
Bridging Cyanobacteria to Neurodegenerative Diseases: A New Potential Source of Bioactive Compounds against Alzheimer’s Disease


 , , ,
, , ,  and
and 
Abstract
1. General Introduction
1.1. Neurodegenerative Diseases and Natural Compounds
1.2. Cyanobacteria
1.3. Objectives of the Review
2. Alzheimer Disease
2.1. Alzheimer Disease Hypothesis and Main Therapeutical Targets
2.1.1. Amyloid Cascade Hypothesis (ACH)
2.1.2. Cholinergic Hypothesis (CH)
2.1.3. Glutaminergic Hypothesis (GH)
2.1.4. Tau Hypothesis (TH)
2.1.5. Inflammation and Oxidative Stress Hypothesis
2.2. Current Therapeutics
3. Cyanobacteria Potential in Alzheimer Disease

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genera/Specie | Compound/Extract | Mechanism/Effect | In Vitro Assays | In Vivo Assays | Reference |
|---|---|---|---|---|---|
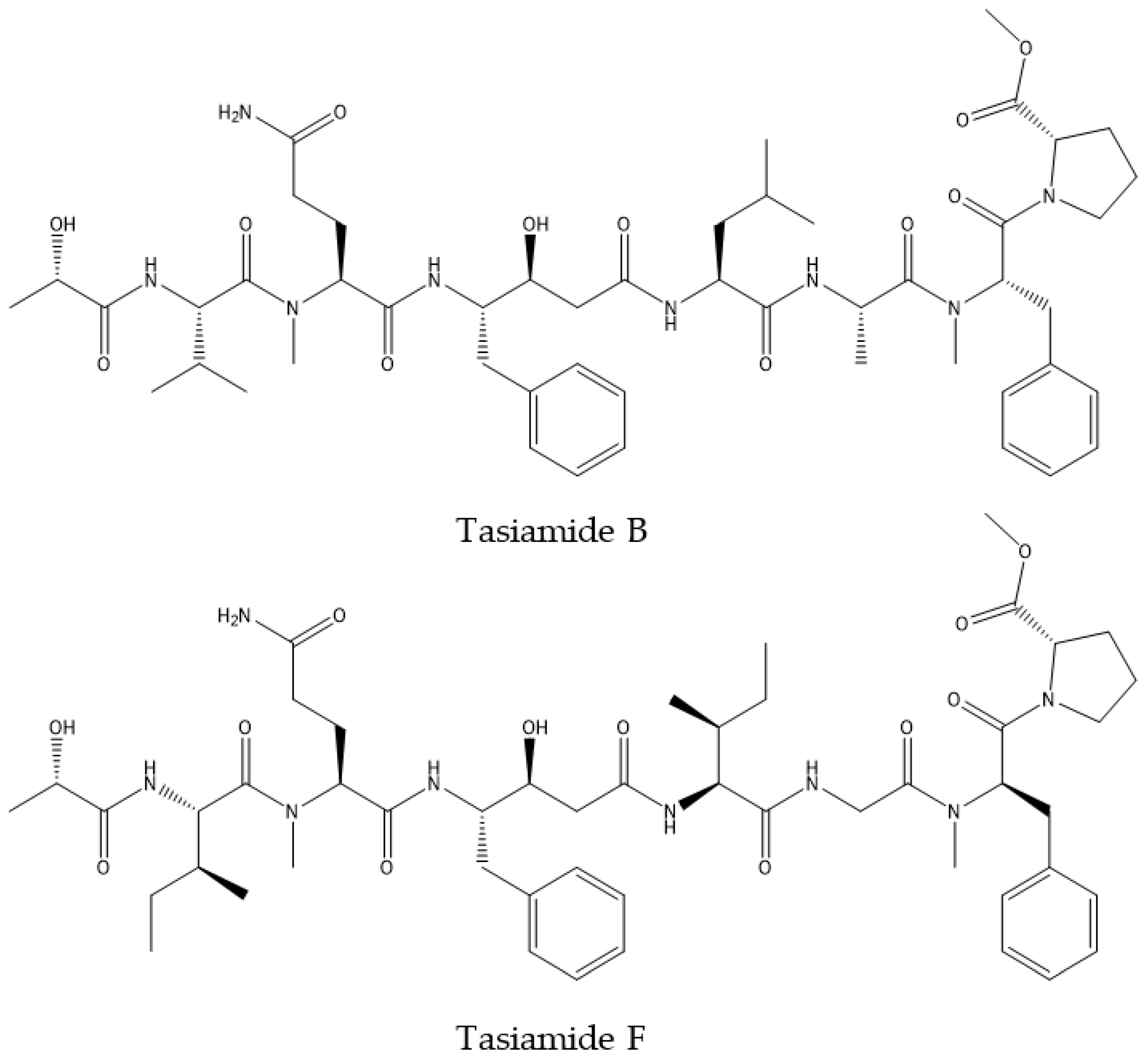
| Symploca sp. | Tasiamide B | BACE1 inhibition | BACE1 inhibition assay H4 cells: HPLAP-APP Reporter Assay; CTF analysis; BBB PAMPA; CHO 2B7 cells: Secreted Aβ Assay. | CF-1 Mice: plasma and brain Aβ levels by anti-Aβ antibody; plasma and brain compounds stability monitoring by HPLC | [98,99] |
| Leptolyngbya sp. | Phycocyanin | BACE1 inhibition | In silico assay of molecular docking | Transgenic Caenorhabditis elegans AD-model | [106] |
| Phycoerythrin | BACE1 inhibition | Thermodynamics of binding using surface plasmon resonance (SPR); isothermal titration calorimetry (ITC); enzyme activity by kinetic parameters. | Caenorhabditis elegans CL4176 transgenic AD model worm: Aβ reduction by Thioflavin-T staining assay | [107] | |
| Calothrix sp. | Methanolic extract | AChE inhibition | AChE inhibitory assay | [103] | |
| Tolypothrix sp. | |||||
| Phormidium cf. amoenum | |||||
| Phormidium sp. | |||||
| Geitlerinema splendidum | |||||
| Spirulina sp. | C-phycocyanin (CPC) | Inhibition of Aβ40/42 fibril formation | EM imaging | [108] | |
| Antioxidant; anti-inflammatory | Wistar rats: bioavailability of C-PC in cortical tissue homogenates; DCFH-DA for ROS levels; Cayman’s protein carbonyl assay; TBARS for lipid peroxidation damage; GPx, GR; GST; SOD; CAT assays; Caspase-12 activation assay; Calpain activation assay; Western Blot for Cox-2-, Nk-kB, IL-6, GAPH, GFAP, NF, MBP, IBA1, CD11b, Nrf2, MT, PGP, Occludin, Claudin, ZO-1, Connexin43, b-actin; Immunohistochemistry for GFAP and DAPI. TUNEL staining | [109] | |||
| Spirulina sp. | PUFAs: | Reversion of age-related impairments in LTP; spatial learning and depolarization-induced glutamate release; decrease in age-related microglial activation and associated oxidative stress; inhibition of the Aβ-induced LTP; inhibition by EPA | Glia cells: ELISA for IL-1β and IFNϒ analysis; IL-1β mRNA analysis on agarose gel | Male Wistar rats: assessment of glutamate release by synaptosomal tissue optical density; ROS quantification of hippocampal homogenate (fluorescence); LTP induction and measurement of excitatory postsynaptic potential (EPSP); Young and aged Wistar rats: morris water maze; analysis of LTP; sphyngomielinase; ELISA for 8-hydroxy-2′ deoxyguanosine; immunohistochemical analysis; analysis of fatty acids; Young, middle-aged and aged male Wistar rats: induction of LTPD in performant path-granule cells synapse with electrode; SDS-PAGE for expression of RAGE, CD40, pJNK and PPARϒ on hippocampal homogenate | [118,119,120,121] |
| Spirulina sp. | Water extract | Antioxidant | ABTS assay DPPH assay | [122] | |
| Spirulina platensis | C-phycocyanin | Antioxidant | DPPH assay | [123] | |
| Water extract (SP) | Improve memory function, prevention of Aβ accumulation, reduction of oxidative stress, enhanced catalase activity. | SAMP8 mice: shuttle box: single trial passive avoidance test. Active (shuttle) avoidance test; ambulatory activity in cubic boxes; measurement of Aβ deposition: Immunostaining of sections of the brain with anti-Aβ antibody; redox status: hippocampus, striatum and cortex homogenates separately; lipid peroxidase, SOD, CAT, and GSH-Px activity assays; lipid peroxidation levels by spectrophotometry | [110] | ||
| Oral administration | Reduction of KA-neuronal death in C3 hippocampal cells; antioxidant | Male SW mice treated with kainic acid (KA): determination of the atrophic and nucleolated pyramidal neurons number and volume of observed in the hippocampal region | [111] | ||
| Spirulina maxima | Methanolic extract | Antioxidant | Brain homogenate incubated with and without extract antioxidant activity by inhibition of peroxidation | Wistar rats: antioxidant activity by lipid peroxidation of liver, plasma, and brain homogenate | [41,114] |
| 70% ethanolic extract | Suppression of the Aβ-induced toxicity in PC12 cells by decrease oxidative stress, cell death; increase the brain-derived neuro- trophic factor (BDNF) and decrease BACE1 | MTT assay; LDH assay; intracellular glutathione; western blot | [115] | ||
| Polyphenolic extracts | Antioxidant | DPPH assay | Male Wistar rats: lipid peroxidation in liver homogenate | [41,116] | |
| Phycocyanobilin | Neuroprotection by inhibition of NADPH oxidase | [117] | |||
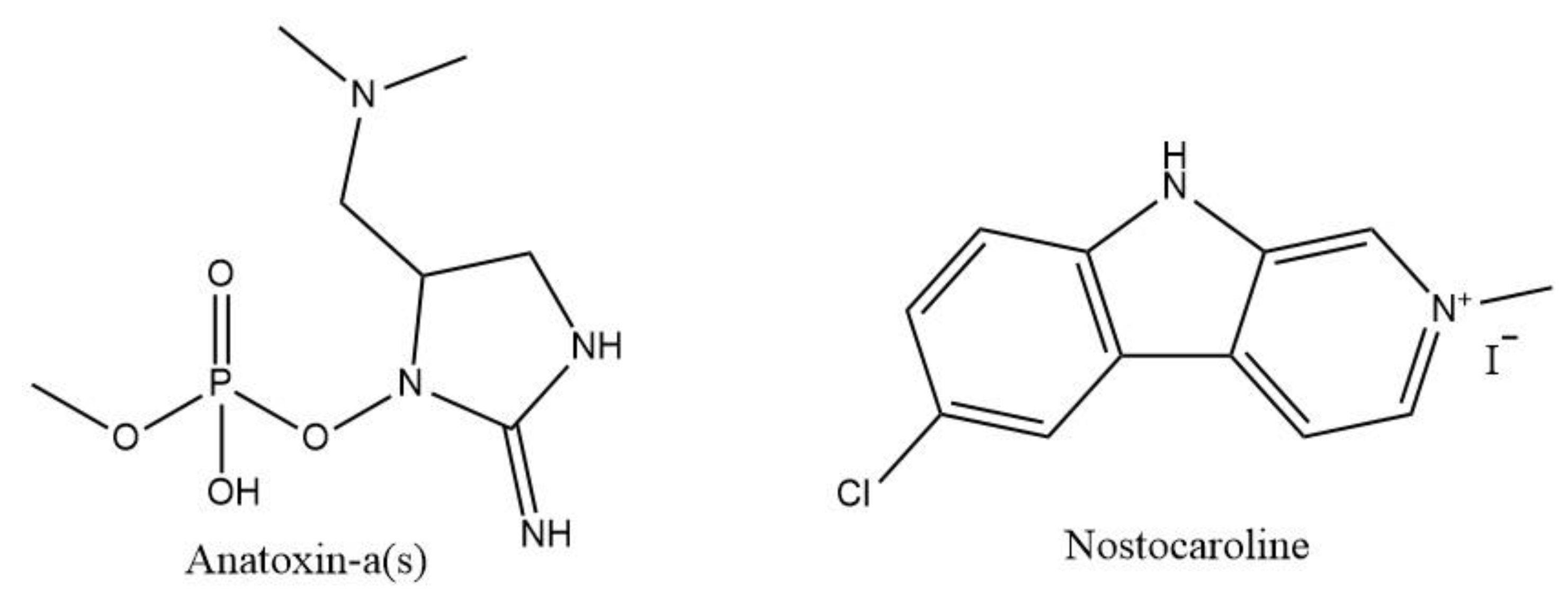
| Nostoc 78-12A | Nostocarboline | BChE inhibition | AChE inhibitory assay | [104] | |
| Nostoc ellipsosporum | Hexane, ethyl acetate and water extract | Antioxidant | Trolox equivalent (TEAC) assay by ABTS radical decolorisation method; Folin Ciocalteu method for total phenolic content | [105] | |
| Synechococcus sp. | |||||
| Lyngbya sp. | Ethanolic fraction | Antioxidant; advanced glycation end-products (AGEs) inhibition | DPPH assay; phosphomolybdenum assay; BSA glycation inhibition assay; nitric oxide inhibition assay for anti-inflammatory activity | C. elegans (N2, Bristol) glucose-induced hyperglycemia: HCS analysis for AGE accumulation in live animals; quantitative analysis of AGE accumulation by spectrofluorimetry; DNSA method for glucose analysis; semi-quantitative RT-PCR analysis for stress responses genes (glod-4, daf-16, daf-2.) C. elegans TJ356: daf16: GFP:HCS of the localization of daf-16 tagged GFP | [124] |
| Tasiamide F | BACE1 inhibition | Antiproteolytic activity | [100] | ||
| Lyngbya majuscula | Hydroalcoholic extract | Antioxidant and neuroprotective | PC12 cells: MTT assay; DPPH assay; Caspase 3 activity; DNA ladder assay; DAPI staining | [125] | |
| Kalkitoxin | Blocking agent to Voltage-gated Sodium channels (VGSC) | Cerebellar granule neurons: LDH activity assay Intracellular Ca2+ analysis whole cell binding assay | [126] | ||
| Anabaena-flos-aquae | Anatoxin-a(s) | AChE and BChE inibithion | AChE inhibitory assay | Sprague-Dawley male rats for blood cells: in vitro cholinesterase assays | [102] |
| Aquose extract (Klamin®) | Anti-inflammatory; protective role against Aβ aggregation | Oxygen Radical Absorvance Capacity (ORAC) assay Folin-ciocalteu Assay LAN5 cells: MTS assay; ROS Generation and Mitochondrial; transmembrane Potential modification through DCFH-DA assay and MitoProbe JC-1 assay kit and MitoSOX Red Reagent; LAN5 cells treated with Aβ oligomers: DCFH-DA assay; MTS assay; Immunostaining for NFkB and Hoescht 33258; ELISA for IL-6 and IL-1β; thioflavin T for Aβ kinetics studies and for the formation and mean size of Aβ aggregates | [41,127] | ||
| Inhibition of H2O2- induced cytotoxicity and ROS generation; neuroprotection towards Aβ oligomers and Aβ oligomer-induced oxidative stress | Oxygen radical absorbance; Folin-Ciocalteu assay for penolic contents; ABTS assay A549 cells: MTS assay in after no treatment and after H2O2 exposure; DCFH-DA assay after H2O2 exposure; LAN5 cells: MTS assay after Aβ treatment oligomers and observation of morphology; DCFH-DA assay after H2O2 or Aβ oligomers exposure | [128] | |||
| Phormidium sp. A09DM | Phycoerythrin | Antioxidant | Fibroblast (3T3-L1) cell line: MTT assay; DCFH-DA staining on H2O2 induced oxidative stress | Caenorhabditis elegans N2: DCFH-DA stain: C. elegans CL4176 transgenic model of AD: DCFH-DA stain; Heat-induced Paralysis assay. Drosophila melanogaster: climbing assay; SOD and CAT assays | [112] |
| Allophycocyanin | Improve lifespan, improve rate of survival against oxidative stress and thermal stress; moderate expression of human Aβ1-42 and associated Aβ-induced paralysis | C. elegans N2 Bristol (wild type): lifespan study; Stress tolerance assay (H2O2-induced oxidative stress and thermal stress by increasing temperature from 20 °C to 35 °C); DCHF DA staining; study of lifespan on knockdown skn-1 and daf-16 worms; C. elegans TJ356: DAF 16:GFP nuclear localization: evaluating the existence of GFP agglomeration in the nuclei after induce heat shock; C. elegans CL4176: paralysis assay after inducement of Aβ1-42 production; Aβ staining by thioflavin T. | [113] | ||
| Trifolium pratense | Biochanin A | Attenuation of the cytotoxic effect of the Aβ25–35 protein by decreasing viability loss, LDH release, and caspase activity in cells; reduction of cytochrome c and Puma; restoration of Bcl-2/Bax and Bcl-xL/Bax ratio preventing mitochondrial dysfunction | PC12 cells: MTT reduction assay after Aβ exposure; LDH activity; annexin V–FITC and PI; Hoechst 33342; caspase-8 caspase-9 and caspase-3 activity assay; rhodamine 123 fluorescent dye (Rh123); Western blot to Bcl-2, Bcl-xL, Bax, Puma and cytochrome c | [129,130] | |
| Microcystis, Anabaena | Microcystin-LR | Inhibition of Ser/Thr Protein Phosphatases (PPP) | Measurement of phosphatases activities | [131,132] | |
| Nodularia | Nodularin |
| Compound | Action | Reference |
|---|---|---|
| Anatoxin a(s) | AChE and BChE inhibition | [102] |
| Biochanin | Prevent mitochondria dysfunction | [130] |
| Kalkitoxin | Inhibition of voltage-gated sodium channels | [126] |
| Microcystis | Inhibition of Ser/Thr protein phosphatases | [131] |
| Nodularin | Inhibition of Ser/Thr protein phosphatases | [131] |
| Nostocarboline | BChE inhibition | [104] |
| Phycocyanin | BACE1 inhibition; antioxidant | [106] |
| Phycoerythrin | BACE1 inhibition; antioxidant | [107] |
| Tasiamide B/F | BACE1 inhibition | [98,100] |
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Gorman, A.M. Neuronal cell death in neurodegenerative diseases: Recurring themes around protein handling: Apoptosis Review Series. J. Cell. Mol. Med. 2008, 12, 2263–2280. [Google Scholar] [CrossRef]
- Park, K. “Genetic Changes” NIH Public Access. Bone 2014, 23, 1–7. [Google Scholar] [CrossRef]
- World Health Organisation. Global Action Plan on the Public Health Response to Dementia 2017–2025; World Health Organisation: Geneva, Switzerland, 2017; Volume 52. [Google Scholar]
- Lane, C.A.; Hardy, J.; Schott, J.M. Alzheimer’s disease. Eur. J. Neurol. 2018, 25, 59–70. [Google Scholar] [CrossRef]
- Xu, W.; Klumbys, E.; Ang, E.L.; Zhao, H. Emerging molecular biology tools and strategies for engineering natural product biosynthesis. Metab. Eng. Commun. 2020, 10, e00108. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.M.; Yang, Z.J.; Xie, Q.; Zhang, Z.K.; Zhang, H.; Ma, J.Y. Natural products for treating colorectal cancer: A mechanistic review. Biomed. Pharmacother. 2019, 117, 109142. [Google Scholar] [CrossRef]
- Andrade, S.; Ramalho, M.J.; Loureiro, J.A.; Do Carmo Pereira, M. Natural compounds for alzheimer’s disease therapy: A systematic review of preclinical and clinical studies. Int. J. Mol. Sci. 2019, 20, 2313. [Google Scholar] [CrossRef]
- Harvey, A.L. Natural products in drug discovery. Drug Discov. Today 2008, 13, 894–901. [Google Scholar] [CrossRef]
- Kim, K.-W.; Roh, J.K.; Wee, H.-J.; Kim, C.N. Natural Product Anticancer Drugs. In Cancer Drug Discovery: Science and History; Springer: Dordrecht, The Netherlands, 2016. [Google Scholar]
- Chari, R.V.J.; Miller, M.L.; Widdison, W.C. Antibody-drug conjugates: An emerging concept in cancer therapy. Angew. Chem. Int. Ed. Engl. 2014, 53, 3796–3827. [Google Scholar] [CrossRef]
- de Claro, R.A.; McGinn, K.; Kwitkowski, V.; Bullock, J.; Khandelwal, A.; Habtemariam, B.; Ouyang, Y.; Saber, H.; Lee, K.; Koti, K.; et al. U.S. Food and Drug Administration approval summary: Brentuximab vedotin for the treatment of relapsed Hodgkin lymphoma or relapsed systemic anaplastic large-cell lymphoma. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2012, 18, 5845–5849. [Google Scholar] [CrossRef]
- Nakamura, T.; Matsumine, A.; Sudo, A. The value of trabectedin in the treatment of soft tissue sarcoma. Ther. Clin. Risk Manag. 2016, 12, 73–79. [Google Scholar] [CrossRef]
- Safavi-Hemami, H.; Brogan, S.E.; Olivera, B.M. Pain therapeutics from cone snail venoms: From Ziconotide to novel non-opioid pathways. J. Proteom. 2019, 190, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Russo, P.; Kisialiou, A.; Lamonaca, P.; Moroni, R.; Prinzi, G.; Fini, M. New drugs from marine organisms in Alzheimer’s disease. Mar. Drugs 2016, 14, 5. [Google Scholar] [CrossRef] [PubMed]
- Kollár, P.; Rajchard, J.; Balounová, Z.; Pazourek, J. Marine natural products: Bryostatins in preclinical and clinical studies. Pharm. Biol. 2014, 52, 237–242. [Google Scholar] [CrossRef]
- Lim, C.S.; Alkon, D.L. PKCε promotes HuD-mediated neprilysin mRNA stability and enhances neprilysin-induced Aβ degradation in brain neurons. PLoS ONE 2014, 9, e97756. [Google Scholar] [CrossRef] [PubMed]
- Soo, R.M.; Hemp, J.; Hugenholtz, P. Free Radical Biology and Medicine Evolution of photosynthesis and aerobic respiration in the cyanobacteria. Free Radic. Biol. Med. 2019, 140, 200–205. [Google Scholar] [CrossRef]
- Silva, B.L.T. Cyanobacteria from Cape Verde Islands: A Contribution to the Diversity and Biotechnological Potential. 2019. Available online: https://hdl.handle.net/10216/125048 (accessed on 11 June 2021).
- Tamaru, Y.; Takani, Y.; Yoshida, T.; Sakamoto, T. Crucial role of extracellular polysaccharides in desiccation and freezing tolerance in the terrestrial cyanobacterium Nostoc commune. Appl. Environ. Microbiol. 2005, 71, 7327–7333. [Google Scholar] [CrossRef]
- Welsh, D.T. Ecological significance of compatible solute accumulation by micro- organisms: From single cells to global climate. FEMS Microbiol. Rev. 2000, 24, 263–290. [Google Scholar] [CrossRef]
- Katoh, H.; Asthana, R.K.; Ohmori, M. Gene expression in the cyanobacterium Anabaena sp. PCC7120 under desiccation. Microb. Ecol. 2004, 47, 164–174. [Google Scholar] [CrossRef]
- Sinha, R.P.; Klisch, M.; Helbling, W.E.; Häder, D.P. Induction of mycosporine-like amino acids (MAAs) in cyanobacteria by solar ultraviolet-B radiation. J. Photochem. Photobiol. B Biol. 2001, 60, 129–135. [Google Scholar] [CrossRef]
- Shashidhar, R.; Kumar, S.A.; Misra, H.S.; Bandekar, J.R. Evaluation of the role of enzymatic and nonenzymatic antioxidant systems in the radiation resistance of Deinococcus. Can. J. Microbiol. 2010, 56, 195–201. [Google Scholar] [CrossRef]
- Fernandes, E.; Silva, E.; Figueira, F.d.S.; Lettnin, A.P.; Carrett-Dias, M.; Filgueira, D.d.M.V.B.; Kalil, S.; Trindade, G.S.; Votto, A.P.d.S. C-Phycocyanin: Cellular targets, mechanisms of action and multi drug resistance in cancer. Pharmacol. Rep. 2018, 70, 75–80. [Google Scholar] [CrossRef]
- Gupta, V.; Ratha, S.K.; Sood, A.; Chaudhary, V.; Prasanna, R. New insights into the biodiversity and applications of cyanobacteria (blue-green algae)—Prospects and challenges. Algal Res. 2013, 2, 79–97. [Google Scholar] [CrossRef]
- Milani, A.; Basirnejad, M.; Shahbazi, S.; Bolhassani, A. Carotenoids: Biochemistry, pharmacology and treatment. Br. J. Pharmacol. 2017, 174, 1290–1324. [Google Scholar] [CrossRef]
- Moodie, L.W.K.; Sepcic, K.; Turk, T.; Frangez, R.; Svenson, J. Natural cholinesterase inhibitors from marine organisms. Nat. Prod. Rep. 2019, 36, 1053–1092. [Google Scholar] [CrossRef]
- Choi, D.Y.; Choi, H. Natural products from marine organisms with neuroprotective activity in the experimental models of Alzheimer’s disease, Parkinson’s disease and ischemic brain stroke: Their molecular targets and action mechanisms. Arch. Pharm. Res. 2015, 38, 139–170. [Google Scholar] [CrossRef]
- Ibrahim, N.H.; Yahaya, M.F.; Mohamed, W.; Teoh, S.L.; Hui, C.K.; Kumar, J. Pharmacotherapy of Alzheimer’s Disease: Seeking Clarity in a Time of Uncertainty. Front. Pharmacol. 2020, 11, 261–277. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K. Cholinesterase inhibitors as Alzheimer’s therapeutics (Review). Mol. Med. Rep. 2019, 20, 1479–1487. [Google Scholar] [CrossRef]
- Grøntvedt, G.R.; Schröder, T.N.; Sando, S.B.; White, L.; Bråthen, G.; Doeller, C.F. Alzheimer’s disease. Curr. Biol. 2018, 28, R645–R649. [Google Scholar] [CrossRef] [PubMed]
- Maresova, P.; Klimova, B.; Novotny, M.; Kuca, K. Alzheimer’s and Parkinson’s Diseases: Expected Economic Impact on Europe—A Call for a Uniform European Strategy. J. Alzheimer’s Dis. 2016, 54, 1123–1133. [Google Scholar] [CrossRef] [PubMed]
- Niu, H.; Álvarez-Álvarez, I.; Guillén-Grima, F.; Aguinaga-Ontoso, I. Prevalencia e incidencia de la enfermedad de Alzheimer en Europa: Metaanálisis. Neurologia 2017, 32, 523–532. [Google Scholar] [CrossRef]
- Coimbra, J.R.M.; Marques, D.F.F.; Baptista, S.J.; Pereira, C.M.F.; Moreira, P.I.; Dinis, T.C.P.; Santos, A.E.; Salvador, J.A.R. Highlights in BACE1 inhibitors for Alzheimer’s disease treatment. Front. Chem. 2018, 6, 178. [Google Scholar] [CrossRef]
- Das, B.; Yan, R. A Close Look at BACE1 Inhibitors for Alzheimer’s Disease Treatment. CNS Drugs 2019, 33, 251–263. [Google Scholar] [CrossRef]
- Yndart, A. Alzheimer ’ s disease: Pathogenesis, diagnostics, and therapeutics. Int. J. Nanomed. 2019, 14, 5541–5554. [Google Scholar]
- Reiss, A.B.; Arain, H.A.; Stecker, M.M.; Siegart, N.M.; Kasselman, L.J. Amyloid toxicity in Alzheimer’s disease. Rev. Neurosci. 2018, 29, 613–627. [Google Scholar] [CrossRef]
- Butterfield, D.A.; Boyd-Kimball, D. Oxidative Stress, Amyloid-β Peptide, and Altered Key Molecular Pathways in the Pathogenesis and Progression of Alzheimer’s Disease. J. Alzheimers Dis. 2018, 62, 1345–1367. [Google Scholar] [CrossRef]
- Cai, Z.; Hussain, M.D.; Yan, L.-J. Microglia, neuroinflammation, and beta-amyloid protein in Alzheimer’s disease. Int. J. Neurosci. 2014, 124, 307–321. [Google Scholar] [CrossRef]
- Martins, R.N.; Villemagne, V.; Sohrabi, H.R.; Chatterjee, P.; Shah, T.M.; Verdile, G.; Fraser, P.; Taddei, K.; Gupta, V.B.; Rainey-Smith, S.R.; et al. Alzheimer’s Disease: A Journey from Amyloid Peptides and Oxidative Stress, to Biomarker Technologies and Disease Prevention Strategies-Gains from AIBL and DIAN Cohort Studies. J. Alzheimer’s Dis. 2018, 62, 965–992. [Google Scholar] [CrossRef]
- Amato, A.; Terzo, S.; Mulè, F. Natural compounds as beneficial antioxidant agents in neurodegenerative disorders: A focus on Alzheimer’s disease. Antioxidants 2019, 8, 608. [Google Scholar] [CrossRef]
- Fan, L.; Mao, C.; Hu, X.; Zhang, S.; Yang, Z.; Hu, Z.; Sun, H.; Fan, Y.; Dong, Y.; Yang, J.; et al. New Insights Into the Pathogenesis of Alzheimer’s Disease. Front. Neurol. 2020, 10, 1312. [Google Scholar] [CrossRef]
- Hampel, H.; Vassar, R.; De Strooper, B.; Hardy, J.; Willem, M.; Singh, N.; Zhou, J.; Yan, R.; Vanmechelen, E.; De Vos, A.; et al. The β-Secretase BACE1 in Alzheimer’s Disease. Biol. Psychiatry 2020, 89, 745–756. [Google Scholar] [CrossRef]
- Moussa-Pacha, N.M.; Abdin, S.M.; Omar, H.A.; Alniss, H.; Al-Tel, T.H. BACE1 inhibitors: Current status and future directions in treating Alzheimer’s disease. Med. Res. Rev. 2020, 40, 339–384. [Google Scholar] [CrossRef]
- Zhang, Y.; Lee, D.H.S. Sink hypothesis and therapeutic strategies for attenuating Aβ levels. Neuroscientist 2011, 17, 163–173. [Google Scholar] [CrossRef]
- Nygaard, H.B. Targeting Fyn Kinase in Alzheimer’s Disease. Biol. Psychiatry 2018, 83, 369–376. [Google Scholar] [CrossRef]
- Francis, P.T.; Palmer, A.M.; Snape, M.; Wilcock, G.K. The cholinergic hypothesis of Alzheimer’s disease: A review of progress. J. Neurol. Neurosurg. Psychiatry 1999, 66, 137–147. [Google Scholar] [CrossRef]
- Maurer, S.V.; Williams, C.L. The Cholinergic System Modulates Memory and Hippocampal Plasticity via Its Interactions with Non-Neuronal Cells. Front. Immunol. 2017, 8, 1489. [Google Scholar] [CrossRef]
- Grossberg, G.T. Cholinesterase inhibitors for the treatment of Alzheimer’s disease:: Getting on and staying on. Curr. Ther. Res. Clin. Exp. 2003, 64, 216–235. [Google Scholar] [CrossRef]
- Perry, E.K.; Perry, R.H.; Blessed, G.; Tomlinson, B.E. Changes in brain cholinesterases in senile dementia of Alzheimer type. Neuropathol. Appl. Neurobiol. 1978, 4, 273–277. [Google Scholar] [CrossRef]
- Ikonomovic, M.D.; Mufson, E.J.; Wuu, J.; Bennett, D.A.; DeKosky, S.T. Reduction of choline acetyltransferase activity in primary visual cortex in mild to moderate Alzheimer’s disease. Arch. Neurol. 2005, 62, 425–430. [Google Scholar] [CrossRef]
- Hampel, H.; Mesulam, M.M.; Cuello, A.C.; Farlow, M.R.; Giacobini, E.; Grossberg, G.T.; Khachaturian, A.S.; Vergallo, A.; Cavedo, E.; Snyder, P.J.; et al. The cholinergic system in the pathophysiology and treatment of Alzheimer’s disease. Brain 2018, 141, 1917–1933. [Google Scholar] [CrossRef]
- Caccamo, A.; Oddo, S.; Billings, L.M.; Green, K.N.; Martinez-Coria, H.; Fisher, A.; LaFerla, F.M. M1 receptors play a central role in modulating AD-like pathology in transgenic mice. Neuron 2006, 49, 671–682. [Google Scholar] [CrossRef]
- Forlenza, O.V.; Spink, J.M.; Dayanandan, R.; Anderton, B.H.; Olesen, O.F.; Lovestone, S. Muscarinic agonists reduce tau phosphorylation in non-neuronal cells via GSK-3beta inhibition and in neurons. J. Neural Transm. 2000, 107, 1201–1212. [Google Scholar] [CrossRef]
- Jones, C.K.; Brady, A.E.; Davis, A.A.; Xiang, Z.; Bubser, M.; Tantawy, M.N.; Kane, A.S.; Bridges, T.M.; Kennedy, J.P.; Bradley, S.R.; et al. Novel selective allosteric activator of the M1 muscarinic acetylcholine receptor regulates amyloid processing and produces antipsychotic-like activity in rats. J. Neurosci. 2008, 28, 10422–10433. [Google Scholar] [CrossRef] [PubMed]
- Boyle, C.D.; Lachowicz, J.E. Orally active and selective benzylidene ketal M2 muscarinic receptor antagonists for the treatment of Alzheimer’s disease. Drug Dev. Res. 2002, 56, 310–320. [Google Scholar] [CrossRef]
- Shirey, J.K.; Brady, A.E.; Jones, P.J.; Davis, A.A.; Bridges, T.M.; Kennedy, J.P.; Jadhav, S.B.; Menon, U.N.; Xiang, Z.; Watson, M.L.; et al. A selective allosteric potentiator of the M1 muscarinic acetylcholine receptor increases activity of medial prefrontal cortical neurons and restores impairments in reversal learning. J. Neurosci. 2009, 29, 14271–14286. [Google Scholar] [CrossRef]
- Seiger, A.; Nordberg, A.; von Holst, H.; Bäckman, L.; Ebendal, T.; Alafuzoff, I.; Amberla, K.; Hartvig, P.; Herlitz, A.; Lilja, A. Intracranial infusion of purified nerve growth factor to an Alzheimer patient: The first attempt of a possible future treatment strategy. Behav. Brain Res. 1993, 57, 255–261. [Google Scholar] [CrossRef]
- Tuszynski, M.H.; Thal, L.; Pay, M.; Salmon, D.P.U.H.S.; Bakay, R.; Patel, P.; Blesch, A.; Vahlsing, H.L.; Ho, G.; Vahlsing, H.; et al. A phase 1 clinical trial of nerve growth factor gene therapy for Alzheimer disease. Nat. Med. 2005, 11, 551–555. [Google Scholar] [CrossRef] [PubMed]
- Ferreira-Vieira, T.H.; Guimaraes, I.M.; Silva, F.R.; Ribeiro, F.M. Alzheimer’s disease: Targeting the Cholinergic System. Curr. Neuropharmacol. 2016, 14, 101–115. [Google Scholar] [CrossRef] [PubMed]
- Malenka, R.C.; Nicoll, R.A. NMDA-receptor-dependent synaptic plasticity: Multiple forms and mechanisms. Trends Neurosci. 1993, 16, 521–527. [Google Scholar] [CrossRef]
- Gupta, K.; Hardingham, G.E.; Chandran, S. NMDA receptor-dependent glutamate excitotoxicity in human embryonic stem cell-derived neurons. Neurosci. Lett. 2013, 543, 95–100. [Google Scholar] [CrossRef]
- Ferreira, I.L.; Bajouco, L.M.; Mota, S.I.; Auberson, Y.P.; Oliveira, C.R.; Rego, A.C. Amyloid beta peptide 1-42 disturbs intracellular calcium homeostasis through activation of GluN2B-containing N-methyl-d-aspartate receptors in cortical cultures. Cell Calcium 2012, 51, 95–106. [Google Scholar] [CrossRef]
- Rönicke, R.; Mikhaylova, M.; Rönicke, S.; Meinhardt, J.; Schröder, U.H.; Fändrich, M.; Reiser, G.; Kreutz, M.R.; Reymann, K.G. Early neuronal dysfunction by amyloid β oligomers depends on activation of NR2B-containing NMDA receptors. Neurobiol. Aging 2011, 32, 2219–2228. [Google Scholar] [CrossRef]
- Texidó, L.; Martín-Satué, M.; Alberdi, E.; Solsona, C.; Matute, C. Amyloid β peptide oligomers directly activate NMDA receptors. Cell Calcium 2011, 49, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Tan, L.; Yu, J.-T.; Tan, L. Tau in Alzheimer’s Disease: Mechanisms and Therapeutic Strategies. Curr. Alzheimer Res. 2018, 15, 283–300. [Google Scholar] [CrossRef] [PubMed]
- Iwata, M.; Watanabe, S.; Yamane, A.; Miyasaka, T.; Misonou, H. Regulatory mechanisms for the axonal localization of tau protein in neurons. Mol. Biol. Cell 2019, 30, 2441–2457. [Google Scholar] [CrossRef] [PubMed]
- Mondragón-Rodríguez, S.; Trillaud-Doppia, E.; Dudilot, A.; Bourgeois, C.; Lauzon, M.; Leclerc, N.; Boehm, J. Interaction of endogenous tau protein with synaptic proteins is regulated by N-methyl-D-aspartate receptor-dependent tau phosphorylation. J. Biol. Chem. 2012, 287, 32040–32053. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.; Noble, W.; Hanger, D.P. Roles of tau protein in health and disease. Acta Neuropathol. 2017, 133, 665–704. [Google Scholar] [CrossRef] [PubMed]
- Jouanne, M.; Rault, S.; Voisin-Chiret, A.-S. Tau protein aggregation in Alzheimer’s disease: An attractive target for the development of novel therapeutic agents. Eur. J. Med. Chem. 2017, 139, 153–167. [Google Scholar] [CrossRef] [PubMed]
- Kutter, S.; Eichner, T.; Deaconescu, A.M.; Kern, D. Regulation of Microtubule Assembly by Tau and not by Pin1. J. Mol. Biol. 2016, 428, 1742–1759. [Google Scholar] [CrossRef]
- Barbier, P.; Zejneli, O.; Martinho, M.; Lasorsa, A.; Belle, V.; Smet-Nocca, C.; Tsvetkov, P.O.; Devred, F.; Landrieu, I. Role of Tau as a Microtubule-Associated Protein: Structural and Functional Aspects. Front. Aging Neurosci. 2019, 11, 204. [Google Scholar] [CrossRef]
- Callahan, L.M.; Vaules, W.A.; Coleman, P.D. Quantitative decrease in synaptophysin message expression and increase in cathepsin D message expression in Alzheimer disease neurons containing neurofibrillary tangles. J. Neuropathol. Exp. Neurol. 1999, 58, 275–287. [Google Scholar] [CrossRef]
- Mudher, A.; Colin, M.; Dujardin, S.; Medina, M.; Dewachter, I.; Naini, S.M.A.; Mandelkow, E.-M.; Mandelkow, E.; Buée, L.; Goedert, M.; et al. What is the evidence that tau pathology spreads through prion-like propagation? Acta Neuropathol. Commun. 2017, 5, 99. [Google Scholar] [CrossRef]
- Haroutunian, V.; Davies, P.; Vianna, C.; Buxbaum, J.D.; Purohit, D.P. Tau protein abnormalities associated with the progression of alzheimer disease type dementia. Neurobiol. Aging 2007, 28, 1–7. [Google Scholar] [CrossRef]
- Bhat, R.V.; Budd Haeberlein, S.L.; Avila, J. Glycogen synthase kinase 3: A drug target for CNS therapies. J. Neurochem. 2004, 89, 1313–1317. [Google Scholar] [CrossRef]
- Schain, M.; Kreisl, W.C. Neuroinflammation in Neurodegenerative Disorders—A Review. Curr. Neurol. Neurosci. Rep. 2017, 17. [Google Scholar] [CrossRef] [PubMed]
- Butterfield, D.A.; Halliwell, B. Oxidative stress, dysfunctional glucose metabolism and Alzheimer disease. Nat. Rev. Neurosci. 2019, 20, 148–160. [Google Scholar] [CrossRef] [PubMed]
- Zündorf, G.; Reiser, G. Calcium dysregulation and homeostasis of neural calcium in the molecular mechanisms of neurodegenerative diseases provide multiple targets for neuroprotection. Antioxid. Redox Signal. 2011, 14, 1275–1288. [Google Scholar] [CrossRef]
- Graffe, M.; Zenisek, D.; Taraska, J.W. A marginal band of microtubules transports and organizes mitochondria in retinal bipolar synaptic terminals. J. Gen. Physiol. 2015, 146, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Ochieng, J.; Tait, L.; Russo, J. Calcium-mediated modulation of microtubule assembly in human breast epithelial cells. Vitr. Cell. Dev. Biol. J. Tissue Cult. Assoc. 1990, 26, 318–324. [Google Scholar] [CrossRef]
- Sinha, K.; Das, J.; Pal, P.B.; Sil, P.C. Oxidative stress: The mitochondria-dependent and mitochondria-independent pathways of apoptosis. Arch. Toxicol. 2013, 87, 1157–1180. [Google Scholar] [CrossRef] [PubMed]
- Birnbaum, J.H.; Wanner, D.; Gietl, A.F.; Saake, A.; Kündig, T.M.; Hock, C.; Nitsch, R.M.; Tackenberg, C. Oxidative stress and altered mitochondrial protein expression in the absence of amyloid-β and tau pathology in iPSC-derived neurons from sporadic Alzheimer’s disease patients. Stem Cell Res. 2018, 27, 121–130. [Google Scholar] [CrossRef]
- Han, Y.; Chen, J.Z. Oxidative stress induces mitochondrial DNA damage and cytotoxicity through independent mechanisms in human cancer cells. BioMed Res. Int. 2013, 2013, 825065. [Google Scholar] [CrossRef] [PubMed]
- Kowalska, M.; Piekut, T.; Prendecki, M.; Sodel, A.; Kozubski, W.; Dorszewska, J. Mitochondrial and Nuclear DNA Oxidative Damage in Physiological and Pathological Aging. DNA Cell Biol. 2020, 39, 1410–1420. [Google Scholar] [CrossRef]
- Attwell, D.; Laughlin, S.B. An energy budget for signaling in the grey matter of the brain. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 2001, 21, 1133–1145. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Oliveros, A.; Jang, M.-H. Dysfunctional Mitochondrial Bioenergetics and Synaptic Degeneration in Alzheimer Disease. Int. Neurourol. J. 2019, 23, S5–S10. [Google Scholar] [CrossRef] [PubMed]
- Ogura, H.; Kosasa, T.; Kuriya, Y.; Yamanishi, Y. Comparison of inhibitory activities of donepezil and other cholinesterase inhibitors on acetylcholinesterase and butylcholinesterase in vitro. Methods Find Exp. Clin. Pharmacol. 2000, 22, 609–613. [Google Scholar] [CrossRef]
- Lilienfeld, S. Galantamine—A novel cholinergic drug with a unique dual mode of action for the treatment of patients with Alzheimer’s disease. CNS Drug Rev. 2002, 8, 159–176. [Google Scholar] [CrossRef]
- U.S. Food and Drug Administration. Reminyl (Galanamine Hydrobromide) Tablets. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/nda/2001/21-169_Reminyl.cfm (accessed on 28 May 2020).
- Liu, J.; Chang, L.; Song, Y.; Li, H.; Wu, Y. The role of NMDA receptors in Alzheimer’s disease. Front. Neurosci. 2019, 13, 43. [Google Scholar] [CrossRef]
- U.S. Food and Drugs Administration. FDA ARICEPT® (Donepezil Hydr Ochloride) Tablets; pp. 1–14, Reference ID: 3096907. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2012/020690s035,021720s008,022568s005lbl.pdf (accessed on 11 May 2021).
- Food and Drug Administration. FDA Exelon Approval Letter; pp. 1–4, Aplication Number: 20823. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/nda/2000/20823_Exelon_Approv.pdf (accessed on 11 May 2021).
- Food and Drug Administration. FDA Full Prescription NAMENDA (Memantine HCl); 2013, pp. 1–18, Reference ID: 3394954. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2013/021487s010s012s014,021627s008lbl.pdf (accessed on 11 May 2021).
- Alzheimer’s Association. Alzheimer’s Association FDA-Approved Treatments for Alzheimer’s. 2019, pp. 1–5. Available online: https://www.alz.org/media/documents/fda-approved-treatments-alzheimers-ts.pdf (accessed on 11 May 2021).
- Holland, A.; Kinnear, S. Interpreting the possible ecological role(s) of cyanotoxins: Compounds for competitive advantage and/or physiological aide? Mar. Drugs 2013, 11, 2239–2258. [Google Scholar] [CrossRef]
- Almeida, J.R.; Freitas, M.; Cruz, S.; Leão, P.N.; Vasconcelos, V.; Cunha, I. Acetylcholinesterase in Biofouling Species: Characterization and Mode of Action of Cyanobacteria-Derived Antifouling Agents. Toxins 2015, 7, 2739–2756. [Google Scholar] [CrossRef]
- Liu, J.; Chen, W.; Xu, Y.; Ren, S.; Zhang, W.; Li, Y. Design, synthesis and biological evaluation of tasiamide B derivatives as BACE1 inhibitors. Bioorganic Med. Chem. 2015, 23, 1963–1974. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, W.; Li, L.; Salvador, L.A.; Chen, T.; Chen, W.; Felsenstein, K.M.; Ladd, T.B.; Price, A.R.; Golde, T.E.; et al. Cyanobacterial peptides as a prototype for the design of potent β-secretase inhibitors and the development of selective chemical probes for other aspartic proteases. J. Med. Chem. 2012, 55, 10749–10765. [Google Scholar] [CrossRef]
- Al-Awadhi, F.H.; Ratnayake, R.; Paul, V.J.; Luesch, H. Tasiamide F, a potent inhibitor of cathepsins D and .E from a marine cyanobacterium. Bioorg. Med. Chem. 2016, 24, 3276–3282. [Google Scholar] [CrossRef]
- Li, Z.; Bao, K.; Xu, H.; Wu, P.; Li, W.; Liu, J.; Zhang, W. Design, synthesis, and bioactivities of tasiamide B derivatives as cathepsin D inhibitors. J. Pept. Sci. 2019, 25, e3154. [Google Scholar] [CrossRef]
- Mahmood, N.A.; Carmichael, W.W. Anatoxin-a(s), an anticholinesterase from the cyanobacterium Anabaena flos-aquae NRC-525-17. Toxicon 1987, 25, 1221–1227. [Google Scholar] [CrossRef]
- Carvalho, L.R.; Costa-Neves, A.; Conserva, G.A.A.; Brunetti, R.L.; Hentschke, G.S.; Malone, C.F.S.; Torres, L.M.B.; Sant’Anna, C.L.; Rangel, M. Biologically active compounds from cyano bacteria extracts: In vivo and in vitro aspects. Braz. J. Pharmacogn. 2013, 23, 471–480. [Google Scholar] [CrossRef]
- Becher, P.G.; Beuchat, J.; Gademann, K.; Jüttner, F. Nostocarboline: Isolation and synthesis of a new cholinesterase inhibitor from Nostoc 78-12A. J. Nat. Prod. 2005, 68, 1793–1795. [Google Scholar] [CrossRef]
- Li, H.B.; Cheng, K.W.; Wong, C.C.; Fan, K.W.; Chen, F.; Jiang, Y. Evaluation of antioxidant capacity and total phenolic content of different fractions of selected microalgae. Food Chem. 2007, 102, 771–776. [Google Scholar] [CrossRef]
- Singh, N.K.; Hasan, S.S.; Kumar, J.; Raj, I.; Pathan, A.A.; Parmar, A.; Shakil, S.; Gourinath, S.; Madamwar, D. Crystal structure and interaction of phycocyanin with β-secretase: A putative therapy for Alzheimer’s Disease. CNS Neurol. Disord. Drug Targets 2014, 13, 691–698. [Google Scholar] [CrossRef]
- Chaubey, M.G.; Patel, S.N.; Rastogi, R.P.; Srivastava, P.L.; Singh, A.K.; Madamwar, D.; Singh, N.K. Therapeutic potential of cyanobacterial pigment protein phycoerythrin: In silico and in vitro study of BACE1 interaction and in vivo Aβ reduction. Int. J. Biol. Macromol. 2019, 134, 368–378. [Google Scholar] [CrossRef]
- Liu, Y.; Jovcevski, B.; Pukala, T.L. C-Phycocyanin from Spirulina Inhibits α-Synuclein and Amyloid-β Fibril Formation but Not Amorphous Aggregation. J. Nat. Prod. 2019, 82, 66–73. [Google Scholar] [CrossRef]
- Mitra, S.; Siddiqui, W.A.; Khandelwal, S. C-Phycocyanin protects against acute tributyltin chloride neurotoxicity by modulating glial cell activity along with its anti-oxidant and anti-inflammatory property: A comparative efficacy evaluation with N-acetyl cysteine in adult rat brain. Chem. Biol. Interact. 2015, 238, 138–150. [Google Scholar] [CrossRef]
- Hwang, J.-H.; Lee, I.-T.; Jeng, K.-C.; Wang, M.-F.; Hou, R.C.-W.; Wu, S.-M.; Chan, Y.-C. Spirulina prevents memory dysfunction, reduces oxidative stress damage and augments antioxidant activity in senescence-accelerated mice. J. Nutr. Sci. Vitaminol. 2011, 57, 186–191. [Google Scholar] [CrossRef]
- Pérez-Juárez, A.; Chamorro, G.; Alva-Sánchez, C.; Paniagua-Castro, N.; Pacheco-Rosado, J. Neuroprotective effect of Arthrospira (Spirulina) platensis against kainic acid-neuronal death. Pharm. Biol. 2016, 54, 1408–1412. [Google Scholar] [CrossRef]
- Sonani, R.R.; Rastogi, R.P.; Singh, N.K.; Thadani, J.; Patel, P.J.; Kumar, J.; Tiwari, A.K.; Devkar, R.V.; Madamwar, D. Phycoerythrin averts intracellular ROS generation and physiological functional decline in eukaryotes under oxidative stress. Protoplasma 2017, 254, 849–862. [Google Scholar] [CrossRef] [PubMed]
- Chaubey, M.G.; Patel, S.N.; Rastogi, R.P.; Madamwar, D.; Singh, N.K. Cyanobacterial pigment protein allophycocyanin exhibits longevity and reduces Aβ-mediated paralysis in C. elegans: Complicity of FOXO and NRF2 ortholog DAF-16 and SKN-1. 3 Biotech 2020, 10, 332. [Google Scholar] [CrossRef] [PubMed]
- Miranda, M.S.; Cintra, R.G.; Barros, S.B.M.; Mancini-Filho, J. Antioxidant activity of the microalga Spirulina maxima. Braz. J. Med. Biol. Res. 1998, 31, 1075–1079. [Google Scholar] [CrossRef] [PubMed]
- Koh, E.J.; Kim, K.J.; Choi, J.; Kang, D.H.; Lee, B.Y. Spirulina maxima extract prevents cell death through BDNF activation against amyloid beta 1-42 (Aβ 1-42 ) induced neurotoxicity in PC12 cells. Neurosci. Lett. 2018, 673, 33–38. [Google Scholar] [CrossRef]
- El-Baky, H.H.A.; El Baz, F.K.; El-Baroty, G.S. Production of phenolic compounds from Spirulina maxima microalgae and its protective effects. Afr. J. Biotechnol. 2009, 8, 7059–7067. [Google Scholar] [CrossRef]
- McCarty, M.F.; Barroso-Aranda, J.; Contreras, F. Oral phycocyanobilin may diminish the pathogenicity of activated brain microglia in neurodegenerative disorders. Med. Hypotheses 2010, 74, 601–605. [Google Scholar] [CrossRef]
- McGahon, B.M.; Martin, D.S.D.; Horrobin, D.F.; Lynch, M.A. Age-related changes in synaptic function: Analysis of the effect of dietary supplementation with ω-3 fatty acids. Neuroscience 1999, 94, 305–314. [Google Scholar] [CrossRef]
- Kelly, L.; Grehan, B.; Della Chiesa, A.; O’Mara, S.M.; Downer, E.; Sahyoun, G.; Massey, K.A.; Nicolaou, A.; Lynch, M.A. The polyunsaturated fatty acids, EPA and DPA exert a protective effect in the hippocampus of the aged rat. Neurobiol. Aging 2011, 32, 2318.e1–2318.e15. [Google Scholar] [CrossRef]
- Olasehinde, T.A.; Olaniran, A.O.; Okoh, A.I.; Koulen, P. Therapeutic potentials of microalgae in the treatment of Alzheimer’s disease. Molecules 2017, 22, 480. [Google Scholar] [CrossRef] [PubMed]
- Dyall, S.C. Long-chain omega-3 fatty acids and the brain: A review of the independent and shared effects of EPA, DPA and DHA. Front. Aging Neurosci. 2015, 7, 52. [Google Scholar] [CrossRef]
- Wu, L.C.; Ho, J.A.A.; Shieh, M.C.; Lu, I.W. Antioxidant and antiproliferative activities of spirulina and Chlorella water extracts. J. Agric. Food Chem. 2005, 53, 4207–4212. [Google Scholar] [CrossRef] [PubMed]
- Pan-utai, W.; Iamtham, S. Extraction, purification and antioxidant activity of phycobiliprotein from Arthrospira platensis. Process Biochem. 2019, 82, 189–198. [Google Scholar] [CrossRef]
- Kumar, R.S.; Shakambari, G.; Ashokkumar, B.; Varalakshmi, P. Inhibition of advanced glycation end products formation and inflammation in C. elegans: Studies of potential of Lyngbya sp. against expression of stress related genes and Live cell imaging. Biocatal. Agric. Biotechnol. 2019, 17, 233–241. [Google Scholar] [CrossRef]
- Manogar, P.; Vijayakumar, S.; Praseetha, P.K. Evaluation of antioxidant and neuroprotective activities of Lyngbya majuscula on human neural tissues. Gene Rep. 2020, 19, 100661. [Google Scholar] [CrossRef]
- LePage, K.T.; Goeger, D.; Yokokawa, F.; Asano, T.; Shioiri, T.; Gerwick, W.H.; Murray, T.F. The neurotoxic lipopeptide kalkitoxin interacts with voltage-sensitive sodium channels in cerebellar granule neurons. Toxicol. Lett. 2005, 158, 133–139. [Google Scholar] [CrossRef]
- Nuzzo, D.; Presti, G.; Picone, P.; Galizzi, G.; Gulotta, E.; Giuliano, S.; Mannino, C.; Gambino, V.; Scoglio, S.; Di Carlo, M. Effects of the aphanizomenon flos-aquae extract (Klamin®) on a neurodegeneration cellular model. Oxid. Med. Cell. Longev. 2018, 2018. [Google Scholar] [CrossRef]
- Nuzzo, D.; Contardi, M.; Kossyvaki, D.; Picone, P.; Cristaldi, L.; Galizzi, G.; Bosco, G.; Scoglio, S.; Athanassiou, A.; Di Carlo, M. Heat-Resistant Aphanizomenon flos-aquae (AFA) Extract (Klamin®) as a Functional Ingredient in Food Strategy for Prevention of Oxidative Stress. Oxid. Med. Cell. Longev. 2019, 2019, 9481390. [Google Scholar] [CrossRef]
- Procházková, T.; Sychrová, E.; Javůrková, B.; Večerková, J.; Kohoutek, J.; Lepšová-Skácelová, O.; Bláha, L.; Hilscherová, K. Phytoestrogens and sterols in waters with cyanobacterial blooms—Analytical methods and estrogenic potencies. Chemosphere 2017, 170, 104–112. [Google Scholar] [CrossRef]
- Tan, J.W.; Kim, M.K. Neuroprotective effects of Biochanin A against β-amyloid-induced neurotoxicity in PC12 cells via a mitochondrial-dependent apoptosis pathway. Molecules 2016, 21, 548. [Google Scholar] [CrossRef]
- Swingle, M.; Ni, L.; Honkanen, R.E. Small-Molecule Inhibitors of Ser/Thr Protein Phosphatases. In Protein Phosphatase Protocols; Methods in Molecular Biology; Springer: Totowa, NJ, USA, 2007; Volume 365, pp. 23–38. [Google Scholar]
- Pereira, S.R.; Vasconcelos, V.M.; Antunes, A. The phosphoprotein phosphatase family of Ser/Thr phosphatases as principal targets of naturally occurring toxins. Crit. Rev. Toxicol. 2011, 41, 83–110. [Google Scholar] [CrossRef]
- Minogue, A.M.; Lynch, A.M.; Loane, D.J.; Herron, C.E.; Lynch, M.A. Modulation of amyloid-β-induced and age-associated changes in rat hippocampus by eicosapentaenoic acid. J. Neurochem. 2007, 103, 914–926. [Google Scholar] [CrossRef]
- Li, T.-T.; Tong, A.-J.; Liu, Y.-Y.; Huang, Z.-R.; Wan, X.-Z.; Pan, Y.-Y.; Jia, R.-B.; Liu, B.; Chen, X.-H.; Zhao, C. Polyunsaturated fatty acids from microalgae Spirulina platensis modulates lipid metabolism disorders and gut microbiota in high-fat diet rats. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2019, 131, 110558. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Grundke-Iqbal, I.; Iqbal, K.; Gong, C.-X. Contributions of protein phosphatases PP1, PP2A, PP2B and PP5 to the regulation of tau phosphorylation. Eur. J. Neurosci. 2005, 22, 1942–1950. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castaneda, A.; Ferraz, R.; Vieira, M.; Cardoso, I.; Vasconcelos, V.; Martins, R. Bridging Cyanobacteria to Neurodegenerative Diseases: A New Potential Source of Bioactive Compounds against Alzheimer’s Disease. Mar. Drugs 2021, 19, 343. https://doi.org/10.3390/md19060343
Castaneda A, Ferraz R, Vieira M, Cardoso I, Vasconcelos V, Martins R. Bridging Cyanobacteria to Neurodegenerative Diseases: A New Potential Source of Bioactive Compounds against Alzheimer’s Disease. Marine Drugs. 2021; 19(6):343. https://doi.org/10.3390/md19060343
Chicago/Turabian StyleCastaneda, Andrea, Ricardo Ferraz, Mónica Vieira, Isabel Cardoso, Vitor Vasconcelos, and Rosário Martins. 2021. "Bridging Cyanobacteria to Neurodegenerative Diseases: A New Potential Source of Bioactive Compounds against Alzheimer’s Disease" Marine Drugs 19, no. 6: 343. https://doi.org/10.3390/md19060343
APA StyleCastaneda, A., Ferraz, R., Vieira, M., Cardoso, I., Vasconcelos, V., & Martins, R. (2021). Bridging Cyanobacteria to Neurodegenerative Diseases: A New Potential Source of Bioactive Compounds against Alzheimer’s Disease. Marine Drugs, 19(6), 343. https://doi.org/10.3390/md19060343