
Marine Alkaloids: Compounds with In Vivo Activity and Chemical Synthesis


 ,
,  ,
,  ,
, 


Abstract
:1. Introduction
2. Marine Alkaloids with Biological Activity In Vivo
2.1. Antitumor Effect
2.2. Carodioprotective Activity
2.3. Antiseizure Activity, Alzheimer’s Disease, and Mental Health
2.4. Anti-Inflammatory Activity
2.5. Antiparasitic and Antimicrobial Activity
2.6. Other Health-Related Effects
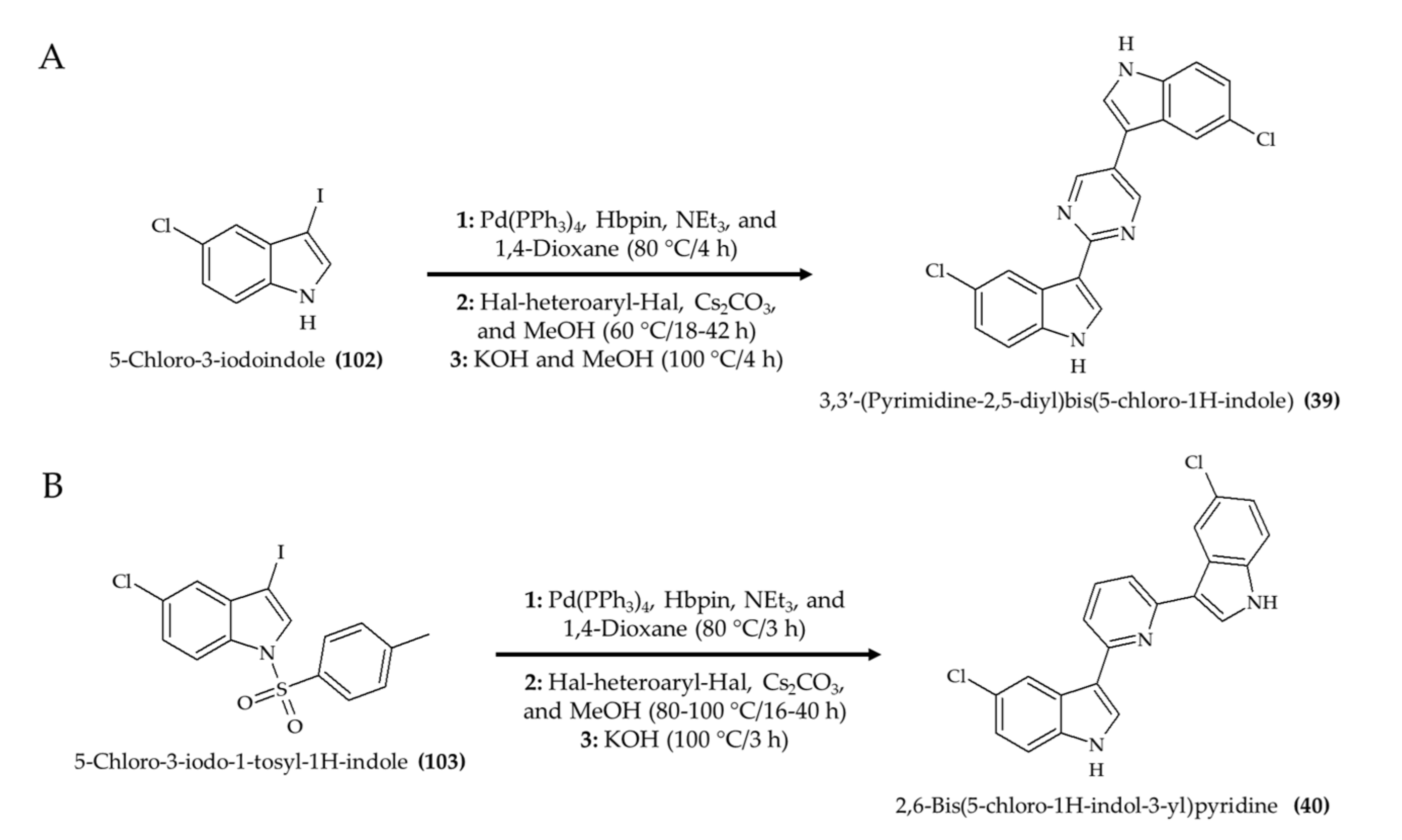
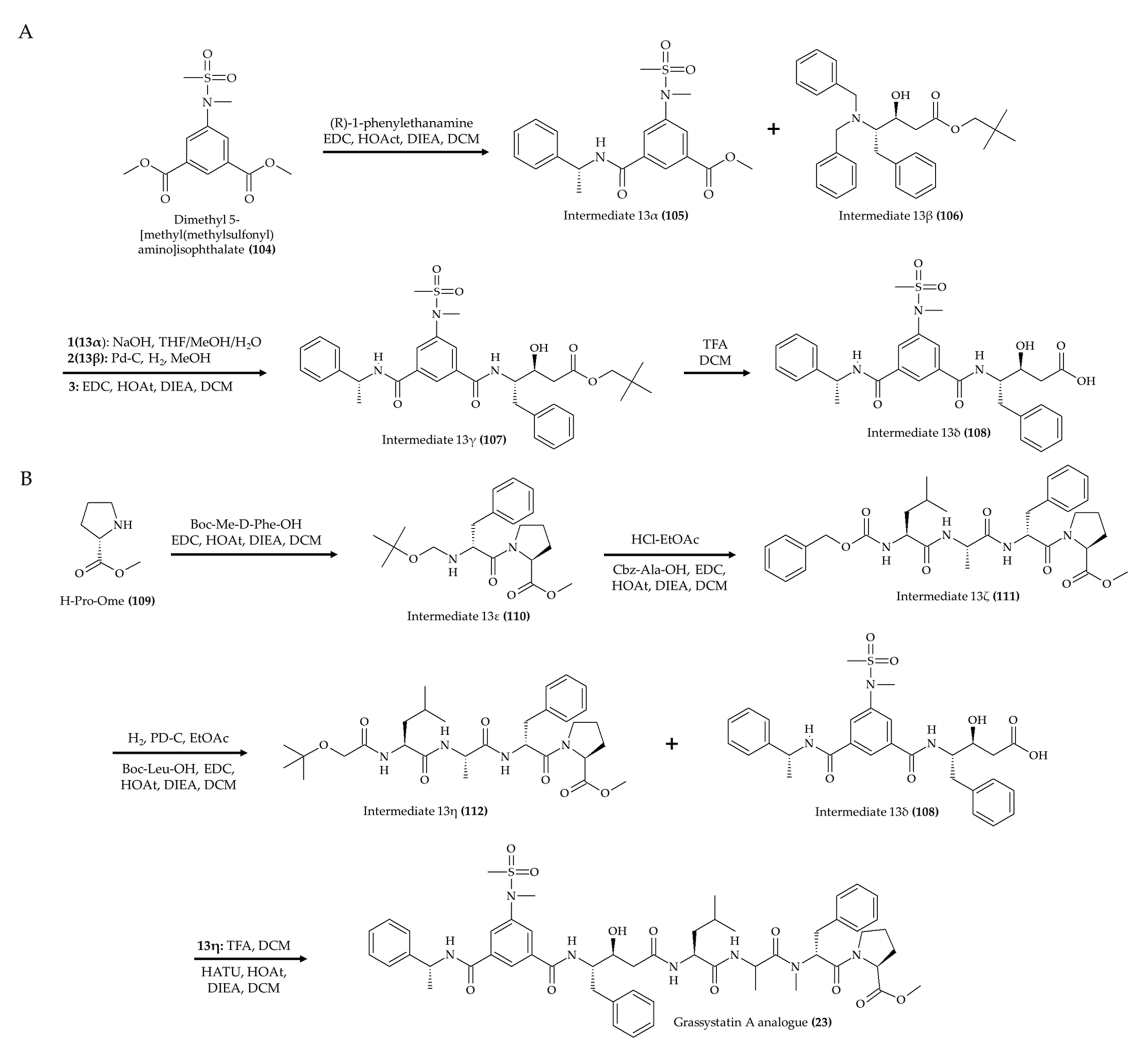
3. Chemical Modification and Synthesis of Marine Alkaloids
4. Animal Model Context and Further Considerations for Bioactive Marine Alkaloids
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kemppainen, L.M.; Kemppainen, T.T.; Reippainen, J.A.; Salmenniemi, S.T.; Vuolanto, P.H. Use of complementary and alternative medicine in Europe: Health-related and sociodemographic determinants. Scand. J. Public Health 2018, 46, 448–455. [Google Scholar] [CrossRef] [Green Version]
- Lucas, S.; Leach, M.; Kumar, S. Complementary and alternative medicine utilisation for the management of acute respiratory tract infection in children: A systematic review. Complement. Ther. Med. 2018, 37, 158–166. [Google Scholar] [CrossRef]
- Thomford, N.E.; Senthebane, D.A.; Rowe, A.; Munro, D.; Seele, P.; Maroyi, A.; Dzobo, K. Natural products for drug discovery in the 21st century: Innovations for novel drug discovery. Int. J. Mol. Sci. 2018, 19, 1578. [Google Scholar] [CrossRef] [Green Version]
- Altmann, K.H. Drugs from the oceans: Marine natural products as leads for drug discovery. Chimia 2017, 71, 646–651. [Google Scholar] [CrossRef] [PubMed]
- Netz, N.; Opatz, T. Marine indole alkaloids. Mar. Drugs 2015, 13, 4814–4914. [Google Scholar] [CrossRef] [Green Version]
- Elissawy, A.M.; Dehkordi, E.S.; Mehdinezhad, N.; Ashour, M.L.; Pour, P.M. Cytotoxic alkaloids derived from marine sponges: A comprehensive review. Biomolecules 2021, 11, 258. [Google Scholar] [CrossRef] [PubMed]
- Souza, C.R.M.; Bezerra, W.P.; Souto, J.T. Marine alkaloids with anti-inflammatory activity: Current knowledge and future perspectives. Mar. Drugs 2020, 18, 147. [Google Scholar] [CrossRef] [Green Version]
- Tempone, A.G.; Pieper, P.; Borborema, S.E.T.; Thevenard, F.; Lago, J.H.G.; Croft, S.L.; Anderson, E.A. Marine alkaloids as bioactive agents against protozoal neglected tropical diseases and malaria. Nat. Prod. Rep. 2021. [Google Scholar] [CrossRef]
- Mak, I.W.Y.; Evaniew, N.; Ghert, M. Lost in translation: Animal models and clinical trials in cancer treatment. Am. J. Transl. Res. 2014, 6, 114–118. [Google Scholar]
- Choi, C.; Cho, Y.; Son, A.; Shin, S.W.; Lee, Y.J.; Park, H.C. Therapeutic potential of (-)-Agelamide D, a diterpene alkaloid from the marine sponge Agelas sp., as a natural radiosensitizer in hepatocellular carcinoma models. Mar. Drugs 2020, 18, 500. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Huang, Y.; Huang, C.H.; Yu, J.C.; Zheng, Y.C.; Chen, Y.; She, Z.G.; Yuan, J. A Marine Alkaloid, Ascomylactam A, suppresses lung tumorigenesis via inducing cell cycle G1/S arrest through ROS/Akt/Rb pathway. Mar. Drugs 2020, 18, 494. [Google Scholar] [CrossRef]
- Sharma, S.; Guru, S.K.; Manda, S.; Kumar, A.; Mintoo, M.J.; Prasad, V.D.; Sharma, P.R.; Mondhe, D.M.; Bharate, S.B.; Bhushan, S. A marine sponge alkaloid derivative 4-chloro fascaplysin inhibits tumor growth and VEGF mediated angiogenesis by disrupting PI3K/Akt/mTOR signaling cascade. Chem. Biol. Interact. 2017, 275, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Medellin, D.C.; Zhou, Q.; Scott, R.; Hill, R.M.; Frail, S.K.; Dasari, R.; Ontiveros, S.J.; Pelly, S.C.; Van Otterlo, W.A.L.; Betancourt, T.; et al. Novel microtubule-targeting 7-Deazahypoxanthines derived from marine alkaloid rigidins with potent in vitro and in vivo anticancer activities. J. Med. Chem. 2016, 59, 480–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Rayburn, E.R.; Velu, S.E.; Nadkarni, D.H.; Murugesan, S.; Zhang, R. In vitro and in vivo anticancer activity of novel synthetic makaluvamine analogues. Clin. Cancer Res. 2009, 15, 3511–3518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.; Xu, Y.; Guo, H.; Liu, Y.; Hu, P.; Yang, X.; Li, X.; Ge, S.; Velu, S.E.; Nadkarni, D.H.; et al. Experimental therapy of ovarian cancer with synthetic makaluvamine analog: In Vitro and In vivo anticancer activity and molecular mechanisms of action. PLoS ONE 2011, 6, e20729. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Xu, H.; Zhang, X.; Voruganti, S.; Murugesan, S.; Nadkarni, D.H.; Velu, S.E.; Wang, M.H.; Wang, W.; Zhang, R. Preclinical evaluation of anticancer efficacy and pharmacological properties of FBA-TPQ, a novel synthetic Makaluvamine analog. Mar. Drugs 2012, 10, 1138–1155. [Google Scholar] [CrossRef]
- Marshall, K.M.; Matsumoto, S.S.; Holden, J.A.; Concepción, G.P.; Tasdemir, D.; Ireland, C.M.; Barrows, L.R. The anti-neoplastic and novel topoisomerase II-mediated cytotoxicity of neoamphimedine, a marine pyridoacridine. Biochem. Pharmacol. 2003, 66, 447–458. [Google Scholar] [CrossRef]
- Nishiya, N.; Oku, Y.; Ishikawa, C.; Fukuda, T.; Dan, S.; Mashima, T.; Ushijima, M.; Furukawa, Y.; Sasaki, Y.; Otsu, K.; et al. Lamellarin 14, a derivative of marine alkaloids, inhibits the T790M/C797S mutant epidermal growth factor receptor. Cancer Sci. 2021, 112, 1963–1974. [Google Scholar] [CrossRef]
- Florean, C.; Schnekenburger, M.; Lee, J.Y.; Kim, K.R.; Mazumder, A.; Song, S.; Kim, J.M.; Grandjenette, C.; Kim, J.G.; Yoon, A.Y.; et al. Discovery and characterization of Isofistularin-3, a marine brominated alkaloid, as a new DNA demethylating agent inducing cell cycle arrest and sensitization to TRAIL in cancer cells. Oncotarget 2016, 7, 24027–24049. [Google Scholar] [CrossRef] [PubMed]
- Roel, M.; Rubiolo, J.A.; Guerra-Varela, J.; Silva, S.B.L.; Thomas, O.P.; Cabezas-Sainz, P.; Sánchez, L.; López, R.; Botana, L.M. Marine guanidine alkaloids crambescidins inhibit tumor growth and activate intrinsic apoptotic signaling inducing tumor regression in a colorectal carcinoma zebrafish xenograft model. Oncotarget 2016, 7, 83071–83087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eguchi, K.; Fujiwara, Y.; Hayashida, A.; Horlad, H.; Kato, H.; Rotinsulu, H.; Losung, F.; Mangindaan, R.E.P.; De Voogd, N.J.; Takeya, M.; et al. Manzamine A, a marine-derived alkaloid, inhibits accumulation of cholesterol ester in macrophages and suppresses hyperlipidemia and atherosclerosis in vivo. Bioorganic Med. Chem. 2013, 21, 3831–3838. [Google Scholar] [CrossRef]
- Yan, T.; Wu, W.; Su, T.; Chen, J.; Zhu, Q.; Zhang, C.; Wang, X.; Bao, B. Effects of a novel marine natural product: Pyrano indolone alkaloid fibrinolytic compound on thrombolysis and hemorrhagic activities in vitro and in vivo. Arch. Pharm. Res. 2015, 38, 1530–1540. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.Q.; Li, P.H.; Chao, Y.X.; Chen, H.; Du, N.; He, Q.X.; Liu, K.C. Alkaloids with cardiovascular effects from the marine-derived fungus Penicillium expansum Y32. Mar. Drugs 2015, 13, 6489–6504. [Google Scholar] [CrossRef]
- Li, P.; Zhang, M.; Li, H.; Wang, R.; Hou, H.; Li, X.; Liu, K.; Chen, H. New prenylated indole homodimeric and pteridine alkaloids from the marine-derived fungus Aspergillus austroafricanus Y32-2. Mar. Drugs 2021, 19, 98. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Tang, X.; Luo, X.; Wang, Q.; Liu, K.; Zhang, Y.; De Voogd, N.J.; Yang, J.; Li, P.; Li, G. Agelanemoechine, a dimeric bromopyrrole alkaloid with a pro-angiogenic effect from the south China sea sponge Agelas nemoechinata. Org. Lett. 2019, 21, 9483–9486. [Google Scholar] [CrossRef] [PubMed]
- Copmans, D.; Kildgaard, S.; Rasmussen, S.A.; Ślezak, M.; Dirkx, N.; Partoens, M.; Esguerra, C.V.; Crawford, A.D.; Larsen, T.O.; De Witte, P.A.M. Zebrafish-based discovery of antiseizure compounds from the north sea: Isoquinoline alkaloids TMC-120A and TMC-120B. Mar. Drugs 2019, 17, 607. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhang, W.; Li, L.; Salvador, L.A.; Chen, T.; Chen, W.; Felsenstein, K.M.; Ladd, T.B.; Price, A.R.; Golde, T.E.; et al. Cyanobacterial peptides as a prototype for the design of potent β-secretase inhibitors and the development of selective chemical probes for other aspartic proteases. J. Med. Chem. 2012, 55, 10749–10765. [Google Scholar] [CrossRef]
- Pan, H.; Qiu, H.; Zhang, K.; Zhang, P.; Liang, W.; Yang, M.; Mou, C.; Lin, M.; He, M.; Xiao, X.; et al. Fascaplysin derivatives are potent multitarget agents against Alzheimer’s disease: In vitro and in vivo evidence. ACS Chem. Neurosci. 2019. [Google Scholar] [CrossRef]
- Zhang, C.; Hu, L.; Liu, D.; Huang, J.; Lin, W. Circumdatin D exerts neuroprotective effects by attenuating LPS-induced pro-inflammatory responses and downregulating acetylcholinesterase activity in vitro and in vivo. Front. Pharmacol. 2020, 11, 760. [Google Scholar] [CrossRef]
- Kochanowska-Karamyan, A.J.; Araujo, H.C.; Zhang, X.; El-Alfy, A.; Carvalho, P.; Avery, M.A.; Holmbo, S.D.; Magolan, J.; Hamann, M.T. Isolation and synthesis of veranamine, an antidepressant lead from the marine sponge Verongula rigida. J. Nat. Prod. 2020, 83, 1092–1098. [Google Scholar] [CrossRef]
- Cesário, H.P.S.F.; Silva, F.C.O.; Ferreira, M.K.A.; de Menezes, J.E.S.A.; dos Santos, H.S.; Nogueira, C.E.S.; de L. Silva, K.S.B.; Hajdu, E.; Silveira, E.R.; Pessoa, O.D.L. Anxiolytic-like effect of brominated compounds from the marine sponge Aplysina fulva on adult zebrafish (Danio rerio): Involvement of the GABAergic system. Neurochem. Int. 2021, 146, 105021. [Google Scholar] [CrossRef]
- Ahmad, T.B.; Rudd, D.; Benkendorff, K.; Mahdi, L.K.; Pratt, K.A.; Dooley, L.; Wei, C.; Kotiw, M. Brominated indoles from a marine mollusc inhibit inflammation in a murine model of acute lung injury. PLoS ONE 2017, 12, e0186904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, Z.; Liu, Q.; Xing, C.; Zhang, Y.; Zhou, Y.; Zhang, J.; Liu, H.; Cao, M.; Yang, X.; Liu, G. Viridicatol isolated from deep-sea Penicillium Griseofulvum alleviates anaphylaxis and repairs the intestinal barrier in mice by suppressing mast cell activation. Mar. Drugs 2020, 18, 517. [Google Scholar] [CrossRef] [PubMed]
- Lucena, A.M.M.; Souza, C.R.M.; Jales, J.T.; Guedes, P.M.M.; De Miranda, G.E.C.; de Moura, A.M.A.; Araújo-Júnior, J.X.; Nascimento, G.J.; Scortecci, K.C.; Santos, B.V.O.; et al. The bisindole alkaloid caulerpin, from seaweeds of the genus Caulerpa, attenuated colon damage in murine colitis model. Mar. Drugs 2018, 16, 318. [Google Scholar] [CrossRef] [Green Version]
- Gui, Y.H.; Liu, L.; Wu, W.; Zhang, Y.; Jia, Z.L.; Shi, Y.P.; Kong, H.T.; Liu, K.C.; Jiao, W.H.; Lin, H.W. Discovery of nitrogenous sesquiterpene quinone derivatives from sponge Dysidea septosa with anti-inflammatory activity in vivo zebrafish model. Bioorg. Chem. 2020, 94, 103435. [Google Scholar] [CrossRef] [PubMed]
- Davis, R.A.; Buchanan, M.S.; Duffy, S.; Avery, V.M.; Charman, S.A.; Charman, W.N.; White, K.L.; Shackleford, D.M.; Edstein, M.D.; Andrews, K.T.; et al. Antimalarial activity of pyrroloiminoquinones from the Australian marine sponge Zyzzya sp. J. Med. Chem. 2012, 55, 5851–5858. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, M.G.; Cardoso, J.F.; Perasoli, F.B.; Branquinho, R.T.; Mourão, R.S.; Tavares, H.D.S.; Xocaira, M.L.C.T.; Guimarães, D.S.M.; Viana, G.H.R.; Varotti, F.D.P.; et al. Nanoemulsion composed of 10-(4,5-dihydrothiazol-2-yl)thio)decan-1-ol), a synthetic analog of 3-alkylpiridine marine alkaloid: Development, characterization, and antimalarial activity. Eur. J. Pharm. Sci. 2020, 151, 105382. [Google Scholar] [CrossRef] [PubMed]
- Mani, L.; Petek, S.; Valentin, A.; Chevalley, S.; Folcher, E.; Aalbersberg, W.; Debitus, C. The in vivo anti-plasmodial activity of haliclonacyclamine A, an alkaloid from the marine sponge, Haliclona sp. Nat. Prod. Res. 2011, 25, 1923–1930. [Google Scholar] [CrossRef] [PubMed]
- Pech-Puch, D.; Pérez-Povedano, M.; Martinez-Guitian, M.; Lasarte-Monterrubio, C.; Vázquez-Ucha, J.C.; Bou, G.; Rodríguez, J.; Beceiro, A.; Jimenez, C. In vitro and in vivo assessment of the efficacy of bromoageliferin, an alkaloid isolated from the sponge Agelas dilatata, against Pseudomonas aeruginosa. Mar. Drugs 2020, 18, 326. [Google Scholar] [CrossRef] [PubMed]
- Andrade, J.T.; Lima, W.G.; Sousa, J.F.; Saldanha, A.A.; De Sá, N.P.; Morais, F.B.; Prates Silva, M.K.; Ribeiro Viana, G.H.; Johann, S.; Soares, A.C.; et al. Design, synthesis, and biodistribution studies of new analogues of marine alkaloids: Potent in vitro and in vivo fungicidal agents against Candida spp. Eur. J. Med. Chem. 2021, 210, 113048. [Google Scholar] [CrossRef]
- Rehberg, N.; Sommer, G.A.; Drießen, D.; Kruppa, M.; Adeniyi, E.T.; Chen, S.; Wang, L.; Wolf, K.; Tasch, B.O.A.; Ioerger, T.R.; et al. Nature-inspired (di)azine-bridged bisindole alkaloids with potent antibacterial in vitro and in vivo efficacy against methicillin-resistant Staphylococcus aureus. J. Med. Chem. 2020, 63, 12623–12641. [Google Scholar] [CrossRef]
- Wang, Q.; Chen, D.; Jin, H.; Ye, Z.; Wang, C.; Chen, K.; Kuek, V.; Xu, K.; Qiu, H.; Chen, P.; et al. Hymenialdisine: A marine natural product that acts on both osteoblasts and osteoclasts and prevents estrogen-dependent bone loss in mice. J. Bone Miner. Res. 2020, 35, 1582–1596. [Google Scholar] [CrossRef]
- Carlile, G.W.; Keyzers, R.A.; Teske, K.A.; Robert, R.; Williams, D.E.; Linington, R.G.; Gray, C.A.; Centko, R.M.; Yan, L.; Anjos, S.M.; et al. Correction of F508del-CFTR trafficking by the sponge alkaloid latonduine is modulated by interaction with PARP. Chem. Biol. 2012, 19, 1288–1299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, W.H.; Li, J.; Zhang, M.M.; Cui, J.; Gui, Y.H.; Zhang, Y.; Li, J.Y.; Liu, K.C.; Lin, H.W. Frondoplysins A and B, unprecedented terpene-alkaloid bioconjugates from Dysidea frondosa. Org. Lett. 2019, 21, 6190–6193. [Google Scholar] [CrossRef] [PubMed]
- Ballane, G.; Cauley, J.A.; Luckey, M.M.; El-Hajj Fuleihan, G. Worldwide prevalence and incidence of osteoporotic vertebral fractures. Osteoporos. Int. 2017, 28, 1531–1542. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.; Liu, J.; Xu, S.; Zhu, Z.; Xu, J. The structural modification of natural products for novel drug discovery. Expert Opin. Drug Discov. 2017, 12, 121–140. [Google Scholar] [CrossRef]
- Lam, K.S. New aspects of natural products in drug discovery. Trends Microbiol. 2007, 15, 279–289. [Google Scholar] [CrossRef]
- Chen, M.; Shao, C.L.; Fu, X.M.; Xu, R.F.; Zheng, J.J.; Zhao, D.L.; She, Z.G.; Wang, C.Y. Bioactive indole alkaloids and phenyl ether derivatives from a marine-derived Aspergillus sp. fungus. J. Nat. Prod. 2013, 76, 547–553. [Google Scholar] [CrossRef]
- Lyakhova, I.A.; Bryukhovetsky, I.S.; Kudryavtsev, I.V.; Khotimchenko, Y.S.; Zhidkov, M.E.; Kantemirov, A.V. Antitumor activity of fascaplysin derivatives on glioblastoma model in vitro. Bull. Exp. Biol. Med. 2018, 164, 666–672. [Google Scholar] [CrossRef]
- Parra, L.L.L.; Bertonha, A.F.; Severo, I.R.M.; Aguiar, A.C.C.; De Souza, G.E.; Oliva, G.; Guido, R.V.C.; Grazzia, N.; Costa, T.R.; Miguel, D.C.; et al. Isolation, derivative synthesis, and structure-activity relationships of antiparasitic bromopyrrole alkaloids from the marine sponge Tedania brasiliensis. J. Nat. Prod. 2018, 81, 188–202. [Google Scholar] [CrossRef]
- Zheng, L.; Gao, T.; Ge, Z.; Ma, Z.; Xu, J.; Ding, W.; Shen, L. Design, synthesis and structure-activity relationship studies of glycosylated derivatives of marine natural product Lamellarin D. Eur. J. Med. Chem. 2021, 214, 113226. [Google Scholar] [CrossRef]
- Shinkre, B.A.; Raisch, K.P.; Fan, L.; Velu, S.E. Analogs of the marine alkaloid makaluvamines: Synthesis, topoisomerase II inhibition, and anticancer activity. Bioorganic Med. Chem. Lett. 2007, 17, 2890–2893. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Ezell, S.J.; Zhang, Y.; Wang, W.; Rayburn, E.R.; Nadkarni, D.H.; Murugesan, S.; Velu, S.E.; Zhang, R. FBA-TPQ, a novel marine-derived compound as experimental therapy for prostate cancer. Invest. New Drugs 2010, 28, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, D.S.M.; de Sousa Luz, L.S.; do Nascimento, S.B.; Silva, L.R.; de Miranda Martins, N.R.; de Almeida, H.G.; de Souza Reis, V.; Maluf, S.E.C.; Budu, A.; Marinho, J.A.; et al. Improvement of antimalarial activity of a 3-alkylpiridine alkaloid analog by replacing the pyridine ring to a thiazole-containing heterocycle: Mode of action, mutagenicity profile, and Caco-2 cell-based permeability. Eur. J. Pharm. Sci. 2019, 138, 105015. [Google Scholar] [CrossRef]
- Li, L.; Abraham, A.D.; Zhou, Q.; Ali, H.; O’Brien, J.V.; Hamill, B.D.; Arcaroli, J.J.; Messersmith, W.A.; LaBarbera, D.V. An improved high yield total synthesis and cytotoxicity study of the marine alkaloid neoamphimedine: An ATP-competitive inhibitor of topoisomerase IIα and potent anticancer agent. Mar. Drugs 2014, 12, 4833–4850. [Google Scholar] [CrossRef] [Green Version]
- LaBarbera, D.V.; Bugni, T.S.; Ireland, C.M. The total synthesis of neoamphimedine. J. Org. Chem. 2007, 72, 8501–8505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuda, T.; Umeki, T.; Tokushima, K.; Xiang, G.; Yoshida, Y.; Ishibashi, F.; Oku, Y.; Nishiya, N.; Uehara, Y.; Iwao, M. Design, synthesis, and evaluation of A-ring-modified lamellarin N analogues as noncovalent inhibitors of the EGFR T790M/L858R mutant. Bioorganic Med. Chem. 2017, 25, 6563–6580. [Google Scholar] [CrossRef] [Green Version]
- Komatsubara, M.; Umeki, T.; Fukuda, T.; Iwao, M. Modular synthesis of lamellarins via regioselective assembly of 3,4,5-differentially arylated pyrrole-2-carboxylates. J. Org. Chem. 2014, 79, 529–537. [Google Scholar] [CrossRef] [PubMed]
- Jakubec, P.; Hawkins, A.; Felzmann, W.; Dixon, D.J. Total synthesis of manzamine A and related alkaloids. J. Am. Chem. Soc. 2012, 134, 17482–17485. [Google Scholar] [CrossRef]
- Martin, S.F.; Humphrey, J.M.; Ali, A.; Hillier, M.C. Enantioselective total syntheses of ircinal A and related manzamine alkaloids. J. Am. Chem. Soc. 1999, 121, 866–867. [Google Scholar] [CrossRef]
- Winkler, J.D.; Axten, J.M. The first total syntheses of ircinol A, ircinal A, and manzamines A and D. J. Am. Chem. Soc. 1998, 120, 6425–6426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humphrey, J.M.; Liao, Y.; Ali, A.; Rein, T.; Wong, Y.L.; Chen, H.J.; Courtney, A.K.; Martin, S.F. Enantioselective total syntheses of manzamine A and related alkaloids. J. Am. Chem. Soc. 2002, 124, 8584–8592. [Google Scholar] [CrossRef]
- Toma, T.; Kita, Y.; Fukuyama, T. Total synthesis of (+)-Manzamine A. J. Am. Chem. Soc. 2010, 132, 10233–10235. [Google Scholar] [CrossRef] [PubMed]
- Kapourchali, F.R.; Surendiran, G.; Chen, L.; Uitz, E.; Bahadori, B.; Moghadasian, M.H. Animal models of atherosclerosis. World J. Clin. Cases WJCC 2014, 2, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Kitajima, S.; Watanabe, T.; Xu, J.; Zhang, J.; Liu, E.; Chen, Y.E. Rabbit models for the study of human atherosclerosis: From pathophysiological mechanisms to translational medicine. Pharmacol. Ther. 2015, 146, 104–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, N.; Lai, L. Genetically modified pig models for human diseases. J. Genet. Genomics 2013, 40, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Kirla, K.T.; Erhart, C.; Groh, K.J.; Stadnicka-Michalak, J.; Eggen, R.I.L.; Schirmer, K.; Kraemer, T. Zebrafish early life stages as alternative model to study ‘designer drugs’: Concordance with mammals in response to opioids. Toxicol. Appl. Pharmacol. 2021, 419, 115483. [Google Scholar] [CrossRef] [PubMed]
- Denayer, T.; Stöhrn, T.; Van Roy, M. Animal models in translational medicine: Validation and prediction. New Horizons Transl. Med. 2014, 2, 5–11. [Google Scholar] [CrossRef] [Green Version]
- McGonigle, P.; Ruggeri, B. Animal models of human disease: Challenges in enabling translation. Biochem. Pharmacol. 2014, 87, 162–171. [Google Scholar] [CrossRef]
- Jung, J. Human tumor xenograft models for preclinical assessment of anticancer drug development. Toxicol. Res. 2014, 30, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Getz, G.S.; Reardon, C.A. Animal models of Atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 1104–1115. [Google Scholar] [CrossRef] [Green Version]
- Teng, M.W.L.; Galon, J.; Fridman, W.H.; Smyth, M.J. From mice to humans: Developments in cancer immunoediting. J. Clin. Invest. 2015, 125, 3338–3346. [Google Scholar] [CrossRef] [Green Version]
- Camacho, P.; Fan, H.; Liu, Z.; He, J.Q. Small mammalian animal models of heart disease. Am. J. Cardiovasc. Dis. 2016, 6, 70–80. [Google Scholar] [PubMed]
- Löscher, W.; Ferland, R.J.; Ferraro, T.N. The relevance of inter- and intrastrain differences in mice and rats and their implications for models of seizures and epilepsy. Epilepsy Behav. 2017, 73, 214–235. [Google Scholar] [CrossRef]
- Xie, Y.; Meijer, A.H.; Schaaf, M.J.M. Modeling inflammation in zebrafish for the development of anti-inflammatory drugs. Front. Cell Dev. Biol. 2021, 8, 620984. [Google Scholar] [CrossRef]
- Zanandrea, R.; Bonan, C.D.; Campos, M.M. Zebrafish as a model for inflammation and drug discovery. Drug Discov. Today 2020, 25, 2201–2211. [Google Scholar] [CrossRef]
- Brown, H.K.; Schiavone, K.; Tazzyman, S.; Heymann, D.; Chico, T.J.A. Zebrafish xenograft models of cancer and metastasis for drug discovery. Expert Opin. Drug Discov. 2017, 12, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Nathan, J.; Kannan, R.R. Antiangiogenic molecules from marine actinomycetes and the importance of using zebrafish model in cancer research. Heliyon 2020, 6, e05662. [Google Scholar] [CrossRef] [PubMed]
- Hason, M.; Bartůnĕk, P. Zebrafish models of cancer-new insights on modeling human cancer in a non-mammalian vertebrate. Genes 2019, 10, 935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letrado, P.; De Miguel, I.; Lamberto, I.; Díez-Martínez, R.; Oyarzabal, J. Zebrafish: Speeding up the cancer drug discovery process. Cancer Res. 2018, 78, 6048–6058. [Google Scholar] [CrossRef] [Green Version]
- Giardoglou, P.; Beis, D. On zebrafish disease models and matters of the heart. Biomedicines 2019, 7, 15. [Google Scholar] [CrossRef] [Green Version]
- Fontana, B.D.; Mezzomo, N.J.; Kalueff, A.V.; Rosemberg, D.B. The developing utility of zebrafish models of neurological and neuropsychiatric disorders: A critical review. Exp. Neurol. 2018, 299, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Cekanova, M.; Rathore, K. Animal models and therapeutic molecular targets of cancer: Utility and limitations. Drug Des. Devel. Ther. 2014, 8, 1911–1922. [Google Scholar] [CrossRef] [Green Version]
- Turner, A.S. Animal models of osteoporosis - Necessity and limitations. Eur. Cells Mater. 2001, 1, 66–81. [Google Scholar] [CrossRef] [PubMed]
- Gut, P.; Reischauer, S.; Stainier, D.Y.R.; Arnaout, R. Little fish, big data: Zebrafish as a model for cardiovascular and metabolic disease. Physiol. Rev. 2017, 97, 889–938. [Google Scholar] [CrossRef] [PubMed]
- Hay, M.; Thomas, D.W.; Craighead, J.L.; Economides, C.; Rosenthal, J. Clinical development success rates for investigational drugs. Nat. Biotechnol. 2014, 32, 40–51. [Google Scholar] [CrossRef]
- He, H.; Liu, L.; Morin, E.E.; Liu, M.; Schwendeman, A. Survey of clinical translation of cancer nanomedicines - Lessons learned from successes and failures. Acc. Chem. Res. 2019, 52, 2673–2683. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound and Source | Experimental Conditions | Effect | Ref. |
|---|---|---|---|
| Anticancer activity | |||
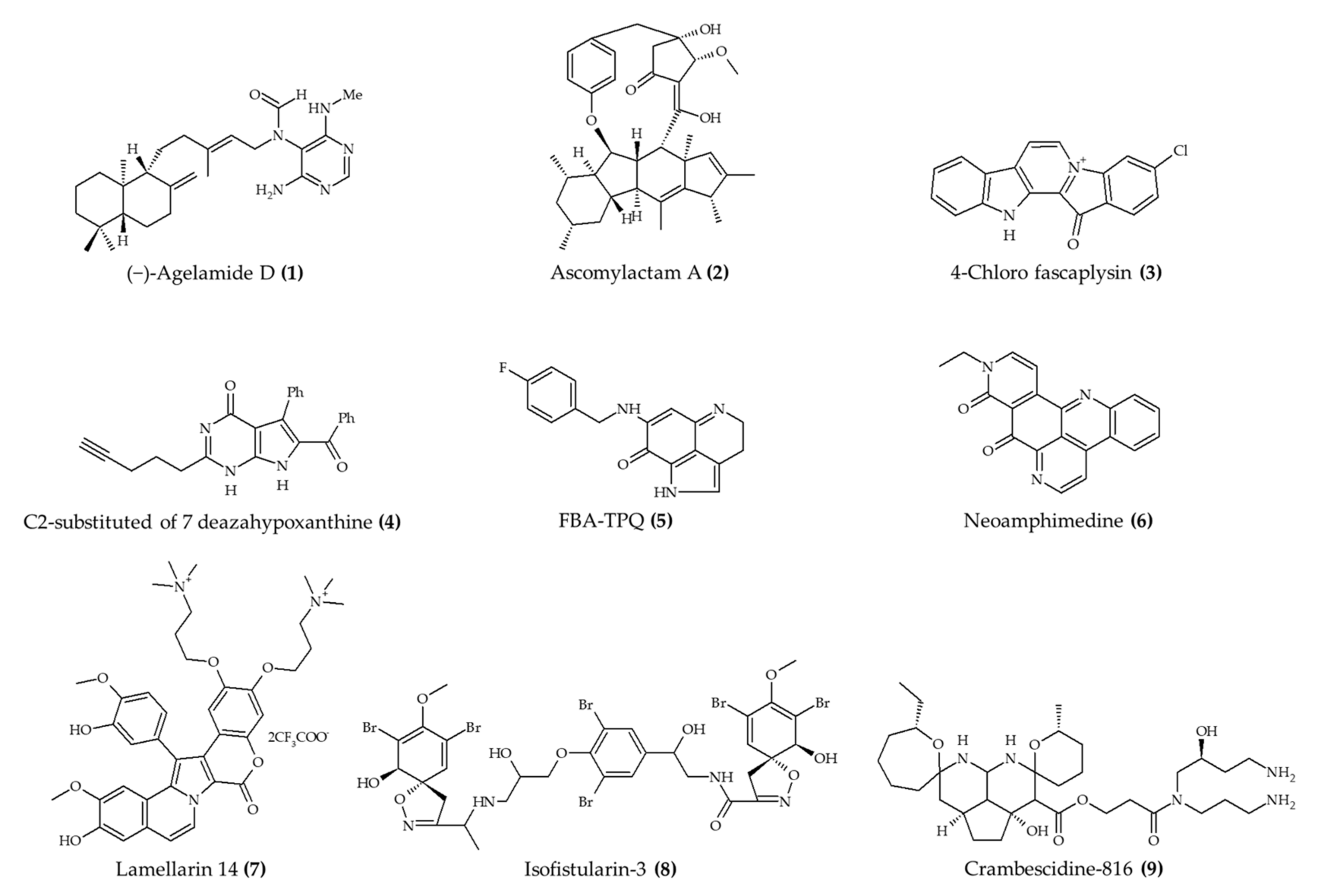
| (−)-Agelamide D (1) from marine sponge Agelas sp. | 6–7-week-old male Balb/c nude mice; intraperitoneal injection; 1.25 mg/kg/day; 3 times a week; mice euthanized after 21 days | Increased the efficacy of radiation therapy in xenograft Hep3B cells with reduced systemic toxicity | [10] |
| Ascomylactam A (2) from mangrove endophytic fungus Ascomycota sp. | Male BALB/c-nu mice; intraperitoneal injection; 3 and 6 (A549 and NCI-H460), 5 or 10 (NCI-H1975) mg/kg/day; every 3 days for 21 days | Suppression of A549 (6 mg/kg/day; volume) and NCI-H460 (6 mg/kg/day; volume and weight) and lung tumor growth; no effect in NCI-H1975 tumor | [11] |
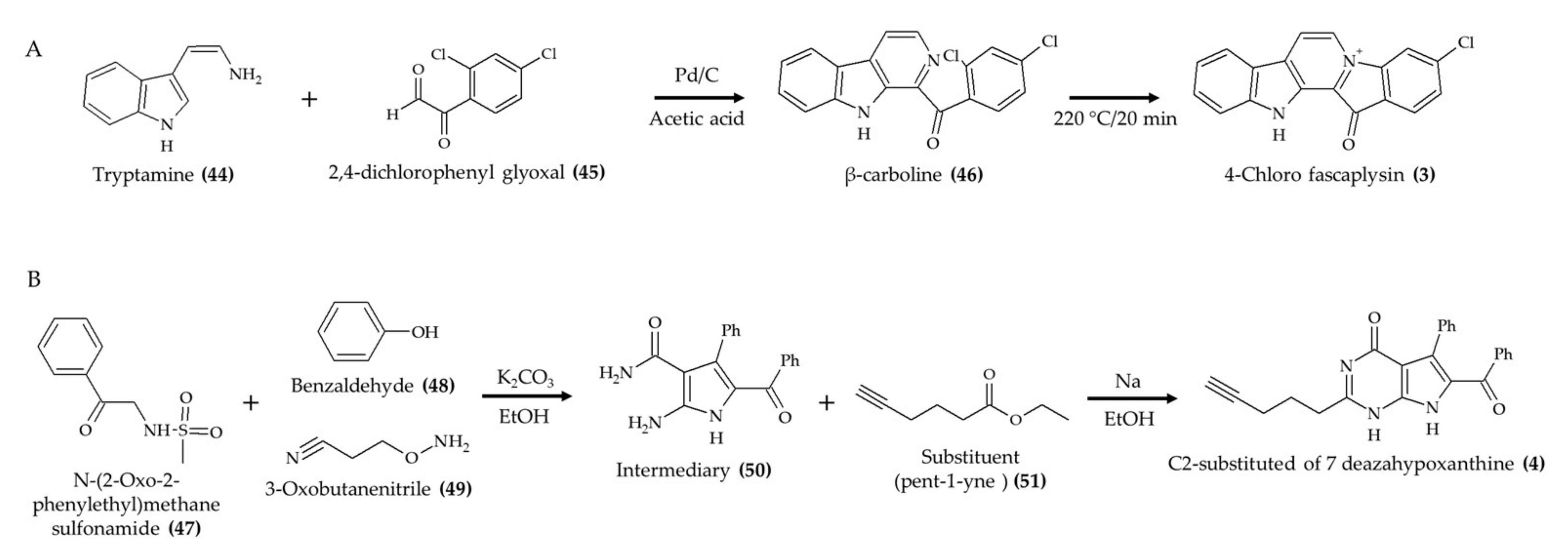
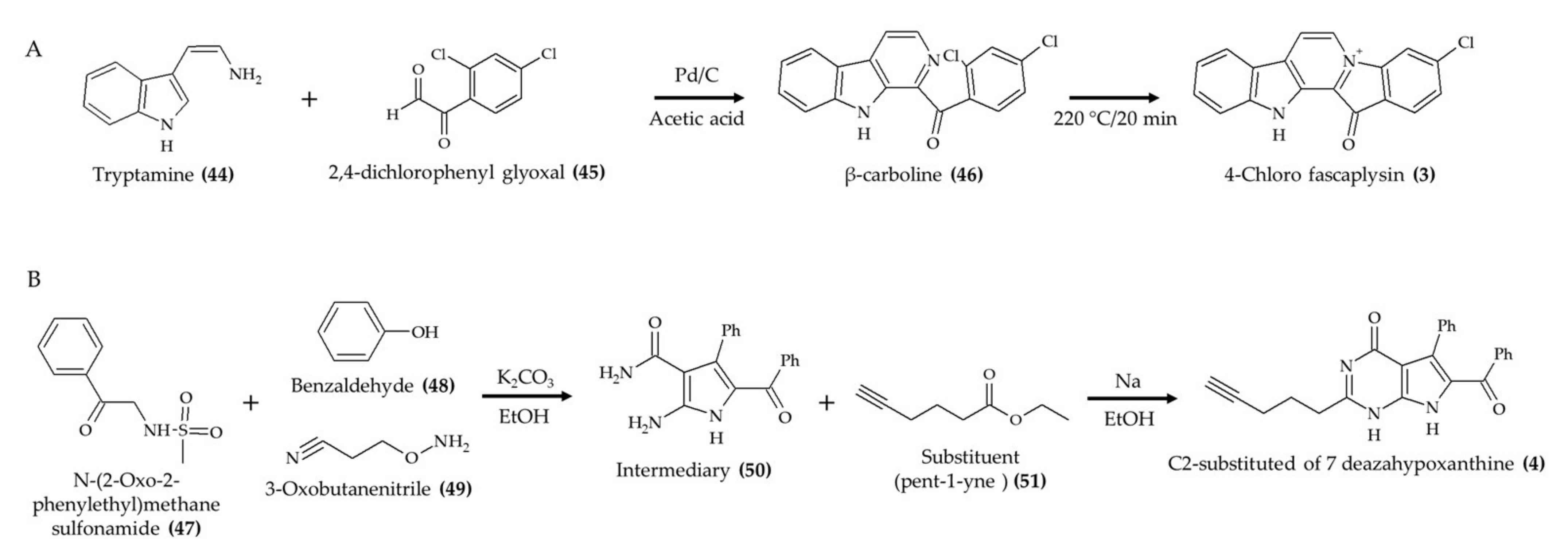
| Synthetic 4-chloro fascaplysin (3) (found in marine sponge) | 4–6-week-old C57B/6J female mice; oral administration; 1, 3, and 5 mg/kg/day for 5 days (tumor angiogenesis assay); 4 and 7 mg/kg/day for 8 days (Ehrlich solid tumor model) | Reduced the formation of VEGF-mediated microvessels and blood vessel formation of xenograft breast cancer cells (1, 3, and 5 mg/kg/day) and Ehrlich solid tumor formation (4 and 7 mg/kg/day) | [12] |
| Synthetic derivative (C2-substituted) of 7-deazahypoxanthine (4) from rigidin D | 4–6-week-old female athymic nude mice; intraperitoneal injection; 3 mg/kg; 5 times per week for 17 days | Reduced tumor growth; no effect on weight | [13] |
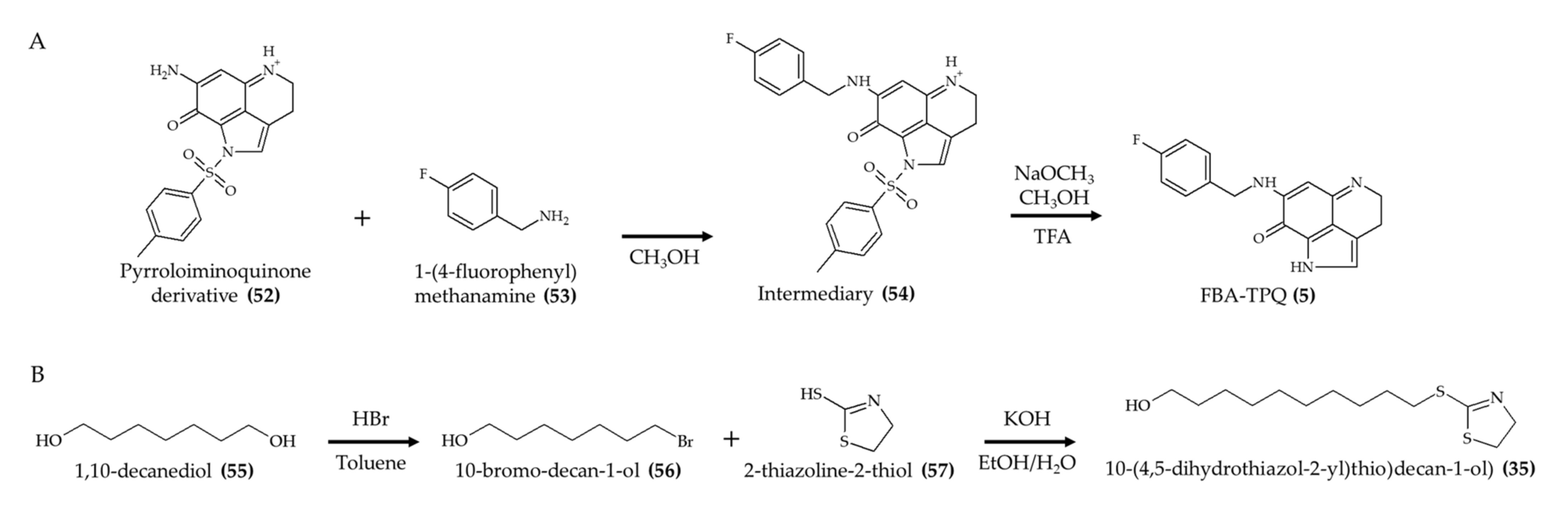
| Synthetic FBA-TPQ (5); makaluvamine analog from marine sponge Zyzzya sp. | 4–6-week-old female athymic nude mice; intraperitoneal injection; 5 mg/kg/day, 3 days/week for 3 weeks; 10 mg/kg, 3 days/week for 2 weeks; and 20 mg/kg, 3 days/week for 1 week | Reduced tumor growth; weight loss with doses of 10 and 20 mg/kg | [14] |
| Synthetic FBA-TPQ (5); makaluvamine analog from marine sponge Zyzzya sp. | 5-week-old female athymic nude mice; intraperitoneal injection; 1 and 10 mg/kg/day; 5 days/week for two and half weeks | Reduced tumor growth and no significant effect on body weight | [15] |
| Synthetic FBA-TPQ (5); makaluvamine analog from marine sponge Zyzzya sp. | 4–6-week-old female athymic nude mice; intraperitoneal injection; 5 and 10 mg/kg/day; 5 days per week for 3 weeks | Reduced tumor growth and induced remission | [16] |
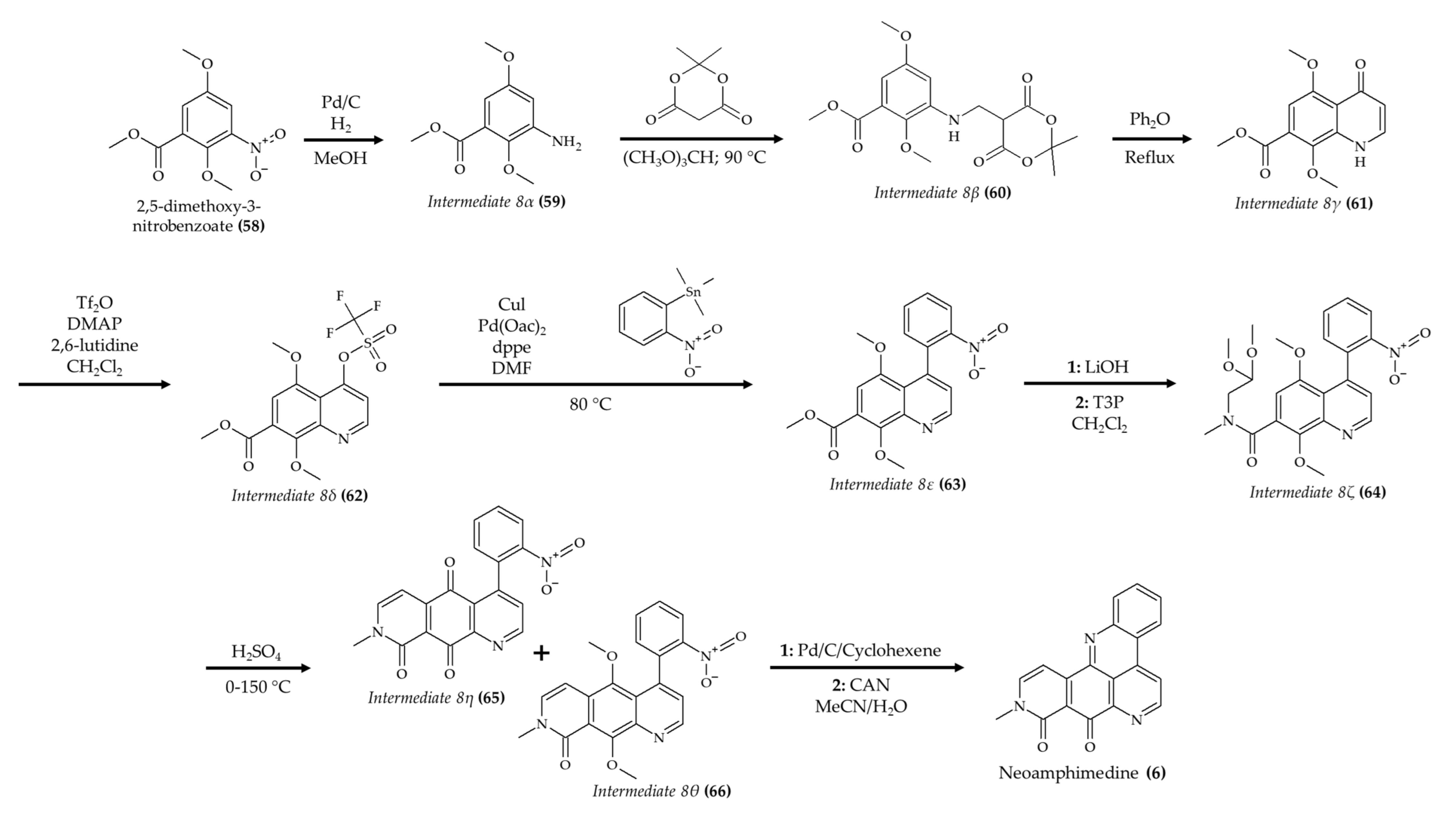
| Neoamphimedine (6) from marine sponge Xestospongia sp. | Nude mice; intraperitoneal injection; 50 mg/kg; 4 q.d. | Reduced tumor growth | [17] |
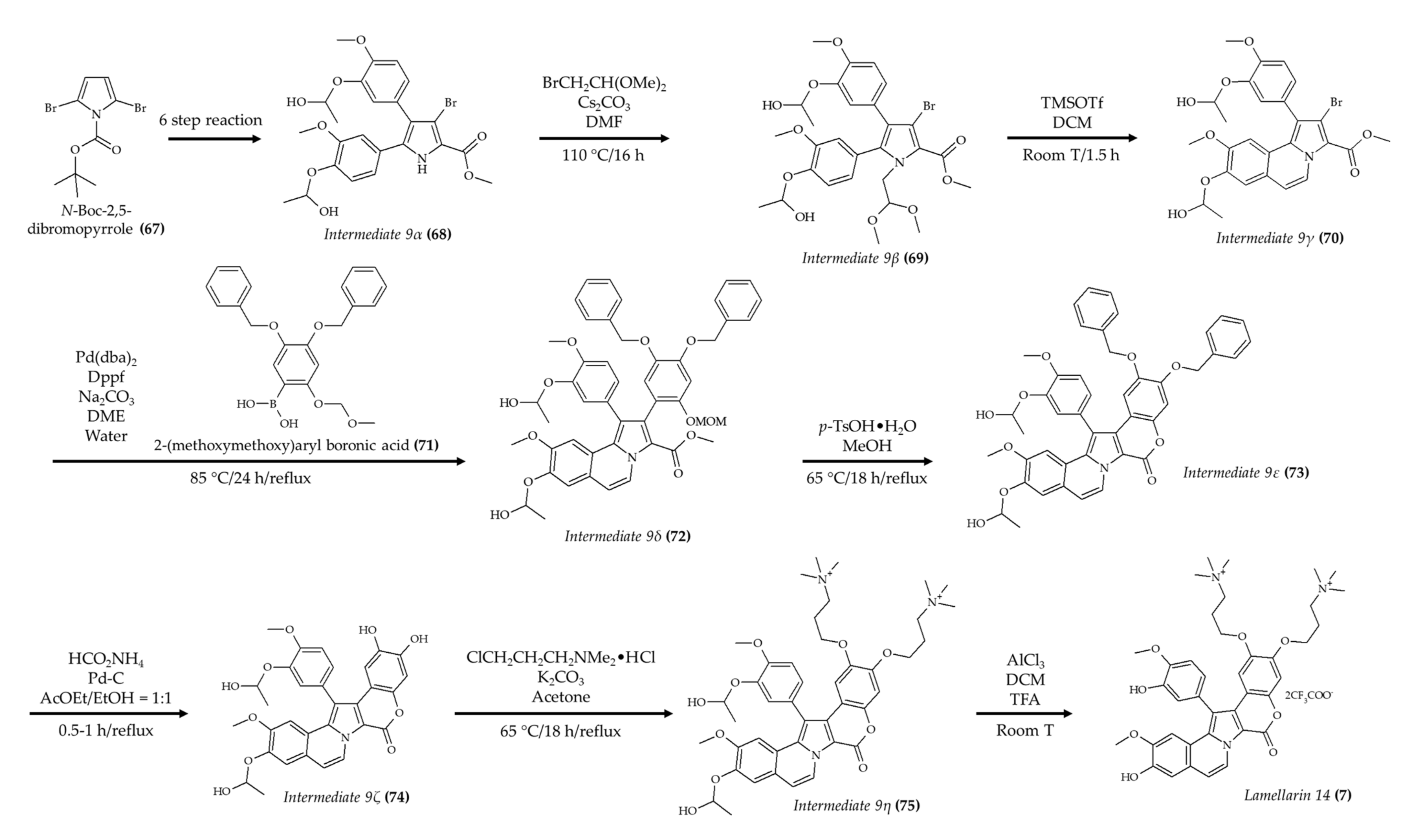
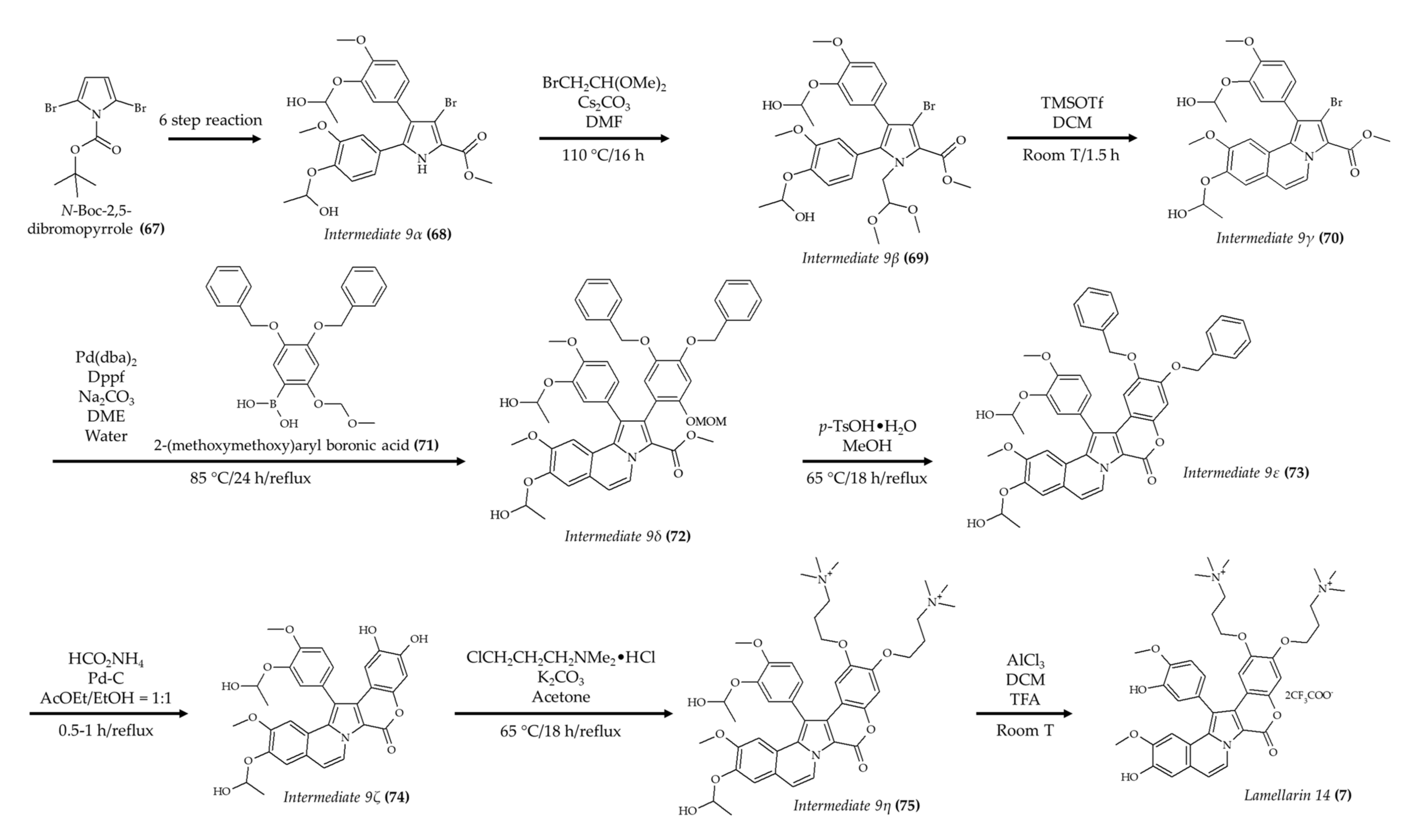
| Synthetic lamellarin 14 (7); lamellarin analog from mollusk Lamellaria sp | BALB/c nu/nu mice; 5 and 10 mg/kg; once a day for 17 days | Reduced tumor growth with no effect on body weight | [18] |
| Isofistularin-3 (8) from marine sponge Aplysina aerophoba | Zebrafish embryos; 15, 20, and 25 μM; incubation for 24 h | Reduced neuroblastoma (15–25 μM) and prostate (20 and 25 μM) cancer development | [19] |
| Crambescidine-816 (9) from marine sponge Crambe crambe | Zebrafish embryos; 0.5, 1, and 2 μM; 48 h after fertilization | Reduced tumor development; no effect on survival rate | [20] |
| Cardioprotective activity | |||
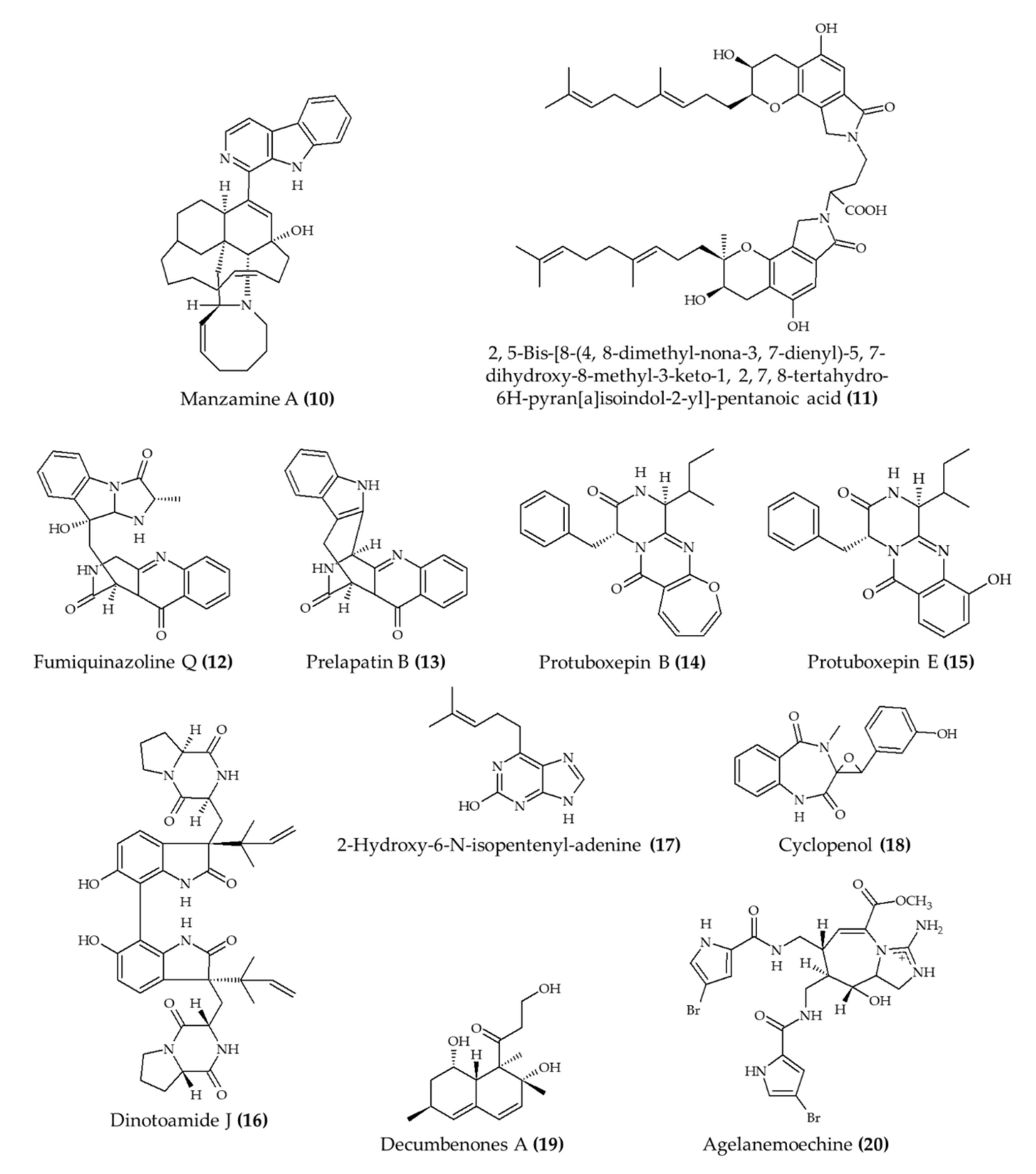
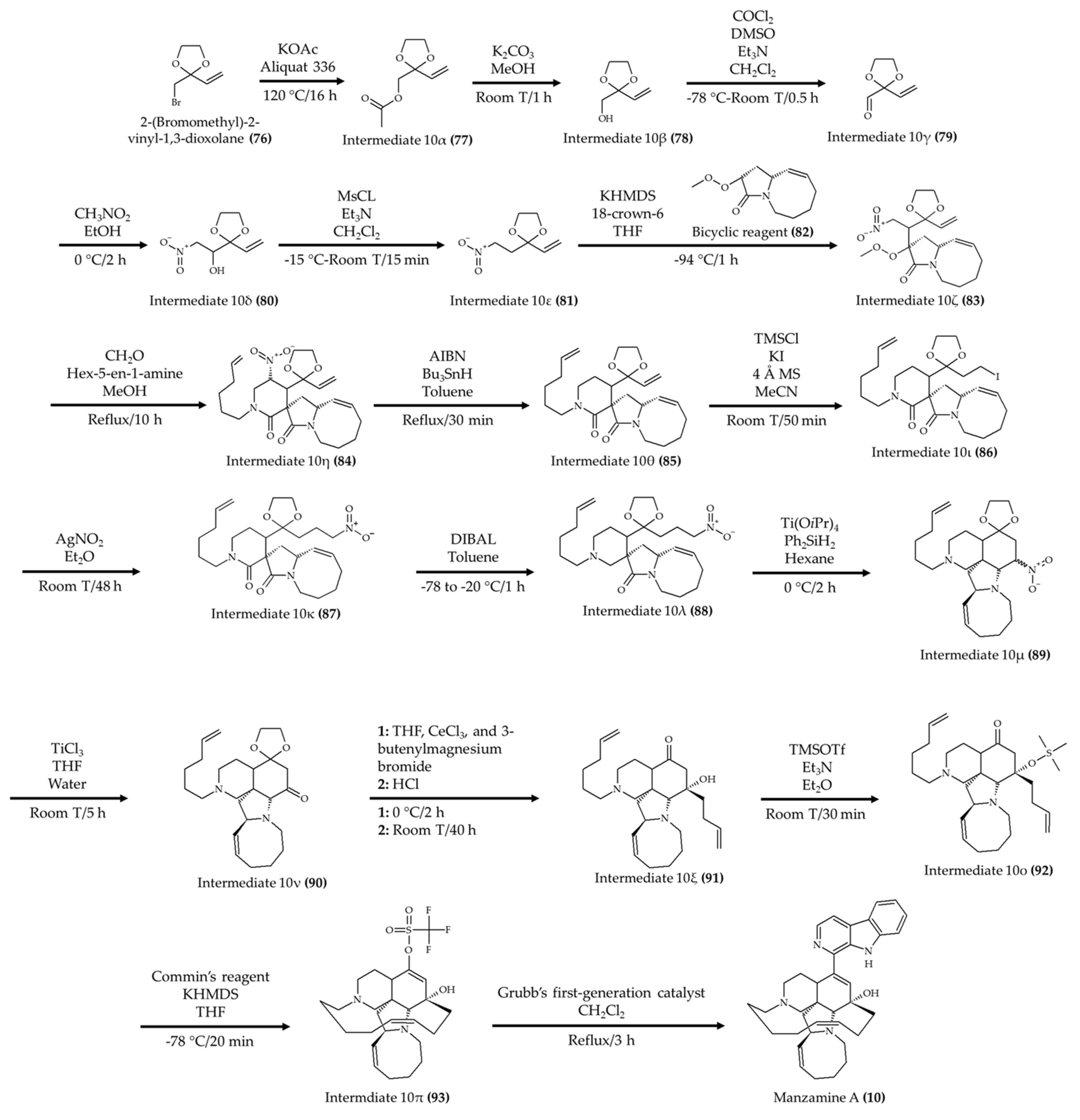
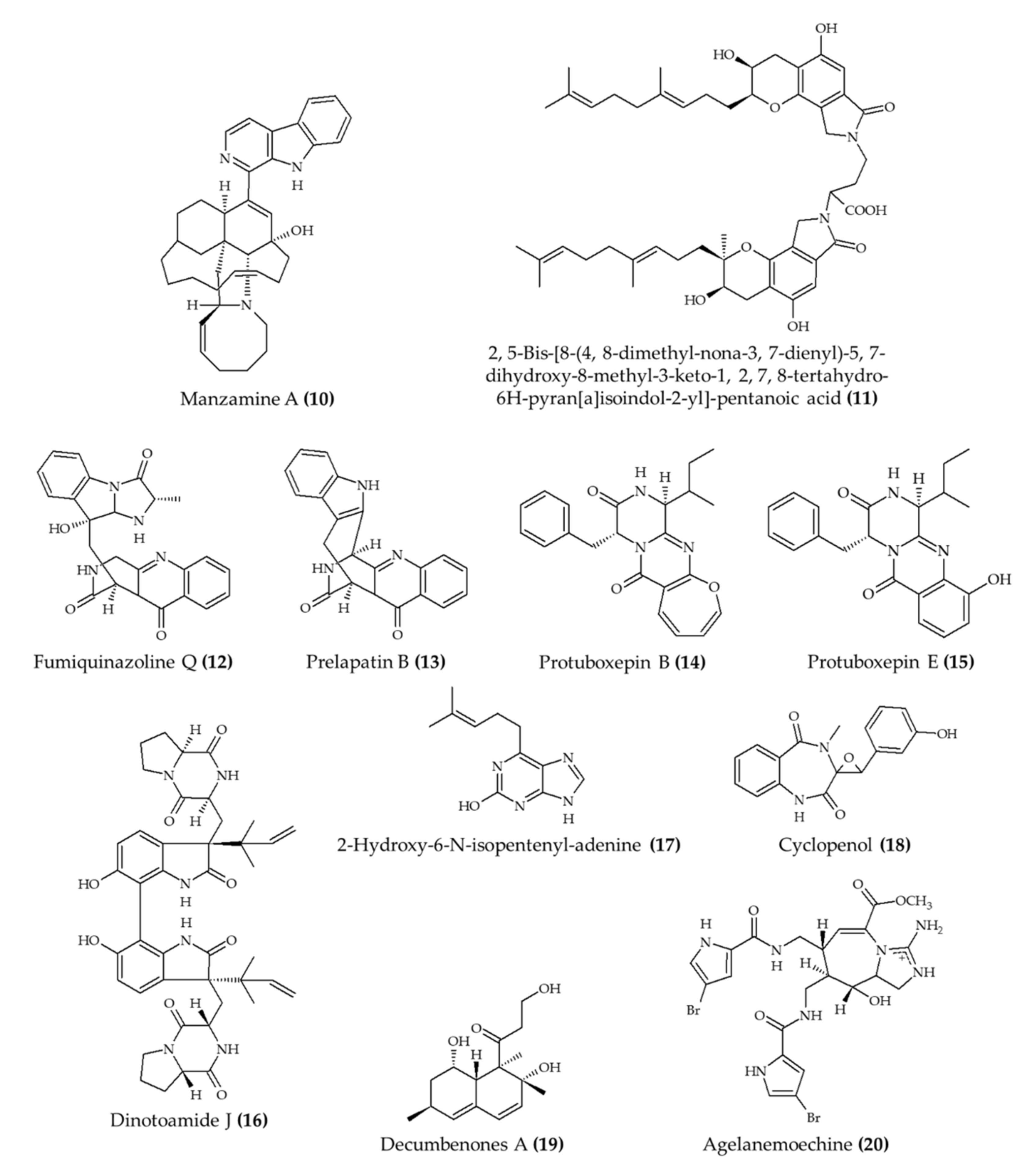
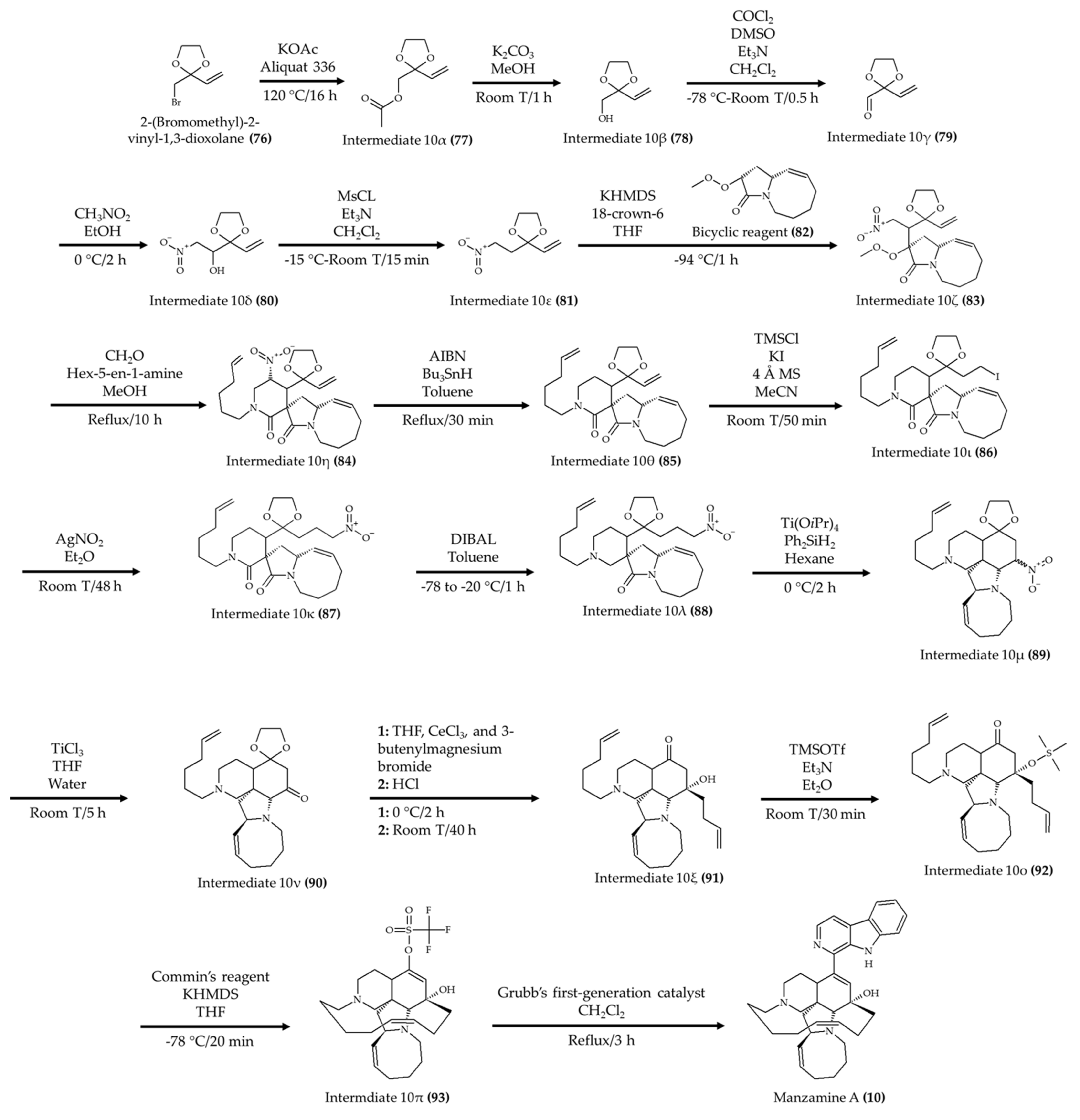
| Manzamine A (10) from marine sponge Acanthostrongylophora ingens | 6-week-old apoE-deficient mice; oral administration; 30 mg/kg for 80 days | Reduced total, free, and LDL cholesterol and triglyceride levels; atherosclerotic lesions were diminished | [21] |
| Fungi fibrinolytic compound 1 (11) from marine fungus Stachbotrys longispora FG216 | Wistar rats; 5, 10, and 25 mg/kg (morphological effect in lungs; euglobulin lysis time); injection in caudal vein | Reduced morphological changes from induced thrombosis and reduced euglobulin lysis time; no effect on fibrinogen degradation concentrations (5–25 mg/kg) | [22] |
| 4 alkaloids (12–15) from marine fungus Penicillium expansum | Zebrafish embryos; 1, 10, and 100 μg/mL; incubation for 24 h | Reduced bradycardia; induced angiogenesis | [23] |
| 4 alkaloids (16–19) from marine fungus Aspergillus austroafricanus | Zebrafish embryos; 30, 70, 120 µg/mL; incubation for 24 h | Induced angiogenesis | [24] |
| Agelanemoechine (20) from marine sponge Agelas nemoechinata | Zebrafish embryos; 1.25, 2.5, 5, 10, and 20 μM | Induced angiogenesis | [25] |
| Compound and Source | Experimental Conditions | Effect | Ref. |
|---|---|---|---|
| Antiseizure activity, Alzheimer’s disease, and mental health | |||
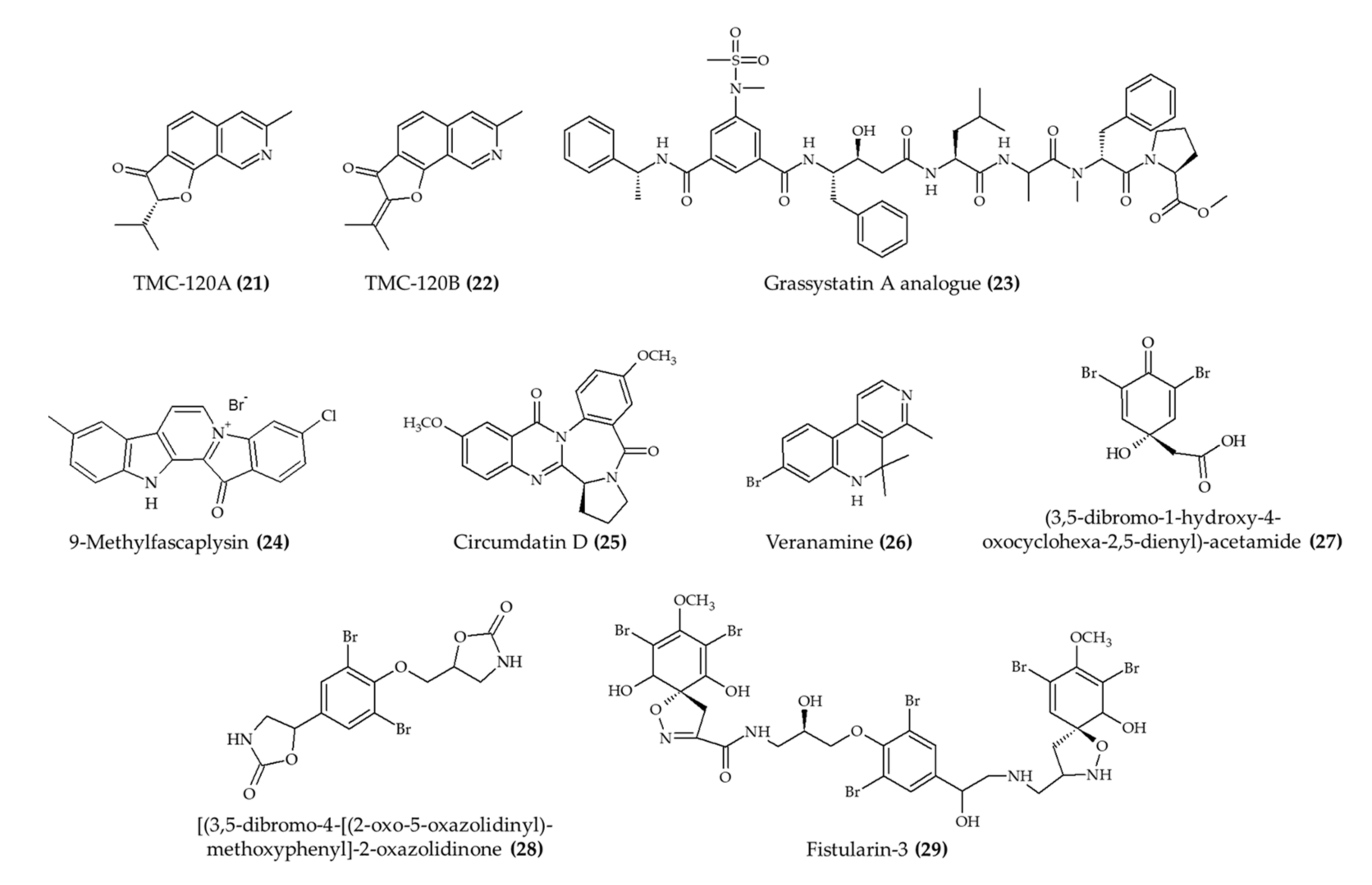
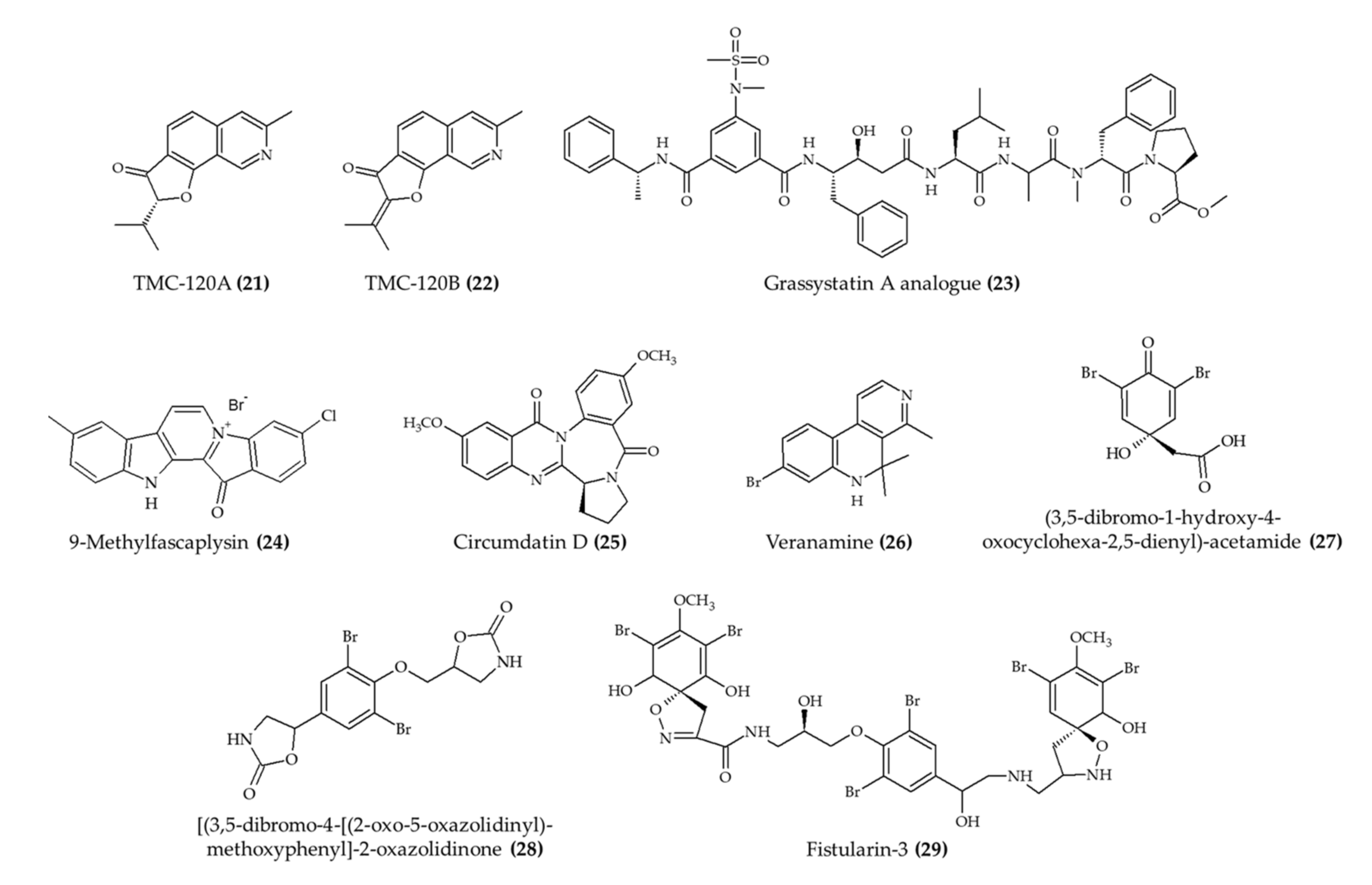
| TMC-120A (21) and TMC-120B (22) isoquinoline alkaloids from marine fungus Aspergillus insuetus | Male NMRI mice; intraperitoneal injection; TMC-120A (1.25, 2.5, 5, and 10 mg/kg) and TMC-120B (2.5, 5, 10, and 20 mg/kg); 30 min before electrical stimulation | Reduced seizure duration (at 10 mg/kg for both alkaloids) | [26] |
| TMC-120A (21) and TMC-120B (22) isoquinoline alkaloids from marine fungus Aspergillus insuetus | Zebrafish larvae; 5, 10, and 20 μg/mL; 2 h incubation | Reduced the proportion of animals and seizure duration (20 μg/mL) | [26] |
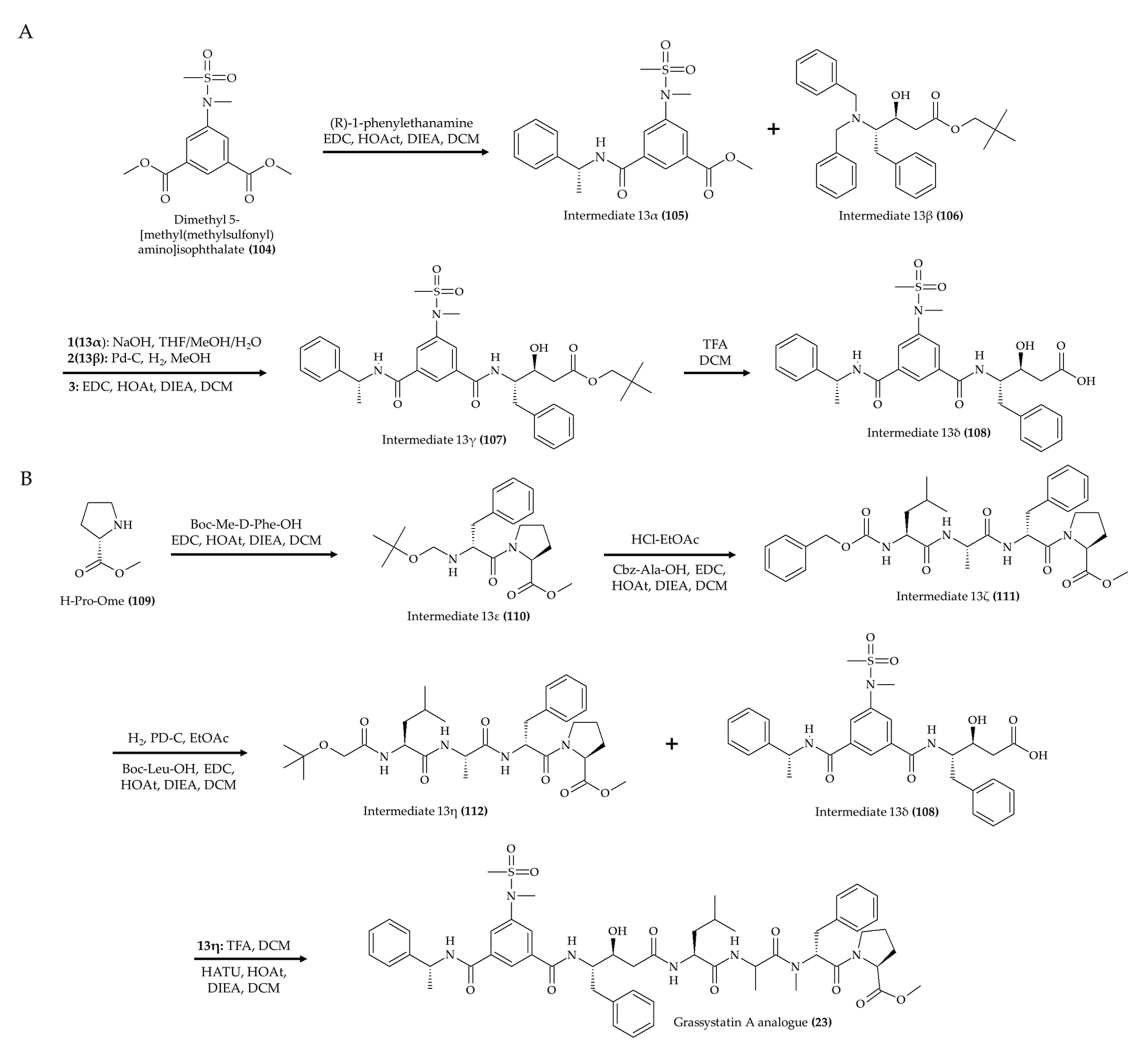
| 1 alkaloid (23); grassystatin A analog from marine cyanobacterium Symploca sp. | 8-week-old CF-1 mice; intraperitoneal injection; 30 mg/kg; single day | Reduced the Aβ40 level in the brain | [27] |
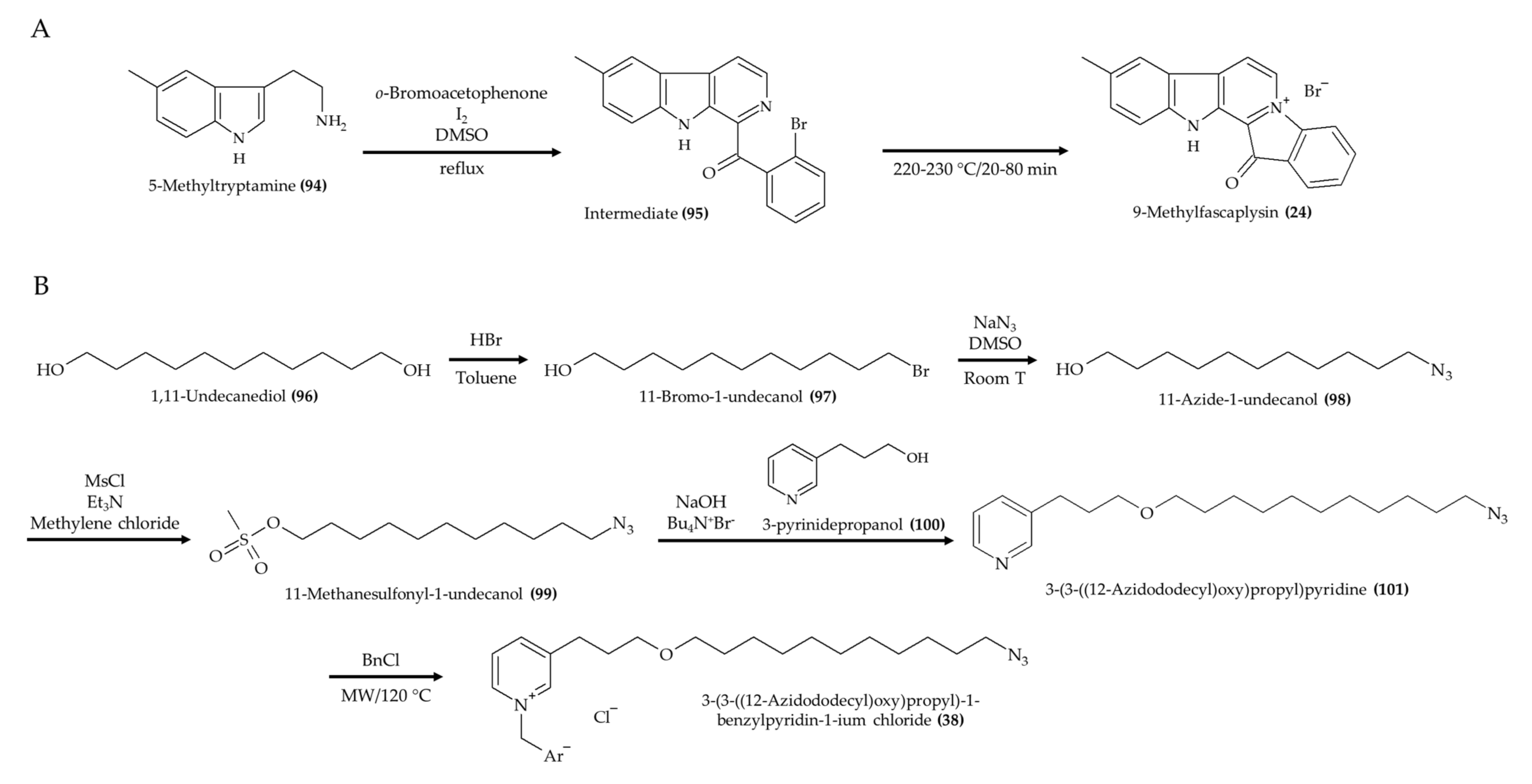
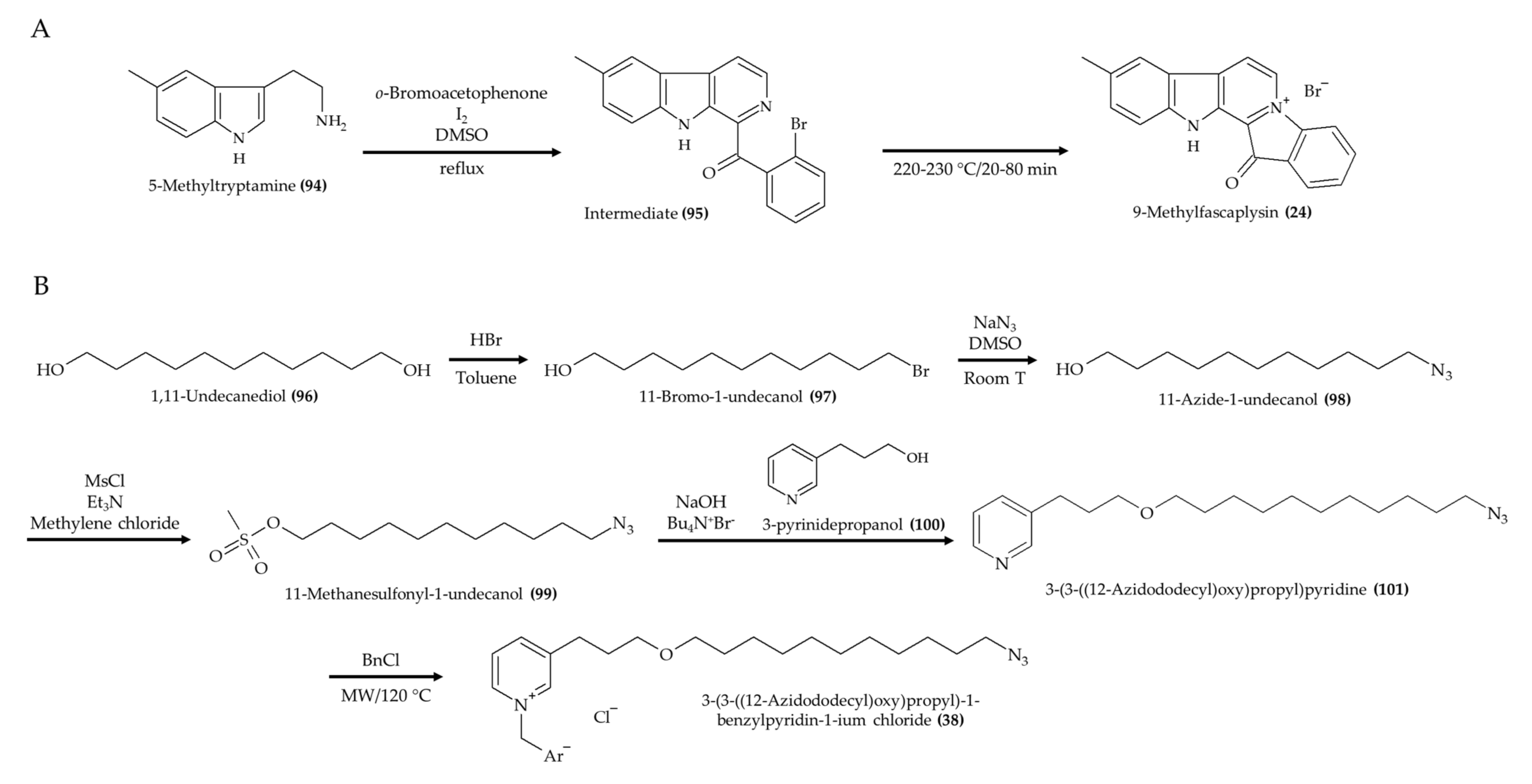
| 9-Methylfascaplysin (24); Fascaplysin analog from marine sponge Fascaplysinopsis sp. | 4-month-old male ICR mice; intrahippocampal injection; scopolamine with 7.3 ng and 21.9 ng; once a day for 10 days | Ameliorated cognitive dysfunction; inhibited Aβ-induced tau hyperphosphorylation; no effect in locomotor function | [28] |
| Circumdatin D (25) from marine fungus Aspergillus ochraceus | Caenorhabditis elegans; 50, 100, and 200 mM incubation for 36 h | Reduced paralysis rate | [29] |
| Veranamine (26) from marine sponge Verongula rigida | Male Swiss Webster mice; intraperitoneal injection; 20 mg/kg; single day | Reduced immobility time; no effect on locomotor activity | [30] |
| 3 alkaloids (27–29) from marine sponge Aplysina fulva | Zebrafish adult; 0.1, 0.5, and 1.0 mg/mL; 1 h incubation | Induced anxiolytic effect and involved the GABAergic system | [31] |
| Anti-inflammatory | |||
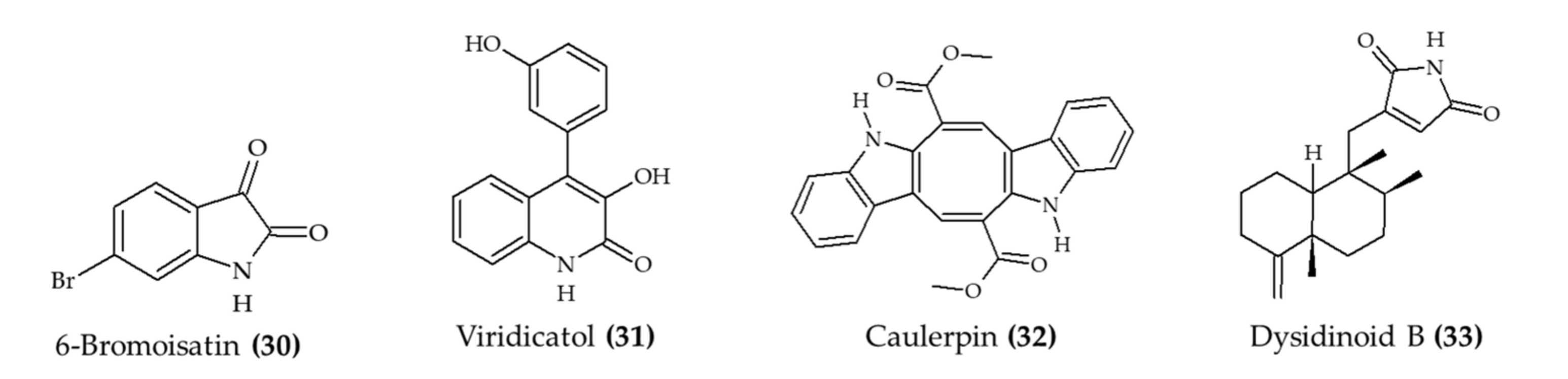
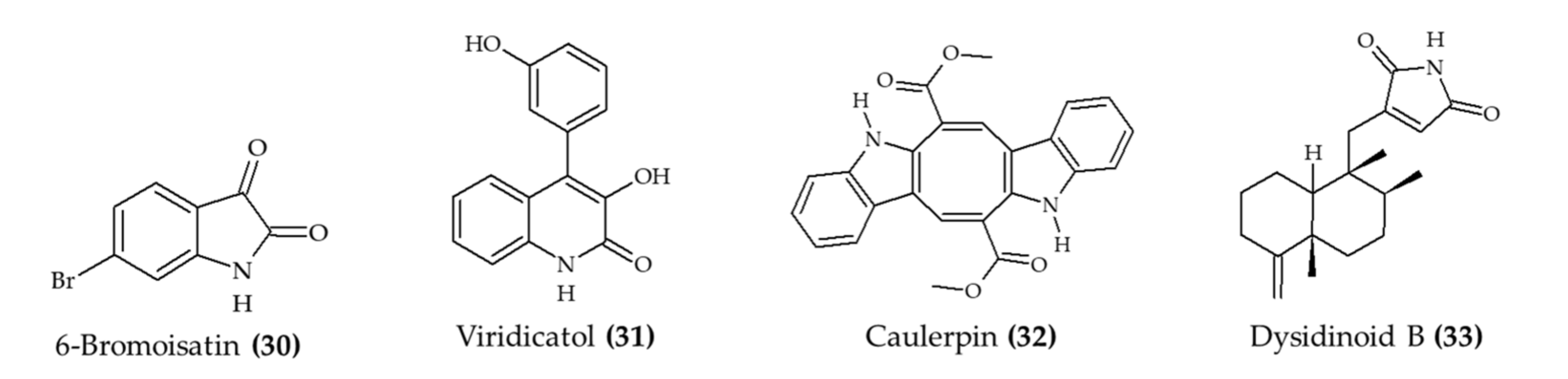
| 6-Bromoisatin (30) isolated from Muricidae mollusk Dicathais orbita | Male and female C57 black/6 mice; oral administration; 0.5 and 0.1 mg/g HBG extract and 0.05 and 0.1 mg/g 6-bromoisatin (30); 48, 24, and 1 h prior to the administration of LPS; mice euthanized after 3 h | Reduce acute lung inflammation, TNFα, IL-1β, and total protein levels in BALF, attenuated physiological changes | [32] |
| Viridicatol (31) isolated from marine fungus Penicillium griseofulvum | 6–7-week-old female BALB/c mice; oral administration; 5, 10, and 20 mg/kg for 13 days | Reduced OVA-specific IgE, serum histamine, mMCP-1, and TNF-α; increased IL10 level | [33] |
| Caulerpin (32) from seaweed Caulerpa racemosa | 6–8-week-old male C57BL/6 mice; oral administration; 0.4, 4, and 40 mg/kg for 7 days | Reduced colon damage and shortening and DAI (4 mg/kg) | [34] |
| Dysidinoid B (33) from marine sponge Dysidea septosa | Zebrafish embryos; 20, 40, and 80 μM; incubation for 2 h | Reduced inflammation (40 and 80 μM) | [35] |
| 3 alkaloids (18, 19, and 31) from marine fungus Aspergillus austroafricanus | Zebrafish embryos; 30, 70, and 120 µg/mL; incubation for 2 h | Induced anti-inflammatory response | [24] |
| Compound and Source | Experimental Conditions | Effect | Ref. |
|---|---|---|---|
| Antiparasitic activity | |||
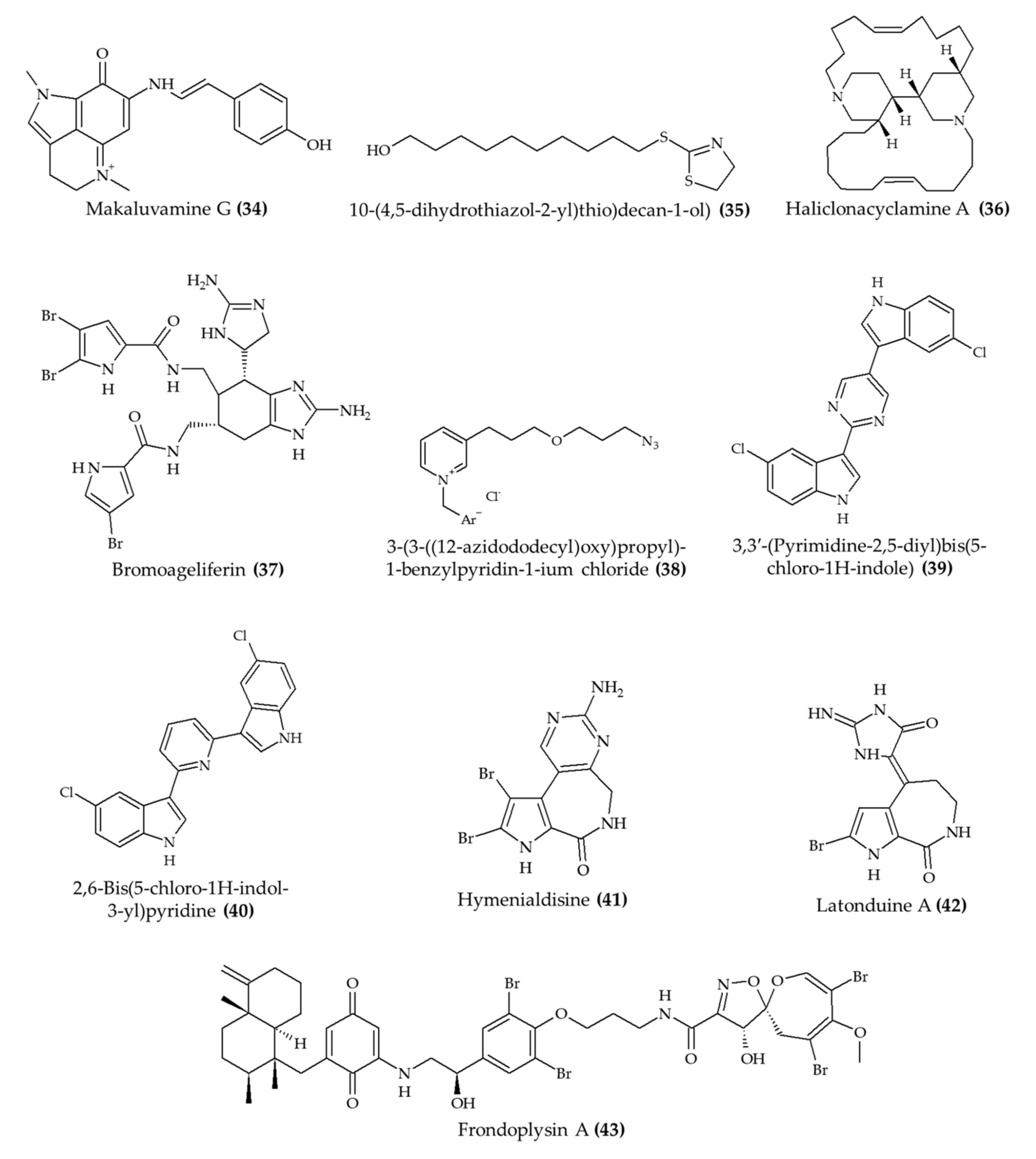
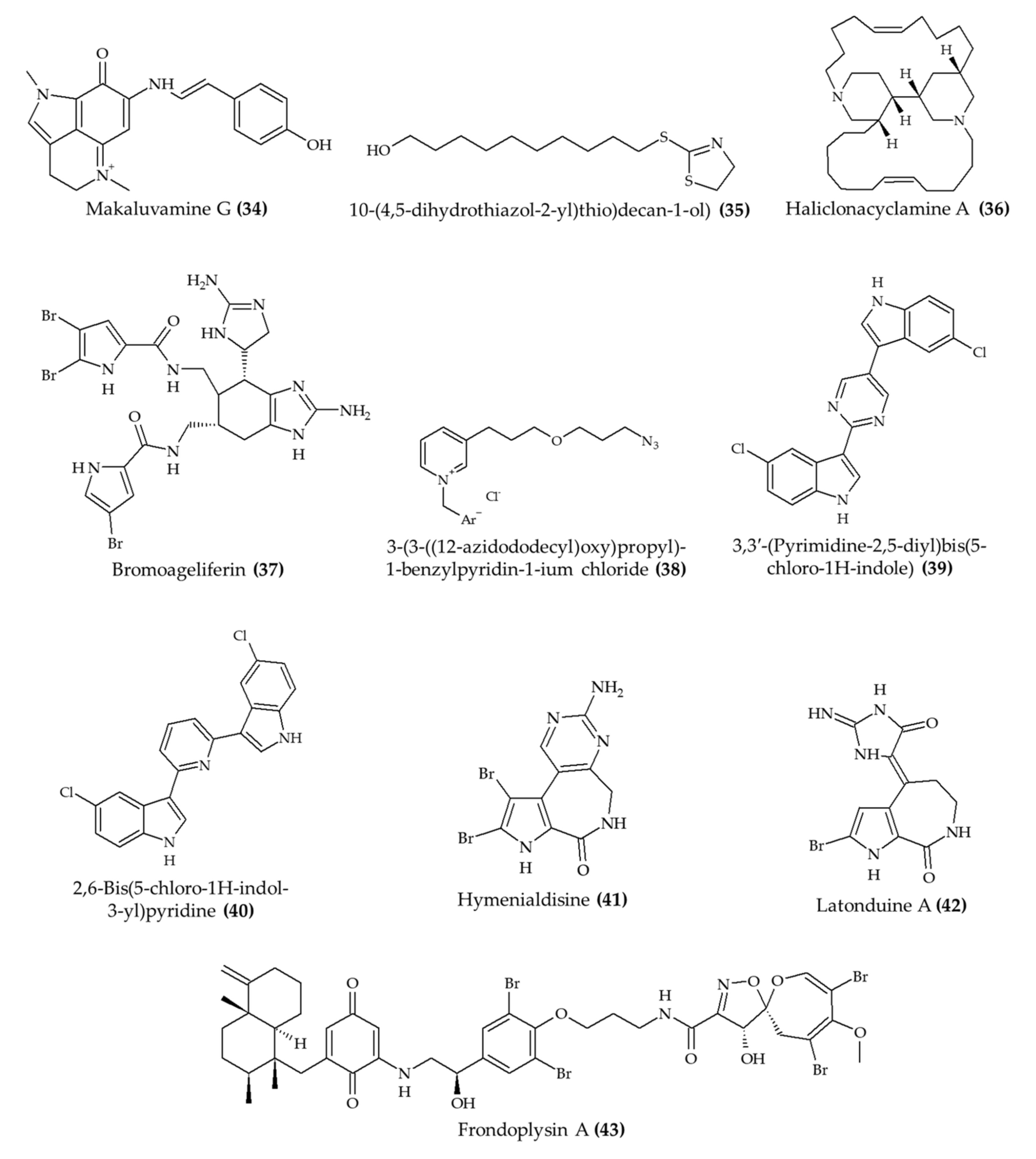
| Makaluvamine G (34) from marine sponge Zyzzya sp. | 7–9-week-old male and female Swiss albino mice; subcutaneous injection; 8 mg/kg/day for 4 days | Reduced the growth of Plasmodium berghei in infected mice | [36] |
| 10-(4,5-Dihydrothiazol-2-yl)thio)decan-1-ol) (thiazoline) (35), a synthetic analog of 3-alkylpiridine | 6–8-week-old female C57BL/6 mice; oral administration; 25 and 50 mg/kg (nanoemulsion) and 25 mg/kg (free compound); 4 h after infection for 4 days; samples collected after 5, 8, and 10 days | Reduced parasitemia for 8 days (especially with 50 mg/kg) | [37] |
| Haliclonacyclamine A (36) from marine sponge Haliclona sp. | Swiss female mice; 0.1, 1, and 10 mg/kg; once a day for 4 days | Reduced parasitemia by 45% after 4 days of treatment (10 mg/kg) | [38] |
| Antimicrobial activity | |||
| Bromoageliferin (37) from marine sponge Agelas dilatata | Galleria mellonella larvae; 2 mg/kg | Increased survival rate after infection with Pseudomonas aeruginosa | [39] |
| 3-(3-((12-Azidododecyl)oxy)propyl)-1-benzylpyridin-1-ium chloride (38), a synthetic 3-alkylpiridine analog | 6–8-week-old Swiss male mice; 0.5 and 1.0 mg/kg; 2, 24, and 48 h postinfection | Reduced microbial infection in kidney and spleen (1.0 mg/kg); no effect in liver | [40] |
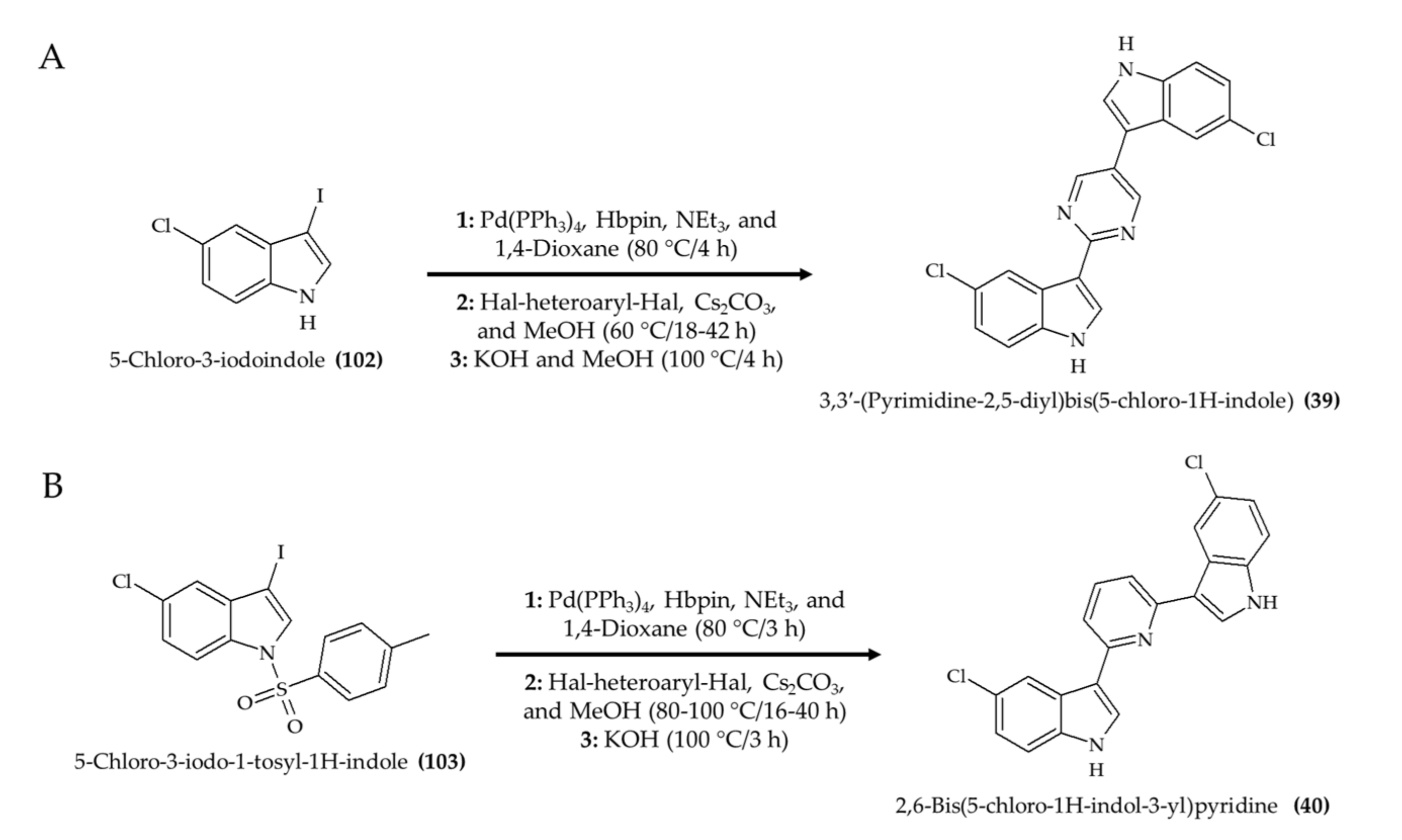
| 2 alkaloids (39 and 40), hyrtinadine A analogs from Red Sea sponge Hyrtios sp. | BALB/c female mice; topical administration and intraperitoneal injection; 5 mg/kg; single application | Improved wound skin wound healing; increased survival rate | [41] |
| Osteoporosis | |||
| Hymenialdisine (41) from marine sponge | 11-week-old female C57BL/6j mice; intraperitoneal injection; 1 mg/kg; every 2 days for 6 weeks | Reduced the loss of bone volume and trabecular thickness | [42] |
| Cystic fibrosis | |||
| Latonduine A (42) from marine sponge Stylissa carteri | 10–12-week-old F508del-CFTR homozygous mice; gavage; 50 mg/kg once daily for 2 days | Reduced salivary secretion | [43] |
| Antioxidant activity | |||
| Frondoplysin A (43) from marine sponge Dysidea frondosa | Zebrafish embryos; 20 μM | Induced antioxidant response | [44] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Munekata, P.E.S.; Pateiro, M.; Conte-Junior, C.A.; Domínguez, R.; Nawaz, A.; Walayat, N.; Movilla Fierro, E.; Lorenzo, J.M. Marine Alkaloids: Compounds with In Vivo Activity and Chemical Synthesis. Mar. Drugs 2021, 19, 374. https://doi.org/10.3390/md19070374
Munekata PES, Pateiro M, Conte-Junior CA, Domínguez R, Nawaz A, Walayat N, Movilla Fierro E, Lorenzo JM. Marine Alkaloids: Compounds with In Vivo Activity and Chemical Synthesis. Marine Drugs. 2021; 19(7):374. https://doi.org/10.3390/md19070374
Chicago/Turabian StyleMunekata, Paulo E. S., Mirian Pateiro, Carlos A. Conte-Junior, Rubén Domínguez, Asad Nawaz, Noman Walayat, Elena Movilla Fierro, and José M. Lorenzo. 2021. "Marine Alkaloids: Compounds with In Vivo Activity and Chemical Synthesis" Marine Drugs 19, no. 7: 374. https://doi.org/10.3390/md19070374
APA StyleMunekata, P. E. S., Pateiro, M., Conte-Junior, C. A., Domínguez, R., Nawaz, A., Walayat, N., Movilla Fierro, E., & Lorenzo, J. M. (2021). Marine Alkaloids: Compounds with In Vivo Activity and Chemical Synthesis. Marine Drugs, 19(7), 374. https://doi.org/10.3390/md19070374