
Intrabody Tetrodotoxin Distribution and Possible Hypothesis for Its Migration in Ribbon Worms Cephalothrix cf. simula (Palaeonemertea, Nemertea)


Abstract
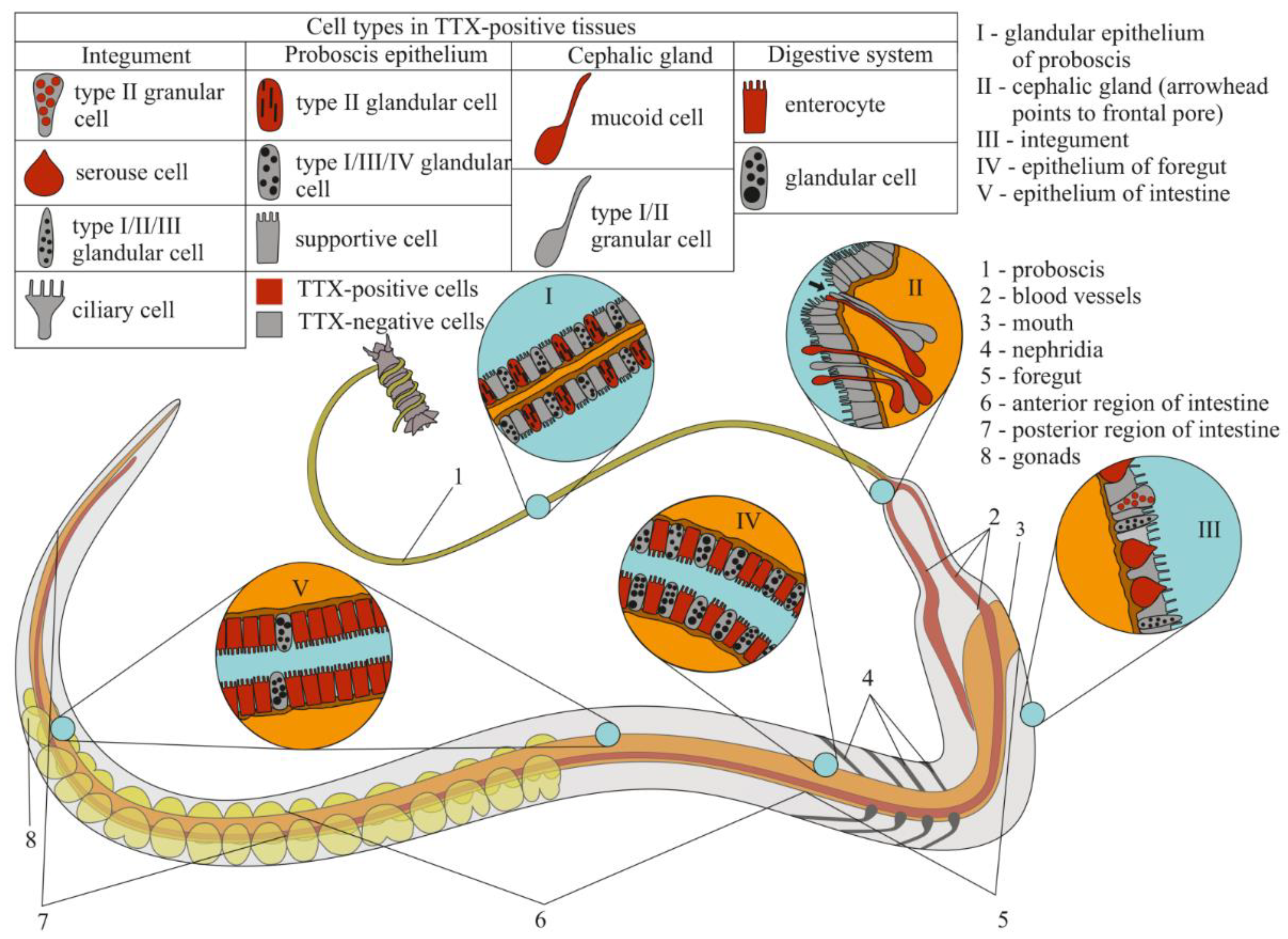
:1. Introduction
2. Results
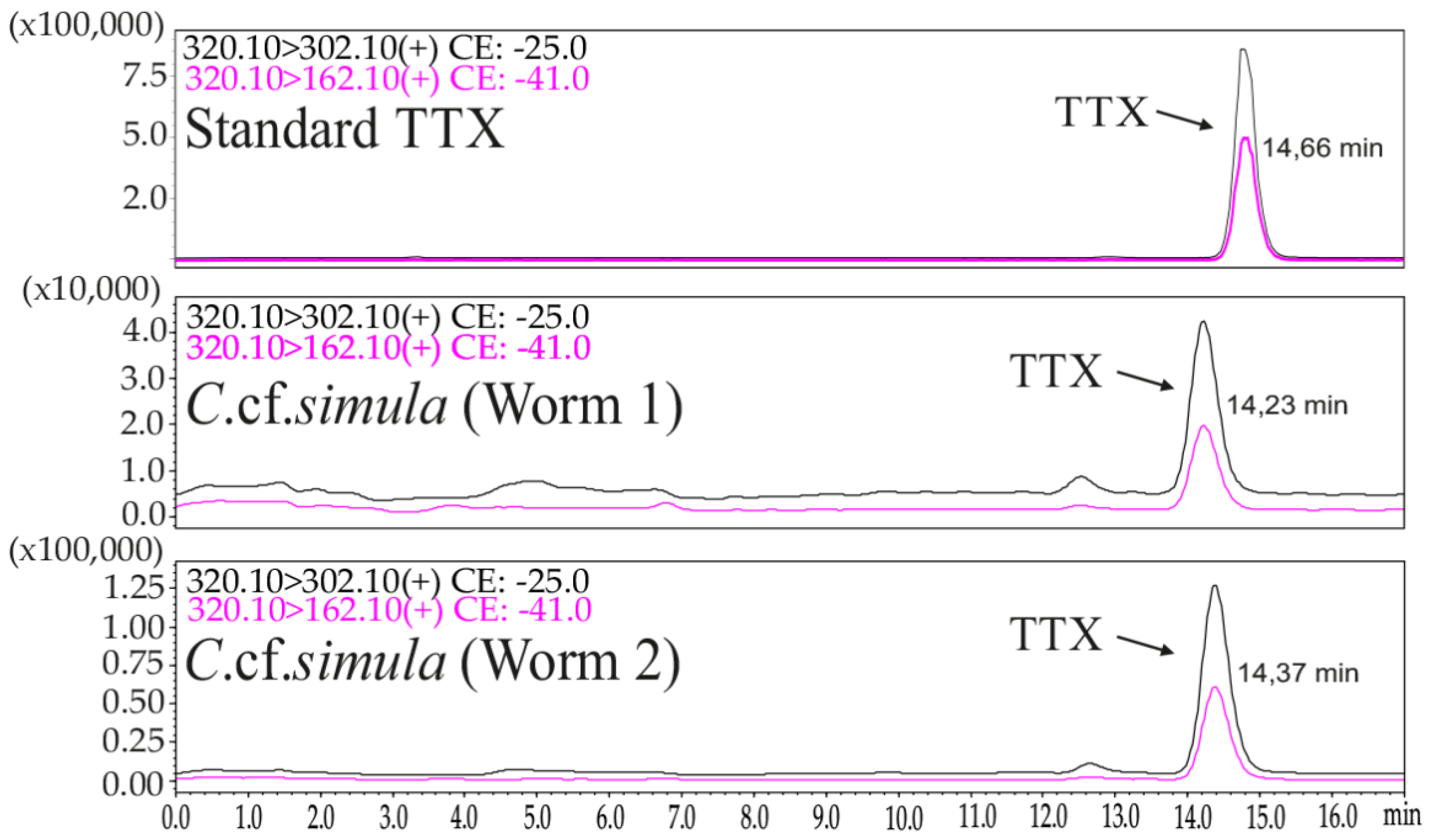
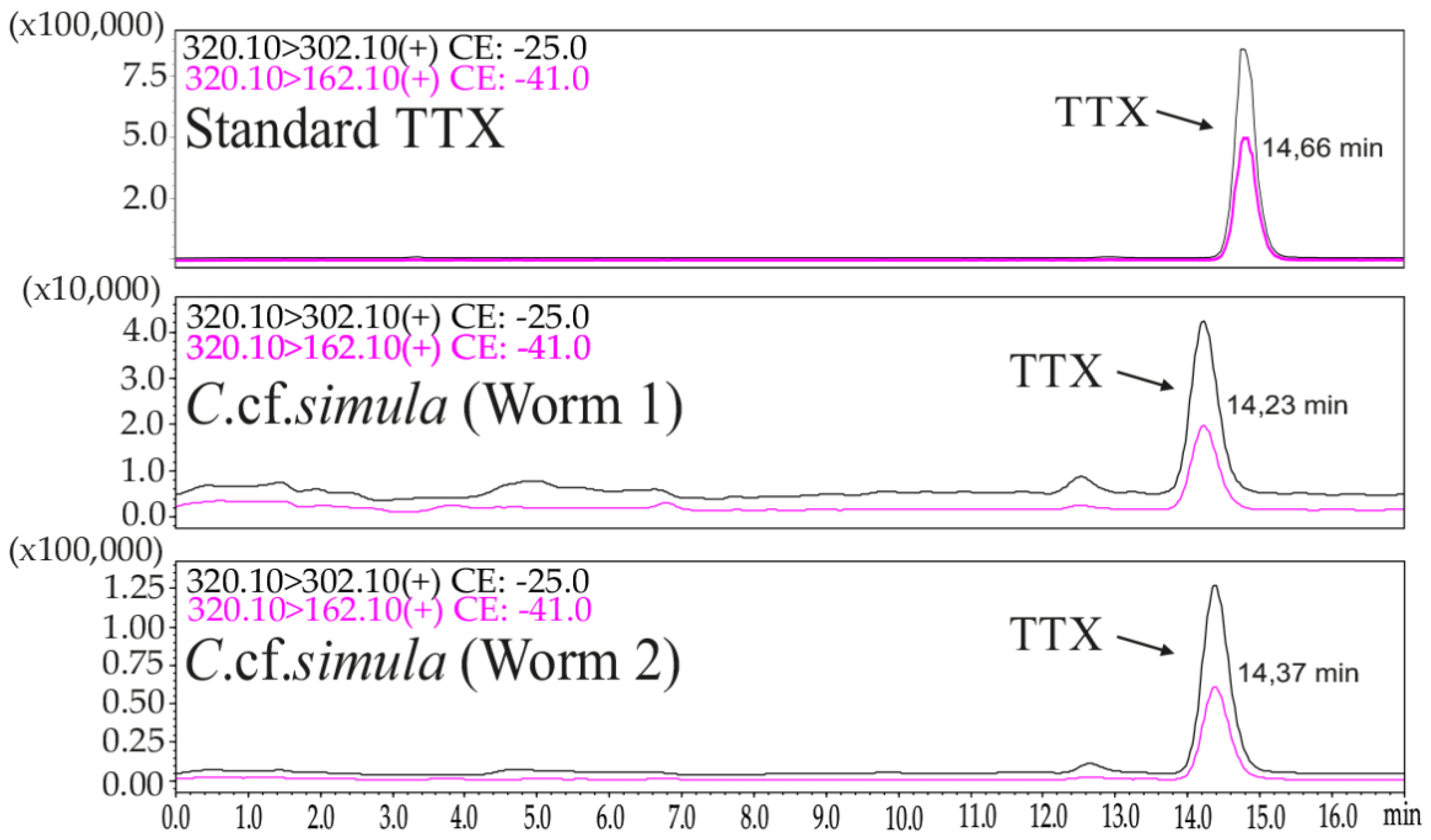
2.1. TTX Measurement by LC–MS/MS
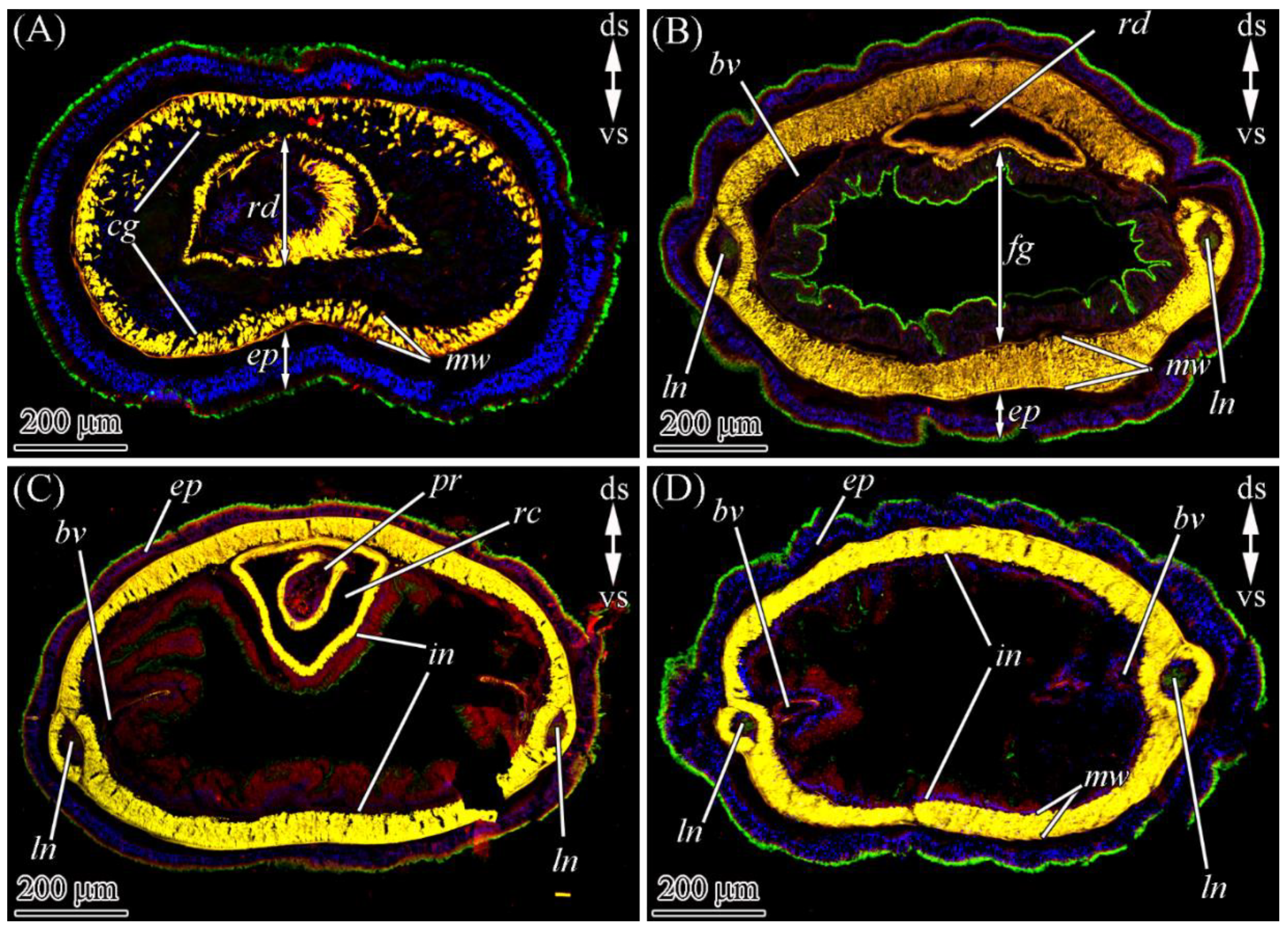
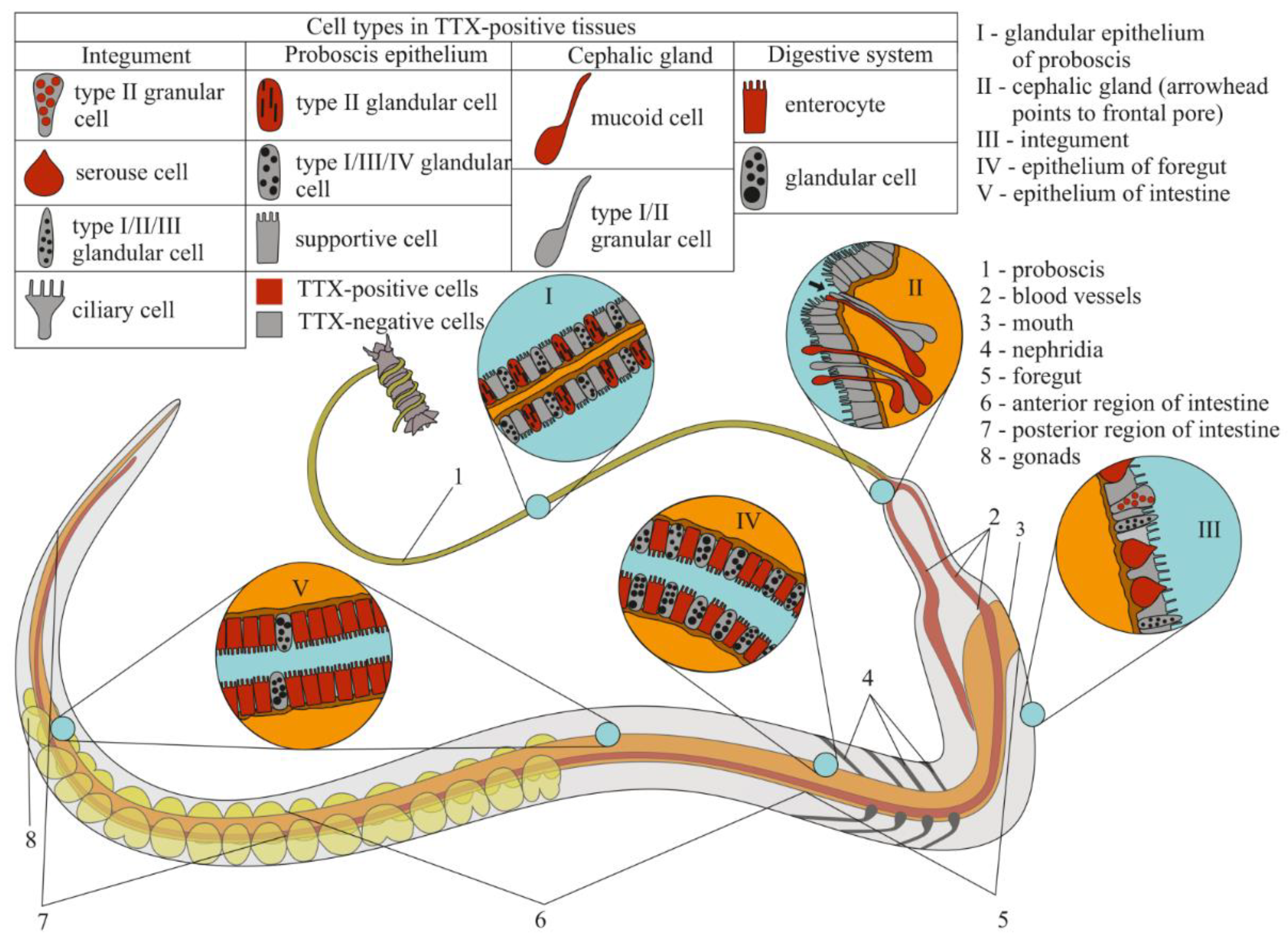
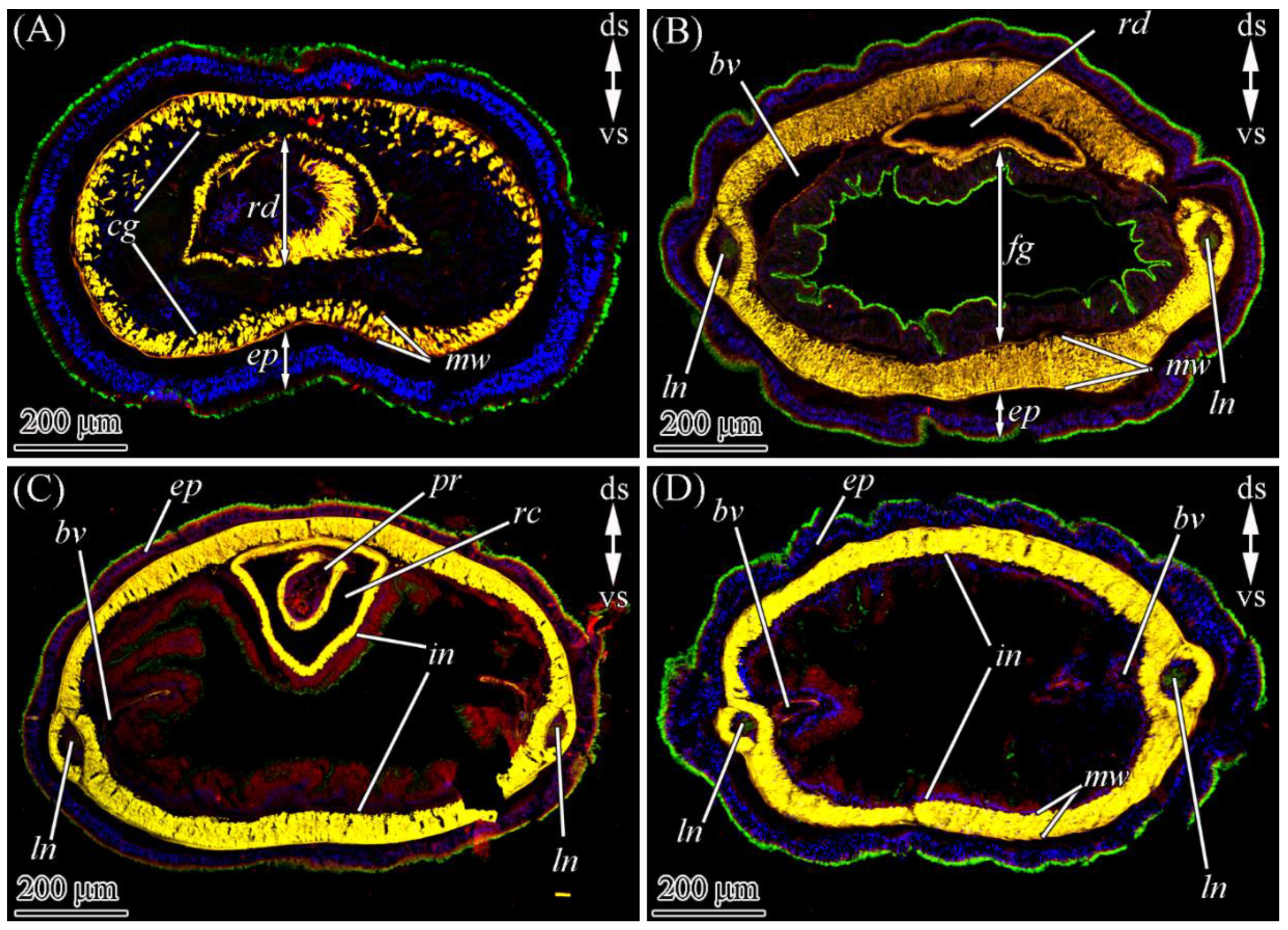
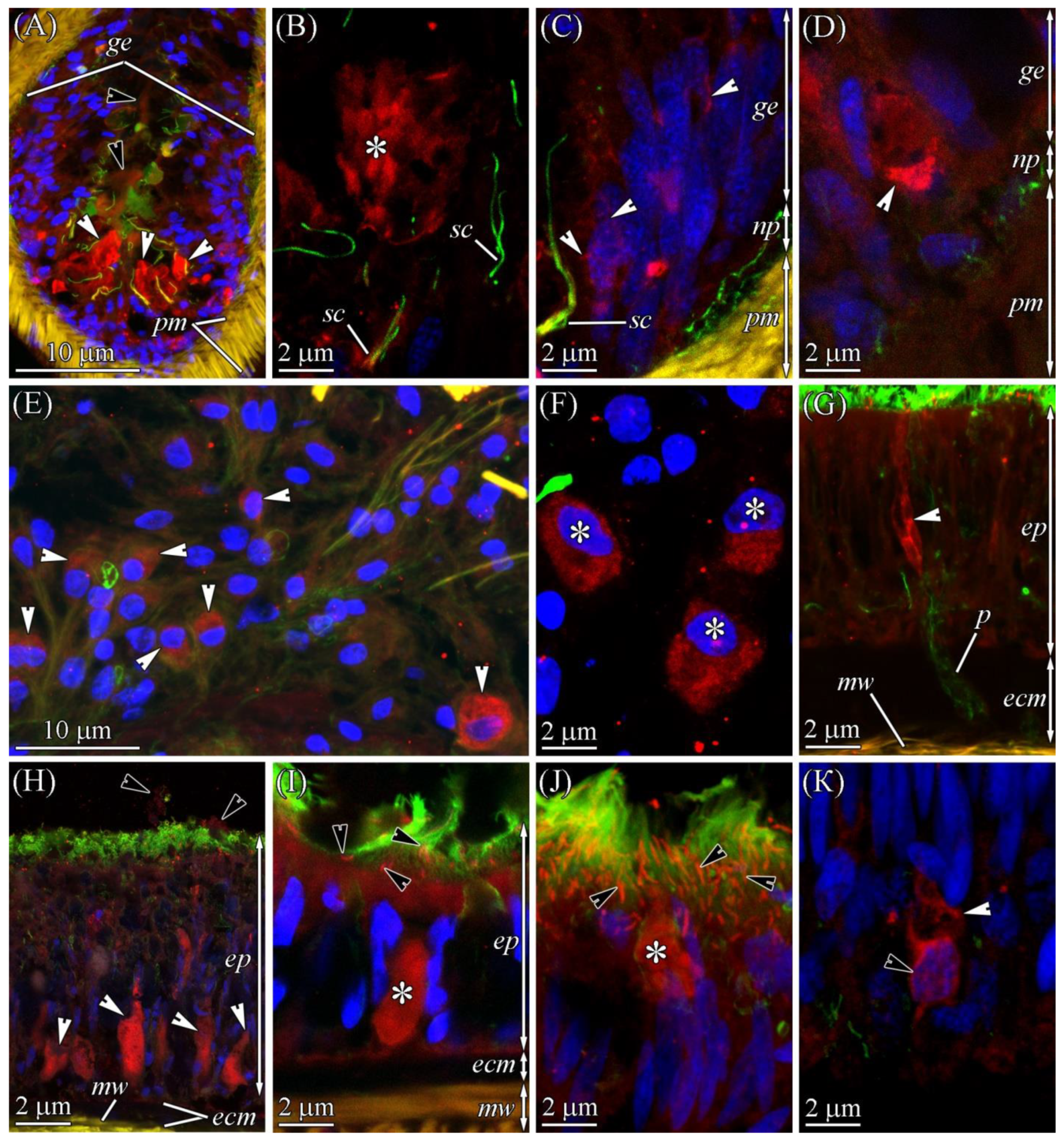
2.2. TTX Distribution by CLSM
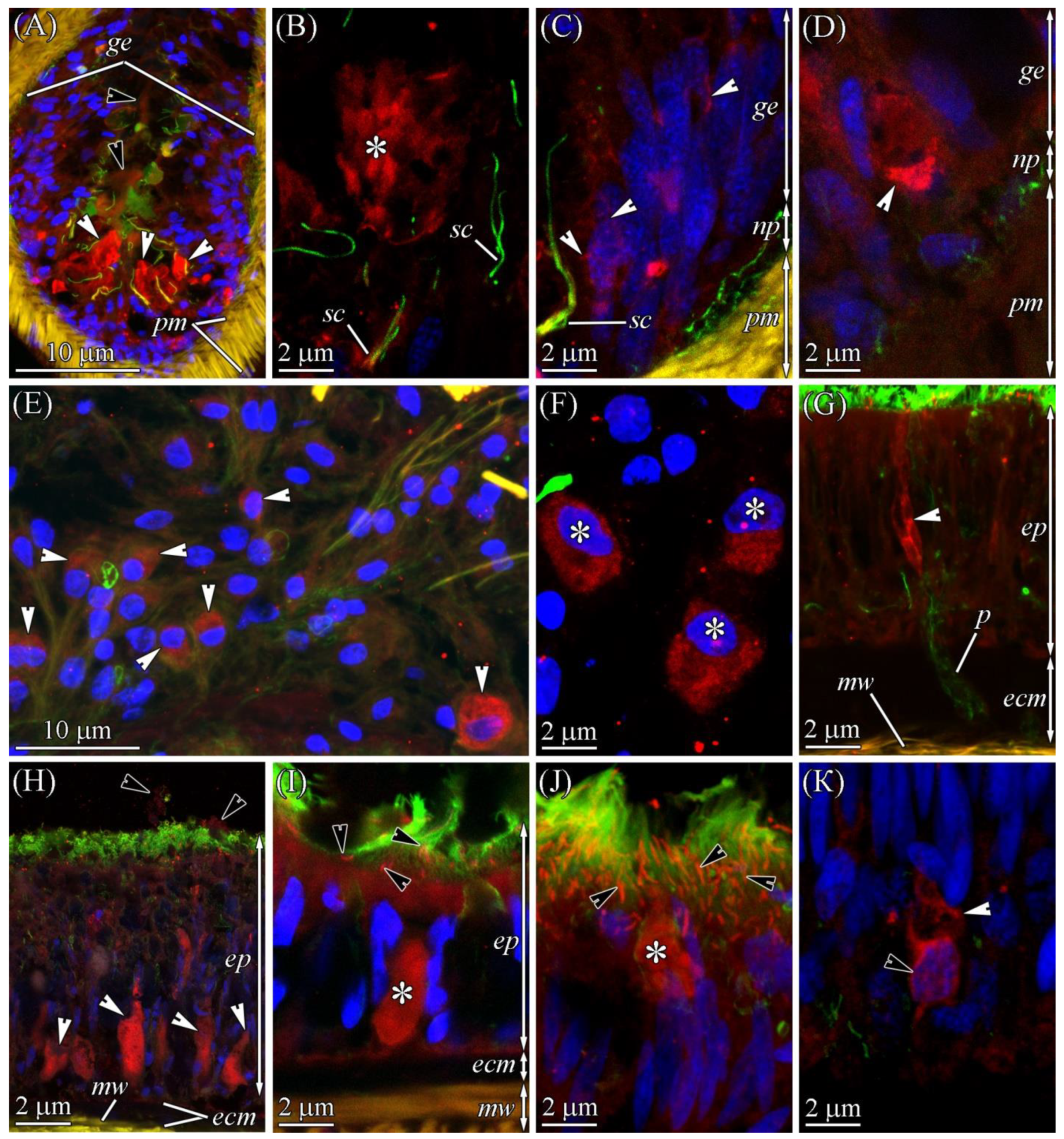
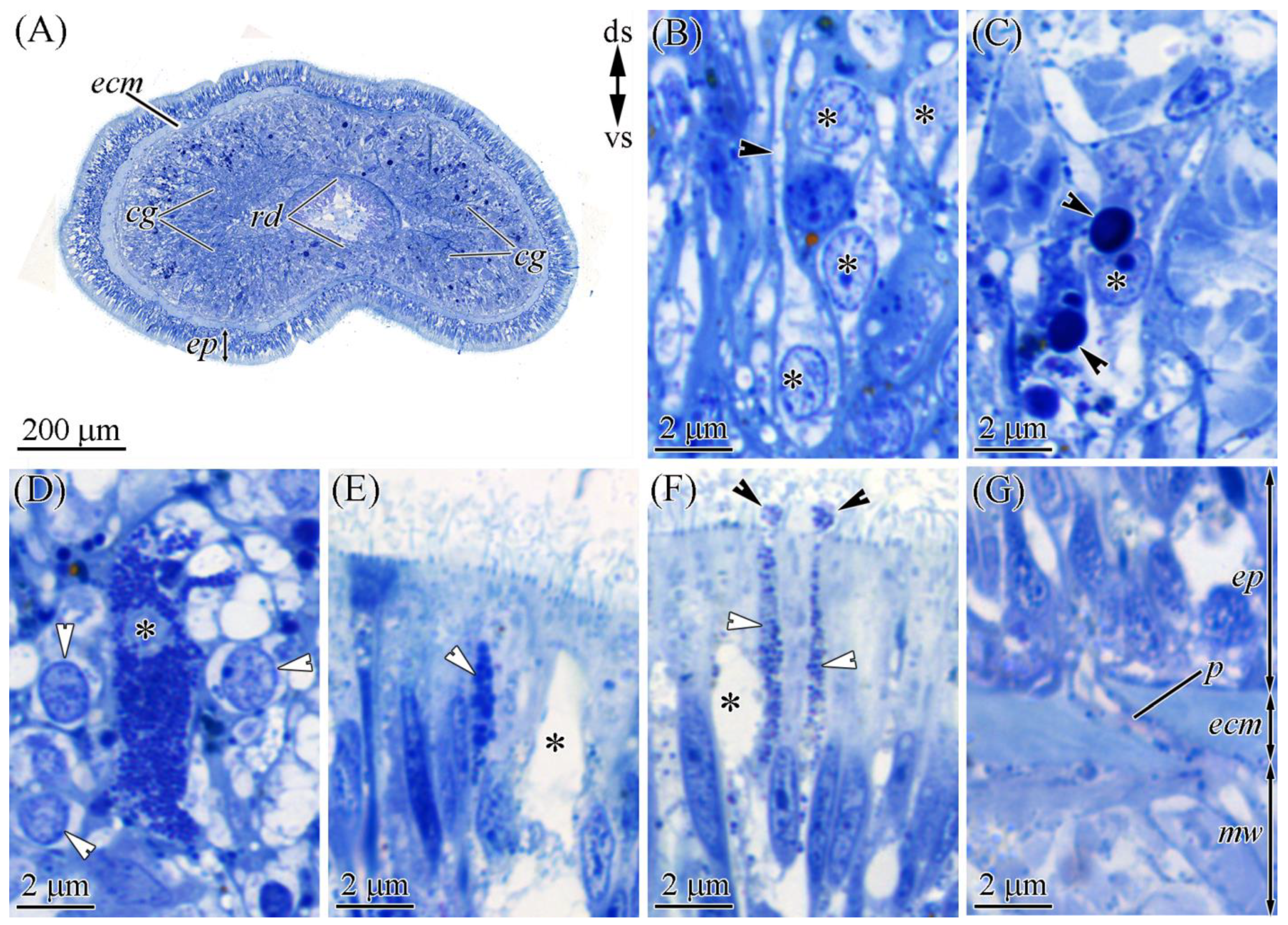
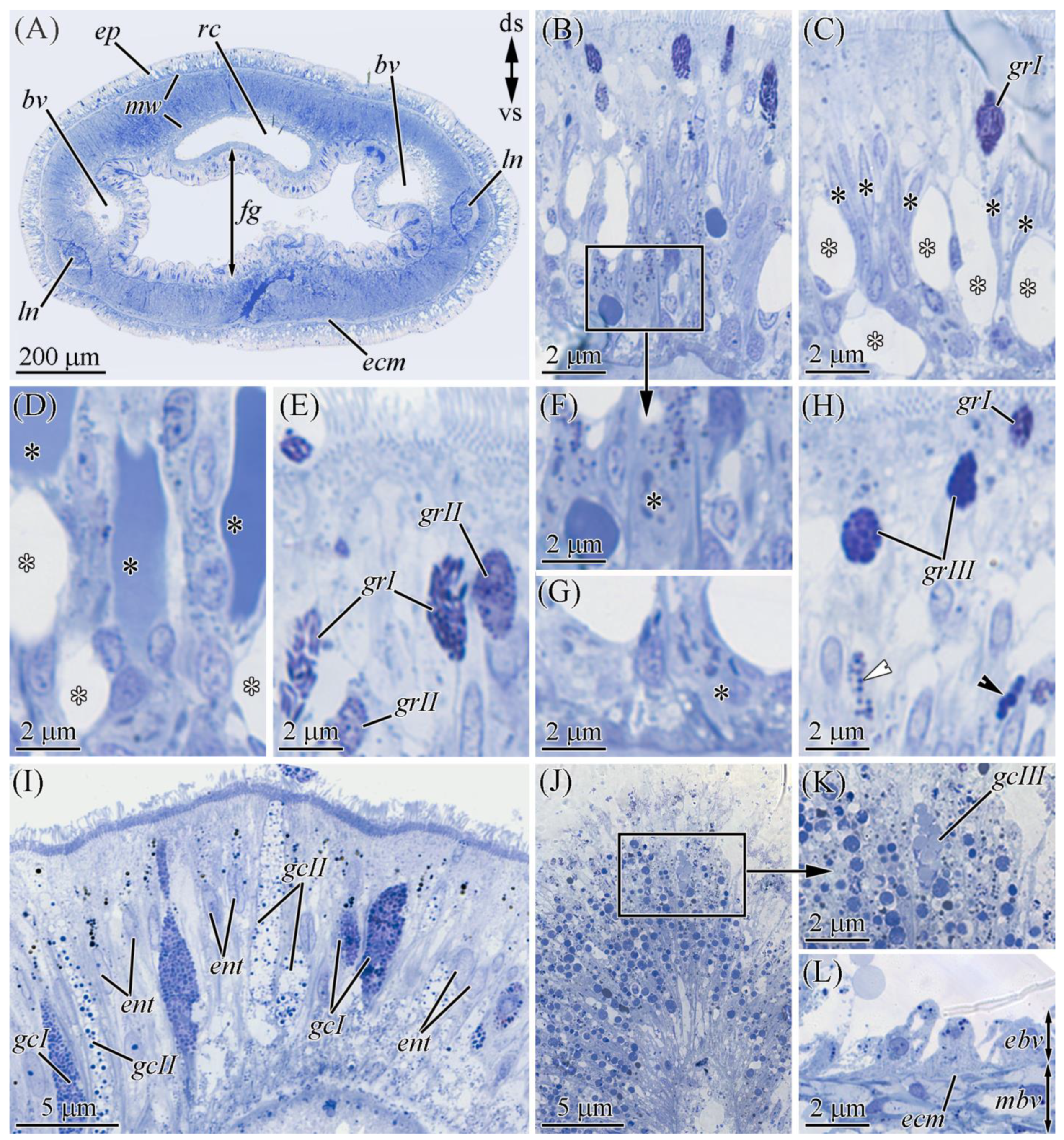
2.2.1. Glandular Epithelium of the Proboscis
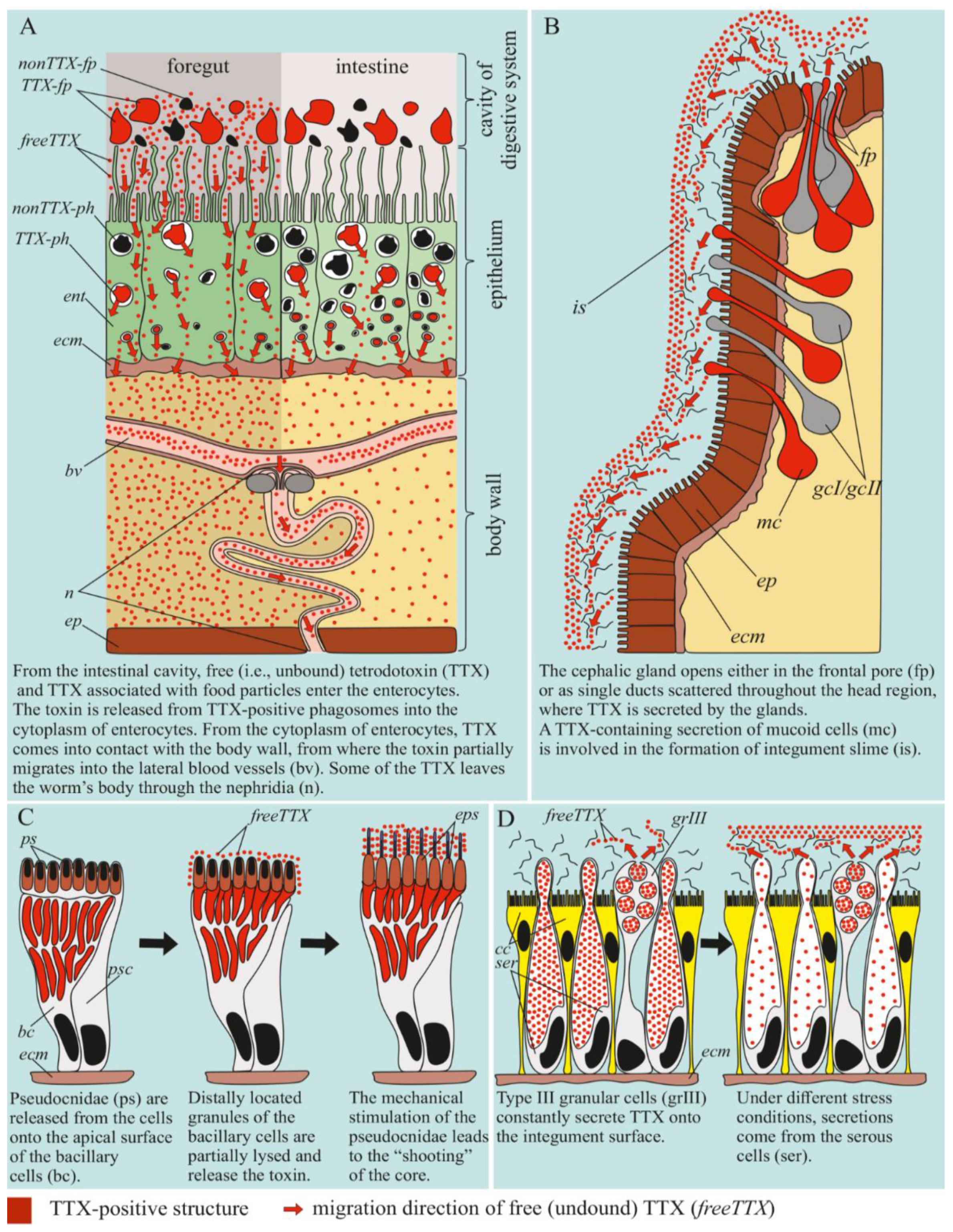
2.2.2. Cephalic Gland
2.2.3. Integument
2.2.4. Digestive System
2.2.5. Blood Vascular and Excretory Systems
2.2.6. Nervous System
2.2.7. Reproductive System
2.2.8. Muscles
3. Discussion
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Strand, M.; Norenburg, J.; Alfaya, J.E.; Ángel Fernández-Álvarez, F.; Andersson, H.S.; Andrade, S.C.S.; Bartolomaeus, T.; Beckers, P.; Bigatti, G.; Cherneva, I.; et al. Nemertean taxonomy—Implementing changes in the higher ranks, dismissing Anopla and Enopla. Zool. Scr. 2019, 48, 118–119. [Google Scholar] [CrossRef] [Green Version]
- Mcdermott, J.J. Observations on the food and feeding behavior of estuarine nemertean worms belonging to the order hoplonemertea. Biol. Bull. 1976, 150, 57–68. [Google Scholar] [CrossRef]
- Mcdermott, J.J.; Roe, P. Food, Feeding Behavior and Feeding Ecology of Nemerteans. Am. Zool. 1985, 25, 113–125. [Google Scholar] [CrossRef]
- Chernyshev, A.V. Nemertean biodiversity in the Sea of Japan and adjacent areas. In Marine Biodiversity and Ecosystem Dynamics of the Northwest Pacific Ocean; Song, S., Adrianov, A.V., Lutaenko, K.A., Xiao-Xia, S., Eds.; Publishing House of Science: Beijing, China, 2014; pp. 119–135. [Google Scholar]
- Kem, W.R. Structure and action of nemertine toxins. Integr. Comp. Biol. 1985, 25, 99–111. [Google Scholar] [CrossRef]
- Göransson, U.; Jacobsson, E.; Strand, M.; Andersson, H.S. The toxins of nemertean worms. Toxins 2019, 11, 120. [Google Scholar] [CrossRef] [Green Version]
- Miyazawa, K.; Higashiyama, M.; Ito, K.; Noguchi, T.; Arakawa, O.; Shida, Y.; Hashimoto, K. Tetrodotoxin in two species of ribbon worm (Nemertini), Lineus fuscoviridis and Tubulanus punctatus. Toxicon 1988, 26, 867–874. [Google Scholar] [CrossRef]
- Asakawa, M.; Ito, K.; Kajihara, H. Highly toxic ribbon worm Cephalothrix simula containing tetrodotoxin in Hiroshima Bay, Hiroshima Prefecture, Japan. Toxins 2013, 5, 376–395. [Google Scholar] [CrossRef]
- Magarlamov, T.Y.; Shokur, O.A.; Chernyshev, A.V. Distribution of tetrodotoxin in the ribbon worm Lineus alborostratus. Toxicon 2016, 112, 29–34. [Google Scholar] [CrossRef]
- Vlasenko, A.E.; Velansky, P.V.; Chernyshev, A.V.; Kuznetsov, V.G.; Magarlamov, T.Y. Tetrodotoxin and its analogues profile in nemertean species from the sea of Japan. Toxicon 2018, 156, 48–51. [Google Scholar] [CrossRef]
- Turner, A.D.; Fenwick, D.; Powell, A.; Dhanji-Rapkova, M.; Ford, C.; Hatfield, R.G.; Santos, A.; Martinez-Urtaza, J.; Bean, T.P.; Baker-Austin, C.; et al. New invasive nemertean species (Cephalothrix simula) in england with high levels of tetrodotoxin and a microbiome linked to toxin metabolism. Mar. Drugs 2018, 16, 452. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.E.; Arakawa, O.; Noguchi, T.; Miyazawa, K.; Shida, Y.; Hashimoto, K. Tetrodotoxin and related substances in a ribbon worm Cephalothrix linearis (Nemertean). Toxicon 1990, 28, 1083–1093. [Google Scholar] [CrossRef]
- Asakawa, M.; Toyoshima, T.; Shida, Y.; Noguchi, T.; Miyazawa, K. Paralytic toxins in a ribbon worm Cephalothrix species (Nemertean) adherent to cultured oysters in Hiroshima Bay, Hiroshima Prefecture, Japan. Toxicon 2000, 38, 763–773. [Google Scholar] [CrossRef]
- Asakawa, M.; Toyoshima, T.; Ito, K.; Bessho, K.; Yamaguchi, C.; Tsunetsugu, S.; Shida, Y.; Kajihara, H.; Mawatari, S.F.; Noguchi, T.; et al. Paralytic toxicity in the ribbon worm Cephalothrix species (Nemertea) in Hiroshima Bay, Hiroshima Prefecture, Japan and the isolation of tetrodotoxin as a main component of its toxins. Toxicon 2003, 41, 747–753. [Google Scholar] [CrossRef]
- Vlasenko, A.E.; Magarlamov, T.Y. Tetrodotoxin and its analogues in Cephalothrix cf. simula (Nemertea: Palaeonemertea) from the Sea of Japan (Peter the Great Gulf): Intrabody distribution and secretions. Toxins 2020, 12, 745. [Google Scholar] [CrossRef] [PubMed]
- Tanu, M.B.; Mahmud, Y.; Arakawa, O.; Takatani, T.; Kajihara, H.; Kawatsu, K.; Hamano, Y.; Asakawa, M.; Miyazawa, K.; Noguchi, T. Immunoenzymatic visualization of tetrodotoxin (TTX) in Cephalothrix species (Nemertea: Anopla: Palaeonemertea: Cephalotrichidae) and Planocera reticulata (Platyhelminthes: Turbellaria: Polycladida: Planoceridae). Toxicon 2004, 44, 515–520. [Google Scholar] [CrossRef]
- Junoy, J.; Montalvo, S.; Roldán, C.; García-Corrales, P. Ultrastructural study of the bacillary, granular and mucoid proboscidial gland cells of Riseriellus occultus (Nemertini, Heteronemertini). Acta Zool. 2000, 81, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Magarlamov, T.Y.; Turbeville, J.M.; Chernyshev, A.V. Pseudocnidae of ribbon worms (nemertea): Ultrastructure, maturation, and functional morphology. PeerJ 2021, 9, e10912. [Google Scholar] [CrossRef] [PubMed]
- Ferraris, J.D. Histological study of cephalic glands in selected nemertina. Trans. Am. Microsc. Soc. 1979, 93, 437–446. [Google Scholar] [CrossRef]
- Norenburg, J.L. Structure of the nemertine integument with consideration of its ecological and phylogenetic significance. Integr. Comp. Biol. 1985, 25, 37–51. [Google Scholar] [CrossRef] [Green Version]
- Bartolomaeus, T.; von Döhren, J. Comparative morphology and evolution of the nephridia in Nemertea. J. Nat. Hist. 2010, 44, 2255–2286. [Google Scholar] [CrossRef]
- Gibson, R. Nemerteans; Hutchinson: London, UK, 1972. [Google Scholar]
- Jennings, J.B.; Gibson, R. Observations on the nutrition of seven species of rhynchocoelan worms. Biol. Bull. 1969, 136, 405–433. [Google Scholar] [CrossRef]
- Noguchi, T.; Arakawa, O. Tetrodotoxin—Distribution and accumulation in aquatic organisms, and cases of human intoxication. Mar. Drugs 2008, 6, 220–242. [Google Scholar] [CrossRef] [Green Version]
- Williams, B.L.; Caldwell, R.L. Intra-organismal distribution of tetrodotoxin in two species of blue-ringed octopuses (Hapalochlaena fasciata and H. lunulata). Toxicon 2009, 54, 345–353. [Google Scholar] [CrossRef]
- Salvitti, L.R.; Wood, S.A.; Winsor, L.; Cary, S.C. Intracellular immunohistochemical detection of tetrodotoxin in Pleurobranchaea maculata (Gastropoda) and Stylochoplana sp. (Turbellaria). Mar. Drugs 2015, 13, 756–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, T.; Nagashima, Y.; Kusuhara, H.; Ishizaki, S.; Shimakura, K.; Shiomi, K. Pharmacokinetics of tetrodotoxin in puffer fish Takifugu rubripes by a single administration technique. Toxicon 2008, 51, 1051–1059. [Google Scholar] [CrossRef]
- Tatsuno, R.; Shikina, M.; Soyano, K.; Ikeda, K.; Takatani, T.; Arakawa, O. Maturation-associated changes in the internal distribution of tetrodotoxin in the female goby Yongeichthys criniger. Toxicon 2013, 63, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Kajihara, H. Rhynchocoel vessel in Cephalotrichidae (Nemertea: Palaeonemertea). J. Nat. Hist. 2010, 44, 2321–2329. [Google Scholar] [CrossRef]
- Ramírez-Carreto, S.; Vera-Estrella, R.; Portillo-Bobadilla, T.; Licea-Navarro, A.; Bernaldez-Sarabia, J.; Rudiño-Piñera, E.; Verleyen, J.J.; Rodríguez, E.; Rodríguez-Almazán, C. Transcriptomic and proteomic analysis of the tentacles and mucus of Anthopleura dowii Verrill, 1869. Mar. Drugs 2019, 17, 436. [Google Scholar] [CrossRef] [Green Version]
- Reverter, M.; Tapissier-Bontemps, N.; Lecchini, D.; Banaigs, B.; Sasal, P. Biological and ecological roles of external fish mucus: A review. Fishes 2018, 3, 41. [Google Scholar] [CrossRef] [Green Version]
- Nagashima, Y.; Arakawa, O. Pufferfish poisoning and tetrodotoxin. In Marine and Freshwater Toxins; Gopalakrishnakone, P., Haddad, V., Jr., Kem, W.R., Tubaro, A., Kim, E., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 1–21. ISBN 9789400766501. [Google Scholar]
- Whitelaw, B.L.; Strugnell, J.M.; Faou, P.; Da Fonseca, R.R.; Hall, N.E.; Norman, M.; Finn, J.; Cooke, I.R. Combined transcriptomic and proteomic analysis of the posterior salivary gland from the southern blue-ringed octopus and the southern sand octopus. J. Proteome Res. 2016, 15, 3284–3297. [Google Scholar] [CrossRef]
- Chernyshev, A.V. Comparative Morphology, Systematics and Phylogeny of the Nemerteans; Dalnauka: Vladivostok, Russia, 2011. [Google Scholar]
- Boie, H.-J. Die Paketdrüsenzone von Lineus rüber O. F. Müller (Nemertini). Z. Morphol. Okol. Tiere 1952, 41, 188–222. [Google Scholar] [CrossRef]
- Oaks, J.A. Ultrastructure of Lineus ruber (rhyncocoela) epidermis. Tissue Cell 1978, 10, 227–242. [Google Scholar] [CrossRef]
- Stricker, S.A.; Cloney, R.A. The ultarstructure of venom-producing cells in Paranemertes peregrina (Nemertea, Hoplonemertea). J. Morphol. 1983, 177, 89–107. [Google Scholar] [CrossRef] [PubMed]
- von Reumont, B.M.; Lüddecke, T.; Timm, T.; Lochnit, G.; Vilcinskas, A.; von Döhren, J.; Nilsson, M.A. Proteo-transcriptomic analysis identifies potential novel toxins secreted by the predatory, orey-piercing ribbon worm Amphiporus lactifloreus. Mar. Drugs 2020, 18, 407. [Google Scholar] [CrossRef] [PubMed]
- Montalvo, S.; Roldán, C.; Junoy, J.; García-Corrales, P. Ultrastructural study of two glandular systems in the proboscidial glandular epithelium of Riseriellus occultus (Nemertea, Heteronemertea). Zoomorphology 1998, 117, 247–257. [Google Scholar] [CrossRef]
- Turbeville, J.M. Ultrastructure of the pseudocnidae of the palaeonemerteans Cephalothrix cf. rufifrons and Carinomella lactea and an assessment of their phylogenetic utility. J. Nat. Hist. 2006, 40, 967–979. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M.; Sugimoto, A.; Terakawa, T.; Shoji, Y.; Miyazawa, T.; Yasumoto, T. Purification, characterization, and cDNA cloning of a novel soluble saxitoxin and tetrodotoxin binding protein from plasma of the puffer fish, Fugu pardalis. Eur. J. Biochem. 2001, 268, 5937–5946. [Google Scholar] [CrossRef]
- Williams, B.L. Behavioral and chemical ecology of marine organisms with respect to tetrodotoxin. Mar. Drugs 2010, 8, 381–398. [Google Scholar] [CrossRef]
- Bane, V.; Brosnan, B.; Barnes, P.; Lehane, M.; Furey, A. High-resolution mass spectrometry analysis of tetrodotoxin (TTX) and its analogues in puffer fish and shellfish. Food Addit. Contam. Part A 2016, 33, 1468–1489. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organ/Tissue | Intracellular Structure | Intensity |
|---|---|---|
| Proboscis | Secretory granules of type II glandular cells | +++ |
| Myocytes | + | |
| Perinuclear area of type II glandular cells | ++ | |
| Glandular epithelium surface and proboscis lumen | + | |
| Cephalic gland | Body and duct of mucoid cells | ++ |
| Epidermis | Secretory granule of serous cells | +++ |
| Secretory granules of type I granular cells | +++ | |
| Perinuclear area of secretory cells | ++ | |
| Microvilli of ciliary cells | +++ | |
| Cytoplasm of ciliary cells | + | |
| Digestive system | Phagosomes of enterocytes | +++ |
| Cytoplasm of enterocytes | ++ | |
| Blood vessels | Cytoplasm of endotheliocytes | + |
| Myocytes | + | |
| Excretory system | Terminal cells of protonephridium | +++ |
| Lateral nerves | Neuropil | ++ |
| Cytoplasm of perikaryon | ++ | |
| Oocyte | Yolk granules | ++ |
| Body wall | Musculature | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malykin, G.V.; Chernyshev, A.V.; Magarlamov, T.Y. Intrabody Tetrodotoxin Distribution and Possible Hypothesis for Its Migration in Ribbon Worms Cephalothrix cf. simula (Palaeonemertea, Nemertea). Mar. Drugs 2021, 19, 494. https://doi.org/10.3390/md19090494
Malykin GV, Chernyshev AV, Magarlamov TY. Intrabody Tetrodotoxin Distribution and Possible Hypothesis for Its Migration in Ribbon Worms Cephalothrix cf. simula (Palaeonemertea, Nemertea). Marine Drugs. 2021; 19(9):494. https://doi.org/10.3390/md19090494
Chicago/Turabian StyleMalykin, Grigorii V., Alexei V. Chernyshev, and Timur Yu. Magarlamov. 2021. "Intrabody Tetrodotoxin Distribution and Possible Hypothesis for Its Migration in Ribbon Worms Cephalothrix cf. simula (Palaeonemertea, Nemertea)" Marine Drugs 19, no. 9: 494. https://doi.org/10.3390/md19090494
APA StyleMalykin, G. V., Chernyshev, A. V., & Magarlamov, T. Y. (2021). Intrabody Tetrodotoxin Distribution and Possible Hypothesis for Its Migration in Ribbon Worms Cephalothrix cf. simula (Palaeonemertea, Nemertea). Marine Drugs, 19(9), 494. https://doi.org/10.3390/md19090494