
Field Validation of the Southern Rock Lobster Paralytic Shellfish Toxin Monitoring Program in Tasmania, Australia

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
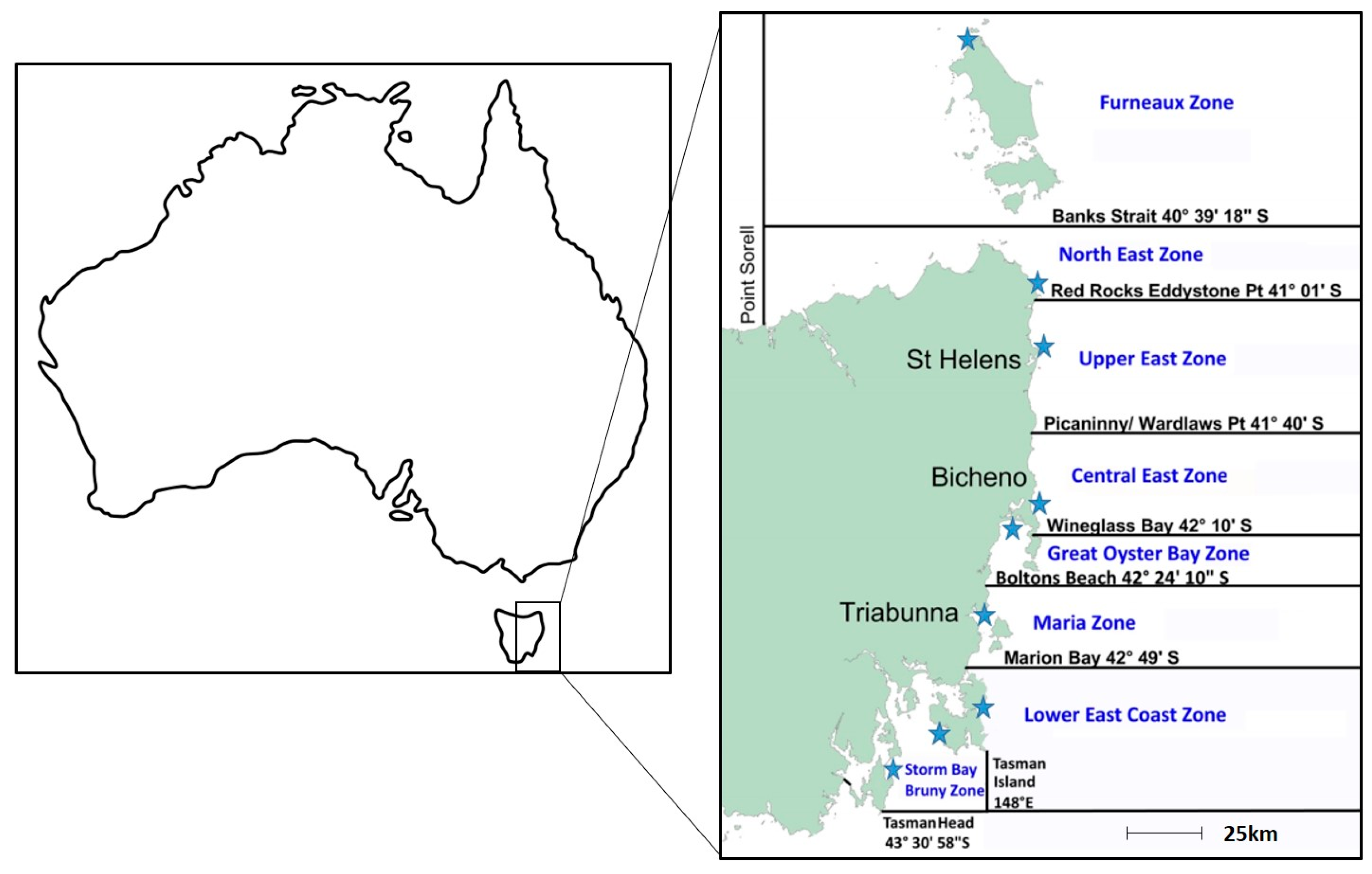
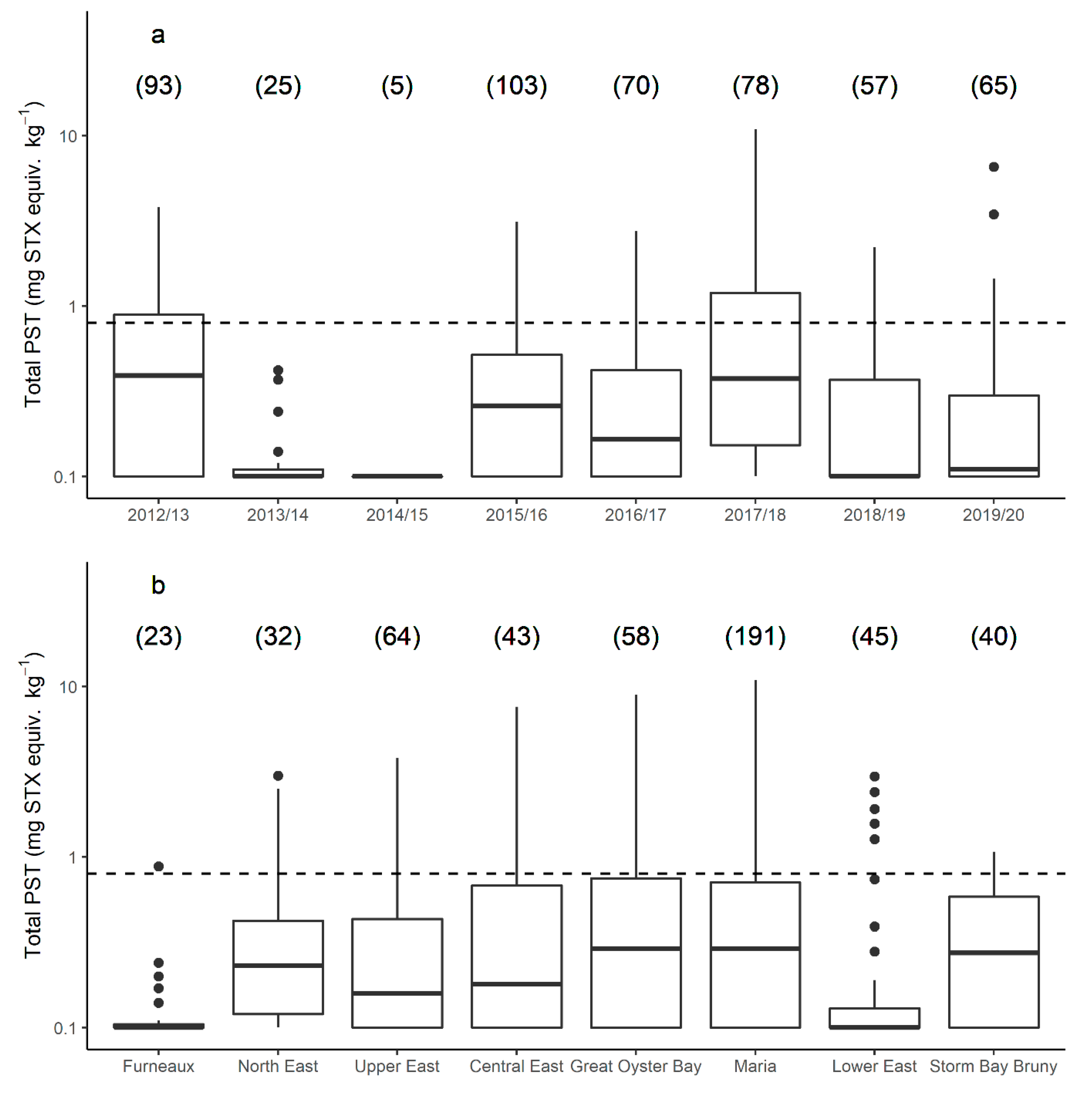
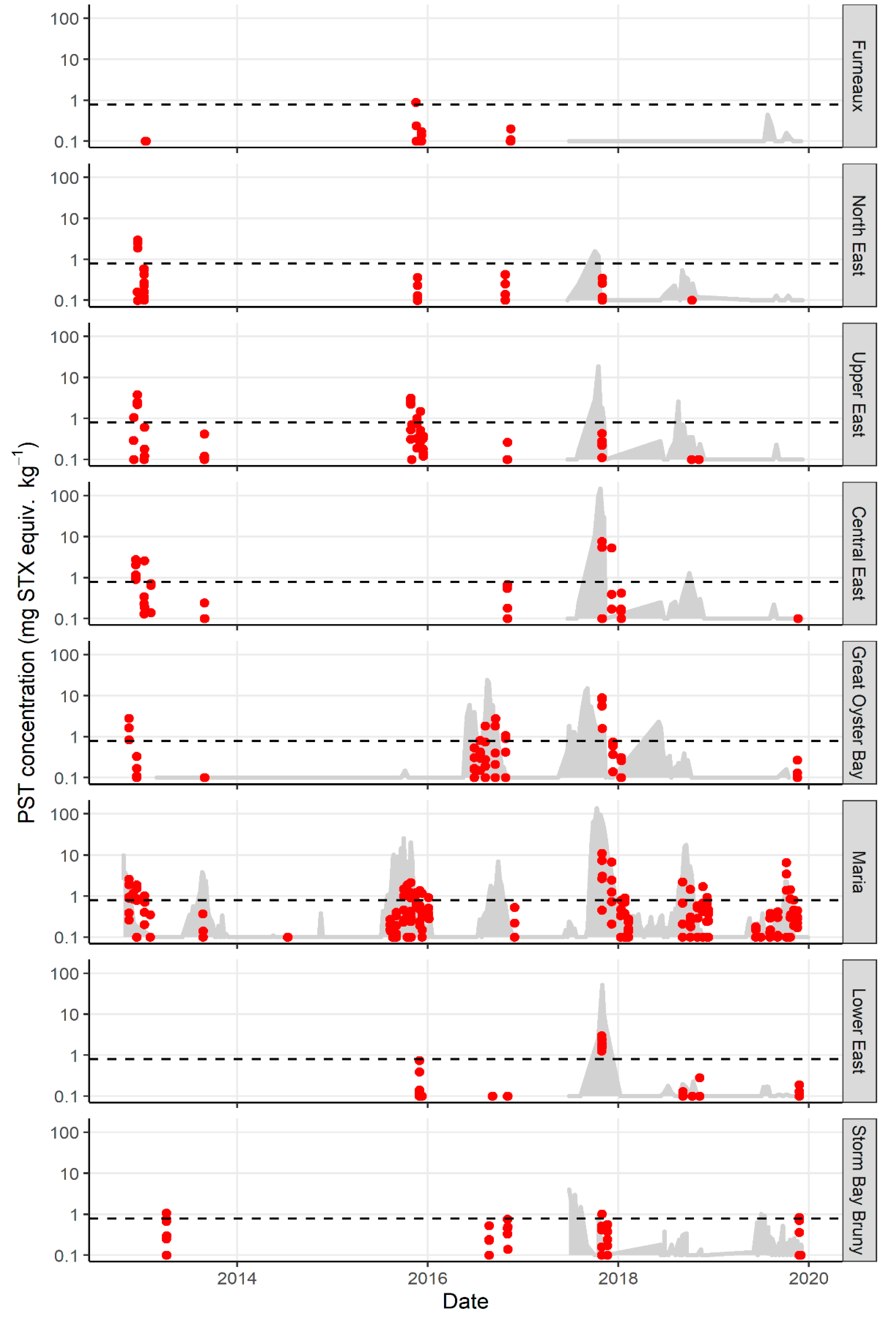
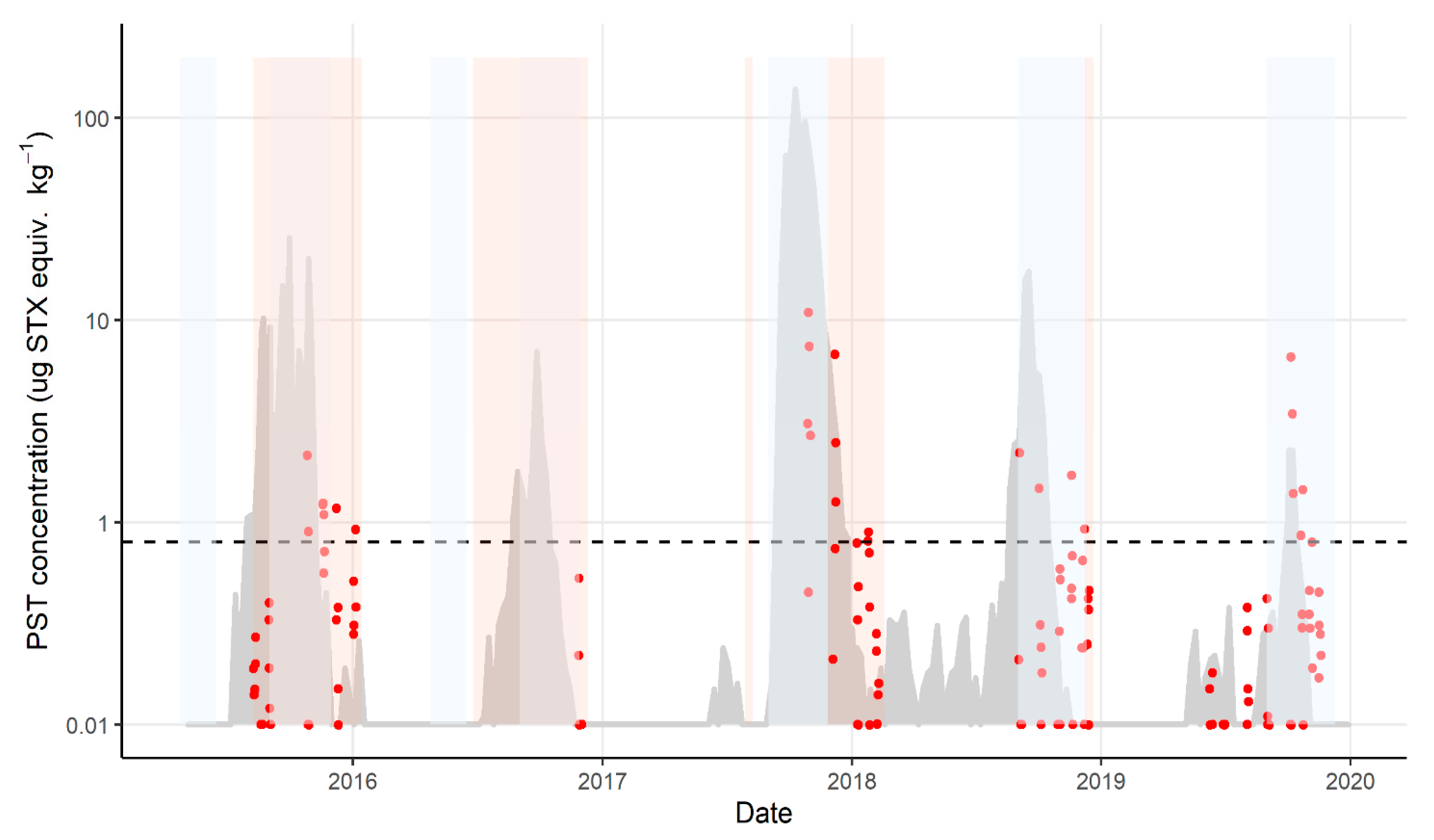
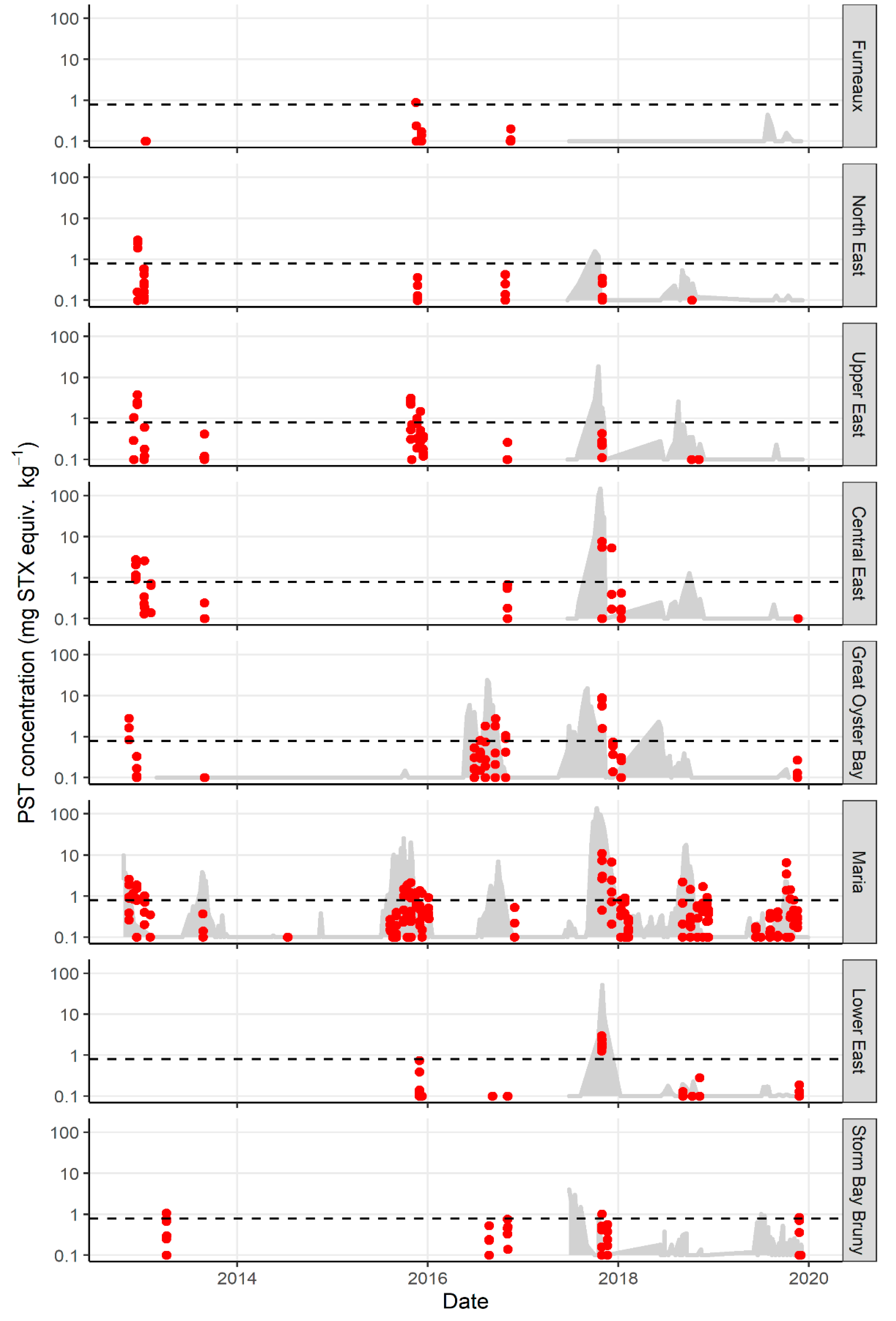
2.1. Variation in PST Levels across Sites and Time
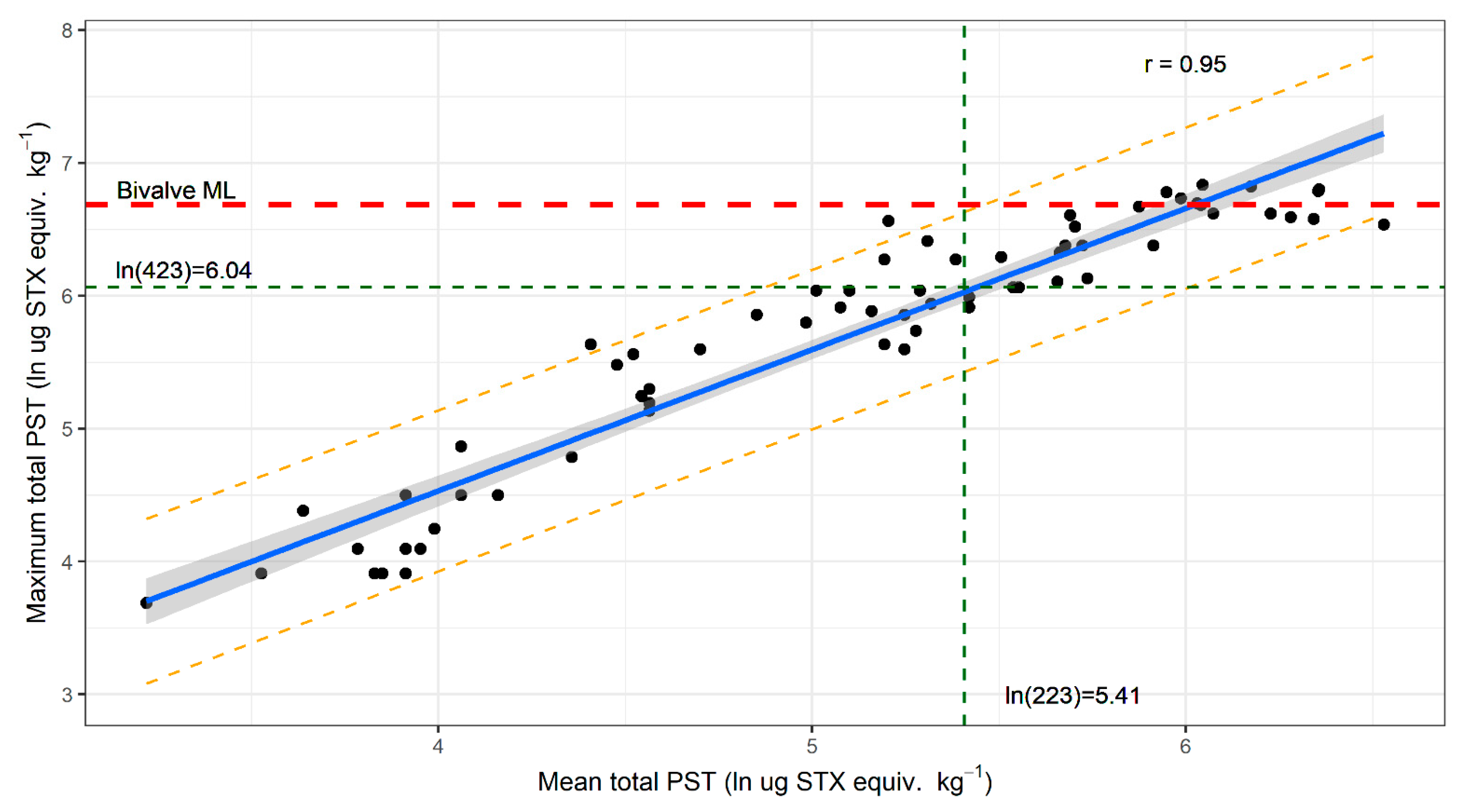
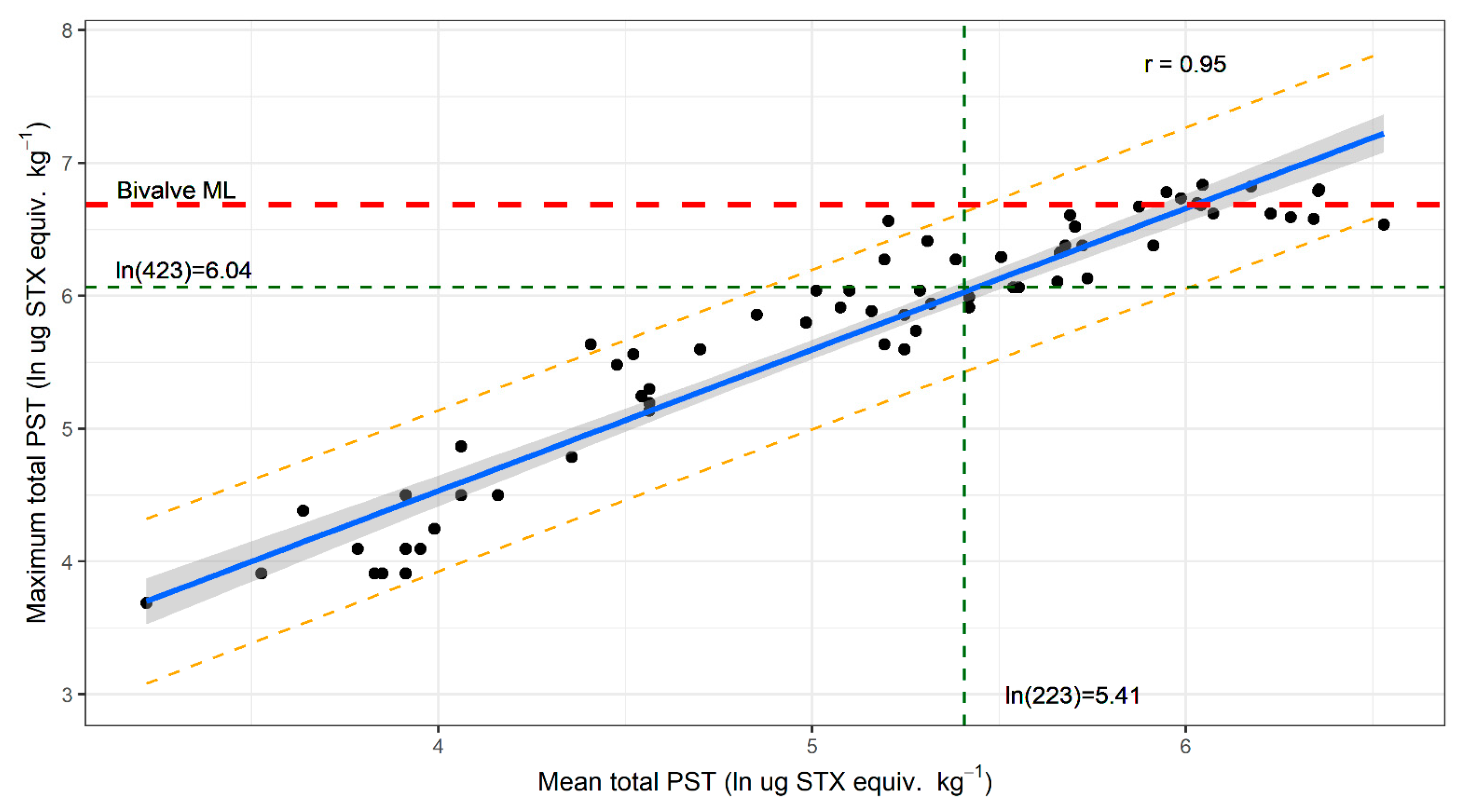
2.2. Variation in PST Accumulation within Sites
2.3. Mussels as a Sentinel Species
2.4. Field PST Accumulation and Depuration Rates of PST
2.5. The Interaction of Fisheries and Biotoxin Closures
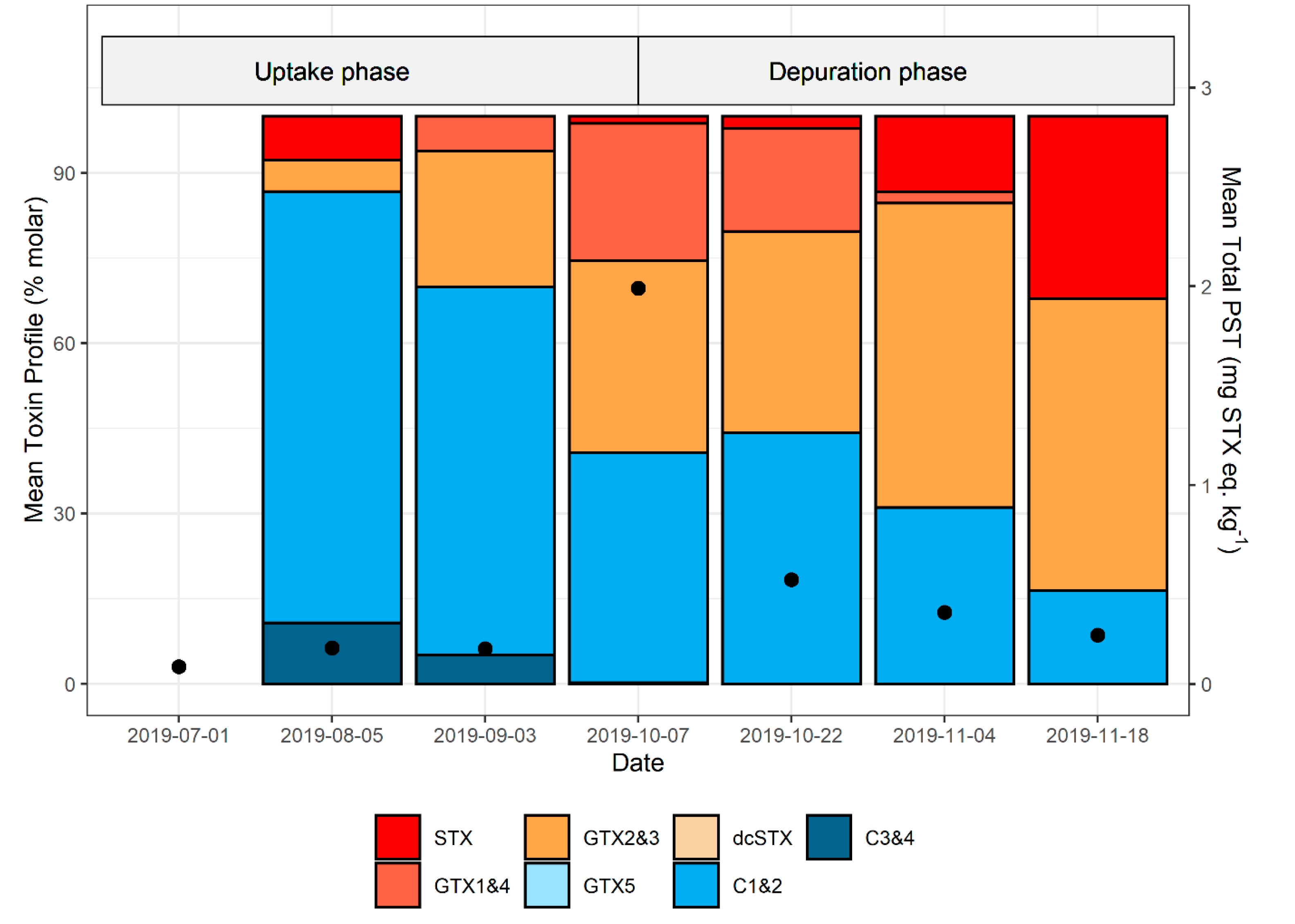
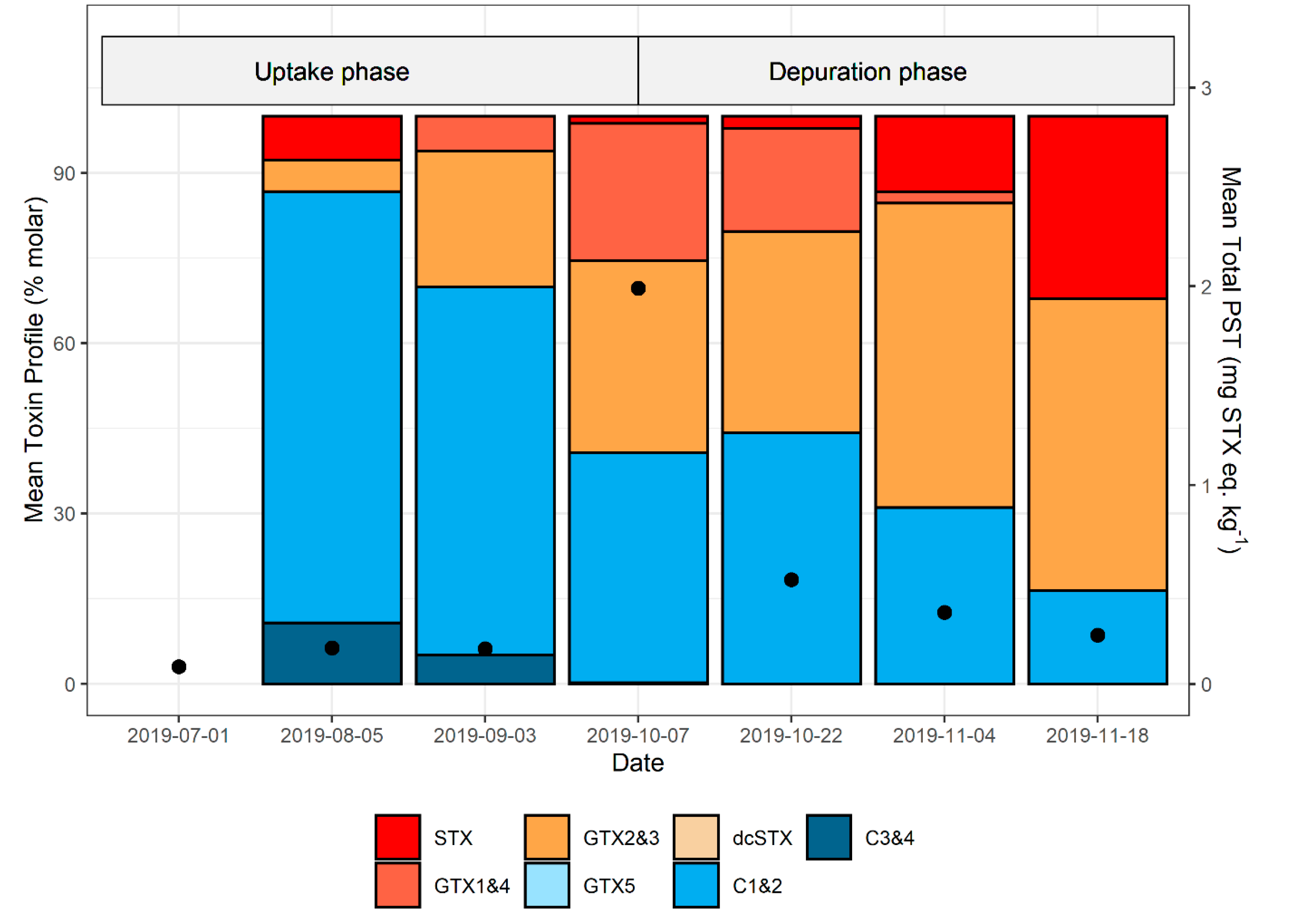
2.6. PST Profiles
3. Discussion
4. Materials and Methods
4.1. Monitoring of PST in J. edwardsii Hepatopancreas
4.2. Mussel Sentinel Sampling
4.3. PST Analysis
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Condie, S.A.; Oliver, E.C.J.; Hallegraeff, G.M. Environmental drivers of unprecedented Alexandrium catenella dinoflagellate blooms off eastern Tasmania, 2012–2018. Harmful Algae 2019, 87, 101628. [Google Scholar] [CrossRef] [PubMed]
- Campbell, A.; Hudson, D.; McLeod, C.; Nicholls, C.; Pointon, A. Tactical Research fund: Review of the 2012–13 Paralytic Shellfish Toxin Event in Tasmania Associated with the Dinoflagellate Alga, Alexandrium tamarense; FRDC Report 2012/060; South Australian Research and Development Institute: Adelaide, Australia, 2013.
- Hallegraeff, G.; Bolch, C.; Campbell, K.; Condie, C.; Dorantes-Aranda, J.; Murray, S.; Turnbull, A.; Ugalde, S. Improved Understanding of Tasmanian Harmful Algal Blooms and Biotoxin Events to Support Seafood Risk Management; Fisheries Research and Development Corporation Australia: Canberra, Australia, 2018; p. 132. [Google Scholar]
- Madigan, T.; Malhi, N.; Tan, J.; McLeod, C.; Stewart, I.; Harwood, T.; Mann, G.; Turnbull, A. Experimental uptake and depuration of paralytic shellfish toxins in Southern Rock Lobster, Jasus edwardsii. Toxicon 2018, 143, 44–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turnbull, A.; Malhi, N.; Tan, J.; Harwood, D.T.; Madigan, T. Fate of paralytic shellfish toxins in Southern Rock Lobster (Jasus edwardsii) during cooking: Concentration, composition, and distribution. J. Food Prot. 2018, 81, 240–245. [Google Scholar] [CrossRef]
- Madigan, T.; Turnbull, A.; Tan, J.; Pearn, R.; McLeod, C. Rock lobster hepatopancreas consumption data for dietary exposure assessment among recreational harvesters in Tasmania and South Australia. Hum. Ecol. Risk Assess. An. Int. J. 2018, 24, 1565–1578. [Google Scholar] [CrossRef]
- McLeod, C.; Kiermeier, A.; Stewart, I.; Tan, J.; Turnbull, A.; Madigan, T. Paralytic shellfish toxins in Australian Southern Rock Lobster (Jasus edwardsii): Acute human exposure from consumption of hepatopancreas. Hum. Ecol. Risk Assess. 2018, 24, 1872. [Google Scholar] [CrossRef]
- Turnbull, A.; Malhi, N.; Seger, A.; Harwood, T.; Jolley, J.; Fitzgibbon, Q.; Hallegraeff, G. Paralytic shellfish toxin uptake, tissue distribution, and depuration in the Southern Rock Lobster Jasus edwardsii Hutton. Harmful Algae 2020, 95, 101818. [Google Scholar] [CrossRef] [PubMed]
- Turnbull, A.; Malhi, N.; Seger, A.; Jolley, J.; Hallegraeff, G.; Fitzgibbon, Q. Accumulation of paralytic shellfish toxins by Southern Rock Lobster Jasus edwardsii causes minimal impact on lobster health. Aquat. Toxicol. 2020, 230, 105704. [Google Scholar] [CrossRef]
- Turnbull, A.; Seger, A.; Jolley, J.; Hallegraeff, G.; Knowles, G.; Fitzgibbon, Q. Lobster Supply Chains Are Not at Risk from Paralytic Shellfish Toxin Accumulation during Wet Storage. Toxins 2021, 13, 129. [Google Scholar] [CrossRef]
- DPIPWE. Rock Lobster Biotoxin Monitoring and Decision Protocols; Tasmanian Department of Primary Industries, Parks, Water and Environment Wild Fisheries Management Branch: Hobart, Australia, 2020.
- Boundy, M.J.; Selwood, A.I.; Harwood, T.D.; McNabb, P.S.; Turner, A.D. Development of a sensitive and selective liquid chromatography–mass spectrometry method for high throughput analysis of paralytic shellfish toxins using graphitised carbon solid phase extraction. J. Chromatogr. A 2015, 1387, 1–12. [Google Scholar] [CrossRef]
- Turner, A.D.; McNabb, P.S.; Harwood, T.; Selwood, A.I.; Boundy, M.J. Single-laboratory validation of a multitoxin ultra-performance LC-hydrophilic interaction LC-MS/MS method for quantitation of paralytic shellfish toxins in bivalve shellfish. J. AOAC Int. 2015, 98, 609–621. [Google Scholar] [CrossRef]
- Turnbull, A.R.; Harwood, D.T.; Boundy, M.J.; Holland, P.T.; Hallegraeff, G.; Malhi, N.; Quilliam, M.A. Paralytic shellfish toxins—Call for uniform reporting units. Toxicon 2020, 178, 59–60. [Google Scholar] [CrossRef]
- Steven, A.H.; Mobsby, D.; Curtotti, R. Australian fisheries and aquaculture statistics 2018. In Fisheries Research and Development Corporation Project 2019/093; Agricultural and Resource Economics and Sciences: Canberra, Australia, 2020. [Google Scholar] [CrossRef]
- Hallegraeff, G.M.; McCausland, M.A.; Brown, R.K. Early warning of toxic dinoflagellate blooms of Gymnodinium catenatum in southern Tasmanian waters. J. Plankton Res. 1995, 17, 1163–1176. [Google Scholar] [CrossRef]
- Hallegraeff, G.; Blackburn, S.; Doblin, M.; Bolch, C. Global toxicology, ecophysiology and population relationships of the chainforming PST dinoflagellate Gymnodinium catenatum. Harmful Algae 2012, 14, 130–143. [Google Scholar] [CrossRef]
- Watson, R.A.; Nowara, G.B.; Tracey, S.R.; Fulton, E.A.; Bulman, C.M.; Edgar, G.J.; Barrett, N.S.; Lyle, J.M.; Frusher, S.D.; Buxton, C.D. Ecosystem model of Tasmanian waters explores impacts of climate-change induced changes in primary productivity. Ecol. Model. 2013, 264, 115–129. [Google Scholar] [CrossRef]
- Edgar, G.J. Australian Marine Life: The Plants and Animals of Temperate Waters; Barnes, B., Ed.; Reed New Holland: Sydney, Australia, 2000. [Google Scholar]
- NZRLIC. National Marine Biotoxin Risk Management Plan. for the New Zealand Rock Lobster Industry; New Zealand Rock Lobster Industry Council: Wellington, New Zealand, 2019. [Google Scholar]
- California Ocean Science Trust. Frequently Asked Questions: Harmful Algal Blooms and California Fisheries. California Ocean Science Trust; 8/5/2016. Available online: https://nrm.dfg.ca.gov/FileHandler.ashx?DocumentID=128577 (accessed on 5 September 2021).
- CDFW. Health Advisories and Closures for California Finfish, Shellfish and Crustaceans. Available online: https://wildlife.ca.gov/Fishing/Ocean/Health-Advisories (accessed on 27 August 2021).
- Government of Canada. Lobster Fishing Areas 27–38 Integrated Fisheries Management Plans. Available online: https://www.dfo-mpo.gc.ca/fisheries-peches/ifmp-gmp/maritimes/2019/inshore-lobster-eng.html#toc4 (accessed on 27 August 2021).
- Lawrence, J.; Loreal, H.; Toyofuku, H.; Hess, P.; Iddya, K.; Ababouch, L. Assessment and Management of Biotoxin Risks in Bivalve Molluscs; FAO: Rome, Italy, 2011. [Google Scholar]
- Seger, A.; Hallegraeff, G.; Stone, D.; Bansemer, M.; Harwood, T.D.; Turnbull, A. Uptake of paralytic shellfish toxins by Blacklip Abalone (Haliotis rubra rubra Leach) from direct exposure to Alexandrium catenella microalgal cells and toxic aquaculture feed. Harmful Algae 2020, 99, 101925. [Google Scholar] [CrossRef]
- Madigan, T.; Tan, J.; Malhi, N.; McLeod, C.; Turnbull, A. Understanding and Reducing the Impact of Paralytic Shellfish Toxins in Southern Rock Lobster; FRDC Report 2013/713; South Australian Research and Development Institute: Adelaide, Australia, 2017.
- Jiang, T.-J.; Niu, T.; Xu, Y.-X. Transfer and metabolism of paralytic shellfish poisoning from scallop (Chlamys nobilis) to spiny lobster (Panulirus stimpsoni). Toxicon 2006, 48, 988–994. [Google Scholar] [CrossRef] [PubMed]
- Etheridge, S.M. Paralytic shellfish poisoning: Seafood safety and human health perspectives. Toxicon 2010, 56, 108–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cembella, A.D.; Desbiens, M. Fate of paralytic shellfish toxins in the American lobster Homarus americanus. J. Shellfish Res. 1994, 13, 302. [Google Scholar]
- Desbiens, M.; Cembella, A.D. Retention and possible transformation of paralytic shellfish toxins in lobster (Homarus americanus). Bull. Aquac. Assoc. Can. 1997, 2, 75–77. [Google Scholar]
- Desbiens, M.; Cembella, A.D. Occurrence and elimination kinetics of PSP toxins in the American lobster (Homarus americanus). In Harmful Marine Algal Blooms; Lassus, P., Arzul, G., Erard-Le Denn, E., Gentien, P., Marcaillou-Le Baut, C., Eds.; Lavoisier Science Publishers: Paris, France, 1995; pp. 433–438. [Google Scholar]
- Haya, K.; Oshima, Y.; Young-Lai, W.W. Profile of paralytic shellfish poisoning toxins in lobsters during uptake and depuration. In Proceedings of the Fourth Canadian Workshop on Harmful Marine Algae, Sidney, BC, Canada, 3–5 May 1994; p. 17. [Google Scholar]
- Haya, K.; Young-Lai, W.W.; Stewart, J.E.; Jellet, J.F. Uptake and excretion of paralytic shellfish toxins by lobster fed scallop digestive glands. In Proceedings of the Third Canandian workshop on harmful marine algae, Mont-Joli, QC, Canada, 12–14 May 1992; p. 38. [Google Scholar]
- Dorantes-Aranda, J.J.; Hayashi, A.; Turnbull, A.R.; Jolley, J.Y.C.; Harwood, D.T.; Hallegraeff, G.M. Detection of paralytic shellfish toxins in Southern Rock Lobster Jasus edwardsii using the qualitative Neogen™ Lateral Flow Immunoassay: Single-Laboratory Validation. J. AOAC Int. 2020, 103, 784–791. [Google Scholar] [CrossRef]
- Harwood, D.T.; Boundy, M.; Selwood, A.I.; van Ginkel, R.; MacKenzie, L.; McNabb, P.S. Refinement and implementation of the Lawrence method (AOAC 2005.06) in a commercial laboratory: Assay performance during an Alexandrium catenella bloom event. Harmful Algae 2013, 24, 20–31. [Google Scholar] [CrossRef]
- Lawrence, J.F.; Niedzwiadek, B.; Menard, C.; De Rojas Astudillo, L.; Bire, R.; Burdaspal, P.A.; Ceredi, A.; Davis, B.; Dias, E.; Eaglesham, G.; et al. Quantitative determination of paralytic shellfish poisoning toxins in shellfish using prechromatographic oxidation and liquid chromatography with fluorescence detection: Collaborative study. J. AOAC Int. 2005, 88, 1714–1732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO; WHO. Technical Paper on Toxicity Equivalency Factors for Marine Biotoxins Associated with Bivalve Molluscs; Food and Agriculture Organization of the United Nations; World Health Organization: Rome, Italy, 2016. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lobster Site & Year | Phase | J. edwardsii | M. galloprovinicalis in Spring Bay | ||
|---|---|---|---|---|---|
| Rate (mg STX. eFquiv. kg−1 day−1) (n) | Residual Variance | Rate (mg STX equiv. kg−1 day−1) (n) | Residual Variance | ||
| Maria Island 2015/16 | Uptake | 0.009 (20) | 0.66 | 0.075(9) | 0.33 |
| Okehampton 2019/20 | Uptake | 0.019 (20) | 1.16 | 0.032 (12) | 0.24 |
| Maria Island 2012/13 | Depuration | −0.031 (16) | 0.99 | −0.043 (12) | 0.26 |
| Maria Island 2017/18 | Depuration | −0.028 (25) | 0.52 | −0.072 (23) * | 0.52 |
| Okehampton 2019/20 | Depuration | −0.017 (20) | 1.20 | −0.103 (7) | 0.17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Turnbull, A.; Dorantes-Aranda, J.J.; Madigan, T.; Jolley, J.; Revill, H.; Harwood, T.; Hallegraeff, G. Field Validation of the Southern Rock Lobster Paralytic Shellfish Toxin Monitoring Program in Tasmania, Australia. Mar. Drugs 2021, 19, 510. https://doi.org/10.3390/md19090510
Turnbull A, Dorantes-Aranda JJ, Madigan T, Jolley J, Revill H, Harwood T, Hallegraeff G. Field Validation of the Southern Rock Lobster Paralytic Shellfish Toxin Monitoring Program in Tasmania, Australia. Marine Drugs. 2021; 19(9):510. https://doi.org/10.3390/md19090510
Chicago/Turabian StyleTurnbull, Alison, Juan José Dorantes-Aranda, Tom Madigan, Jessica Jolley, Hilary Revill, Tim Harwood, and Gustaaf Hallegraeff. 2021. "Field Validation of the Southern Rock Lobster Paralytic Shellfish Toxin Monitoring Program in Tasmania, Australia" Marine Drugs 19, no. 9: 510. https://doi.org/10.3390/md19090510
APA StyleTurnbull, A., Dorantes-Aranda, J. J., Madigan, T., Jolley, J., Revill, H., Harwood, T., & Hallegraeff, G. (2021). Field Validation of the Southern Rock Lobster Paralytic Shellfish Toxin Monitoring Program in Tasmania, Australia. Marine Drugs, 19(9), 510. https://doi.org/10.3390/md19090510