
Crypthecodinium cohnii Growth and Omega Fatty Acid Production in Mediums Supplemented with Extract from Recycled Biomass

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Experiments with Glucose as the Carbon Source
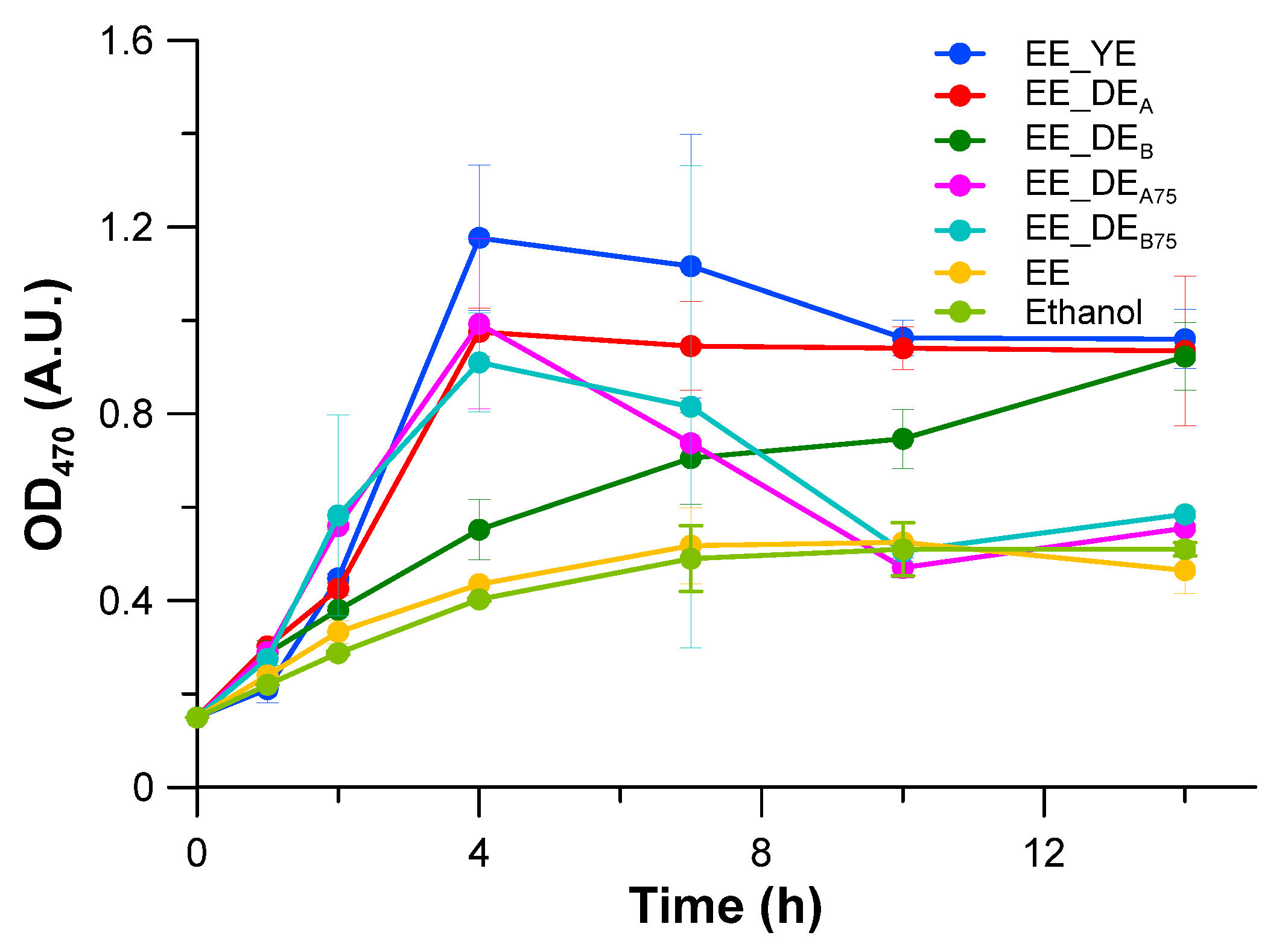
2.2. Experiments with Ethanol as the Carbon Source
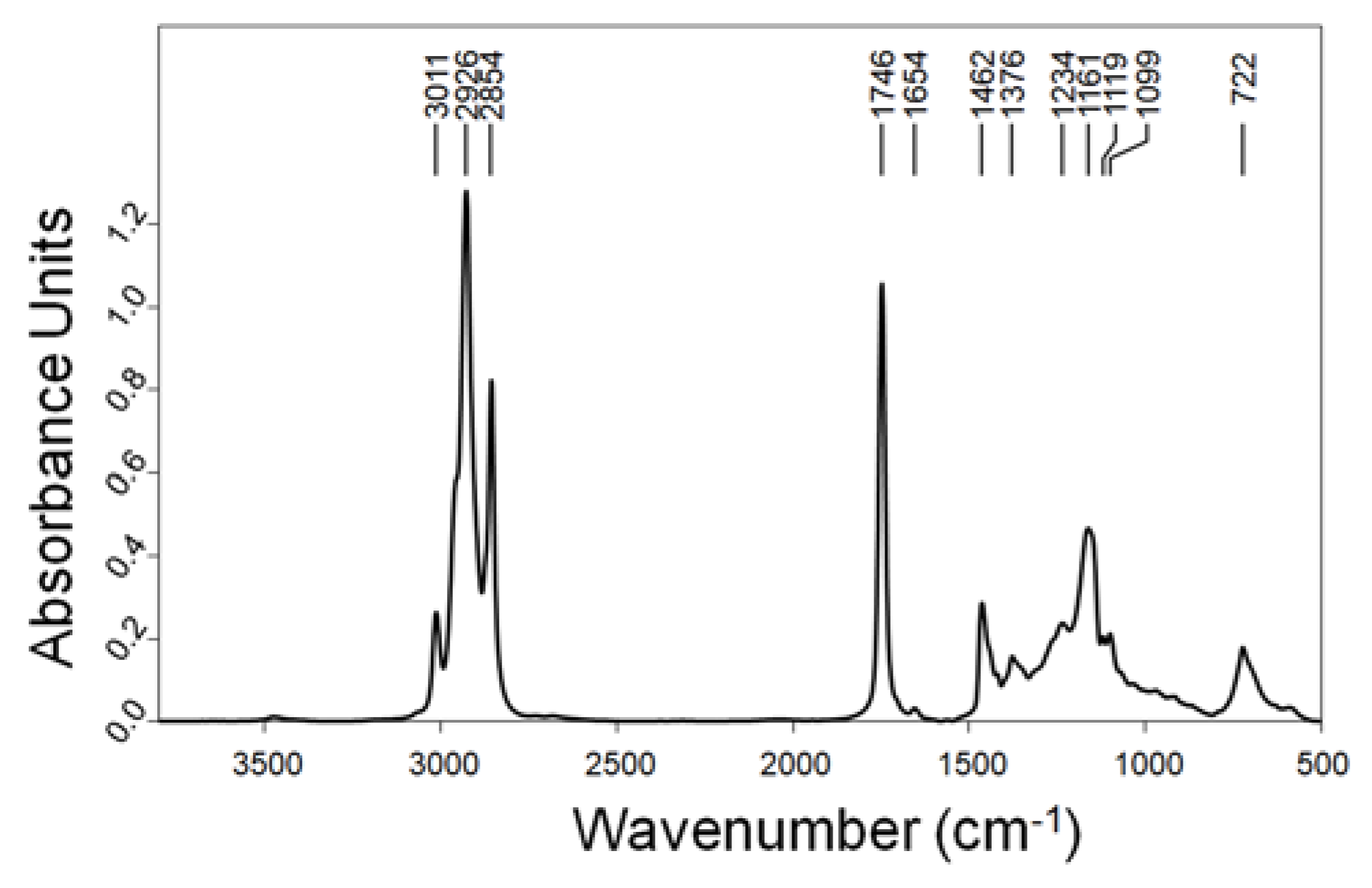
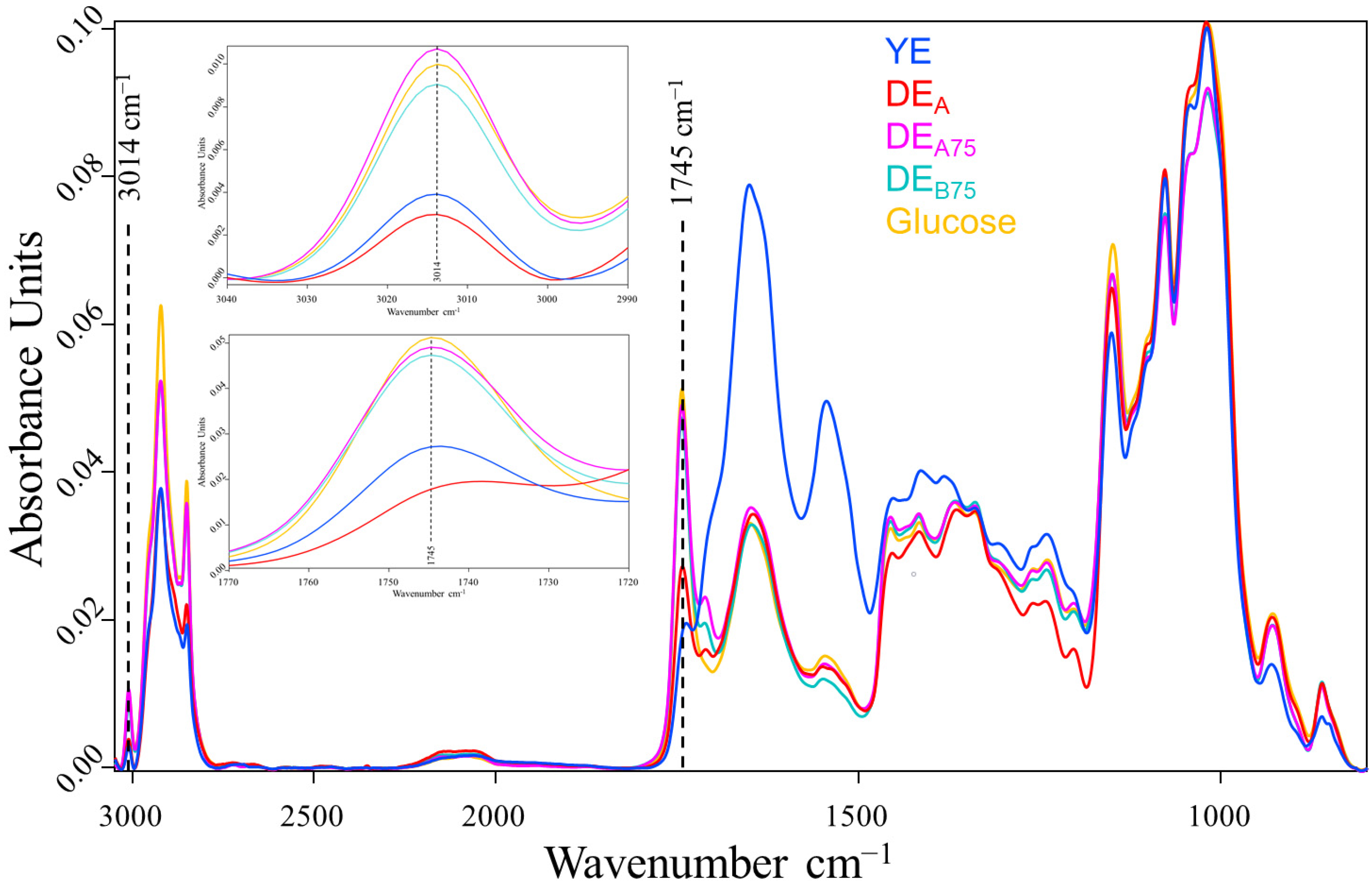
2.3. Evaluation of Lipid/FA and PUFA Accumulation in C. cohnii Biomass by FTIR
3. Discussion
4. Materials and Methods
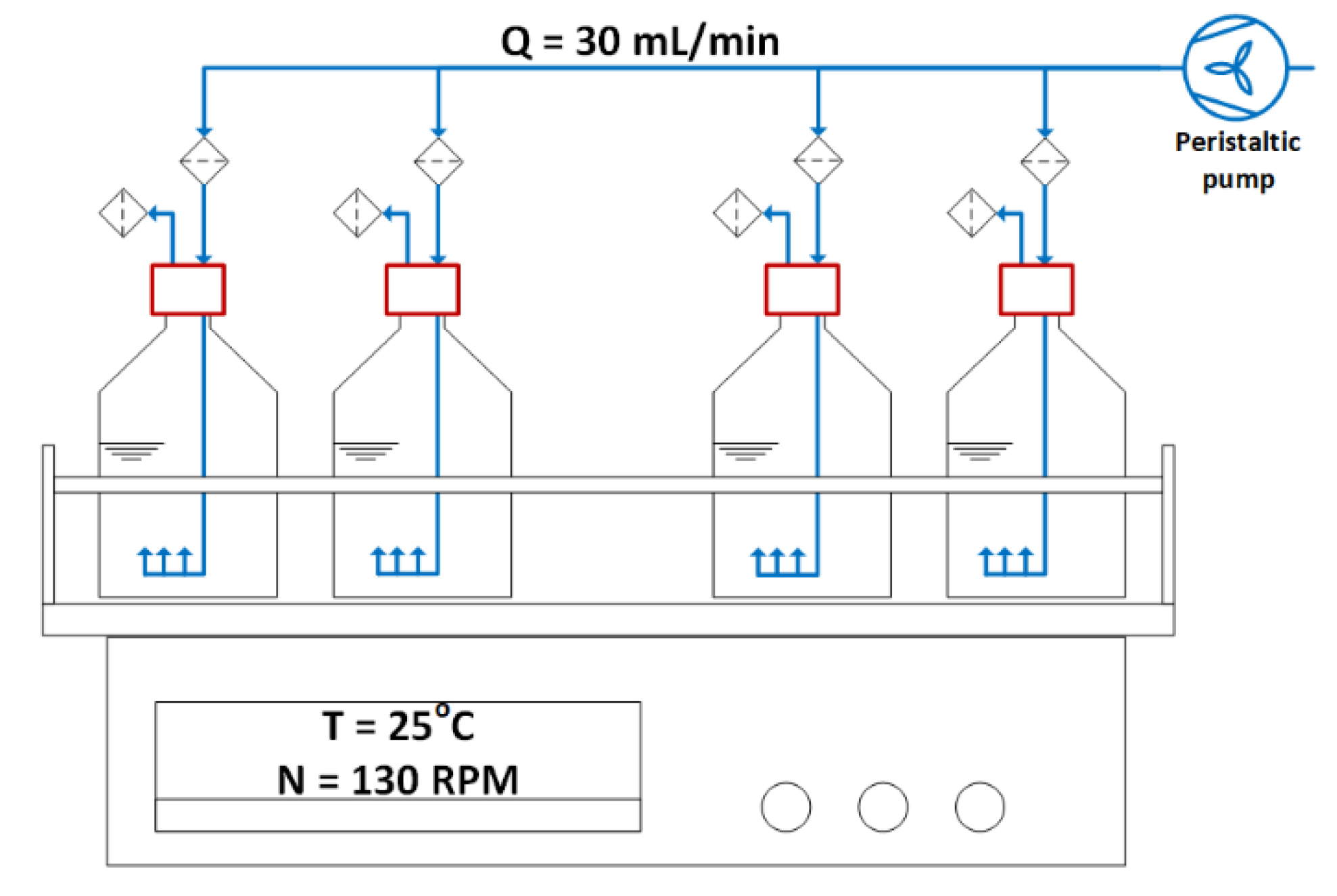
4.1. Cultivation Conditions
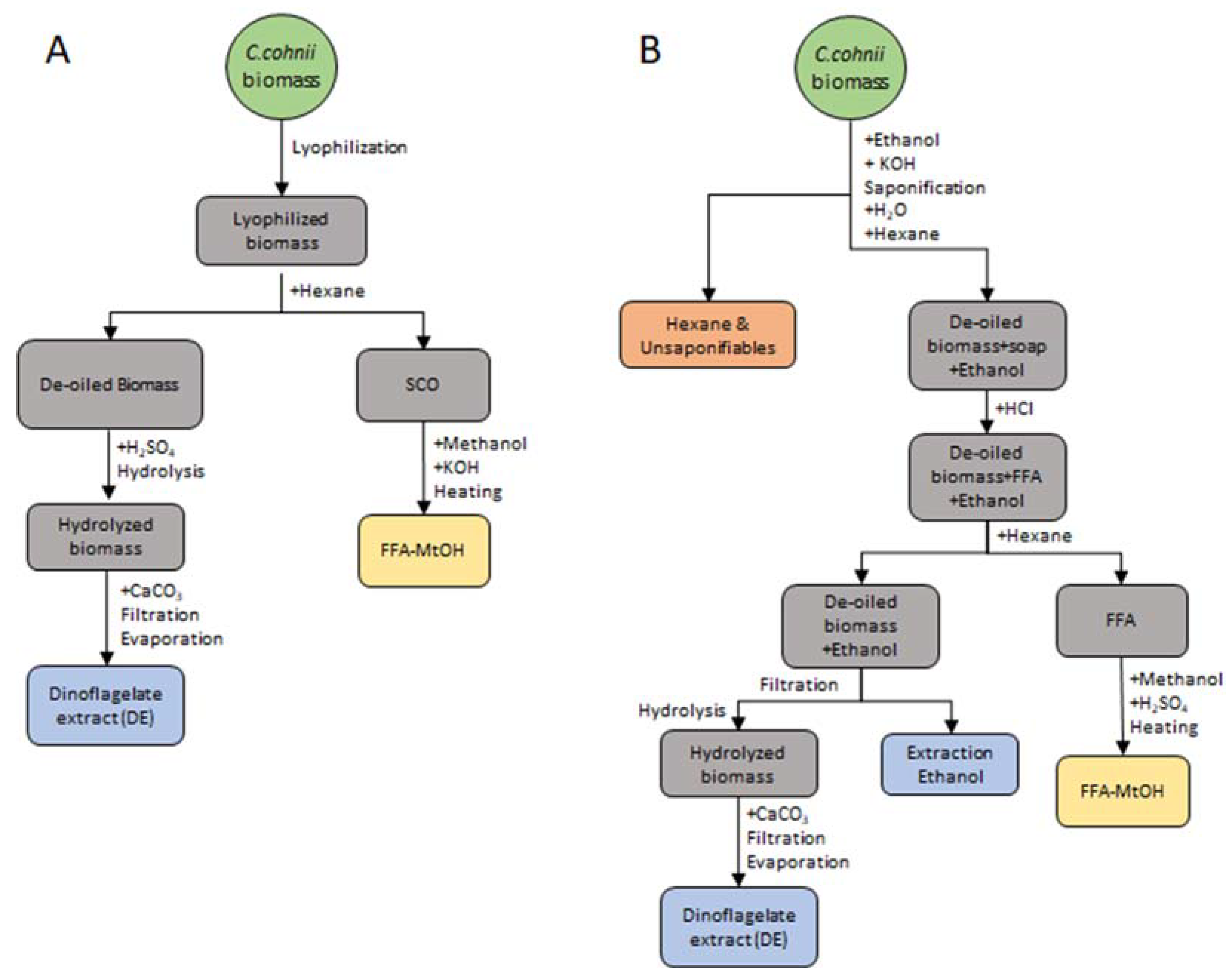
4.2. Dinoflagellate Extract and Extraction Ethanol Obtainment
4.3. Optical Density and Glucose Concentration Measurements
4.4. Determination of Biomass Dry Cell Weight
4.5. Fourier Transform Infrared Spectroscopy
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Adarme-Vega, T.C.; Thomas-Hall, S.R.; Schenk, P.M. Towards sustainable sources for omega-3 fatty acids production. Curr. Opin. Biotechnol. 2014, 26, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Stramarkou, M.; Oikonomopoulou, V.; Chalima, A.; Boukouvalas, C.; Topakas, E.; Krokida, M. Optimization of green extractions for the recovery of docosahexaenoic acid (DHA) from Crypthecodinium cohnii. Algal Res. 2021, 58, 102374. [Google Scholar] [CrossRef]
- da Silva, T.L.; Moniz, P.; Silva, C.; Reis, A. The dark side of microalgae biotechnology: A heterotrophic biorefinery platform directed to ω-3 rich lipid production. Microorganisms 2019, 7, 670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taborda, T.; Moniz, P.; Reis, A.; da Silva, T.L. Evaluating low-cost substrates for Crypthecodinium cohnii lipids and DHA production, by flow cytometry. J. Appl. Phycol. 2021, 33, 263–274. [Google Scholar] [CrossRef]
- Ji, X.-J.; Ren, L.-J.; Huang, H. Omega-3 Biotechnology: A Green and Sustainable Process for Omega-3 Fatty Acids Production. Front. Bioeng. Biotechnol. 2015, 3, 190–209. [Google Scholar] [CrossRef] [Green Version]
- Sharma, J.; Sarmah, P.; Bishnoi, N.R. Market Perspective of EPA and DHA Production from Microalgae. In Nutraceutical Fatty Acids from Oleaginous Microalgae; Scrivener Publishing LLC: Beverly, MA, USA, 2020; pp. 281–297. [Google Scholar] [CrossRef]
- Braunwald, T.; French, W.T.; Claupein, W.; Graeff-Hönninger, S. Economic assessment of microbial biodiesel production using heterotrophic yeasts. Int. J. Green Energy 2016, 13, 274–282. [Google Scholar] [CrossRef]
- Jovanovic, S.; Dietrich, D.; Becker, J.; Kohlstedt, M.; Wittmann, C. Microbial production of polyunsaturated fatty acids—high-value ingredients for aquafeed, superfoods, and pharmaceuticals. Curr. Opin. Biotechnol. 2021, 69, 199–211. [Google Scholar] [CrossRef]
- Petrie, J.R.; Shrestha, P.; Zhou, X.R.; Mansour, M.P.; Liu, Q.; Belide, S.; Nichols, P.D.; Singh, S.P. Metabolic Engineering Plant Seeds with Fish Oil-Like Levels of DHA. PLoS ONE 2012, 7, 2–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos-Sánchez, N.F.; Valadez-Blanco, R.; Hernández-Carlos, B.; Torres-Ariño, A.; Guadarrama-Mendoza, P.C.; Salas-Coronado, R. Lipids rich in ω-3 polyunsaturated fatty acids from microalgae. Appl. Microbiol. Biotechnol. 2016, 100, 8667–8684. [Google Scholar] [CrossRef]
- Graham, I.A.; Larson, T.; Napier, J.A. Rational metabolic engineering of transgenic plants for biosynthesis of omega-3 polyunsaturates. Curr. Opin. Biotechnol. 2007, 18, 142–147. [Google Scholar] [CrossRef]
- Mendes, A.; Reis, A.; Vasconcelos, R.; Guerra, P.; Lopes Da Silva, T. Crypthecodinium cohnii with emphasis on DHA production: A review. J. Appl. Phycol. 2009, 21, 199–214. [Google Scholar] [CrossRef]
- Xiao, R.; Li, X.; Zheng, Y. Comprehensive Study of Cultivation Conditions and Methods on Lipid Accumulation of a Marine Protist, Thraustochytrium striatum. Protist 2018, 169, 451–465. [Google Scholar] [CrossRef]
- Lv, M.; Wang, F.; Zeng, L.; Bi, Y.; Cui, J.; Liu, L.; Bi, Y.; Chen, L.; Zhang, W. Identification and metabolomic analysis of a starch-deficient Crypthecodinium cohnii mutant reveals multiple mechanisms relevant to enhanced growth and lipid accumulation. Algal Res. 2020, 50, 102001. [Google Scholar] [CrossRef]
- Rumiani, L.A.; Jalili, H.; Amrane, A. Enhanced docosahexaenoic acid production by Crypthecodinium cohnii under combined stress in two-stage cultivation with date syrup based medium. Algal Res. 2018, 34, 75–81. [Google Scholar] [CrossRef]
- Paz, A.; Karnaouri, A.; Templis, C.C.; Papayannakos, N.; Topakas, E. Valorization of exhausted olive pomace for the production of omega-3 fatty acids by Crypthecodinium cohnii. Waste Manag. 2020, 118, 435–444. [Google Scholar] [CrossRef]
- De Swaaf, M.E.; Sijtsma, L.; Pronk, J.T. High-cell-density fed-batch cultivation of the docosahexaenoic acid producing marine alga Crypthecodinium cohnii. Biotechnol. Bioeng. 2003, 81, 666–672. [Google Scholar] [CrossRef]
- De Swaaf, M.E.; Pronk, J.T.; Sijtsma, L. Fed-batch cultivation of the docosahexaenoic-acid-producing marine alga Crypthecodinium cohnii on ethanol. Appl. Microbiol. Biotechnol. 2003, 61, 40–43. [Google Scholar] [CrossRef]
- de Swaaf, M.E.; de Rijk, T.C.; Eggink, G.; Sijtsma, L. Optimisation of docosahexaenoic acid production in batch cultivations by Crypthecodinium cohnii. Prog. Ind. Microbiol. 1999, 35, 185–192. [Google Scholar] [CrossRef]
- Karnaouri, A.; Asimakopoulou, G.; Kalogiannis, K.G.; Lappas, A.A.; Topakas, E. Efficient production of nutraceuticals and lactic acid from lignocellulosic biomass by combining organosolv fractionation with enzymatic/fermentative routes. Bioresour. Technol. 2021, 341, 125846. [Google Scholar] [CrossRef] [PubMed]
- Chalima, A.; Taxeidis, G.; Topakas, E. Optimization of the production of docosahexaenoic fatty acid by the heterotrophic microalga Crypthecodinium cohnii utilizing a dark fermentation effluent. Renew. Energy 2020, 152, 102–109. [Google Scholar] [CrossRef]
- Safdar, W.; Shamoon, M.; Zan, X.; Haider, J.; Sharif, H.R.; Shoaib, M.; Song, Y. Growth kinetics, fatty acid composition and metabolic activity changes of Crypthecodinium cohnii under different nitrogen source and concentration. AMB Express 2017, 7, 85. [Google Scholar] [CrossRef]
- Safdar, W.; Zan, X.; Shamoon, M.; Sharif, H.R.; Mukama, O.; Tang, X.; Song, Y. Effects of twenty standard amino acids on biochemical constituents, docosahexaenoic acid production and metabolic activity changes of Crypthecodinium cohnii. Bioresour. Technol. 2017, 238, 738–743. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Chen, F.; Liang, S.Z. Production potential of docosahexaenoic acid by the heterotrophic marine dinoflagellate Crypthecodinium cohnii. Process Biochem. 1999, 34, 633–637. [Google Scholar] [CrossRef]
- Gong, Y.; Liu, J.; Jiang, M.; Liang, Z.; Jin, H.; Hu, X.; Wan, X.; Hu, C. Improvement of omega-3 docosahexaenoic acid production by marine dinoflagellate Crypthecodinium cohnii using rapeseed meal hydrolysate and waste molasses as feedstock. PLoS ONE 2015, 10, e0125368. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Chen, F. Effects of medium glucose concentration and pH on docosahexaenoic acid content of heterotrophic Crypthecodinium cohnii. Process Biochem. 2000, 35, 1205–1209. [Google Scholar] [CrossRef]
- Diao, J.; Song, X.; Zhang, X.; Chen, L.; Zhang, W. Genetic Engineering of Crypthecodinium cohnii to increase growth and lipid accumulation. Front. Microbiol. 2018, 9, 492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grube, M.; Bekers, M.; Upite, D.; Kaminska, E. IR-spectroscopic studies of Zymomonas mobilis and levan precipitate. Vib. Spectrosc. 2002, 28, 277–285. [Google Scholar] [CrossRef]
- Fuchino, K.; Kalnenieks, U.; Rutkis, R.; Grube, M.; Bruheim, P. Metabolic profiling of glucose-fed metabolically active resting Zymomonas mobilis strains. Metabolites 2020, 10, 81. [Google Scholar] [CrossRef] [Green Version]
- Grube, M.; Kalnenieks, U.; Muter, O. Metabolic response of bacteria to elevated concentrations of glyphosate-based herbicide. Ecotoxicol. Environ. Saf. 2019, 173, 373–380. [Google Scholar] [CrossRef]
- Grube, M.; Shvirksts, K.; Krafft, C.; Kokorevicha, S.; Zandberga, E.; Abols, A.; Line, A.; Kalnenieks, U. Miniature diamond-anvil cells for FTIR-microspectroscopy of small quantities of biosamples. Analyst 2018, 143, 3595–3599. [Google Scholar] [CrossRef]
- Killeen, D.P.; Marshall, S.N.; Burgess, E.J.; Gordon, K.C.; Perry, N.B. Raman Spectroscopy of Fish Oil Capsules: Polyunsaturated Fatty Acid Quantitation Plus Detection of Ethyl Esters and Oxidation. J. Agric. Food Chem. 2017, 65, 3551–3558. [Google Scholar] [CrossRef]
- Karunathilaka, S.R.; Choi, S.H.; Mossoba, M.M.; Yakes, B.J.; Brückner, L.; Ellsworth, Z.; Srigley, C.T. Rapid classification and quantification of marine oil omega-3 supplements using ATR-FTIR, FT-NIR and chemometrics. J. Food Compos. Anal. 2019, 77, 9–19. [Google Scholar] [CrossRef]
- Guillén, M.D.; Cabo, N. Characterization of edible oils and lard by fourier transform infrared spectroscopy. Relationships between composition and frequency of concrete bands in the fingerprint region. JAOCS J. Am. Oil Chem. Soc. 1997, 74, 1281–1286. [Google Scholar] [CrossRef]
- Dean, A.P.; Sigee, D.C.; Estrada, B.; Pittman, J.K. Using FTIR spectroscopy for rapid determination of lipid accumulation in response to nitrogen limitation in freshwater microalgae. Bioresour. Technol. 2010, 101, 4499–4507. [Google Scholar] [CrossRef]
- Vongsvivut, J.; Heraud, P.; Zhang, W.; Kralovec, J.A.; McNaughton, D.; Barrow, C.J. Quantitative determination of fatty acid compositions in micro-encapsulated fish-oil supplements using Fourier transform infrared (FTIR) spectroscopy. Food Chem. 2012, 135, 603–609. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, R.; Lourenço, S.; Lopes, A.; Andrade, C.; Câmara, J.S.; Castilho, P.; Perestrelo, R. Evaluation of fatty acids profile as a useful tool towards valorization of by-products of agri-food industry. Foods 2021, 10, 2867. [Google Scholar] [CrossRef] [PubMed]
- Ripoche, A.; Guillard, A.S. Determination of fatty acid composition of pork fat by Fourier transform infrared spectroscopy. Meat Sci. 2001, 58, 299–304. [Google Scholar] [CrossRef]
- Meng, W.; Jiang, Y.; Rothschild, D.; Lipke, M.; Hall, G.; Wang, L. Modeling the structure and infrared spectra of omega-3 fatty acid esters. J. Chem. Phys. 2020, 153. [Google Scholar] [CrossRef] [PubMed]
- Ami, D.; Posteri, R.; Mereghetti, P.; Porro, D.; Doglia, S.M.; Branduardi, P. Fourier transform infrared spectroscopy as a method to study lipid accumulation in oleaginous yeasts. Biotechnol. Biofuels 2014, 7, 12. [Google Scholar] [CrossRef] [Green Version]
- Berzins, K.; Muiznieks, R.; Baumanis, M.R.; Strazdina, I.; Shvirksts, K.; Prikule, S.; Galvanauskas, V.; Pleissner, D.; Pentjuss, A.; Grube, M.; et al. Kinetic and stoichiometric modeling based analysis of docosahexaenoic acid (DHA) production potential by C.cohnii from glycerol, glucose and ethanol. Mar. Drugs 2022. Under review. [Google Scholar]
- Shapaval, V.; Afseth, N.K.; Vogt, G.; Kohler, A. Fourier transform infrared spectroscopy for the prediction of fatty acid profiles in Mucor fungi grown in media with different carbon sources. Microb. Cell Fact. 2014, 13, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Vongsvivut, J.; Heraud, P.; Gupta, A.; Puri, M.; McNaughton, D.; Barrow, C.J. FTIR microspectroscopy for rapid screening and monitoring of polyunsaturated fatty acid production in commercially valuable marine yeasts and protists. Analyst 2013, 138, 6016–6031. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, S.; Yoshida, H. Noninvasive analyses of polyunsaturated fatty acids in human oral mucosa in vivo by fourier-transform infrared spectroscopy. Biopolymers 2004, 74, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.-T.; Shimamura, T.; Ishida, N.; Takahashi, H. Investigation of utilization of the algal biomass residue after oil extraction to lower the total production cost of biodiesel. J. Biosci. Bioeng. 2012, 114, 330–333. [Google Scholar] [CrossRef] [PubMed]
- Halim, R.; Danquah, M.K.; Webley, P.A. Extraction of oil from microalgae for biodiesel production: A review. Biotechnol. Adv. 2012, 30, 709–732. [Google Scholar] [CrossRef] [PubMed]
- Mendes, A.; Da Silva, T.L.; Reis, A. DHA concentration and purification from the marine heterotrophic microalga Crypthecodinium cohnii CCMP 316 by winterization and urea complexation. Food Technol. Biotechnol. 2007, 45, 38–44. [Google Scholar]
- Dubencovs, K.; Liepins, J.; Suleiko, A.; Suleiko, A.; Vangravs, R.; Kassaliete, J.; Scerbaka, R.; Grigs, O. Optimization of synthetic media composition for Kluyveromyces marxianus fed-batch cultivation. Fermentation 2021, 7, 62. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Carbon Source | Nitrogen Source | Fermentation Mode | µmax, h−1 | Biomass, g·L−1 | Yx/s, g·g−1 | DHA, g·L−1 | Lipid,% of DCW | Ref. |
|---|---|---|---|---|---|---|---|---|
| Acetate | Ammonium sulphate * | Batch | - | ~7.7 | - | - | - | [21] |
| Yeast extract * | - | ~6.0 | - | - | 18.67 ** | |||
| 0.025 | 7.03 | - | 0.03118 ± 0.00160 | 12.43 ± 0.62 | [4] | |||
| Acetic acid | Ammonium sulphate * | Fed-batch | - | - | - | 0.1016 | - | [21] |
| Yeast extract * | - | - | - | 0.1629 | - | |||
| 0.053 | 109 | 0.13 | 19 | 55.69 ** | [17] | |||
| Ethanol | Yeast extract | Fed-batch | 0.05 | 83 | 0.31 | 11.7 | 42.17 ** | [18] |
| Galacturonic acid | Yeast extract | Batch | - | 3.07 ± 0.04 | - | 0.05273 ± 0.00015 | 46.58 ** | [16] |
| Glucose | Sodium nitrate * | Batch | - | 23.7 ± 0.61 | 0.38 | 0.99 | 18.14 ** | [22] |
| Threonine * | - | 25.3 | 0.95 ± 0.09 | - | 28.46 ** | [23] | ||
| Yeast extract, tryptone | 0.067 | 2.046 | 0.499 | 0.159 | - | [24] | ||
| Yeast extract | 0.017 | 2.66 | - | 0.01634 ± 0.00168 | 14.70 ± 0.07 | [4] | ||
| - | 6.4 | - | 1.4 | - | [20] | |||
| - | 27.7 | - | 1.6 | 13.36 | [19] | |||
| Glycerol (crude) | Yeast extract | Batch | 0.018 | 5.05 | - | 0.02696 ± 0.00107 | 14.70 ± 0.73 | [4] |
| Glycerol (pure) | Yeast extract | Batch | 0.019 | 6.33 | - | 0.01307 ± 0.00072 | 11.04 ± 0.50 | |
| Molasses | Yeast extract | Batch | 0.013 | 3.91 | - | 0.01956 ± 0.00100 | 11.12 ± 0.56 | |
| Molasses (crude waste) | Rapeseed meal hydrolysate | Batch | - | 3.43 | - | 0.00872 | - | [25] |
| Organosolv pulps | Yeast extract | Batch | - | 5.2 | - | 0.8 | - | [20] |
| Medium | Component, g·L−1 | µmax, h−1 | Yx/s, g·g−1 | ||||
|---|---|---|---|---|---|---|---|
| Sea Salts | Glucose | YE | DEA | DEB | |||
| YE | 12.5 | 10.0 | 1.0 | - | - | 0.930 | 0.446 |
| DEA | 12.5 | 10.0 | - | 1.0 | - | 1.012 | 0.601 |
| DEB | 12.5 | 10.0 | - | - | 1.0 | 0.655 | 0.398 |
| DEA75 | 12.5 | 10.0 | 0.25 | 0.75 | - | 0.900 | 0.403 |
| DEB75 | 12.5 | 10.0 | 0.25 | - | 0.75 | 0.901 | 0.371 |
| Glucose | 12.5 | 10.0 | - | - | - | 0.615 | 0.397 |
| Medium | Component, g·L−1 | µmax, h−1 | Yx/s, g·g−1 | |||||
|---|---|---|---|---|---|---|---|---|
| Sea Salts | EE | Ethanol | YE | DEA | DEB | |||
| EE_YE | 12.5 | 5.9 | - | 1.0 | - | - | 0.757 | 0.282 |
| EE_DEA | 12.5 | 5.9 | - | - | 1.0 | - | 0.701 | 0.234 |
| EE_DEB | 12.5 | 5.9 | - | - | - | 1.0 | 0.651 | 0.221 |
| EE_DEA75 | 12.5 | 5.9 | - | 0.25 | 0.75 | - | 0.658 | 0.238 |
| EE_DEB75 | 12.5 | 5.9 | - | 0.25 | - | 0.75 | 0.606 | 0.218 |
| EE | 12.5 | 5.9 | - | - | - | - | 0.470 | 0.124 |
| Ethanol | 12.5 | - | 4.7 | - | - | - | 0.383 | 0.122 |
| Medium | Component, g·L−1 | ||||||
|---|---|---|---|---|---|---|---|
| Sea Salts | Glucose | EE | Ethanol | YE | DEA | DEB | |
| YE | 12.5 | 10.0 | - | - | 1.0 | - | - |
| DEA | 12.5 | 10.0 | - | - | - | 1.0 | - |
| DEB | 12.5 | 10.0 | - | - | - | - | 1.0 |
| DEA75 | 12.5 | 10.0 | - | - | 0.25 | 0.75 | - |
| DEB75 | 12.5 | 10.0 | - | - | 0.25 | - | 0.75 |
| Glucose | 12.5 | 10.0 | - | - | - | - | - |
| EE_YE | 12.5 | - | 5.9 | - | 1.0 | - | - |
| EE_DEA | 12.5 | - | 5.9 | - | - | 1.0 | - |
| EE_DEB | 12.5 | - | 5.9 | - | - | - | 1.0 |
| EE_DEA75 | 12.5 | - | 5.9 | - | 0.25 | 0.75 | - |
| EE_DEB75 | 12.5 | - | 5.9 | - | 0.25 | - | 0.75 |
| EE | 12.5 | - | 5.9 | - | - | - | - |
| Ethanol | 12.5 | - | - | 4.7 | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Didrihsone, E.; Dubencovs, K.; Grube, M.; Shvirksts, K.; Suleiko, A.; Suleiko, A.; Vanags, J. Crypthecodinium cohnii Growth and Omega Fatty Acid Production in Mediums Supplemented with Extract from Recycled Biomass. Mar. Drugs 2022, 20, 68. https://doi.org/10.3390/md20010068
Didrihsone E, Dubencovs K, Grube M, Shvirksts K, Suleiko A, Suleiko A, Vanags J. Crypthecodinium cohnii Growth and Omega Fatty Acid Production in Mediums Supplemented with Extract from Recycled Biomass. Marine Drugs. 2022; 20(1):68. https://doi.org/10.3390/md20010068
Chicago/Turabian StyleDidrihsone, Elina, Konstantins Dubencovs, Mara Grube, Karlis Shvirksts, Anastasija Suleiko, Arturs Suleiko, and Juris Vanags. 2022. "Crypthecodinium cohnii Growth and Omega Fatty Acid Production in Mediums Supplemented with Extract from Recycled Biomass" Marine Drugs 20, no. 1: 68. https://doi.org/10.3390/md20010068
APA StyleDidrihsone, E., Dubencovs, K., Grube, M., Shvirksts, K., Suleiko, A., Suleiko, A., & Vanags, J. (2022). Crypthecodinium cohnii Growth and Omega Fatty Acid Production in Mediums Supplemented with Extract from Recycled Biomass. Marine Drugs, 20(1), 68. https://doi.org/10.3390/md20010068