
Characterization of Protein Hydrolysates from Fish Discards and By-Products from the North-West Spain Fishing Fleet as Potential Sources of Bioactive Peptides


 ,
,  , ,
, , 

Abstract

1. Introduction
2. Results and Discussion
2.1. Chemical Composition of FPH
2.2. Antioxidant Activity of FPH
2.2.1. DPPH Scavenging Activity
2.2.2. ABTS•+ Radical Scavenging Activity
2.2.3. Reducing Power
2.3. Metal Chelating Activity of FPH
2.3.1. Cu2+ Chelating Activity
2.3.2. Fe2+ Chelating Activity
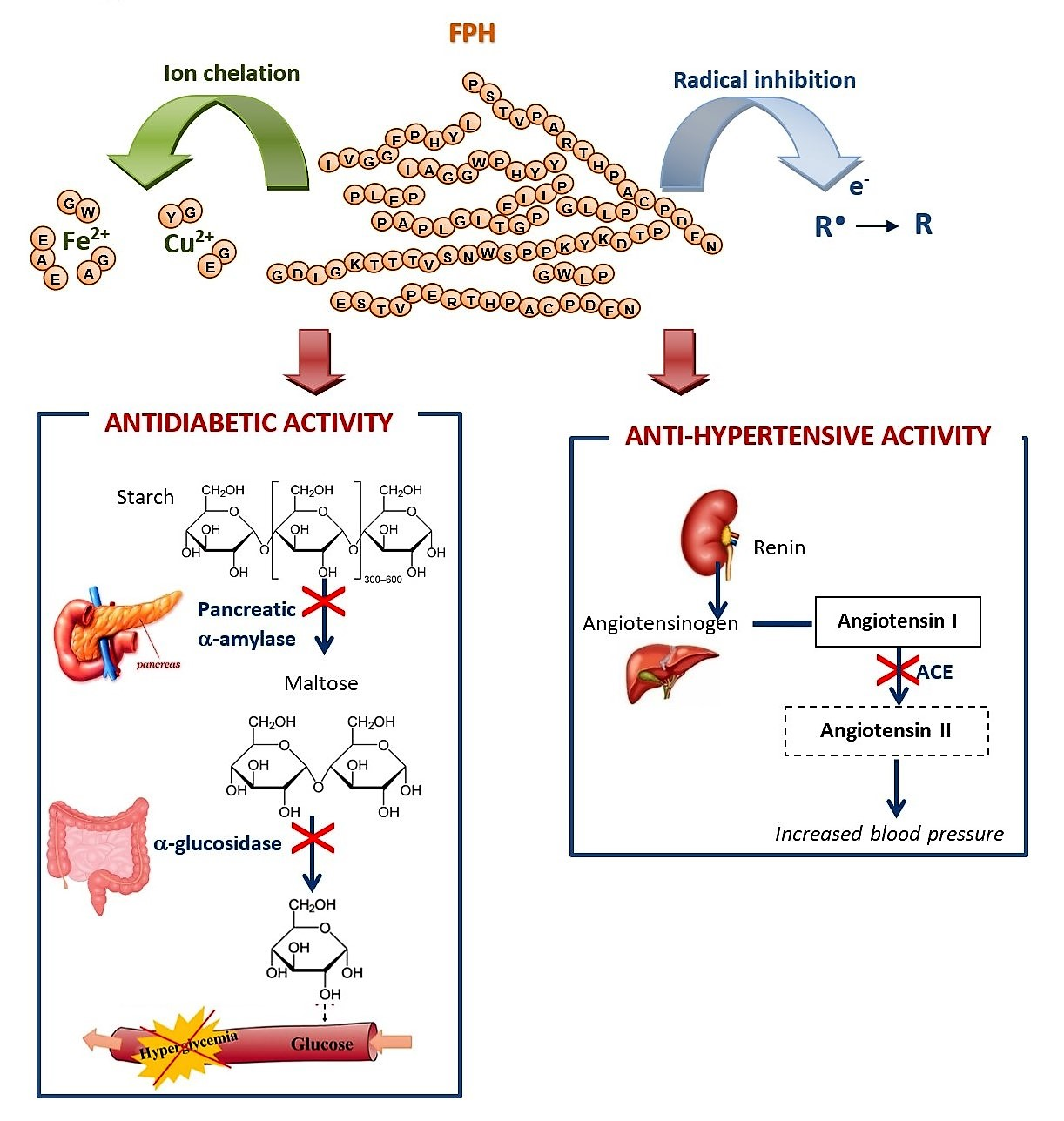
2.4. Anti-Diabetic Activity
α-Amylase and α-Glucosidase Inhibitory Activities
2.5. ACE Inhibitory Activity
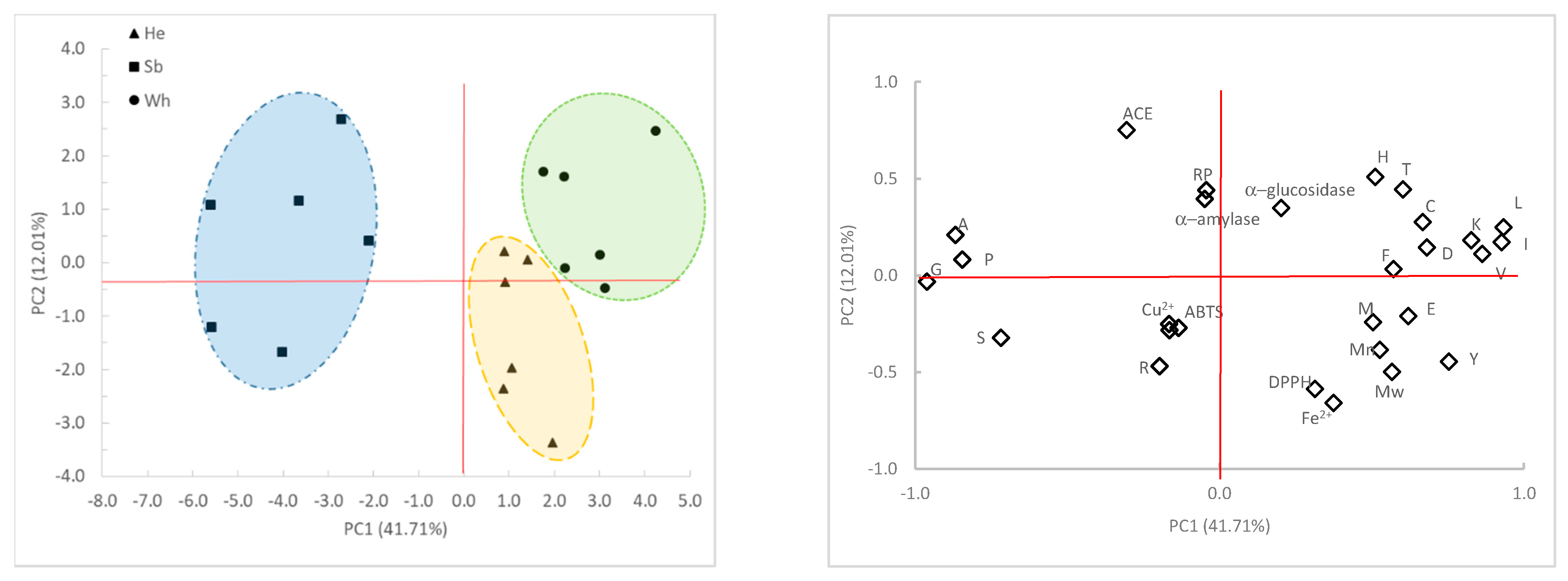
2.6. Principal Components Analysis
3. Material and Methods
3.1. Fish Materials
3.2. Production of Fish Protein Hydrolysates
3.3. Chemical Characterization of FPH
3.4. Amino Acid Analysis
3.5. Molecular Weight Analysis
3.6. Biological Activities
3.6.1. DPPH Radical Scavenging Activity
3.6.2. ABTS Radical Scavenging Activity
3.6.3. Reducing Power
3.6.4. Cu2+ Chelating Activity
3.6.5. Fe2+ Chelation Activity
3.6.6. α-Amylase Inhibitory Activity
3.6.7. α-Glucosidase inhibitory activity
3.6.8. ACE Inhibitory Activity
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture-Meeting the Sustainable Development Goals; Food and Agriculture Organization of the United Nations: Rome, Italy, 2018; pp. 2–75. [Google Scholar]
- FAO. The State of World Fisheries and Aquaculture. Sustainability in Action; Food and Agriculture Organization of the United Nations: Rome, Italy, 2020; pp. 1–73. [Google Scholar]
- Prokopov, I.A.; Kovaleva, E.L.; Minaeva, E.D.; Pryakhina, E.A.; Savin, E.V.; Gamayunova, A.V.; Pozharitskaya, O.N.; Makarov, V.G.; Shikov, A.N. Animal-derived medicinal products in Russia: Current nomenclature and specific aspects of quality control. J. Ethnopharmacol. 2019, 240, 111933. [Google Scholar] [CrossRef] [PubMed]
- Le Gouic, A.V.; Harnedy, P.A.; FitzGerald, R.J. Bioactive Peptides from Fish Protein By-Products. In Bioactive Molecules in Food; Mérillon, J.-M., Ramawat, K.G., Eds.; Reference Series in Phytochemistry; Springer Nature Switzerland AG: Berlin/Heidelberg, Germany, 2019. [Google Scholar] [CrossRef]
- Idowu, A.T.; Igiehon, O.O.; Idowu, S.; Olatunde, O.O.; Benjakul, S. Bioactivity Potentials and General Applications of Fish Protein Hydrolysates. Int. J. Pept. Res. Ther. 2021, 27, 109–118. [Google Scholar] [CrossRef]
- Shikov, A.N.; Flisyuk, E.V.; Obluchinskaya, E.D.; Pozharitskaya, O.N. Pharmacokinetics of Marine-Derived Drugs. Mar. Drugs 2020, 18, 557. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.S.S.; Nazeer, R.A.; Jaiganesh, R. Purification and identification of antioxidant peptides from the skin protein hydrolysate of two marine fishes, horse mackerel (Magalaspis cordyla) and croaker (Otolithes ruber). Amino Acids 2012, 42, 1641–1649. [Google Scholar] [CrossRef] [PubMed]
- Naqash, S.Y.; Nazeer, R.A. Antioxidant and functional properties of protein hydrolysates from pink perch (Nemipterus japonicus) muscle. J. Food Sci. Technol. 2013, 50, 972–978. [Google Scholar] [CrossRef]
- Pires, C.; Clemente, T.; Batista, I. Functional and antioxidative properties of protein hydrolysates from Cape hake by-products prepared by three different methodologies. J. Sci. Food Agric. 2012, 93, 771–780. [Google Scholar] [CrossRef]
- Wijesekara, I.; Kim, S. Angiotensin-I-converting enzyme (ACE) inhibitors from marine resources: Propects in the pharmaceutical industry. Mar. Drugs 2010, 8, 1080–1093. [Google Scholar] [CrossRef]
- Wang, J.; Hu, J.; Cui, J.; Bai, X.; Du, Y.; Miyaguchi, Y. Purification and identification of a ACE inhibitory peptide from oyster proteins hydrolysate and the antihypertensive effect of hydrolysate in spontaneously hypertensive rats. Food Chem. 2008, 111, 302–308. [Google Scholar] [CrossRef]
- Wu, H.; He, H.; Chen, X.; Sun, C.; Zhang, Y.; Zhou, B. Purifcation and identification of novel angiotensin-I-converting enzyme inhibitory peptides from shark meat hydrolysate. Process. Biochem. 2008, 43, 457–461. [Google Scholar] [CrossRef]
- Ahn, C.B.; Jeon, Y.J.; Kim, Y.T.; Je, J.Y. Angiotensin I converting enzyme (ACE) inhibitory peptides from salmon byproduct protein hydrolysate by Alcalase hydrolysis. Process. Biochem. 2012, 47, 2240–2245. [Google Scholar] [CrossRef]
- Salampessy, J.; Reddy, N.; Phillips, M. Isolation and characterization of nutraceutically potential ACE-inhibitory peptides from leatherjacket (Meuchenia sp.) protein hydrolysates. LBWTAP 2017, 80, 430–436. [Google Scholar] [CrossRef]
- Urakova, I.N.; Pozharitskaya, O.N.; Demchenko, D.V.; Shikov, A.N.; Makarov, V.G. The biological activities of fish peptides and methods of their isolation. Russ. J. Mar. Biol 2012, 38, 417–422. [Google Scholar] [CrossRef]
- Rivero-Pino, F.; Espejo-Carpio, F.J.; Guadix, E.M. Antidiabetic Food-Derived Peptides for Functional Feeding: Production, Functionality and In Vivo Evidences. Foods 2020, 9, 983. [Google Scholar] [CrossRef] [PubMed]
- Chalamaiah, M.; Kumar, B.D.; Hemalatha, R.; Jyothirmayi, T. Fish protein hydrolysates: Proximate composition, amino acid composition, antioxidant activities and applications: A review. Food Chem. 2012, 135, 3020–3038. [Google Scholar] [CrossRef]
- Barkia, A.; Bougatef, A.; Khaled, H.B.; Nasri, M. Antioxidant activities of sardinelle heads and/or viscera protein hydrolysates prepared by enzymatic treatment. J. Food Biochem. 2009, 34, 303–320. [Google Scholar] [CrossRef]
- Bougatef, A.; Hajji, M.; Balti, R.; Lassoued, I.; Triki-Ellouz, Y.; Nasri, M. Antioxidant and free radical-scavenging activities of smooth hound (Mustelus mustelus) muscle protein hydrolysates obtained by gastrointestinal proteases. Food Chem. 2009, 114, 1198–1205. [Google Scholar] [CrossRef]
- Jridi, M.; Lassoued, I.; Nasri, R.; Ayadi, M.A.; Nasri, M.; Souissi, N. Characterization and potential use of cuttlefish skin gelatin hydrolysates prepared by different microbial proteases. BioMed Res. Int. 2014, 461728. [Google Scholar] [CrossRef]
- Chi, C.F.; Wang, B.; Deng, Y.Y.; Wang, Y.M.; Deng, S.G.; Ma, J.Y. Isolation and characterization of three antioxidant pentapeptides from protein hydrolysate of monkfish (Lophius litulon) muscle. Int. Food Res. J. 2014, 55, 222–228. [Google Scholar] [CrossRef]
- Jiang, H.; Tong, T.; Sun, J.; Xu, Y.; Zhao, Z.; Liao, D. Purification and characterization of antioxidative peptides from round scad (Decapterus maruadsi) muscle protein hydrolysate. Food Chem. 2014, 154, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Klompong, V.; Benjakul, S.; Kantachote, D.; Shahidi, F. Antioxidative activity and functional properties of protein hydrolysate of yellow stripe trevally (Selaroides eptolepis) as influenced by the degree of hydrolysis and enzyme type. Food Chem. 2007, 102, 317–1327. [Google Scholar] [CrossRef]
- Nakajima, K.; Yoshie-Stark, Y.; Ogushi, M. Comparison of ACE inhibitory and DPPH radical scavenging activities of fish muscle hydrolysates. Food Chem. 2009, 114, 844–851. [Google Scholar] [CrossRef]
- Jao, C.-L.; Ko, W.C. 1,1-Diphenyl-2-picrylhydrazyl (DPPH) radical scavenging by protein hydrolyzates from tuna cooking juice. Fish. Sci. 2002, 68, 430–435. [Google Scholar] [CrossRef]
- Jun, S.Y.; Park, P.J.; Jung, W.K.; Kim, S.K. Purification and characterization of an antioxidative peptide from enzymatic hydrolysate of yellowfin sole (Limanda aspera) frame protein. Eur. Food Res. Technol. 2004, 219, 20–26. [Google Scholar]
- Raghavan, S.; Kristinsson, H.G.; Leeuwenburgh, C. Radical scavenging and reducing ability of tilapia (Oreochromis niloticus) protein hydrolysates. J. Agric. Food Chem. 2008, 56, 10359–10367. [Google Scholar] [CrossRef]
- Teixeira, B.; Pires, C.; Nunes, M.L.; Batista, I. Effect of in vitro gastrointestinal digestion on the antioxidant activity of protein hydrolysates prepared from Cape hake by-products. Int. J. Food Sci. Technol. 2016, 51, 2528–2536. [Google Scholar] [CrossRef]
- Najafian, L.; Babji, A.S. A review of fish-derived antioxidant and antimicrobial peptides: Their production, assessment, and applications. Peptides 2012, 33, 178–185. [Google Scholar] [CrossRef]
- Chalamaiah, M.; Jyothirmayi, T.; Diwan, P.V.; Kumar, B.D. Antioxidant activity and functional properties of enzymatic protein hydrolysates from common carp (Cyprinus carpio) roe (egg). J. Food Sci. Technol. 2015, 52, 5817–5825. [Google Scholar] [CrossRef]
- Chai, T.T.; Tong, S.R.; Law, Y.C.; Ismail, N.I.M.; Manan, F.A.; Wong, F.C. Anti-Oxidative, Metal Chelating and Radical Scavenging Effects of Protein Hydrolysates from Blue-spotted Stingray. Trop. J. Pharm. Res. 2015, 14, 1349–1355. [Google Scholar] [CrossRef]
- Bkhairia, I.; Rabeb, B.S.; Rim, N.; Mourad, J.; Sofiane, G. In-vitro antioxidant and functional properties of protein hydrolysates from golden grey mullet prepared by commercial, microbial and visceral proteases. J. Food Sci. Technol. 2016, 53, 2902–2912. [Google Scholar] [CrossRef]
- Foh, M.B.K.; Amadou, I.; Foh, B.M.; Kamara, M.T.; Xia, W. Functionality and Antioxidant Properties of Tilapia (Oreochromis niloticus) as Influenced by the Degree of Hydrolysis. Int. J. Mol. Sci. 2010, 11, 1851–1869. [Google Scholar] [CrossRef]
- Apak, R.; Güçlü, K.; Demirata, B.; Özyürek, M.; Çelik, S.E.; Bektaşoğlu, B.; Berker, K.I.; Özyurt, D. Comparative Evaluation of Various Total Antioxidant Capacity Assays Applied to Phenolic Compounds with the CUPRAC Assay. Molecules 2007, 12, 1496–1547. [Google Scholar] [CrossRef]
- Yildirim, A.; Mavi, A.; Kara, A.A. Determination of antioxidant and antimicrobial activities of Rumex crispus L. extracts. J. Agric. Food Chem. 2001, 49, 4083–4089. [Google Scholar] [CrossRef]
- Ktari, N.; Jridi, M.; Bkhairia, I.; Sayari, N.; Salah, R.B.; Nasri, M. Functionalities and antioxidant properties of protein hydrolysates from muscle of zebra blenny (Salaria basilisca) obtained with different crude protease extracts. Int. Food Res. J. 2012, 49, 747–756. [Google Scholar] [CrossRef]
- García-Moreno, P.J.; Batista, I.; Pires, C.; Bandarra, N.M.; Espejo-Carpio, F.J.; Guadix, A.; Guadix, E.M. Antioxidant activity of protein hydrolysates obtained from discarded Mediterranean fish species. Food Res. Int. 2014, 65, 469–476. [Google Scholar] [CrossRef]
- Sayari, N.; Sila, A.; Haddar, A.; Balti, R.; Ellouz-Chaabouni, E.; Bougatef, A. Valorisation of smooth hound (Mustelus mustelus) waste biomass through recovery of functional, antioxidative and antihypertensive bioactive peptides. Environ. Sci. Pollut. Res. 2016, 23, 366–376. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.Y.; Tang, Y.; Zhu, B.W.; Qin, L.; Li, D.M.; Yang, J.F.; Lei, K.; Murata, Y. Antioxidant activity of hydrolysates obtained from scallop (Patinopecten yessoensis) and abalone (Haliotis discus hannai Ino) muscle. Food Chem. 2012, 132, 815–822. [Google Scholar] [CrossRef]
- Chen, X.; Hu, X.; Li, L.; Yang, X.; Wu, Y.; Lin, W.; Zhao, Y.; Ma, H.; Wei, Y. Antioxidant properties of tilapia component protein hydrolysates and the membrane ultrafiltration fractions. Adv. Mat. Res. 2015, 1073–1076, 1812–1817. [Google Scholar] [CrossRef]
- Zhang, T.; Li, Y.; Miao, M.; Jiang, B. Purification and characterisation of a new antioxidant peptide from chickpea (Cicer arietium L.) protein hydrolysates. Food Chem. 2011, 128, 28–33. [Google Scholar]
- You, L.; Zhao, M.; Regenstein, J.M.; Ren, J. In vitro antioxidant activity and in vivo anti-fatigue effect of loach (Misgurnus anguillicaudatus) peptides prepared by papain digestion. Food Chem. 2011, 124, 188–194. [Google Scholar] [CrossRef]
- Zhu, L.; Chen, J.; Tang, X.; Xiong, Y.L. Reducing, radical scavenging, and chelation properties of in vitro digests of Alcalase-treated zein hydrolysate. J. Agric. Food Chem. 2008, 56, 2714–2721. [Google Scholar] [CrossRef]
- Kong, B.; Xiong, Y.L. Antioxidant activity of zein hydrolysates in a liposome system and the possible mode of action. J. Agric Food Chem. 2006, 54, 6059–6068. [Google Scholar] [CrossRef] [PubMed]
- Jai ganesh, R.; Nazeer, R.A.; Kumar, N.S. Purification and identification of antioxidant peptide from black pomfret, Parastromateus niger (Bloch, 1975) viscera protein hydrolysate. Food Sci. Biotechnol. 2011, 20, 1087. [Google Scholar] [CrossRef]
- Je, J.Y.; Lee, K.H.; Lee, M.H.; Ahn, C.B. Antioxidant and antihypertensive protein hydrolysates produced from tuna liver by enzymatic hydrolysis. Food Res. Int. 2009, 42, 1266–1272. [Google Scholar] [CrossRef]
- Intarasirisawat, R.; Benjakul, S.; Wu, J.; Visessanguan, W. Isolation of antioxidative and ACE inhibitory peptides from protein hydrolysate of skipjack (Katsuwana pelamis) roe. J. Funct. Foods 2013, 5, 1854–1862. [Google Scholar] [CrossRef]
- Ngoh, Y.Y.; Gan, C.Y. Enzyme-assisted extraction and identifification of antioxidative and a-amylase inhibitory peptides from Pinto beans (Phaseolus vulgaris cv. Pinto). Food Chem. 2016, 190, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Siala, R.; Khabir, A.; Lassoued, I.; Abdelhedi, O.; Elfeki, A.; Vallaeys, T.; Nasri, M. Functional and antioxidant properties of protein hydrolysates from grey triggerfish muscle and in vivo evaluation of hypoglycemic and hypolipidemic activities. Appl. Environ. Microbiol. 2016, 4, 105–119. [Google Scholar]
- Salem, R.B.S.; Ktari, N.; Bkahairia, I.; Nasri, R.; Mora, L.; Kallel, R.; Hamdi, S.; Jamoussi, K.; Boudaouara, T.; Feki, A.; et al. In vitro and in vivo anti-diabetic and anti-hyperlipidemic effects of protein hydrolysates from octopus vulgaris in alloxanic rats. Food Res. Int. 2018, 106, 952–963. [Google Scholar] [CrossRef]
- Mendenieks, L.; Vasiljevic, T. Underutilised fish as sources of bioactive peptides with potential health benefits. Food Aust. 2008, 60, 581–588. [Google Scholar]
- Matsui, T.; Oki, T.; Osajima, Y. Isolation and Identification of peptidic a-glucosidase inhibitors derived from sardine muscle hydrolyzate. Z Naturforsch C J. Biosci. 1999, 54, 259–263. [Google Scholar] [CrossRef]
- Wang, R.; Zhao, H.; Pan, X.; Orfila, C.; Lu, W.; Ma, Y. Preparation of bioactive peptides with antidiabetic, antihypertensive, and antioxidant activities and identification of α-glucosidase inhibitory peptides from soy protein. Food Sci. Nutr. 2019, 7, 1848–1856. [Google Scholar] [CrossRef]
- Awosika, T.O.; Aluko, R.E. Inhibition of the in vitro activities of α-amylase, α-glucosidase and pancreatic lipase by yellow field pea (Pisum sativum L.) protein hydrolysates. Int. J. Food Sci. 2019, 54, 2021–2034. [Google Scholar] [CrossRef]
- Karimi, A.; Azizi, M.H.; Gavlighi, H.A. Fractionation of hydrolysate from corn germ protein by ultrafiltration: In vitro antidiabetic and antioxidant activity. Food Sci. Nutr. 2020, 8, 2395–2405. [Google Scholar] [CrossRef] [PubMed]
- Arise, R.O.; Idi, J.J.; Mic-Braimoh, I.M.; Korode, E.; Ahmed, R.N.; Osemwegie, O. In vitro Angiotesin-1-converting enzyme, α-amylase and α-glucosidase inhibitory and antioxidant activities of Luffa cylindrical (L.) M. Roem seed protein hydrolysate. Heliyon 2019, 5, e01634. [Google Scholar] [CrossRef]
- Bougatef, A.; Nedjar-Arroume, N.; Ravallec-Plé, R.; Leroy, Y.; Guillochon, D.; Barkia, A.; Nasri, M. Angiotensin I-converting enzyme (ACE) inhibitory activities of sardinelle (Sardinella aurita) by-products protein hydrolysates obtained by treatment with microbial and visceral fish serine proteases. Food Chem. 2008, 111, 350–356. [Google Scholar] [CrossRef] [PubMed]
- Pires, C.; Teixeira, B.; Cardoso, C.; Mendes, R.; Nunes, M.L.; Batista, I. Cape hake protein hydrolysates prepared from alkaline solubilised proteins pre-treated with citric acid and calcium ions: Functional properties and ACE inhibitory activity. Process. Biochem. 2015, 50, 1006–1015. [Google Scholar] [CrossRef]
- Ngo, D.H.; Ryu, B.; Kim, S.K. Active peptides from skate (Okamejei kenojei) skin gelatin diminish angiotensin-I converting enzyme activity and intracellular free radical-mediated oxidation. Food Chem. 2014, 143, 246–255. [Google Scholar] [CrossRef]
- Kim, H.J.; Park, K.H.; Shin, J.H.; Lee, J.S.; Jeu, M.S.; Lee, D.H.; Kim, J.S. Antioxidant and ACE inhibiting activities of the rockfish Sebastes hubbsi skin gelatin hydrolysates produced by sequential two-step enzymatic hydrolysis. Fish. Aquat. Sci. 2011, 14, 1–10. [Google Scholar] [CrossRef]
- Lee, S.H.; Qian, Z.J.; Kim, S.K. A novel angiotensin I converting enzyme inhibitory peptide from tuna frame protein hydrolysate and its antihypertensive effect in spontaneously hypertensive rats. Food Chem. 2010, 118, 96–102. [Google Scholar] [CrossRef]
- Neves, A.C.; Harnedy, P.A.; O´Keeffe, M.B.; Alashi, M.A.; Aluko, R.E.; FitzGerald, R.J. Peptide identification in a salmon gelatin hydrolysate with antihypertensive, dipeptidyl peptidase IV inhibitory and antioxidant activities. Food Res. Int. 2017, 100, 112–120. [Google Scholar] [CrossRef]
- Alemán, A.; Perez-Santin, E.; Bordenave-Juchereau, S.; Arnaudin, I.; Gomez-Guillen, M.C.; Montero, P. Squid gelatin hydrolysates with antihypertensive, anticancer and antioxidant activity. Food Res. Int. 2011, 44, 1044–1051. [Google Scholar] [CrossRef]
- Vázquez, J.A.; Fernández-Compás, A.; Blanco, M.; Rodríguez-Amado, I.; Moreno, H.; Borderías, J.; Pérez-Martín, R.I. Development of bioprocesses for the integral valorisation of fish discards. Biochem. Eng. J. 2019, 144, 198–208. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 16th ed.; Association of Official Analytical Chemistry: Washington, DC, USA, 1997. [Google Scholar]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Miller, E.L.; Bimbo, A.P.; Walters, D.E.; Barlow, S.M.; Sheridan, B. Determination of nitrogen solubility in dilute pepsin hydrochloric acid solution of fish meal: Interlaboratory study. J. AOAC Int. 2002, 85, 1374–1381. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.; Spackman, D.H.; Stein, W.H. Chromatography of amino acids on sulfonated polystyrene resins. An improved system. Anal. Chem. 1958, 30, 1185–1190. [Google Scholar] [CrossRef]
- Vazquez, J.A.; Rodríguez-Amado, I.; Sotelo, C.G.; Sanz, N.; Pérez-Martin, R.I.; Valcárcel, J. Production, Characterization, and Bioactivity of Fish Protein Hydrolysates from Aquaculture Turbot (Scophthalmus maximus) Wastes. Biomolecules 2020, 10, 310. [Google Scholar] [CrossRef]
- Shimada, K.; Fujikawa, K.; Yahara, K.; Nakamura, T. Antioxidative properties of xanthan on the antioxidantion of soybean oil in cyclodextrin emulsion. J. Agric. Food Chem. 1992, 40, 945–948. [Google Scholar] [CrossRef]
- Picot, L.; Ravallec, R.; Péron, M.; Vandanjon, L.; Jaouen, P.; Derouiniot, M.; Guérard, F.; Chabeaud, A.; LeGal, Y. Impact of ultrafiltration and nanofiltration of an industrial fish protein hydrolysate on its bioactive properties. J. Sci. Food Agric. 2010, 90, 1819–1826. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Oyaizu, M. Antioxidative activities of browning products of glucosamine fractionated by organic solvent and thin-layer chromatography. Nippon Shokuhin Kogyo Gakkaishi 1988, 35, 771–775. [Google Scholar] [CrossRef]
- Saiga, A.; Tanabe, S.; Nishimura, T. Antioxidant Activity of Peptides Obtained from Porcine Myofibrillar Proteins by Protease Treatment. J. Agric. Food Chem. 2003, 51, 3661–3667. [Google Scholar] [CrossRef]
- Torres-Fuentes, C.; Alaiz, M.; Vioque, J. Affinity purification and characterisation of chelating peptides from chickpea protein hydrolysates. Food Chem. 2011, 129, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Welch, E.A.; Welch, B. Role of ferritin as a lipid oxidation catalyst in muscle food. J. Agric. Food Chem. 1990, 38, 674–677. [Google Scholar]
- Hansawasdi, C.; Kawabata, J.; Kasai, T. α-Amylase Inhibitors from Roselle (Hibiscus sabdariffa Linn.) Tea. Biosci. Biotechnol. Biochem. 2000, 64, 1041–1043. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.I.; Apostolidis, E.; Shetty, K. In vitro studies of eggplant (Solanum Melongena) phenolics as inhibitors of key enzymes relelvant for type 2 diabetes and hypertension. Bioresour. Technol. 2008, 99, 2981–2988. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| FPH | Y (%) | Mo (%) | OM (%) | Ash (%) | TP (%) | TF (%) | Dig (%) | Mn (Da) | Mw (Da) |
|---|---|---|---|---|---|---|---|---|---|
| BW_Sb | 97.3 ± 1.2 | 5.0 ± 0.2 a,b,c | 79.6 ± 0.3 | 15.4 ± 0.6 d | 73.5 ± 2.9 | 5.0 ± 2.4 a,b,c,d,e | 93.4 ± 0.4 c,d,e,f,g | 632 ± 26 a,b | 940 ± 49 a |
| RS_Sb | 93.2 ± 0.8 | 3.5 ± 0.1 a,b | 82.1 ± 3.4 | 14.4 ± 3.4 b,c,d | 74.7 ± 2.6 | 6.2 ± 1.8 a,b,c,d,e | 94.4 ± 1.0 d,e,f,g,h | 557 ± 38 a | 848 ± 80 a |
| Po_Sb | 95.7 ± 2.8 | 4.1 ± 0.9 a,b,c | 83.1 ± 0.7 | 12.9 ± 0.1 a,b,c | 76.6 ± 3.0 | 4.8 ± 0.3 a,b,c,d,e | 95.8 ± 1.1 g,h | 634 ± 47 a,b | 954 ± 75 a |
| Gu_Sb | 94.7 ± 2.6 | 6.7 ± 2.1 c,d | 81.5 ± 2.2 | 11.9 ± 0.2 a | 74.5 ± 1.9 | 4.4 ± 0.6 a,b,c,d,e | 92.7 ± 1.1 b,c,d,e | 631 ± 100 a,b | 941 ± 107 a |
| M_Sb | 90.2 ± 1.9 | 3.3 ± 1.1 a | 82.7 ± 1.7 | 14.0 ± 0.6 a,b,c,d | 75.2 ± 4.0 | 3.6 ± 0.1 a,b,c,d | 93.9 ± 0.8 d,e,f,g | 733 ± 76 a,b,c | 1168 ± 133 a,b,c |
| AHM_Sb | 88.3 ± 1.4 | 5.0 ± 2.5 a,b,c | 83.0 ± 2.6 | 12.1 ± 0.1 a,b | 72.1 ± 4.1 | 7.9 ± 1.1 c,d,e | 92.8 ± 0.5 b,c,d,e,f | 1016 ± 141 c,d,e | 1890 ± 113 e |
| BW_He | 87.7 ± 1.7 | 5.1 ± 0.7 a,b,c | 81.0 ± 0.7 | 14.0 ± 0.2 a,b,c,d | 70.1 ± 3.0 | 9.4 ± 5.8 e | 92.3 ± 0.5 a,b,c,d,e | 1081 ± 78 d,e | 2353 ± 98 f |
| RS_He | 87.5 ± 1.5 | 5.2 ± 1.2 a,b,c,d | 81.8 ± 2.3 | 13.0 ± 1.1 a,b,c,d | 70.2 ± 3.1 | 8.0 ± 5.1 d,e | 89.7 ± 0.9 a | 1099 ± 125 d,e | 2490 ± 181 f,g |
| Po_He | 90.4 ± 0.8 | 4.9 ± 0.3 a,b,c | 80.5 ± 0.2 | 14.7 ± 0.5 c,d | 71.1 ± 4.1 | 2.3 ± 0.7 a,b,c | 91.7 ± 0.7 a,b,c,d | 992 ± 84 c,d,e | 2329 ± 191 f |
| Gu_He | 85.3 ± 1.2 | 5.6 ± 0.3 a,b,c,d | 80.4 ± 1.5 | 14.1 ± 1.2 a,b,c,d | 71.9 ± 4.8 | 3.3 ± 0.4 a,b,c,d | 90.7 ± 0.3 a,b,c | 1247 ± 134 e | 2823 ± 122 g |
| M_He | 86.1 ± 0.7 | 6.3 ± 0.4 b,c,d | 79.7 ± 0.6 | 14.1 ± 0.2 a,b,c,d | 69.8 ± 2.0 | 2.5 ± 0.2 a,b,c,d | 92.0 ± 0.1 a,b,c,d,e | 1785 ± 127 f | 2875 ± 225 g |
| AHM_He | 90.3 ± 2.2 | 5.1 ± 1.0 a,b,c | 81.1 ± 1.2 | 13.9 ± 0.2 a,b,c,d | 70.4 ± 1.8 | 7.2 ± 1.7 b,c,d,e | 90.3 ± 0.5 a,b | 1203 ± 136 e | 2359 ± 237 f |
| BW_Wh | 93.1 ± 0.5 | 5. 2 ± 0.8 a,b,c,d | 81.2 ± 0.4 | 13.6 ± 0.4 a,b,c,d | 74.7 ± 2.8 | 3.8 ± 0.5 a,b,c,d | 97.2 ± 0.4 h | 779 ± 117 a,b,c | 1428 ± 137 c,d |
| RS_Wh | 88.5 ± 0.9 | 5.5 ± 1.0 a,b,c,d | 81.7 ± 0.9 | 12.8 ± 0.0 a,b,c | 75.8 ± 1.9 | 1.5 ± 0.7 a | 94.6 ± 2.5 e,f,g,h | 823 ± 117 a,b,c,d | 1394 ± 173 b,c,d |
| Po_Wh | 91.6 ± 0.7 | 8.0 ± 1.4 d | 79.3 ± 2.2 | 12.7 ± 0.8 a,b,c | 73.0 ± 3.8 | 2.9 ± 0.3 a,b,c,d | 93.2 ± 0.4 b,c,d,e,f,g | 651 ± 100 a,b | 978 ± 126 a,b |
| Gu_Wh | 83.4 ± 1.1 | 5.1 ± 1.5 a,b,c | 80.9 ± 1.6 | 14.0 ± 0.1 a,b,c,d | 75.8 ± 3.2 | 1.7 ± 0.6 a,b | 94.4 ± 0.7 d,e,f,g,h | 1016 ± 85 c,d,e | 1809 ± 85 d,e |
| M_Wh | 90.9 ± 3.1 | 5.0 ± 0.5 a,b,c | 81.7 ± 0.5 | 13.3 ± 0.0 a,b,c,d | 74.4 ± 0.2 | 4.2 ± 0.1 a,b,c,d,e | 95.7 ± 0.9 f,g,h | 1232 ± 96 e | 2350 ± 177 f |
| AHM_Wh | 90.0 ± 2.1 | 6.4 ± 0.1 c,d | 79.8 ± 0.4 | 13.8 ± 0.5 a,b,c,d | 71.9 ± 1.8 | 3.0 ± 2.5 a,b,c,d | 94.3 ± 2.8 d,e,f.g.h | 874 ± 120 b,c,d | 1669 ± 130 d,e |
| NEAA (%) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| FPH | Asp | Ser | Glu | Ala | Gly | Cys | Tyr | Arg | Pro | Hyp |
| BW_Sb | 9.85 ± 0.071 c,d,e | 5.38 ± 0.184 b,c,d,e | 15.01 ± 0.438 c,d,e | 6.35 ± 0.007 a | 7.18 ± 0.248 a,b | 0.39 ± 0.064 a,b,c | 3.72 ± 0.212 d,e,f | 7.01 ± 0.106 c | 4.37 ± 0.438 a,b,c | 4.14 ± 0.304 c,d |
| RS_Sb | 9.84 ± 0.233 c,d,e | 5.12 ± 0.262 b,c,d | 15.32 ± 0.410 c,d,e | 6.23 ± 0.226 a | 6.99 ± 0.346 a,b | 0.41 ± 0.042 a,b,c,d | 4.13 ± 0.283 f,g | 6.78 ± 0.856 b,c | 4.47 ± 0.587 a,b,c | 3.98 ± 0.156 c,d |
| Po_Sb | 10.13 ± 0.028 d,e | 5.12 ± 0.049 b,c,d | 13.83 ± 0.057 a,b,c,d | 7.68 ± 0.035 c,d | 8.29 ± 0.113 b | 0.36 ± 0.007 a,b | 3.13 ± 0 b,c,d,e | 6.095 ± 0.021 a,b,c | 4.22 ± 0.014 a,b | 4.42 ± 0.283 d |
| Gu_Sb | 9.95 ± 0.057 c,d,e | 5.17 ± 0.354 b,c,d,e | 15.34 ± 1.789 c,d,e | 6.64 ± 0.481 a,b | 7.44 ± 1.322 a,b | 0.39 ± 0.007 a,b,c | 3.42 ± 0.078 b,c,d,e,f | 6.23 ± 0.141 a,b,c | 4.92 ± 0.014 b,c,d,e | 4.08 ± 0.304 c,d |
| M_Sb | 9.88 ± 0.141 c,d,e | 5.26 ± 0.474 b,c,d,e | 16.33 ± 0.481 e | 6.50 ± 0.290 a,b | 6.73 ± 0.325 a,b | 0.36 ± 0.028 a,b | 3.77 ± 0.417 e,f | 6.31 ± 0.191 a,b,c | 4.89 ± 0.021 b,c,d,e | 4.28 ± 0.028 c,d |
| AHM_Sb | 9.45 ± 0.233 b,c | 5.34 ± 0.106 b,c,d,e | 14.12 ± 0.276 a,b,c,d | 6.57±0.035 a,b | 6.73 ± 0.163 a,b | 0.45 ± 0.000 b,c,d | 4.78 ± 0.566 g | 6.76 ± 0.226 b,c | 4.62 ± 0.092 a,b,c,d | 3.50 ± 0.304 a,b,c,d |
| BW_He | 9.77 ± 0.035 b,c,d,e | 5.81 ± 0.071 e | 14.45 ± 0.170 b,c,d,e | 8.25 ± 0.240 d,e,f | 10.88 ± 0.219 c | 0.35 ± 0.021 a,b | 2.87 ± 0.021 a,b,c | 6.25 ± 0.035 a,b,c | 5.43 ± 0.007 c,d,e | 1.99 ± 0.226 a |
| RS_He | 9.13 ± 0.021 b | 5.38 ± 0.078 b,c,d,e | 12.88 ± 0.191 a,b | 8.21 ± 0.035 d,e,f | 11.64 ± 0.049 c | 0.27 ± 0.007 a | 2.81 ± 0.035 a,b,c | 6.52 ± 0.078 b,c | 6.45 ± 0.148 f | 2.21 ± 0.198 a,b |
| Po_He | 9.64 ± 0.035 b,c,d | 5.50 ± 0.064 c,d,e | 13.50 ± 0.007 a,b,c | 8.47 ± 0.134 d,e,f | 12.36 ± 0.255 c | 0.32 ± 0.007 a,b | 2.76 ± 0.007 a,b,c | 6.63 ± 0.064 b,c | 6.31 ± 0.057 f | 2.78 ± 0.113 a,b,c |
| Gu_He | 9.61 ± 0.057 b,c,d | 5.15 ± 0.007 b,c,d,e | 12.55 ± 0.127 a,b | 8.10 ± 0.035 d,e | 11.46 ± 0.113 c | 0.33 ± 0.000 a,b | 2.64 ± 0.056 a,b | 5.85 ± 0.127 a,b | 5.11 ± 0.212 b,c,d,e | 9.41 ± 0.926 e |
| M_He | 9.12 ± 0.361 b | 5.86 ± 0.078 e | 12.49 ± 0.007 a,b | 8.85 ± 0.071 e,f | 15.05 ± 0.057 d | 0.26 ± 0.014 a | 2.29 ± 0.021 a | 6.66 ± 0.304 b,c | 6.24 ± 0.34 f | 3.24 ± 0.276 a,b,c,d |
| AHM_He | 7.98 ± 0.156 a | 5.85 ± 0.049 e | 12.36 ± 0.240 a | 9.01 ± 0.106 f | 15.94 ± 0.233 d | 0.29 ± 0.042 a | 2.81 ± 0.085 a,b,c | 6.47 ± 0.198 b,c | 5.93 ± 0.099 e,f | 3.60 ± 0.304 b,c,d |
| BW_Wh | 10.39 ± 0.064 e | 5.13 ± 0.092 b,c,d | 14.9 ± 0.198 c,d,e | 7.26 ± 0.099 b,c | 5.96 ± 0.127 a | 0.54 ± 0.014 d,e | 3.57 ± 0.113 c,d,e,f | 6.00 ± 0.057 a,b | 3.72 ± 0.205 a | 3.04 ± 0.375 a,b,c,d |
| RS_Wh | 9.86 ± 0.071 c,d,e | 4.82 ± 0.127 a,b,c | 13.39 ± 0.354 a,b,c | 6.98 ± 0.035 a,b,c | 5.78 ± 0.106 a | 0.61 ± 0.028 f | 3.47 ± 0.042 c,d,e,f | 5.41 ± 0.071 a | 3.68 ± 0.156 a | 3.14 ± 0.184 a,b,c,d |
| Po_Wh | 10.23 ± 0.049 d,e | 5.02 ± 0.014 b,c | 15.81 ± 0.268 d,e | 7.25 ± 0.042 b,c | 5.92 ± 0.035 a | 0.41 ± 0.014 a,b,c,d | 2.94 ± 0.021 a,b,c,d | 6.37 ± 0.042 a,b,c | 4.48 ± 0.028 a,b,c | 2.94 ± 0.212 a,b,c,d |
| Gu_Wh | 10.02 ± 0.014 c,d,e | 4.77 ± 0.042 a,b | 13.87 ± 0.071 a,b,c,d | 7.17 ± 0.085 b,c | 6.17 ± 0.014 a | 0.55 ± 0.021 d,e | 3.56 ± 0.049 c,d,e,f | 6.14 ± 0.057 a,b,c | 4.07 ± 0.191 a,b | 3.305 ± 0.205 kb,c,d |
| M_Wh | 9.38 ± 0.375 b,c | 4.31 ± 0.134 a | 13.74 ± 0.375 a,b,c | 6.96 ± 0.247 a,b,c | 8.63 ± 1.386 a | 0.85 ± 0.120 e | 3.42 ± 0.297 b,c,d,e,f | 6.71 ± 0.233 b,c | 5.63 ± 0.559 d,e,f | 2.97 ± 0.148 a,b,c,d |
| AHM_Wh | 10.01 ± 0.262 c,d,e | 4.955 ± 0.177 a,b,c | 13.84 ± 0.368 a,b,c,d | 6.92 ± 0.424 a,b,c | 6.27 ± 0.078 a | 0.53 ± 0.028 c,d,e | 3.38 ± 0.035 b,c,d,e,f | 6.29 ± 0.163 a,b,c | 4.19 ± 0.304 a,b | 3.68 ± 0.891 b,c,d |
| FPH | EAA (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Thr | Val | Met | Ile | Leu | Phe | His | Lys | TEAA/TAA | |
| BW_Sb | 4.16 ± 0.099 a,b,c | 4.20 ± 0.028 d,e | 3.56 ± 0.057 c,d | 3.37 ± 0.537 a.b.c.d | 7.10 ± 0.079 b,c | 4.16 ± 0.127 a,b,c | 2.22 ± 0.297 a,b,c | 7.82 ± 0.042 f,g,h | 36.59 ± 1.068 |
| RS_Sb | 4.02 ± 0.283 a,b,c | 4.23 ± 0.113 d,e | 3.69 ± 0.127 c,d | 3.40 ± 0.417 a,b,c,d | 7.14 ± 0.049 b,c,d,e | 4.49 ± 0.587 b,c | 2.14 ± 0.495 a,b,c | 7.59 ± 0.071 e,f,g | 36.69 ± 0.742 |
| Po_Sb | 4.27 ± 0.042 b,c,d | 4.21 ± 0.021 d,e | 3.66 ± 0.014 c,d | 3.33 ± 0.021 a,b,c,d | 7.41 ± 0.021 c,d,e,f | 4.79 ± 0.028 c | 1.95 ± 0.007 a,b | 7.09 ± 0.042 c,d,e,f | 36.69 e ± 0.156 |
| Gu_Sb | 4.37 ± 0.163 b,c,d | 4.21 ± 0.283 d,e | 3.27 ± 0.085 b,c,d | 3.94 ± 0.764 c,d | 7.59 ± 0.205 d,e,f,g | 3.86 ± 0.495 a,b,c | 2.07 ± 0.403 a,b,c | 7.04 ± 0.325 b,c,d,e | 36.335 ± 0.389 |
| M_Sb | 4.29 ± 0.042 b,c,d | 4.12 ± 0.141 d,e | 3.47 ± 0.361 c,d | 3.80 ± 0.969 c,d | 7.52 ± 0.028 c,d,ef,g | 3.26 ± 0.361 a | 2.23 ± 0.127 a,b,c | 7.03 ± 0.311 b,c,d,e | 35.71 ± 0.318 |
| AHM_Sb | 4.29 ± 0.057 b,c,d | 4.39 ± 0.071 d,e | 3.93 ± 0.460 d | 3.53 ± 0.255 b,c,d | 7.12 ± 0.247 b,c,d | 4.38 ± 0.226 b,c | 2.57 ± 0.049 b,c,d,e | 7.48 ± 0.368 d,e,f | 37.68 ± 0.813 |
| BW_He | 4.45 ± 0.134 b,c,d | 3.88 ± 0.071 b,c,d,e | 3.22 ± 0.255 b,c,d | 2.96 ± 0.085 a,b,c,d | 6.76 ± 0.035 b | 3.97 ± 0.120 a,b,c | 1.95 ± 0.000 a,b | 6.77 ± 0.092 a,b,c,d | 33.94 ± 0.368 |
| RS_He | 4.36 ± 0.057 b,c,d | 4.03 ± 0.014 c,d,e | 3.05 ± 0.078 b,c | 3.13 ± 0.014 a,b,c,d | 6.83 ± 0.049 b | 3.86 ± 0.134 a,b,c | 2.72 ± 0.184 c,d,e | 6.56 ± 0.050 a,b,c | 34.52 ± 0.184 |
| Po_He | 4.18 ± 0.042 a,b,c | 3.27 ± 0.000 a,b | 3.24 ± 0.092 b,c,d | 2.42 ± 0.035 a,b,c | 6.13 ± 0.064 a | 3.98 ± 0.021 a,b,c | 1.80 ± 0.000 a | 6.73 ± 0.042 a,b,c,d | 31.73 ± 0.226 |
| Gu_He | 3.47 ± 0.078 a | 2.93 ± 0.035 a | 3.31 ± 0.085 b,c,d | 2.06 ± 0.049 a,b | 5.87 ± 0.042 a | 4.00 ± 0.021 a,b,c | 1.93 ± 0.014 a,b | 6.21 ± 0.240 a,b | 29.76 ± 0.410 |
| M_He | 4.02 ± 0.078 a,b,c | 3.30 ± 0.042 a,b,c | 3.30 ± 0.191 b,c,d | 2.06 ± 0.120 a,b | 5.72 ± 0.071 a | 3.70 ± 0.184 a,b | 1.91 ± 0.035 a,b | 5.93 ± 0.233 a | 29.92 ± 0.728 |
| AHM_He | 4.0 ± 0.014 a,b | 3.67 ± 0.269 b,c,d | 2.12 ± 0.057 a | 1.99 ± 0.092 a | 5.68 ± 0.064 a | 4.05 ± 0.290 a,b,c | 2.29 ± 0.177 a,b,c,d | 5.94 ± 0.346 a | 29.73 ± 0.587 |
| BW_Wh | 4.40 ± 0.113 b,c,d | 4.52 ± 0.014 e,f | 3.63 ± 0.106 c,d | 3.72 ± 0.049 c,d | 8.36 ± 0.049 h | 4.79 ± 0.134 c | 2.02 ± 0.049 a,b,c | 8.52 ± 0.078 h,i | 39.93 ± 0.226 |
| RS_Wh | 4.73 ± 0.092 c,d | 5.25 ± 0.049 f | 3.57 ± 0.120 c,d | 4.32 ± 0.028 d | 8.34 ± 0.049 h | 4.58 ± 0.163 b,c | 4.34 ± 0.092 f | 7.72 ± 0.092 f,g,h | 42.82 ± 0.021 |
| Po_Wh | 4.99 ± 0.049 d | 4.00 ± 0.057 b,c,d,e | 2.61 ± 0.049 a,b | 3.71 ± 0.078 c,d | 7.89 ± 0.042 g,h | 4.12 ± 0.042 a,b,c | 2.62 ± 0.01 b,c,d,e | 8.86 ± 0.035 i | 38.78 ± 0.184 |
| Gu_Wh | 4.45 ± 0.120 b,c,d | 4.50 ± 0.078 e | 3.54 ± 0.134 c,d | 3.75 ± 0.099 c,d | 7.97 ± 0.021 g,h | 4.66 ± 0.106 b,c | 3.23 ± 0.071 e | 8.39 ± 0.057 g,h,i | 40.465 ± 0.686 |
| M_Wh | 4.62 ± 0.141 b,c,d | 4.16 ± 0.071 d,e | 3.06 ± 0.184 b,c | 4.01 ± 0.113 d | 7.60 ± 0.276 e,f,g | 4.55 ± 0.163 b,c | 2.39 ± 0.212 a,b,c,d | 7.19 ± 0.445 c,d,e,f | 37.57 ± 0.346 |
| AHM_Wh | 4.49 ± 0.594 b,c,d | 4.22 ± 0.629 d,e | 3.21 ± 0.403 b,c,d | 3.45 ± 0.707 a,b,c,d | 7.65 ± 0.198 f,g | 4.49 ± 0.269 b,c | 2.99 ± 0.014 d,e | 8.68 ± 0.113 i | 39.17 ± 0.608 |
| FPH | DPPH | ABTS | Reducing Power | Cu2+ | Fe2+ |
|---|---|---|---|---|---|
| % Inhibition (3 mg/mL) | EC50 (mg/mL) | A0.5 (mg/mL) | EC50 (mg/mL) | ||
| BW_He | 18.91 ± 0.119 h | 4.00 ± 0.057 h | 3.39 ± 0.034 b | 5.66 ± 0.095 h | 0.38 ± 0.009 d,e,f |
| M_He | 4.46 ± 0.536 b | 2.17 ± 0.055 b | 6.35 ± 0.037 m | 3.13 ± 0.095 b,c,d,e | 0.35 ± 0.023 b,c,d,e,f |
| RS_He | 31.87 ± 0.655 j | 2.45 ± 0.059 b,c,d | 4.58 ± 0.131 j | 3.73 ± 0.057 f | 0.49 ± 0.095 g,h |
| Po_He | 14.04 ± 0.335 g | 4.01 ± 0.066 h | 3.98 ± 0.013 f,g | 3.47 ± 0.061 d,e,f | 0.32 ± 0.040 a,b,c,d,e |
| Gu_He | 8.65 ± 0.549 d,e | 1.12 ± 0.026 a | 3.54 ± 0.029 b,c | 3.41 ± 0.025 d,e,f | 0.26 ± 0.002 a |
| AHM_He | 29.22 ± 0.067 j | 4.89 ± 0.091 i | 3.19 ± 0.057 a | 2.49 ± 0.019 a | 0.53 ± 0.012 h |
| BW_Sb | 2.83 ± 0.293 a,b | 4.76 ± 0.571 i | 4.07 ± 0.011 g,h | 5.20 ± 0.051 h | 0.28 ± 0.011 a,b |
| M_Sb | NA | 2.95 ± 0.132 e,f | 5.79 ± 0.031 l | 3.40 ± 0.060 c,d,e,f | 0.26 ± 0.005 a |
| RS_Sb | NA | 2.72 ± 0.079 d,e | 4.57 ± 0.022 j | 3.32 ± 0.021 c,d,e,f | 0.28 ± 0.004 a,b |
| Po_Sb | 14.75 ± 0.170 g | 3.68 ± 0.008 g,h | 4.22 ± 0.149 h,i | 3.34 ± 0.021 c,d,e,f | 0.27 ± 0.004 a |
| Gu_Sb | 0.76 ± 0.340 a | 2.22 ± 0.073 b,c | 4.37 ± 0.003 i | 3.66 ± 0.025 e,f | 0.31 ± 0.019 a,b,c,d |
| AHM_Sb | 25.10 ± 0.829 i | 4.93 ± 0.018 i | 3.51 ± 0.009 b,c | 3.22 ± 0.031 c,d,e,f | 0.40 ± 0.002 e,f |
| BW_Wh | 10.80 ± 1.041 e,f | 3.30 ± 0.037 f,g | 3.56 ± 0.014 b,c | 4.67 ± 0.031 g | 0.36 ± 0.011 a,d,e,f |
| M_Wh | 12.29 ± 0.382 c,d | 2.67 ± 0.131 c,d,e | 5.10 ± 0.010 k | 3.07 ± 0.028 b,c,d | 0.27 ± 0.008 a,b |
| RS_Wh | 12.29 ± 0.382 f,g | 2.24 ± 0.094 b,c | 4.97 ± 0.010 k | 2.75 ± 0.022 a,b,c | 0.43 ± 0.015 f,g |
| Po_Wh | 18.26 ± 0.130 h | 3.81 ± 0.024 h | 3.84 ± 0.049 e,f | 3.14 ± 0.035 b,c,d,e,f | 0.31 ± 0.017 a,b,c,d |
| Gu_Wh | 5.22 ± 1.147 b,c | 1.47 ± 0.020 a | 3.65 ± 0.018 c,d | 3.37 ± 0.037 c,d,e,f | 0.38 ± 0.014 c,d,e,f |
| AHM_Wh | 18.44 ± 0.850 h | 4.56 ± 0.039 i | 3.75 ± 0.011 d,e | 2.67 ± 0.023 a,b | 0.29 ± 0.010 a,b,c |
| FPH | IC50 (mg/mL) | % Inhibition (5 mg/mL) | |
|---|---|---|---|
| α-Amylase | α-Glucosidase | ACE | |
| BW_He | 5.70 ± 0.67 a | --- | 61.20 ± 7.83 a |
| M_He | 18.49 ± 1.90 b,c,d | 216.9 ± 26.3 g,h,i | 76.58 ± 2.72 a,b,c |
| RS_He | --- | 155.5 ± 4.1 d,e,f,g | 64.22 ± 2.46 a,b |
| Po_He | 60.29 ± 9.46 g | 154.4 ± 3.5 d,e,f,g | 79.35 ± 17.01 a,b,c |
| Gu_He | 17.42 ± 0.49 a,b,c,d | 247.8 ± 19.9 h,i,j | 69.28 ± 5.10 a,b,c |
| AHM_He | 24.96 ± 6.73 c,d,e | 83.1 ± 4.0 b,c | 60.77 ± 2.44 a |
| BW_Sb | 13.96 ± 0.71 a,b | --- | 77.68 ± 2.15 a,b,c |
| M_Sb | 33.66 ± 2.93 e,f | 103.7 ± 15.2 b,c,d | 79.40 ± 2.01 a,b,c |
| RS_Sb | 27.14 ± 0.96 c,d,e | 21.8 ± 8.9 a | 77.79 ± 1.06 a,b,c |
| Po_Sb | 33.27 ± 1.04 e | 133.7 ± 2.9 a,c,d,e | 82.10 ± 2.72 b,c |
| Gu_Sb | 27.82 ± 9.75 c,d,e | 214.5 ± 9.0 g,h,i | 82.41 ± 2.19 b,c |
| AHM_Sb | 29.33 ± 2.34 d,e | 75.1 ± 1.5 b | 70.71 ± 3.09 a,b,c |
| BW_Wh | 18.51 ± 1.53 b,c,d | 156.9 ± 7.2 d,e,f,g | 82.69 ± 12.81 b,c |
| M_Wh | 44.06 ± 2.65 f | 300.0 ± 25.8 j | 85.95 ± 0.56 c |
| RS_Wh | 23.50 ± 1.49 b,c,d,e | 256.7 ± 58.1 i,j | 77.07 ± 1.02 a,b,c |
| Po_Wh | --- | 187.7 ± 36.1 e,f,g,h | 66.60 ± 6.22 a,b,c |
| Gu_Wh | 16.71 ± 0.25 a,b,c | 203.5 ± 14.8 f,g,h,i | 72.78 ± 5.57 a,b,c |
| AHM_Wh | 84.37 ± 5.21 h | 143.3 ± 0.8 c,d,e,f | 75.58 ± 4.48 a,b,c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Henriques, A.; Vázquez, J.A.; Valcarcel, J.; Mendes, R.; Bandarra, N.M.; Pires, C. Characterization of Protein Hydrolysates from Fish Discards and By-Products from the North-West Spain Fishing Fleet as Potential Sources of Bioactive Peptides. Mar. Drugs 2021, 19, 338. https://doi.org/10.3390/md19060338
Henriques A, Vázquez JA, Valcarcel J, Mendes R, Bandarra NM, Pires C. Characterization of Protein Hydrolysates from Fish Discards and By-Products from the North-West Spain Fishing Fleet as Potential Sources of Bioactive Peptides. Marine Drugs. 2021; 19(6):338. https://doi.org/10.3390/md19060338
Chicago/Turabian StyleHenriques, Andreia, José A. Vázquez, Jesus Valcarcel, Rogério Mendes, Narcisa M. Bandarra, and Carla Pires. 2021. "Characterization of Protein Hydrolysates from Fish Discards and By-Products from the North-West Spain Fishing Fleet as Potential Sources of Bioactive Peptides" Marine Drugs 19, no. 6: 338. https://doi.org/10.3390/md19060338
APA StyleHenriques, A., Vázquez, J. A., Valcarcel, J., Mendes, R., Bandarra, N. M., & Pires, C. (2021). Characterization of Protein Hydrolysates from Fish Discards and By-Products from the North-West Spain Fishing Fleet as Potential Sources of Bioactive Peptides. Marine Drugs, 19(6), 338. https://doi.org/10.3390/md19060338