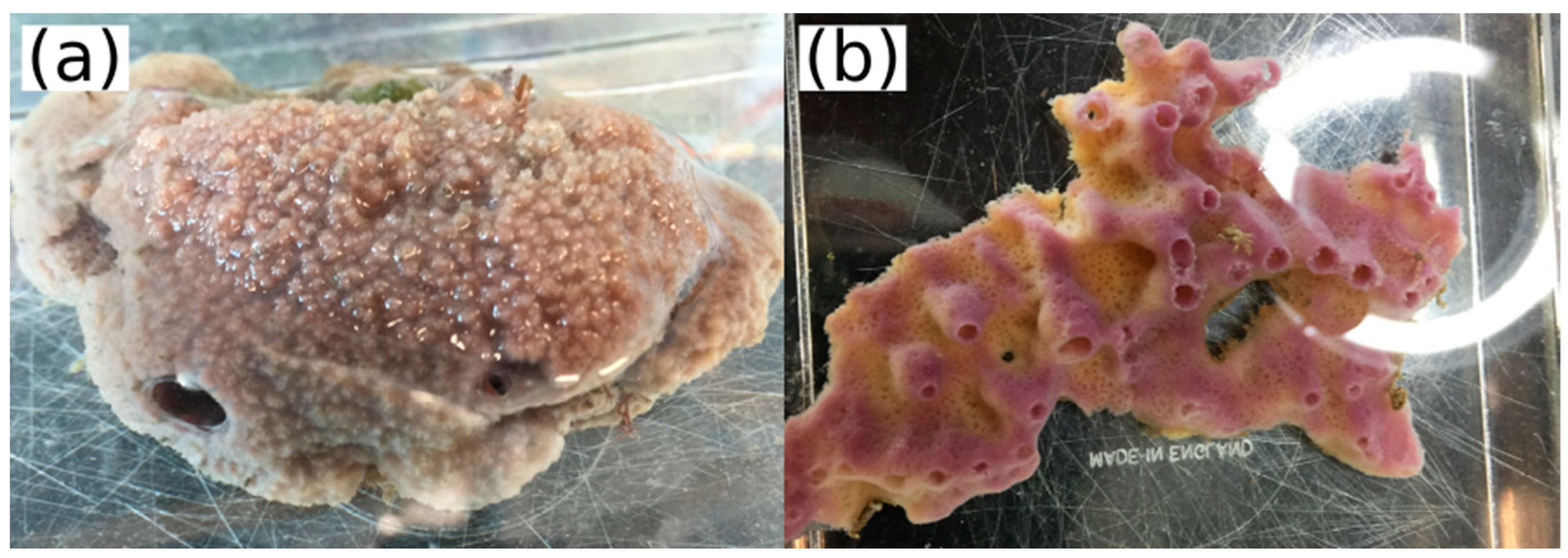
2.1. Transcriptome Sequencing and ALP Identification
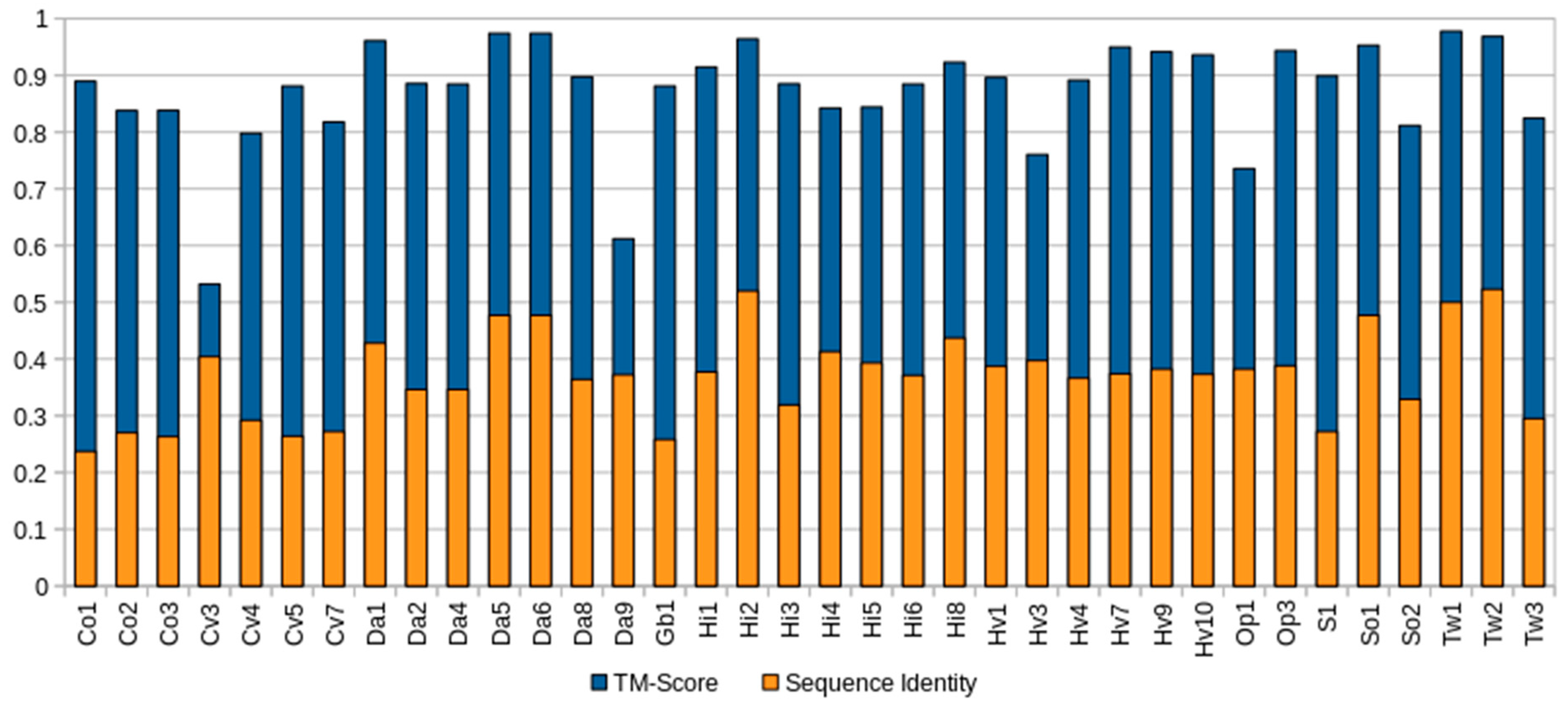
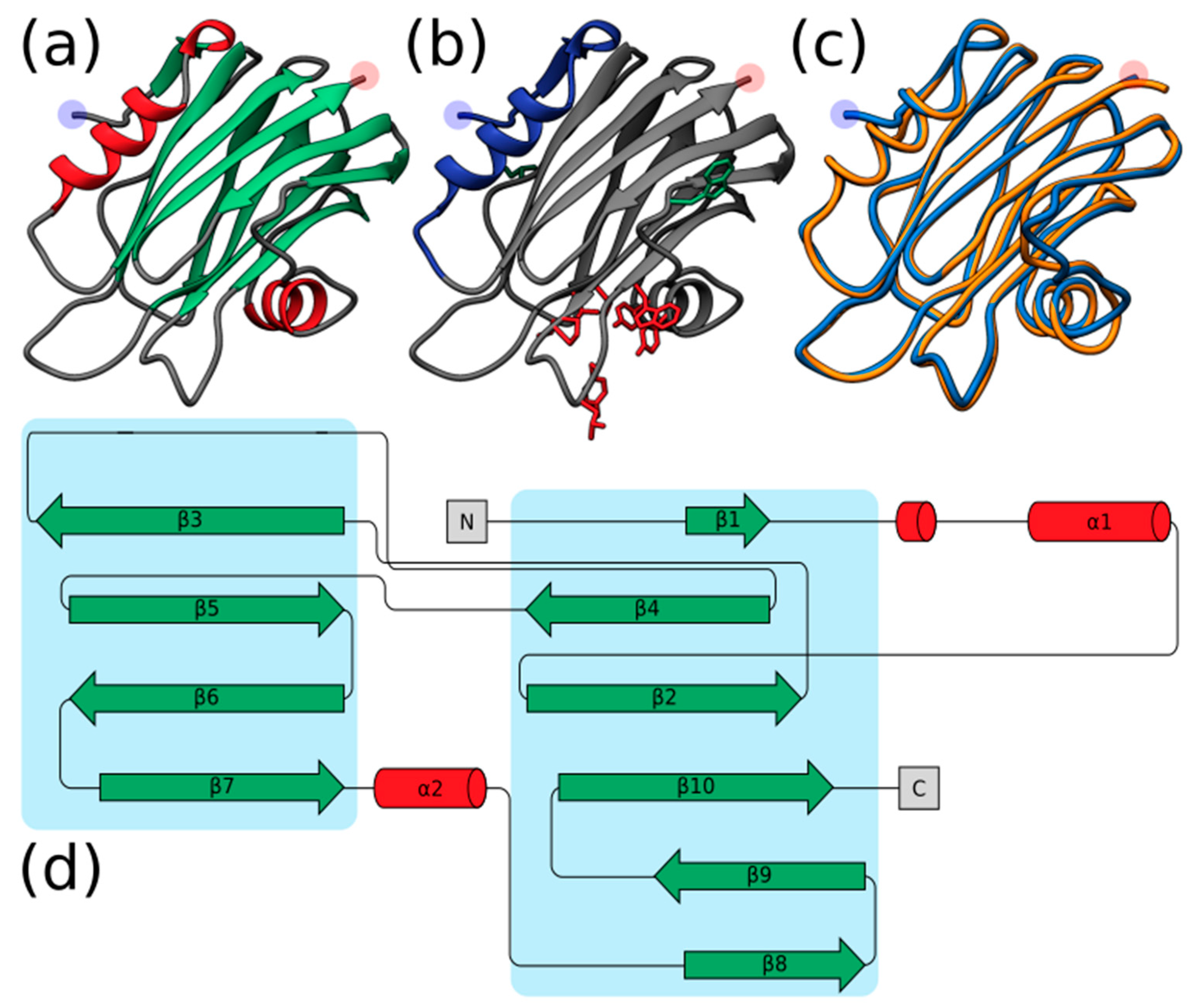
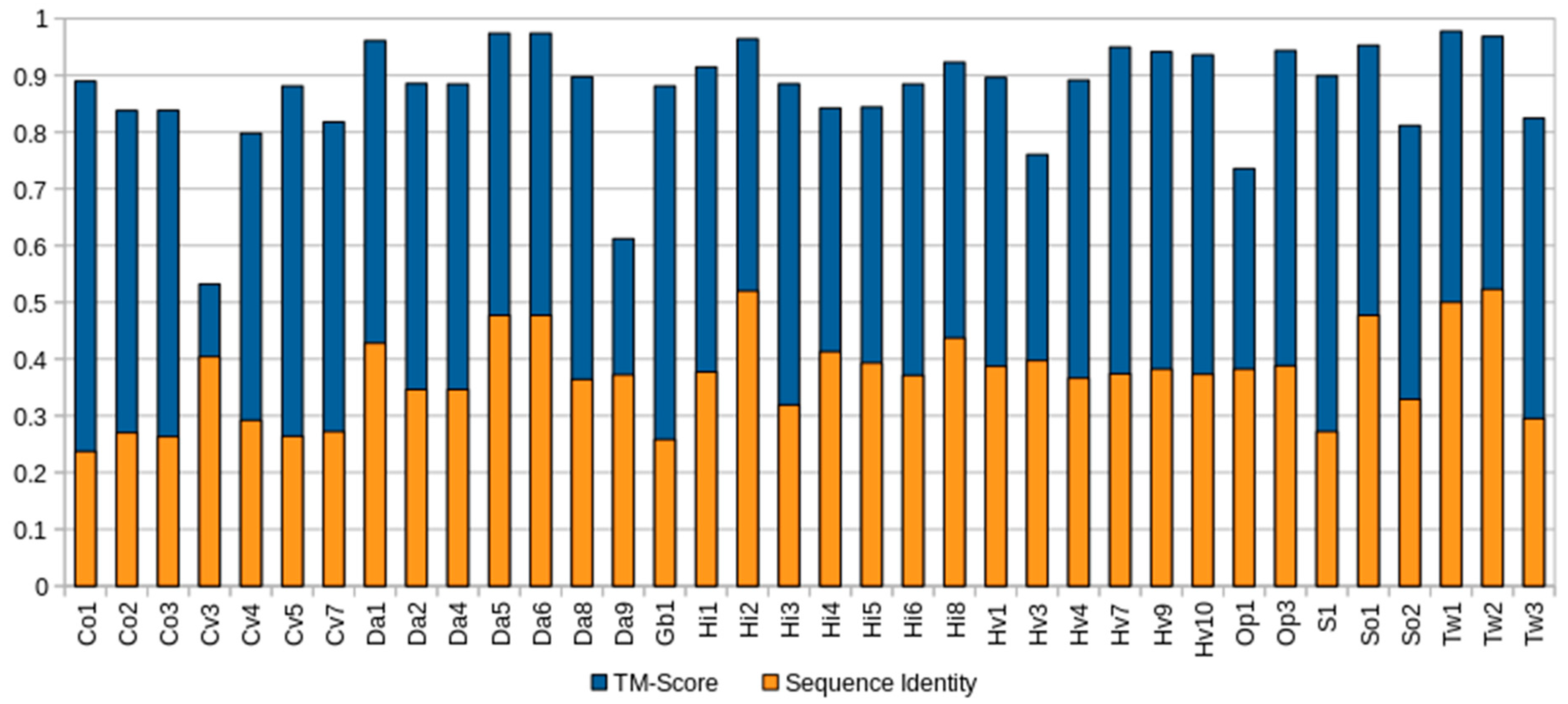
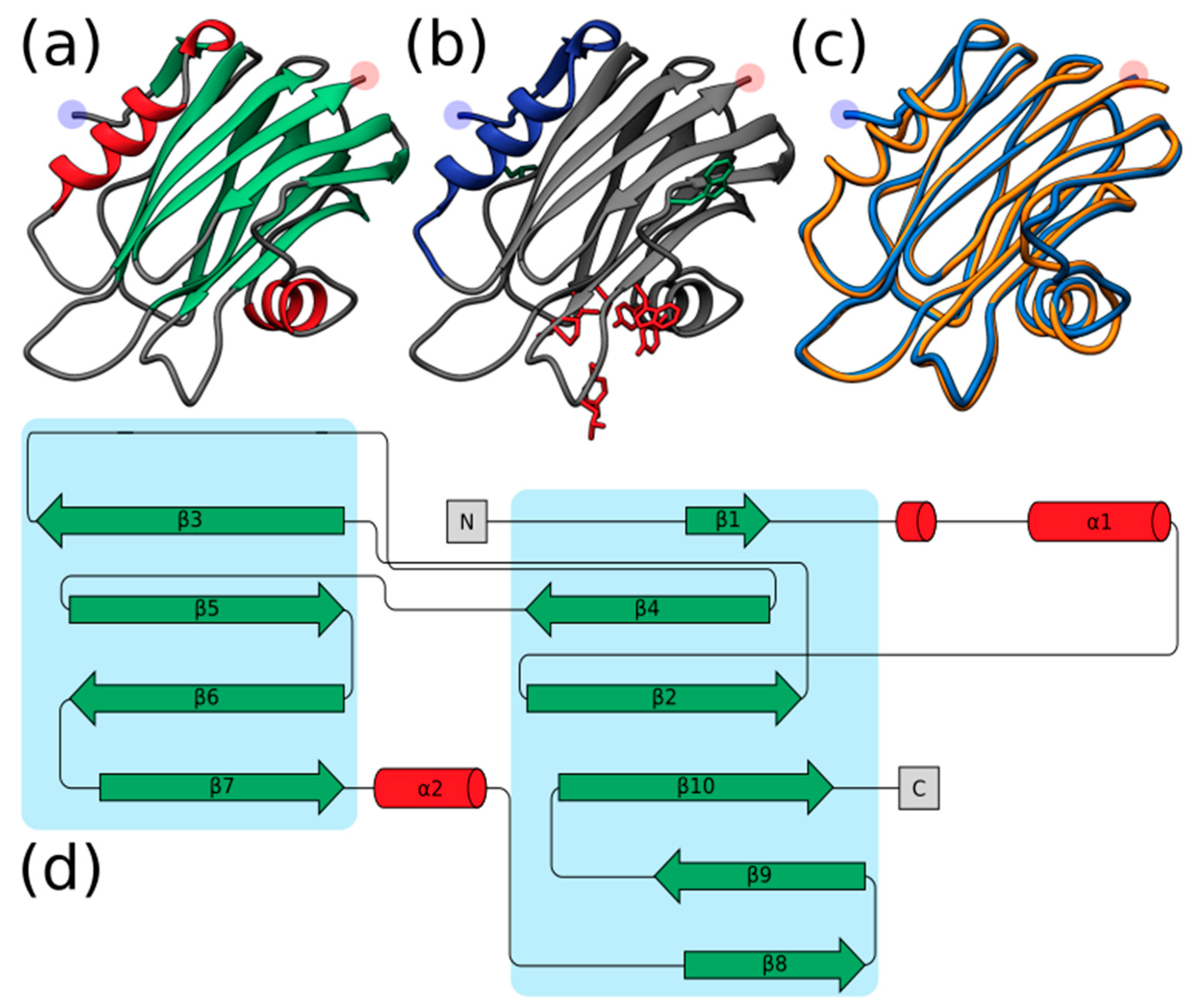
2.2. Structural Prediction
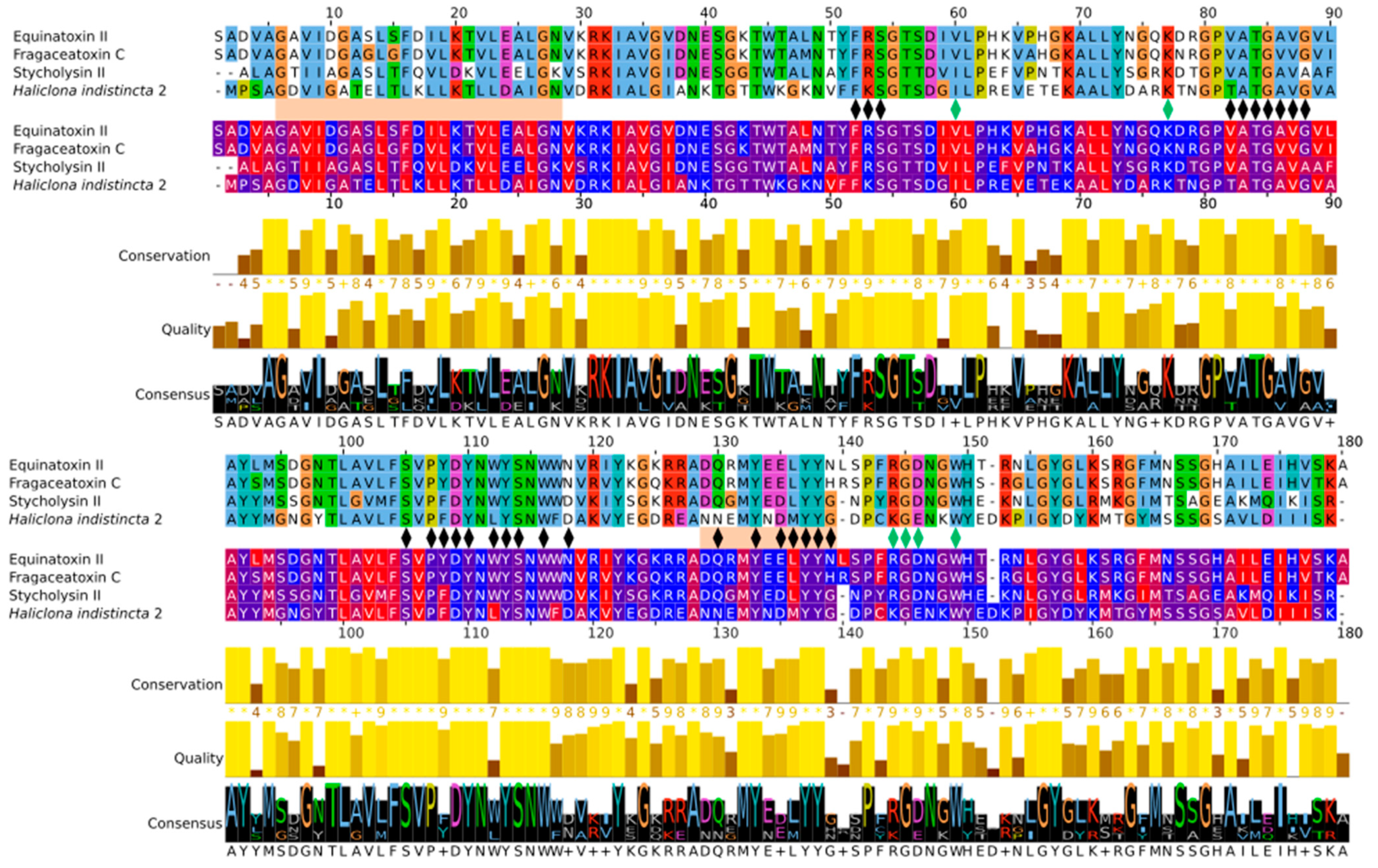
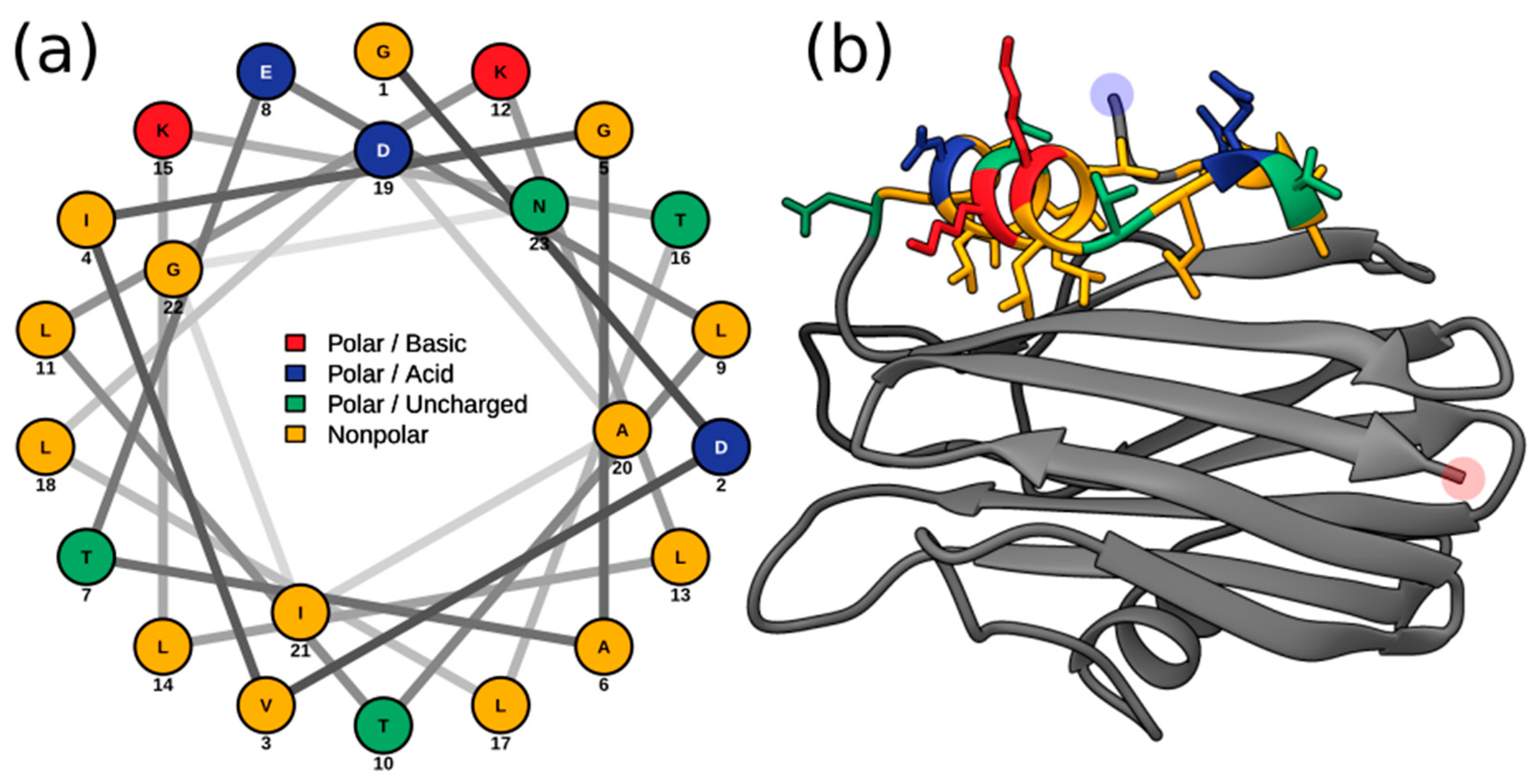
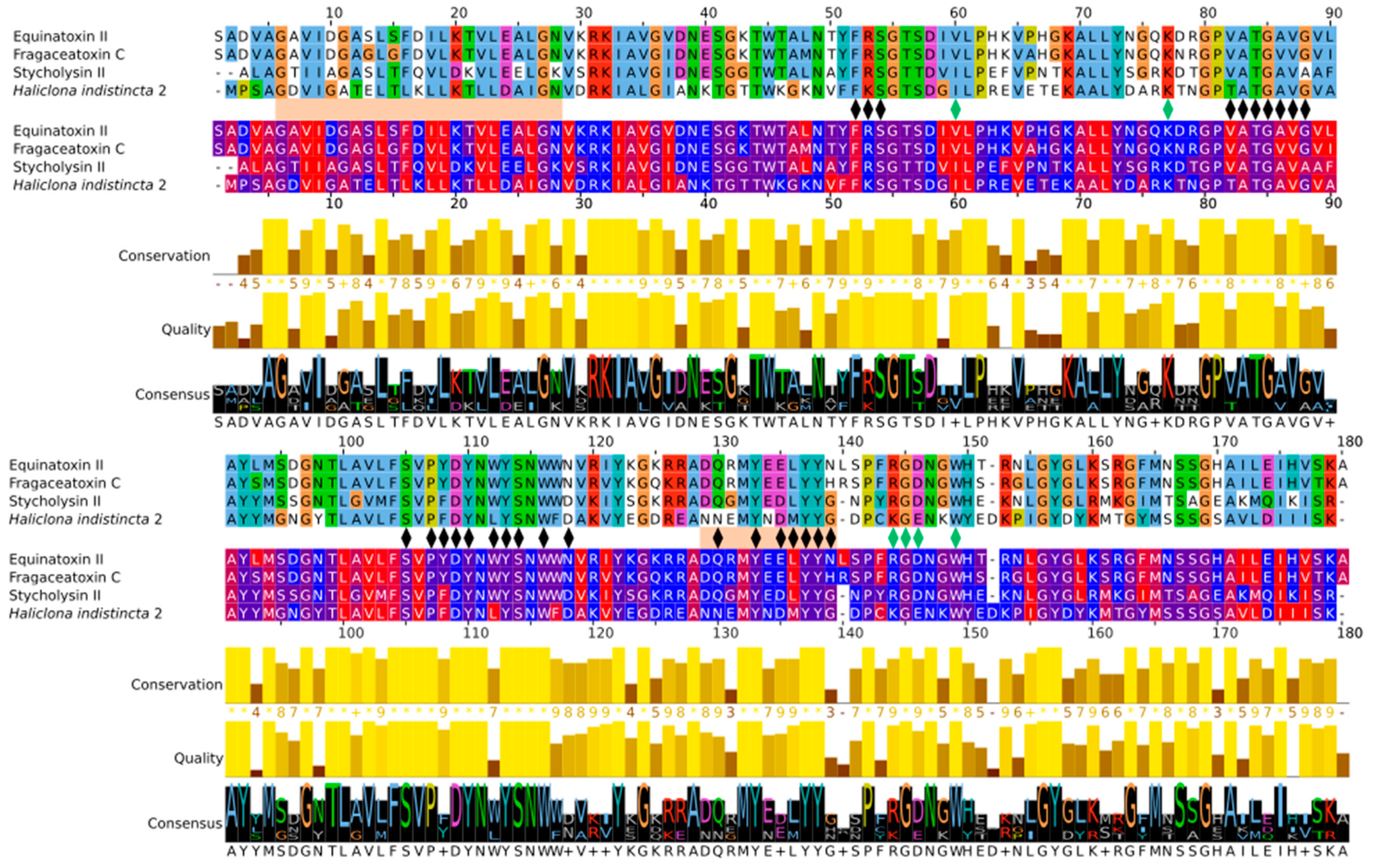
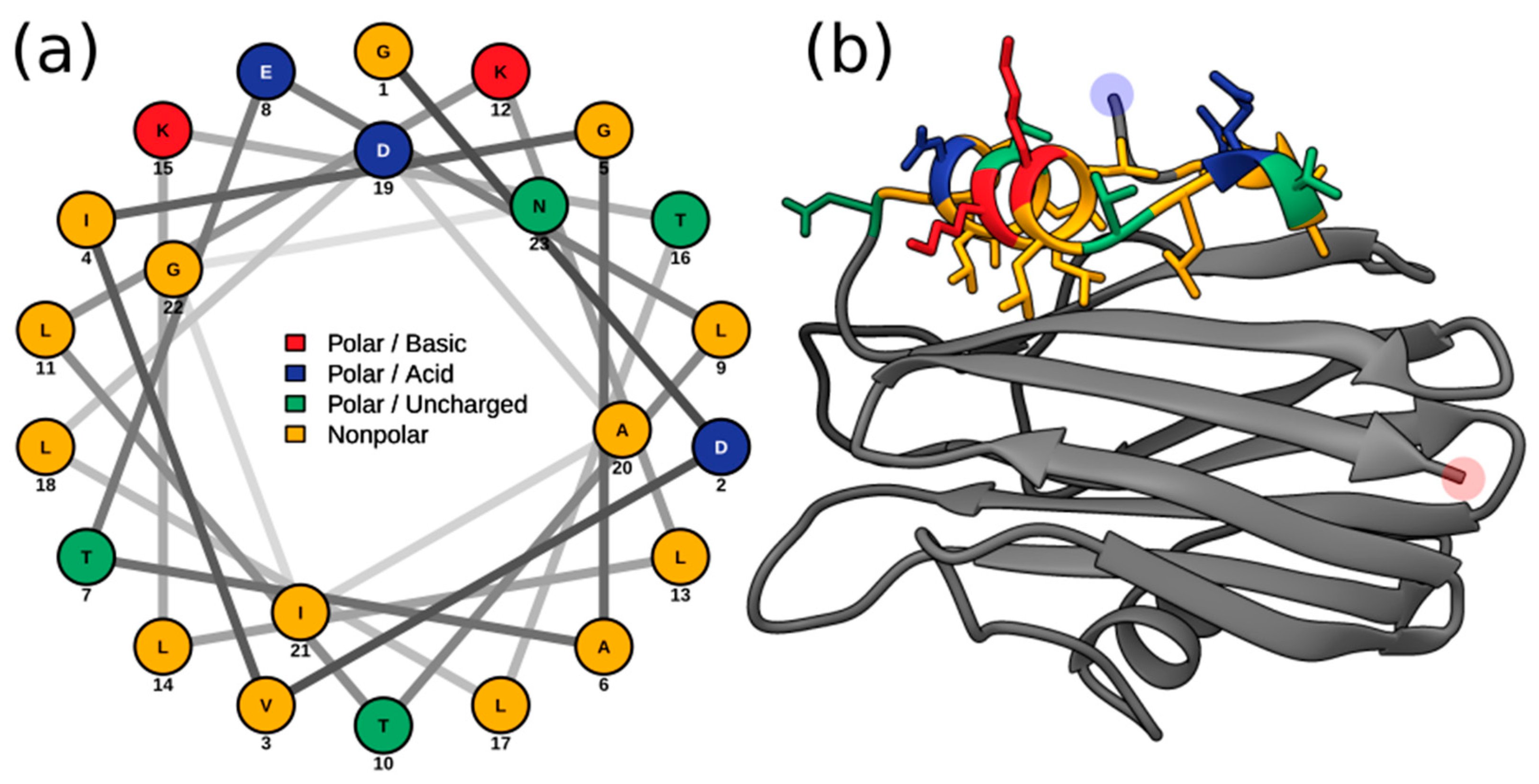
2.3. Multiple Sequence Alignment and Residue Analysis
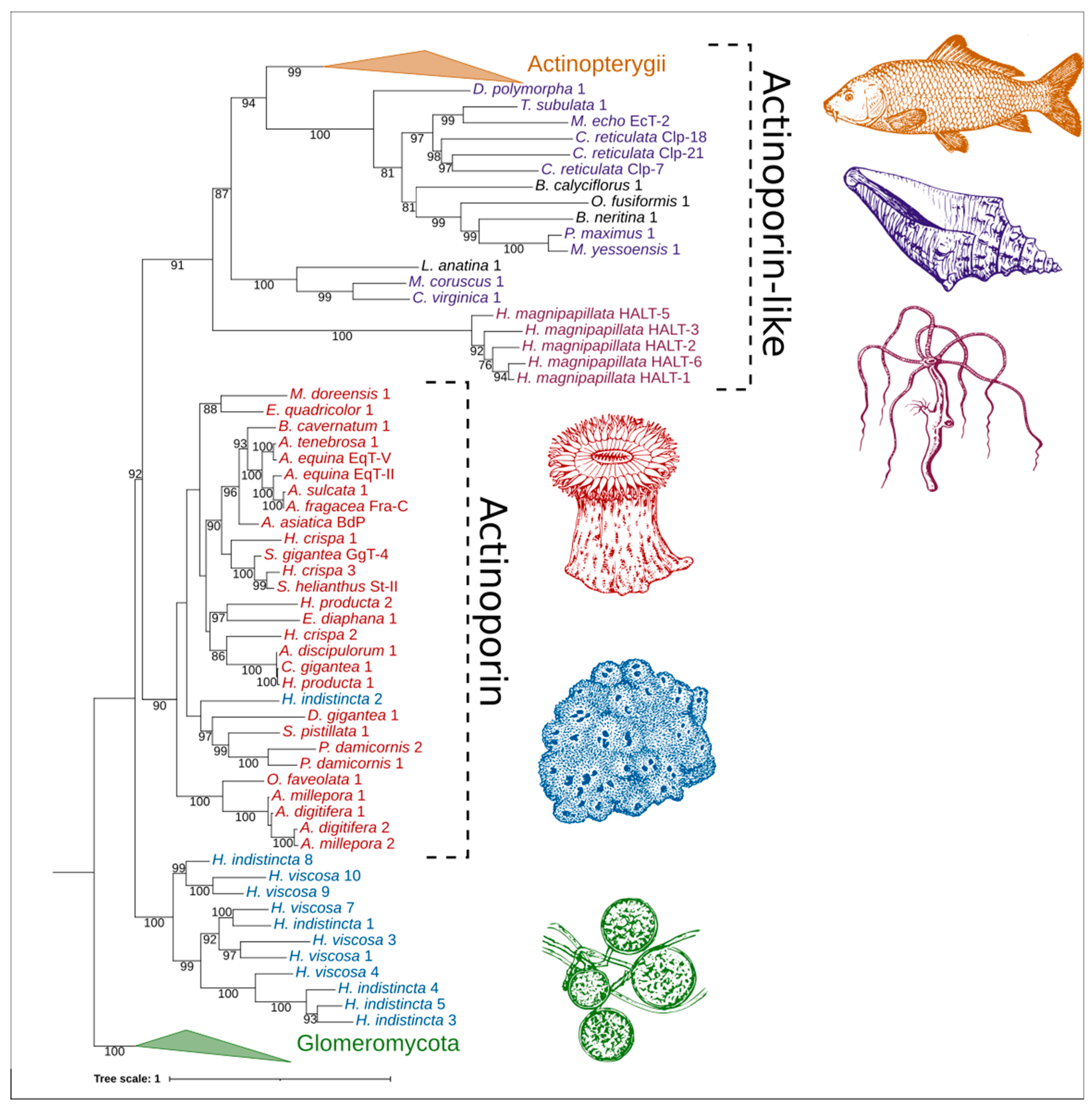
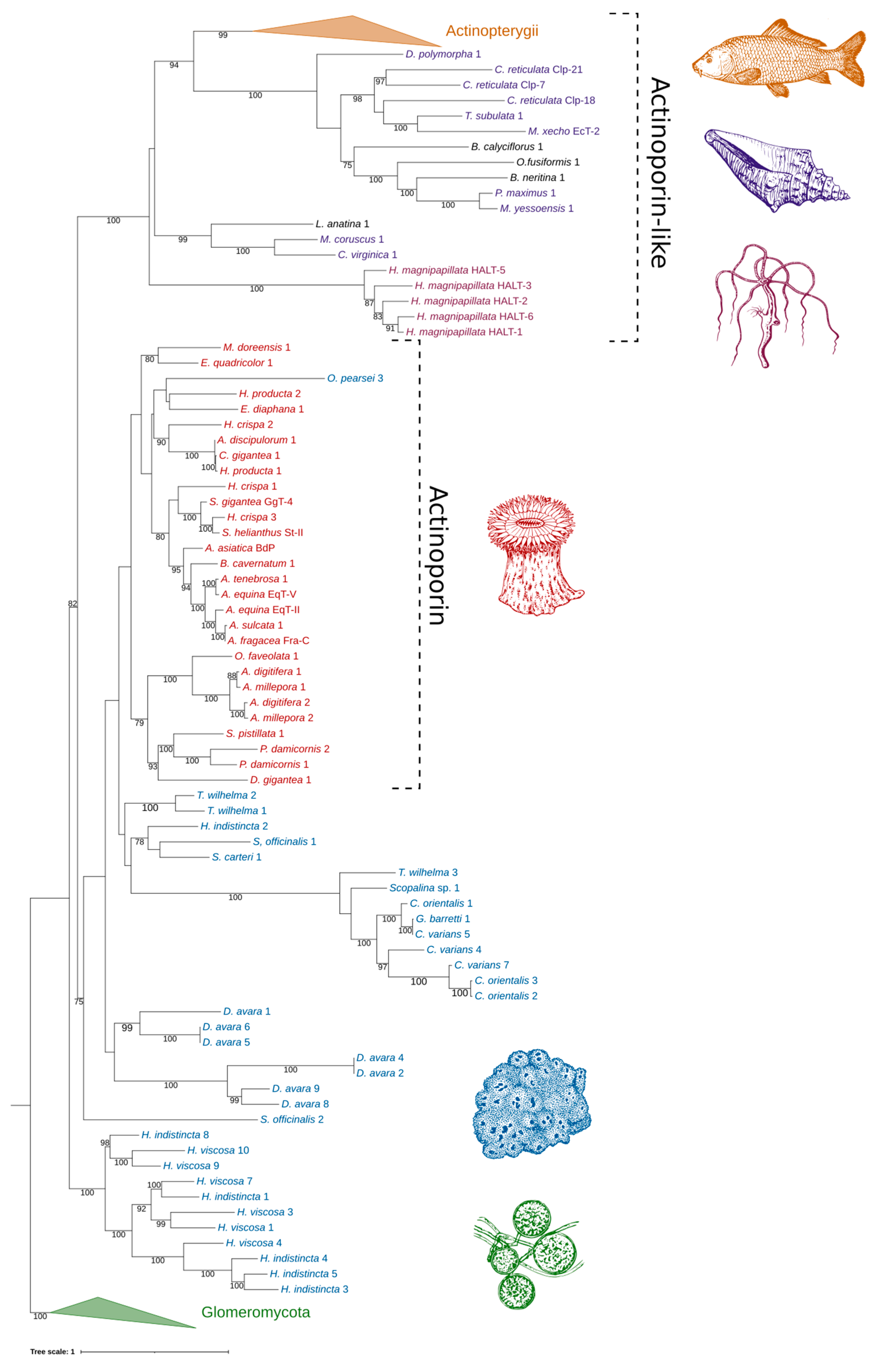
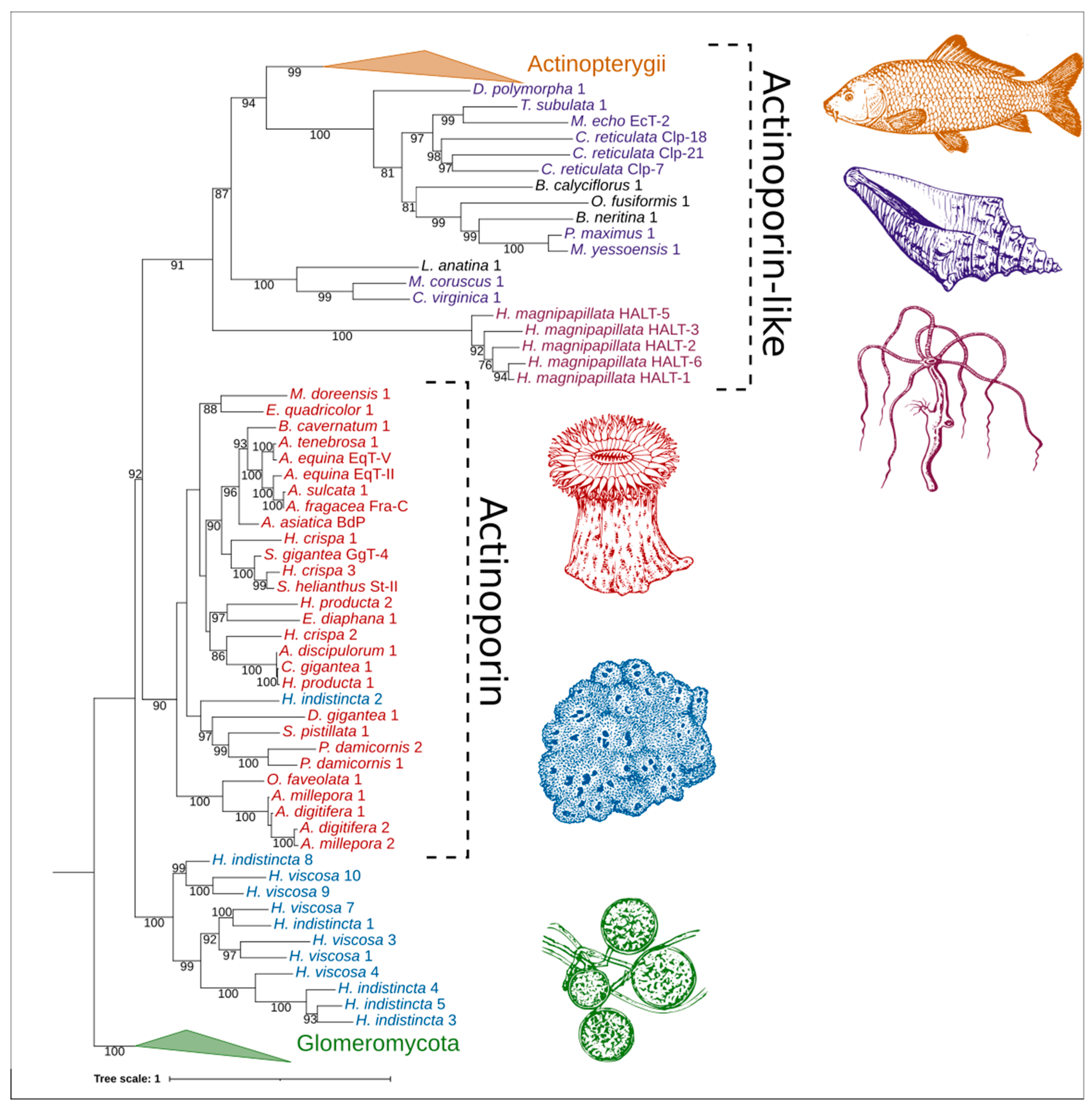
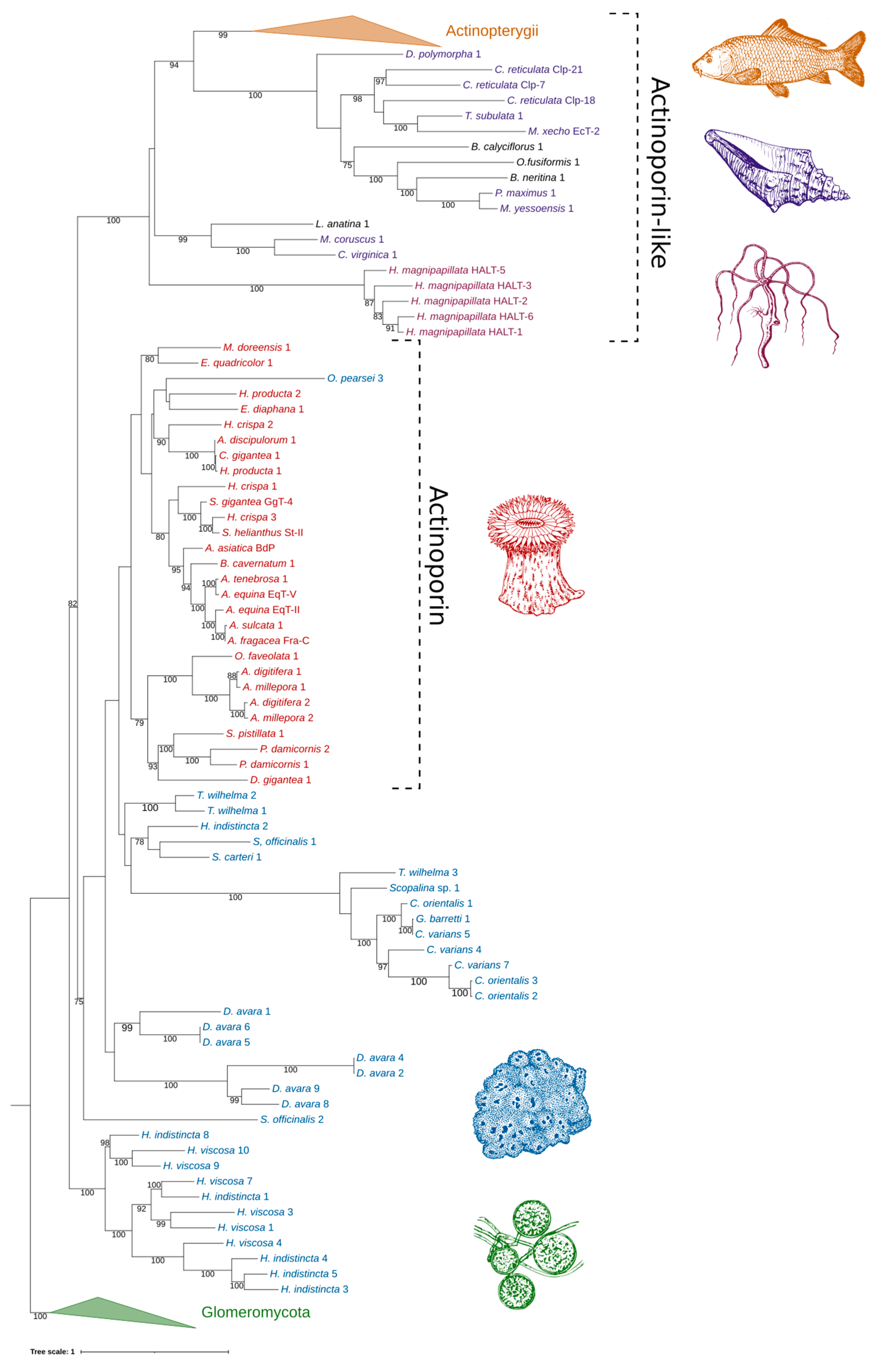
2.4. Phylogenetic Analysis
3. Discussion
4. Materials and Methods
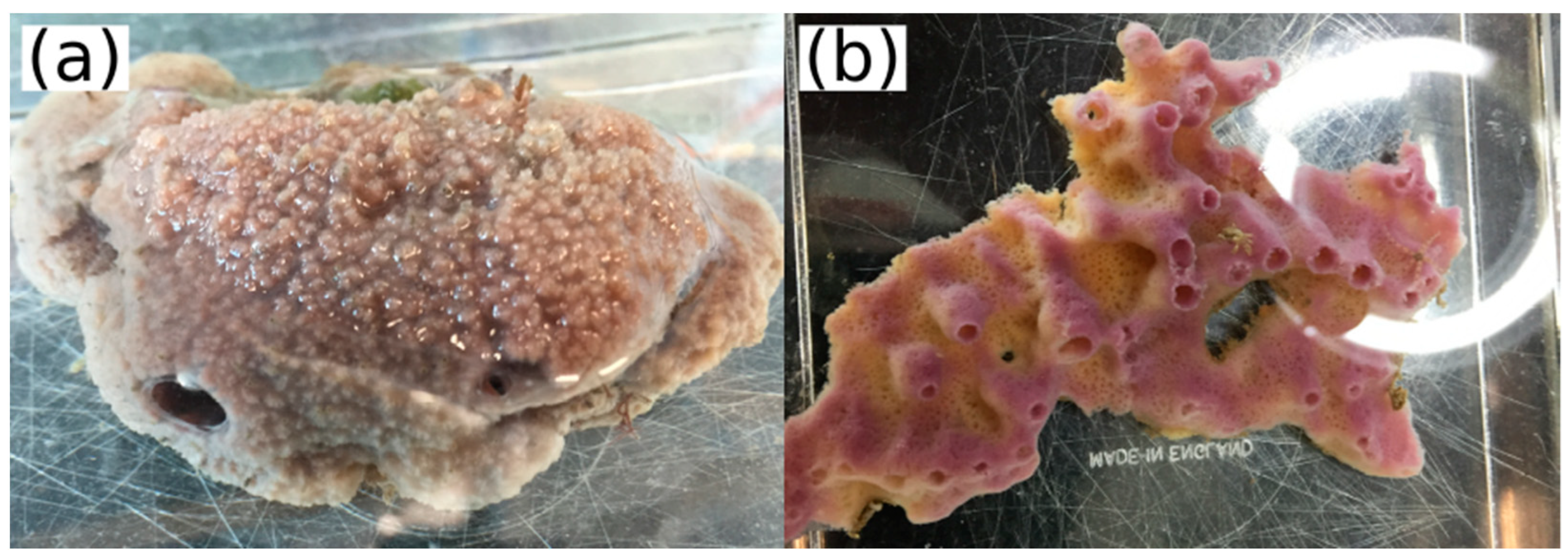
4.1. Sample Collection
4.2. RNA Extraction
4.3. Transcriptome Sequencing
4.4. Transcriptome Assembly
4.5. Identification of Novel Actinoporin-like Proteins from Sea Sponges
4.6. Sequence Analysis and Structural Prediction
4.7. Phylogenetic Analysis of Actinoporin-like Proteins from Sea Sponges
4.8. Generation of Figures
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- Kem, W.R. Sea Anemone Toxins: Structure and Action. Biol. Nematocysts 1988, 375–405. [Google Scholar]
- Anderluh, G.; Maček, P. Cytolytic Peptide and Protein Toxins from Sea Anemones (Anthozoa: Actiniaria). Toxicon 2002, 40, 111–124. [Google Scholar] [CrossRef]
- Athanasiadis, A.; Anderluh, G.; Maček, P.; Turk, D. Crystal Structure of the Soluble Form of Equinatoxin II, a Pore-Forming Toxin from the Sea Anemone Actinia Equina. Structure 2001, 9, 341–346. [Google Scholar] [CrossRef] [Green Version]
- Mancheño, J.M.; Martín-Benito, J.; Martínez-Ripoll, M.; Gavilanes, J.G.; Hermoso, J.A. Crystal and Electron Microscopy Structures of Sticholysin II Actinoporin Reveal Insights into the Mechanism of Membrane Pore Formation. Structure 2003, 11, 1319–1328. [Google Scholar] [CrossRef] [Green Version]
- Mechaly, A.E.; Bellomio, A.; Gil-Cartón, D.; Morante, K.; Valle, M.; González-Mañas, J.M.; Guérin, D.M.A. Structural Insights into the Oligomerization and Architecture of Eukaryotic Membrane Pore-Forming Toxins. Structure 2011, 19, 181–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakrač, B.; Gutiérrez-Aguirre, I.; Podlesek, Z.; Sonnen, A.F.-P.; Gilbert, R.J.C.; Maček, P.; Lakey, J.H.; Anderluh, G. Molecular Determinants of Sphingomyelin Specificity of a Eukaryotic Pore-Forming Toxin. J. Biol. Chem. 2008, 283, 18665–18677. [Google Scholar] [CrossRef] [Green Version]
- Bakrač, B.; Kladnik, A.; Maček, P.; McHaffie, G.; Werner, A.; Lakey, J.H.; Anderluh, G. A Toxin-Based Probe Reveals Cytoplasmic Exposure of Golgi Sphingomyelin. J. Biol. Chem. 2010, 285, 22186–22195. [Google Scholar] [CrossRef] [Green Version]
- Malovrh, P.; Viero, G.; Serra, M.D.; Podlesek, Z.; Lakey, J.H.; Maček, P.; Menestrina, G.; Anderluh, G. A Novel Mechanism of Pore Formation: Membrane Penetration by the N-Terminal Amphipathic Region of Equinatoxin. J. Biol. Chem. 2003, 278, 22678–22685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojko, N.; Kristan, K.Č.; Viero, G.; Žerovnik, E.; Maček, P.; Serra, M.D.; Anderluh, G. Membrane Damage by an α-Helical Pore-Forming Protein, Equinatoxin II, Proceeds through a Succession of Ordered Step. J. Biol. Chem. 2013, 288, 23704–23715. [Google Scholar] [CrossRef] [Green Version]
- Drechsler, A.; Potrich, C.; Sabo, J.K.; Frisanco, M.; Guella, G.; Dalla Serra, M.; Anderluh, G.; Separovic, F.; Norton, R.S. Structure and Activity of the N-Terminal Region of the Eukaryotic Cytolysin Equinatoxin II. Biochemistry 2006, 45, 1818–1828. [Google Scholar] [CrossRef] [PubMed]
- Lam, Y.H.; Hung, A.; Norton, R.S.; Separovic, F.; Watts, A. Solid-State NMR and Simulation Studies of Equinatoxin II N-Terminus Interaction with Lipid Bilayers. Proteins Struct. Funct. Bioinform. 2010, 78, 858–872. [Google Scholar] [CrossRef]
- García-Linares, S.; Richmond, R.; García-Mayoral, M.F.; Bustamante, N.; Bruix, M.; Gavilanes, J.G.; Martínez-del-Pozo, Á. The Sea Anemone Actinoporin (Arg-Gly-Asp) Conserved Motif Is Involved in Maintaining the Competent Oligomerization State of These Pore-Forming Toxins. FEBS J. 2014, 281, 1465–1478. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Caaveiro, J.M.M.; Morante, K.; González-Mañas, J.M.; Tsumoto, K. Structural Basis for Self-Assembly of a Cytolytic Pore Lined by Protein and Lipid. Nat. Commun. 2015, 6, 6337. [Google Scholar] [CrossRef] [Green Version]
- Črnigoj Kristan, K.; Viero, G.; Dalla Serra, M.; Maček, P.; Anderluh, G. Molecular Mechanism of Pore Formation by Actinoporins. Toxicon 2009, 54, 1125–1134. [Google Scholar] [CrossRef] [PubMed]
- Rojko, N.; Dalla Serra, M.; Maček, P.; Anderluh, G. Pore Formation by Actinoporins, Cytolysins from Sea Anemones. Biochim. Biophys. Acta (BBA)-Biomembr. 2016, 1858, 446–456. [Google Scholar] [CrossRef] [PubMed]
- Mutter, N.L.; Soskine, M.; Huang, G.; Albuquerque, I.S.; Bernardes, G.J.L.; Maglia, G. Modular Pore-Forming Immunotoxins with Caged Cytotoxicity Tailored by Directed Evolution. ACS Chem. Biol. 2018, 13, 3153–3160. [Google Scholar] [CrossRef]
- Wloka, C.; Mutter, N.L.; Soskine, M.; Maglia, G. Alpha-Helical Fragaceatoxin C Nanopore Engineered for Double-Stranded and Single-Stranded Nucleic Acid Analysis. Angew. Chem. Int. Ed. 2016, 55, 12494–12498. [Google Scholar] [CrossRef]
- Makino, A.; Abe, M.; Murate, M.; Inaba, T.; Yilmaz, N.; Hullin-Matsuda, F.; Kishimoto, T.; Schieber, N.L.; Taguchi, T.; Arai, H.; et al. Visualization of the Heterogeneous Membrane Distribution of Sphingomyelin Associated with Cytokinesis, Cell Polarity, and Sphingolipidosis. FASEB J. 2015, 29, 477–493. [Google Scholar] [CrossRef]
- Laborde, R.J.; Sanchez-Ferras, O.; Luzardo, M.C.; Cruz-Leal, Y.; Fernández, A.; Mesa, C.; Oliver, L.; Canet, L.; Abreu-Butin, L.; Nogueira, C.V.; et al. Novel Adjuvant Based on the Pore-Forming Protein Sticholysin II Encapsulated into Liposomes Effectively Enhances the Antigen-Specific CTL-Mediated Immune Response. J. Immunol. 2017, 198, 2772–2784. [Google Scholar] [CrossRef] [Green Version]
- Macrander, J.; Daly, M. Evolution of the Cytolytic Pore-Forming Proteins (Actinoporins) in Sea Anemones. Toxins 2016, 8, 368. [Google Scholar] [CrossRef] [Green Version]
- Ben-Ari, H.; Paz, M.; Sher, D. The Chemical Armament of Reef-Building Corals: Inter- and Intra-Specific Variation and the Identification of an Unusual Actinoporin in Stylophora Pistilata. Sci. Rep. 2018, 8, 251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez-Aguirre, I.; Trontelj, P.; Maček, P.; Lakey, J.H.; Anderluh, G. Membrane Binding of Zebrafish Actinoporin-like Protein: AF Domains, a Novel Superfamily of Cell Membrane Binding Domains. Biochem. J. 2006, 398, 381–392. [Google Scholar] [CrossRef] [Green Version]
- von Reumont, B.M.; Campbell, L.I.; Richter, S.; Hering, L.; Sykes, D.; Hetmank, J.; Jenner, R.A.; Bleidorn, C. A Polychaete’s Powerful Punch: Venom Gland Transcriptomics of Glycera Reveals a Complex Cocktail of Toxin Homologs. Genome Biol. Evol. 2014, 6, 2406–2423. [Google Scholar] [CrossRef] [PubMed]
- Gerdol, M.; Luo, Y.-J.; Satoh, N.; Pallavicini, A. Genetic and Molecular Basis of the Immune System in the Brachiopod Lingula Anatina. Dev. Comp. Immunol. 2018, 82, 7–30. [Google Scholar] [CrossRef]
- Gerdol, M.; Cervelli, M.; Oliverio, M.; Modica, M.V. Piercing Fishes: Porin Expansion and Adaptation to Hematophagy in the Vampire Snail Cumia Reticulata. Mol. Biol. Evol. 2018, 35, 2654–2668. [Google Scholar] [CrossRef]
- Kawashima, Y.; Nagai, H.; Ishida, M.; Nagashima, Y.; Shiomi, K. Primary Structure of Echotoxin 2, an Actinoporin-like Hemolytic Toxin from the Salivary Gland of the Marine Gastropod Monoplex Echo. Toxicon 2003, 42, 491–497. [Google Scholar] [CrossRef]
- Hoang, Q.T.; Cho, S.H.; McDaniel, S.F.; Ok, S.H.; Quatrano, R.S.; Shin, J.S. An Actinoporin Plays a Key Role in Water Stress in the Moss Physcomitrella Patens. New Phytol. 2009, 184, 502–510. [Google Scholar] [CrossRef]
- Takara, T.; Nakagawa, T.; Isobe, M.; Okino, N.; Ichinose, S.; Omori, A.; Ito, M. Purification, Molecular Cloning, and Application of a Novel Sphingomyelin-Binding Protein (Clamlysin) from the Brackishwater Clam, Corbicula Japonica. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2011, 1811, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Glasser, E.; Rachamim, T.; Aharonovich, D.; Sher, D. Hydra Actinoporin-like Toxin-1, an Unusual Hemolysin from the Nematocyst Venom of Hydra Magnipapillata Which Belongs to an Extended Gene Family. Toxicon 2014, 91, 103–113. [Google Scholar] [CrossRef]
- Luter, H.M.; Bannister, R.J.; Whalan, S.; Kutti, T.; Pineda, M.-C.; Webster, N.S. Microbiome Analysis of a Disease Affecting the Deep-Sea Sponge Geodia Barretti. FEMS Microbiol. Ecol. 2017, 93, fix074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Syue, S.-T.; Hsu, C.-H.; Soong, K. Testing of How and Why the Terpios Hoshinota Sponge Kills Stony Corals. Sci. Rep. 2021, 11, 7661. [Google Scholar] [CrossRef]
- Padilla Verdín, C.J.; Carballo, J.L.; Camacho, M.L. Qualitative Assessment of Sponge-Feeding Organisms from the Mexican Pacific Coast. Open Mar. Biol. J. 2010, 4. [Google Scholar] [CrossRef] [Green Version]
- Laport, M.S.; Santos, O.C.S.; Muricy, G. Marine Sponges: Potential Sources of New Antimicrobial Drugs. Curr. Pharm. Biotechnol. 2009, 10, 86–105. [Google Scholar] [CrossRef]
- Qi, S.-H.; Ma, X. Antifouling Compounds from Marine Invertebrates. Mar. Drugs 2017, 15, 263. [Google Scholar] [CrossRef] [Green Version]
- Calcabrini, C.; Catanzaro, E.; Bishayee, A.; Turrini, E.; Fimognari, C. Marine Sponge Natural Products with Anticancer Potential: An Updated Review. Mar. Drugs 2017, 15, 310. [Google Scholar] [CrossRef] [Green Version]
- Müller, W.E.G.; Wang, X.; Binder, M.; von Lintig, J.; Wiens, M.; Schröder, H.C. Differential Expression of the Demosponge (Suberites Domuncula) Carotenoid Oxygenases in Response to Light: Protection Mechanism Against the Self-Produced Toxic Protein (Suberitine). Mar. Drugs 2012, 10, 177–199. [Google Scholar] [CrossRef] [Green Version]
- do Nascimento-Neto, L.G.; Cabral, M.G.; Carneiro, R.F.; Silva, Z.; Arruda, F.V.S.; Nagano, C.S.; Fernandes, A.R.; Sampaio, A.H.; Teixeira, E.H.; Videira, P.A. Halilectin-3, a Lectin from the Marine Sponge Haliclona Caerulea, Induces Apoptosis and Autophagy in Human Breast Cancer MCF7 Cells Through Caspase-9 Pathway and LC3-II Protein Expression. Anti-Cancer Agents Med. Chem. (Former. Curr. Med. Chem.—Anti-Cancer Agents) 2018, 18, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Scarfì, S.; Pozzolini, M.; Oliveri, C.; Mirata, S.; Salis, A.; Damonte, G.; Fenoglio, D.; Altosole, T.; Ilan, M.; Bertolino, M.; et al. Identification, Purification and Molecular Characterization of Chondrosin, a New Protein with Anti-Tumoral Activity from the Marine Sponge Chondrosia Reniformis Nardo 1847. Mar. Drugs 2020, 18, 409. [Google Scholar] [CrossRef] [PubMed]
- Mangel, A.; Leitão, J.M.; Batel, R.; Zimmermann, H.; Müller, W.E.G.; Schröder, H.C. Purification and Characterization of a Pore-Forming Protein from the Marine Sponge Tethya Lyncurium. Eur. J. Biochem. 1992, 210, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Wiens, M.; Korzhev, M.; Krasko, A.; Thakur, N.L.; Perović-Ottstadt, S.; Breter, H.J.; Ushijima, H.; Diehl-Seifert, B.; Müller, I.M.; Müller, W.E.G. Innate Immune Defense of the Sponge Suberites Domuncula against Bacteria Involves a MyD88-Dependent Signaling Pathway: Induction of a Perforin-like Molecule. J. Biol. Chem. 2005, 280, 27949–27959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nichols, S.A.; Roberts, B.W.; Richter, D.J.; Fairclough, S.R.; King, N. Origin of Metazoan Cadherin Diversity and the Antiquity of the Classical Cadherin/β-Catenin Complex. Proc. Natl. Acad. Sci. USA 2012, 109, 13046–13051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ereskovsky, A.V.; Richter, D.J.; Lavrov, D.V.; Schippers, K.J.; Nichols, S.A. Transcriptome Sequencing and Delimitation of Sympatric Oscarella Species (O. carmela and O. pearsei sp. Nov) from California, USA. PLoS ONE 2017, 12, e0183002. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Camacho, J.M.; Doonan, L.; McCormack, G.P. Evolution of the Main Skeleton-Forming Genes in Sponges (Phylum Porifera) with Special Focus on the Marine Haplosclerida (Class Demospongiae). Mol. Phylogenet. Evol. 2019, 131, 245–253. [Google Scholar] [CrossRef]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 Improves Signal Peptide Predictions Using Deep Neural Networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Anderluh, G.; Barlič, A.; Potrich, C.; Maček, P.; Menestrina, G. Lysine 77 Is a Key Residue in Aggregation of Equinatoxin II, a Pore-Forming Toxin from Sea Anemone Actinia Equina. J. Membr. Biol. 2000, 173, 47–55. [Google Scholar] [CrossRef]
- Morante, K.; Caaveiro, J.M.M.; Viguera, A.R.; Tsumoto, K.; González-Mañas, J.M. Functional Characterization of Val60, a Key Residue Involved in the Membrane-Oligomerization of Fragaceatoxin C, an Actinoporin from Actinia Fragacea. FEBS Lett. 2015, 589, 1840–1846. [Google Scholar] [CrossRef] [Green Version]
- Peraro, M.D.; van der Goot, F.G. Pore-Forming Toxins: Ancient, but Never Really out of Fashion. Nat. Rev. Microbiol. 2016, 14, 77–92. [Google Scholar] [CrossRef] [PubMed]
- Spicer, B.A.; Conroy, P.J.; Law, R.H.P.; Voskoboinik, I.; Whisstock, J.C. Perforin—A Key (Shaped) Weapon in the Immunological Arsenal. Semin. Cell Dev. Biol. 2017, 72, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Basulto, A.; Pérez, V.M.; Noa, Y.; Varela, C.; Otero, A.J.; Pico, M.C. Immunohistochemical Targeting of Sea Anemone Cytolysins on Tentacles, Mesenteric Filaments and Isolated Nematocysts of Stichodactyla Helianthus. J. Exp. Zool. Part A Comp. Exp. Biol. 2006, 305A, 253–258. [Google Scholar] [CrossRef]
- Koutsouveli, V.; Cárdenas, P.; Santodomingo, N.; Marina, A.; Morato, E.; Rapp, H.T.; Riesgo, A. The Molecular Machinery of Gametogenesis in Geodia Demosponges (Porifera): Evolutionary Origins of a Conserved Toolkit across Animals. Mol. Biol. Evol. 2020, 37, 3485–3506. [Google Scholar] [CrossRef]
- Srivastava, M.; Simakov, O.; Chapman, J.; Fahey, B.; Gauthier, M.E.A.; Mitros, T.; Richards, G.S.; Conaco, C.; Dacre, M.; Hellsten, U.; et al. The Amphimedon Queenslandica Genome and the Evolution of Animal Complexity. Nature 2010, 466, 720–726. [Google Scholar] [CrossRef]
- Hong, Q.; Gutiérrez-Aguirre, I.; Barlič, A.; Malovrh, P.; Kristan, K.; Podlesek, Z.; Maček, P.; Turk, D.; González-Mañas, J.M.; Lakey, J.H.; et al. Two-Step Membrane Binding by Equinatoxin II, a Pore-Forming Toxin from the Sea Anemone, Involves an Exposed Aromatic Cluster and a Flexible Helix. J. Biol. Chem. 2002, 277, 41916–41924. [Google Scholar] [CrossRef] [Green Version]
- Anderluh, G.; Razpotnik, A.; Podlesek, Z.; Maček, P.; Separovic, F.; Norton, R.S. Interaction of the Eukaryotic Pore-Forming Cytolysin Equinatoxin II with Model Membranes: 19F NMR Studies. J. Mol. Biol. 2005, 347, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Valle, A.; Alvarado-Mesén, J.; Lanio, M.E.; Álvarez, C.; Barbosa, J.A.R.G.; Pazos, I.F. The Multigene Families of Actinoporins (Part I): Isoforms and Genetic Structure. Toxicon 2015, 103, 176–187. [Google Scholar] [CrossRef]
- Jiang, X.; Chen, H.; Yang, W.; Liu, Y.; Liu, W.; Wei, J.; Tu, H.; Xie, X.; Wang, L.; Xu, A. Functional Expression and Characterization of an Acidic Actinoporin from Sea Anemone Sagartia Rosea. Biochem. Biophys. Res. Commun. 2003, 312, 562–570. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Trinity: Reconstructing a Full-Length Transcriptome without a Genome from RNA-Seq Data. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De Novo Transcript Sequence Reconstruction from RNA-Seq Using the Trinity Platform for Reference Generation and Analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- O’Donovan, C.; Martin, M.J.; Gattiker, A.; Gasteiger, E.; Bairoch, A.; Apweiler, R. High-Quality Protein Knowledge Resource: SWISS-PROT and TrEMBL. Brief. Bioinform. 2002, 3, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER Web Server: Interactive Sequence Similarity Searching. Nucleic Acids Res. 2011, 39, W29–W37. [Google Scholar] [CrossRef] [Green Version]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The Protein Families Database. Nucleic Acids Res. 2014, 42, D222–D230. [Google Scholar] [CrossRef] [Green Version]
- Simão, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: Assessing Genome Assembly and Annotation Completeness with Single-Copy Orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef] [Green Version]
- Kenny, N.J.; Francis, W.R.; Rivera-Vicéns, R.E.; Juravel, K.; de Mendoza, A.; Díez-Vives, C.; Lister, R.; Bezares-Calderón, L.A.; Grombacher, L.; Roller, M.; et al. Tracing Animal Genomic Evolution with the Chromosomal-Level Assembly of the Freshwater Sponge Ephydatia Muelleri. Nat. Commun. 2020, 11, 3676. [Google Scholar] [CrossRef]
- Kenny, N.J.; Plese, B.; Riesgo, A.; Itskovich, V.B. Symbiosis, Selection, and Novelty: Freshwater Adaptation in the Unique Sponges of Lake Baikal. Mol. Biol. Evol. 2019, 36, 2462–2480. [Google Scholar] [CrossRef] [PubMed]
- Francis, W.R.; Eitel, M.; Vargas, S.; Adamski, M.; Haddock, S.H.D.; Krebs, S.; Blum, H.; Erpenbeck, D.; Wörheide, G. The Genome of the Contractile Demosponge Tethya Wilhelma and the Evolution of Metazoan Neural Signalling Pathways. bioRxiv 2017, 120998. [Google Scholar] [CrossRef] [Green Version]
- Ryu, T.; Seridi, L.; Moitinho-Silva, L.; Oates, M.; Liew, Y.J.; Mavromatis, C.; Wang, X.; Haywood, A.; Lafi, F.F.; Kupresanin, M.; et al. Hologenome Analysis of Two Marine Sponges with Different Microbiomes. BMC Genom. 2016, 17, 158. [Google Scholar] [CrossRef] [Green Version]
- Fortunato, S.A.V.; Adamski, M.; Ramos, O.M.; Leininger, S.; Liu, J.; Ferrier, D.E.K.; Adamska, M. Calcisponges Have a ParaHox Gene and Dynamic Expression of Dispersed NK Homeobox Genes. Nature 2014, 514, 620–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finoshin, A.D.; Adameyko, K.I.; Mikhailov, K.V.; Kravchuk, O.I.; Georgiev, A.A.; Gornostaev, N.G.; Kosevich, I.A.; Mikhailov, V.S.; Gazizova, G.R.; Shagimardanova, E.I.; et al. Iron Metabolic Pathways in the Processes of Sponge Plasticity. PLoS ONE 2020, 15, e0228722. [Google Scholar] [CrossRef] [Green Version]
- Manousaki, T.; Koutsouveli, V.; Lagnel, J.; Kollias, S.; Tsigenopoulos, C.S.; Arvanitidis, C.; Magoulas, A.; Dounas, C.; Dailianis, T. A de Novo Transcriptome Assembly for the Bath Sponge Spongia Officinalis, Adjusting for Microsymbionts. BMC Res. Notes 2019, 12, 813. [Google Scholar] [CrossRef] [Green Version]
- González-Aravena, M.; Kenny, N.J.; Osorio, M.; Font, A.; Riesgo, A.; Cárdenas, C.A. Warm Temperatures, Cool Sponges: The Effect of Increased Temperatures on the Antarctic Sponge Isodictya sp. PeerJ 2019, 7, e8088. [Google Scholar] [CrossRef] [Green Version]
- Leiva, C.; Taboada, S.; Kenny, N.J.; Combosch, D.; Giribet, G.; Jombart, T.; Riesgo, A. Population Substructure and Signals of Divergent Adaptive Selection despite Admixture in the Sponge Dendrilla Antarctica from Shallow Waters Surrounding the Antarctic Peninsula. Mol. Ecol. 2019, 28, 3151–3170. [Google Scholar] [CrossRef] [Green Version]
- Pita, L.; Hoeppner, M.P.; Ribes, M.; Hentschel, U. Differential Expression of Immune Receptors in Two Marine Sponges upon Exposure to Microbial-Associated Molecular Patterns. Sci. Rep. 2018, 8, 16081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Revilla-i-Domingo, R.; Schmidt, C.; Zifko, C.; Raible, F. Establishment of Transgenesis in the Demosponge Suberites Domuncula. Genetics 2018, 210, 435–443. [Google Scholar] [CrossRef] [Green Version]
- Kenny, N.J.; de Goeij, J.M.; de Bakker, D.M.; Whalen, C.G.; Berezikov, E.; Riesgo, A. Towards the Identification of Ancestrally Shared Regenerative Mechanisms across the Metazoa: A Transcriptomic Case Study in the Demosponge Halisarca Caerulea. Mar. Genom. 2018, 37, 135–147. [Google Scholar] [CrossRef] [Green Version]
- Leys, S. Ephydatia Muelleri Trinity Transcriptome. Available online: https://era.library.ualberta.ca/items/d643c77d-aefd-45f3-8ddb-985189eebcf2 (accessed on 2 April 2021).
- Leys, S. Eunapius Fragilis Trinity Transcriptome. Available online: https://era.library.ualberta.ca/items/6139a88f-895d-44a7-bd0d-e22d455d2785 (accessed on 2 April 2021).
- Simion, P.; Philippe, H.; Baurain, D.; Jager, M.; Richter, D.J.; Di Franco, A.; Roure, B.; Satoh, N.; Quéinnec, É.; Ereskovsky, A.; et al. A Large and Consistent Phylogenomic Dataset Supports Sponges as the Sister Group to All Other Animals. Curr. Biol. 2017, 27, 958–967. [Google Scholar] [CrossRef] [Green Version]
- Díez-Vives, C.; Moitinho-Silva, L.; Nielsen, S.; Reynolds, D.; Thomas, T. Expression of Eukaryotic-like Protein in the Microbiome of Sponges. Mol. Ecol. 2017, 26, 1432–1451. [Google Scholar] [CrossRef]
- Borisenko, I.; Adamski, M.; Ereskovsky, A.; Adamska, M. Surprisingly Rich Repertoire of Wnt Genes in the Demosponge Halisarca Dujardini. BMC Evol. Biol. 2016, 16, 123. [Google Scholar] [CrossRef] [Green Version]
- Guzman, C.; Conaco, C. Comparative Transcriptome Analysis Reveals Insights into the Streamlined Genomes of Haplosclerid Demosponges. Sci. Rep. 2016, 6, 18774. [Google Scholar] [CrossRef] [PubMed]
- Alié, A.; Hayashi, T.; Sugimura, I.; Manuel, M.; Sugano, W.; Mano, A.; Satoh, N.; Agata, K.; Funayama, N. The Ancestral Gene Repertoire of Animal Stem Cells. Proc. Natl. Acad. Sci. USA 2015, 112, E7093–E7100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, F.; Ding, S.; Ou, H.; Wang, D.; Chen, J.; Miyamoto, M.M. Transcriptome Changes during the Life Cycle of the Red Sponge, Mycale Phyllophila (Porifera, Demospongiae, Poecilosclerida). Genes 2015, 6, 1023–1052. [Google Scholar] [CrossRef] [Green Version]
- Whelan, N.V.; Kocot, K.M.; Moroz, L.L.; Halanych, K.M. Error, Signal, and the Placement of Ctenophora Sister to All Other Animals. Proc. Natl. Acad. Sci. USA 2015, 112, 5773–5778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riesgo, A.; Peterson, K.; Richardson, C.; Heist, T.; Strehlow, B.; McCauley, M.; Cotman, C.; Hill, M.; Hill, A. Transcriptomic Analysis of Differential Host Gene Expression upon Uptake of Symbionts: A Case Study with Symbiodinium and the Major Bioeroding Sponge Cliona Varians. BMC Genom. 2014, 15, 376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riesgo, A.; Farrar, N.; Windsor, P.J.; Giribet, G.; Leys, S.P. The Analysis of Eight Transcriptomes from All Poriferan Classes Reveals Surprising Genetic Complexity in Sponges. Mol. Biol. Evol. 2014, 31, 1102–1120. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The Conserved Domain Database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, N.; Westbrook, M.J.; Young, S.L.; Kuo, A.; Abedin, M.; Chapman, J.; Fairclough, S.; Hellsten, U.; Isogai, Y.; Letunic, I.; et al. The Genome of the Choanoflagellate Monosiga Brevicollis and the Origin of Metazoans. Nature 2008, 451, 783–788. [Google Scholar] [CrossRef] [Green Version]
- Fairclough, S.R.; Chen, Z.; Kramer, E.; Zeng, Q.; Young, S.; Robertson, H.M.; Begovic, E.; Richter, D.J.; Russ, C.; Westbrook, M.J.; et al. Premetazoan Genome Evolution and the Regulation of Cell Differentiation in the Choanoflagellate Salpingoeca Rosetta. Genome Biol. 2013, 14, R15. [Google Scholar] [CrossRef] [Green Version]
- Ryan, J.F.; Pang, K.; Schnitzler, C.E.; Nguyen, A.-D.; Moreland, R.T.; Simmons, D.K.; Koch, B.J.; Francis, W.R.; Havlak, P.; Smith, S.A.; et al. The Genome of the Ctenophore Mnemiopsis Leidyi and Its Implications for Cell Type Evolution. Science 2013, 342, 1242592. [Google Scholar] [CrossRef] [Green Version]
- Moroz, L.L.; Kocot, K.M.; Citarella, M.R.; Dosung, S.; Norekian, T.P.; Povolotskaya, I.S.; Grigorenko, A.P.; Dailey, C.; Berezikov, E.; Buckley, K.M.; et al. The Ctenophore Genome and the Evolutionary Origins of Neural Systems. Nature 2014, 510, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, M.; Begovic, E.; Chapman, J.; Putnam, N.H.; Hellsten, U.; Kawashima, T.; Kuo, A.; Mitros, T.; Salamov, A.; Carpenter, M.L.; et al. The Trichoplax Genome and the Nature of Placozoans. Nature 2008, 454, 955–960. [Google Scholar] [CrossRef] [Green Version]
- Eitel, M.; Francis, W.R.; Varoqueaux, F.; Daraspe, J.; Osigus, H.-J.; Krebs, S.; Vargas, S.; Blum, H.; Williams, G.A.; Schierwater, B.; et al. Comparative Genomics and the Nature of Placozoan Species. PLoS Biol. 2018, 16, e2005359. [Google Scholar] [CrossRef]
- Richter, D.J.; Fozouni, P.; Eisen, M.B.; King, N. Gene Family Innovation, Conservation and Loss on the Animal Stem Lineage. eLife 2018, 7, e34226. [Google Scholar] [CrossRef]
- Pruitt, K.D.; Tatusova, T.; Maglott, D.R. NCBI Reference Sequence (RefSeq): A Curated Non-Redundant Sequence Database of Genomes, Transcripts and Proteins. Nucleic Acids Res. 2005, 33, D501–D504. [Google Scholar] [CrossRef] [Green Version]
- Artimo, P.; Jonnalagedda, M.; Arnold, K.; Baratin, D.; Csardi, G.; de Castro, E.; Duvaud, S.; Flegel, V.; Fortier, A.; Gasteiger, E.; et al. ExPASy: SIB Bioinformatics Resource Portal. Nucleic Acids Res. 2012, 40, W597–W603. [Google Scholar] [CrossRef]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 Web Portal for Protein Modeling, Prediction and Analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uziela, K.; Menéndez Hurtado, D.; Shu, N.; Wallner, B.; Elofsson, A. ProQ3D: Improved Model Quality Assessments Using Deep Learning. Bioinformatics 2017, 33, 1578–1580. [Google Scholar] [CrossRef] [Green Version]
- McGuffin, L.J.; Aldowsari, F.M.F.; Alharbi, S.M.A.; Adiyaman, R. ModFOLD8: Accurate Global and Local Quality Estimates for 3D Protein Models. Nucleic Acids Res. 2021, 49, W425–W430. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Skolnick, J. TM-Align: A Protein Structure Alignment Algorithm Based on the TM-Score. Nucleic Acids Res. 2005, 33, 2302–2309. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A Visualization System for Exploratory Research and Analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Stivala, A.; Wybrow, M.; Wirth, A.; Whisstock, J.C.; Stuckey, P.J. Automatic Generation of Protein Structure Cartoons with Pro-Origami. Bioinformatics 2011, 27, 3315–3316. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version 2—a Multiple Sequence Alignment Editor and Analysis Workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gautier, R.; Douguet, D.; Antonny, B.; Drin, G. HELIQUEST: A Web Server to Screen Sequences with Specific α-Helical Properties. Bioinformatics 2008, 24, 2101–2102. [Google Scholar] [CrossRef]
- Mól, A.R.; Castro, M.S.; Fontes, W. NetWheels: A Web Application to Create High Quality Peptide Helical Wheel and Net Projections. bioRxiv 2018, 416347. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wang, J.; Huang, Y.; Xiao, Y. 3dRNA v2.0: An Updated Web Server for RNA 3D Structure Prediction. Int. J. Mol. Sci. 2019, 20, 4116. [Google Scholar] [CrossRef] [Green Version]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree of Life (ITOL) v5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- GNU Image Manipulation Program 2.10.28. Available online: https://www.gimp.org/ (accessed on 30 December 2020).
- Inkscape 0.92. Available online: https://inkscape.org/ (accessed on 24 January 2020).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Total (Mbp) | Number of Contigs | Number of Trinity ‘Genes’ Excluding Isoforms | Contig N50 (Kbp) | GC (%) |
|---|---|---|---|---|---|
| H. cinerea | 156.12 | 123,111 | 64,261 | 2.81 | 39.59 |
| H. indistincta | 106.87 | 101,413 | 48,788 | 2.03 | 44.10 |
| H. oculata | 142.18 | 122,855 | 70,008 | 2.46 | 38.56 |
| H. simulans | 104.12 | 106,366 | 55,501 | 1.89 | 39.87 |
| H. viscosa | 94.09 | 105,831 | 59,949 | 1.73 | 44.50 |
| Species | Total (aa) | Number of Complete ORFs | Number of 5′ Partial ORFs | Number of 3′ Partial ORFs |
|---|---|---|---|---|
| H. cinerea | 34,243,765 | 58,606 | 14,476 | 6129 |
| H. indistincta | 26,553,534 | 29,232 | 15,794 | 7290 |
| H. oculata | 30,530,592 | 49,528 | 16,191 | 5687 |
| H. simulans | 24,643,377 | 31,081 | 16,112 | 7561 |
| H. viscosa | 22,299,218 | 24,435 | 12,776 | 7166 |
| Species | Complete (%) | Single (%) | Duplicate (%) | Fragmented (%) | Missing (%) |
|---|---|---|---|---|---|
| H. cinerea | 97.6 | 23.9 | 73.7 | 0.8 | 1.6 |
| H. indistincta | 99.6 | 45.1 | 54.5 | 0.0 | 0.4 |
| H. oculata | 98.5 | 31.4 | 67.1 | 0.4 | 1.1 |
| H. simulans | 97.6 | 38.0 | 59.6 | 2.0 | 0.4 |
| H. viscosa | 95.3 | 55.3 | 40.0 | 4.3 | 0.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sandoval, K.; McCormack, G.P. Actinoporin-like Proteins Are Widely Distributed in the Phylum Porifera. Mar. Drugs 2022, 20, 74. https://doi.org/10.3390/md20010074
Sandoval K, McCormack GP. Actinoporin-like Proteins Are Widely Distributed in the Phylum Porifera. Marine Drugs. 2022; 20(1):74. https://doi.org/10.3390/md20010074
Chicago/Turabian StyleSandoval, Kenneth, and Grace P. McCormack. 2022. "Actinoporin-like Proteins Are Widely Distributed in the Phylum Porifera" Marine Drugs 20, no. 1: 74. https://doi.org/10.3390/md20010074
APA StyleSandoval, K., & McCormack, G. P. (2022). Actinoporin-like Proteins Are Widely Distributed in the Phylum Porifera. Marine Drugs, 20(1), 74. https://doi.org/10.3390/md20010074