
Sphingolipids of Asteroidea and Holothuroidea: Structures and Biological Activities





Abstract
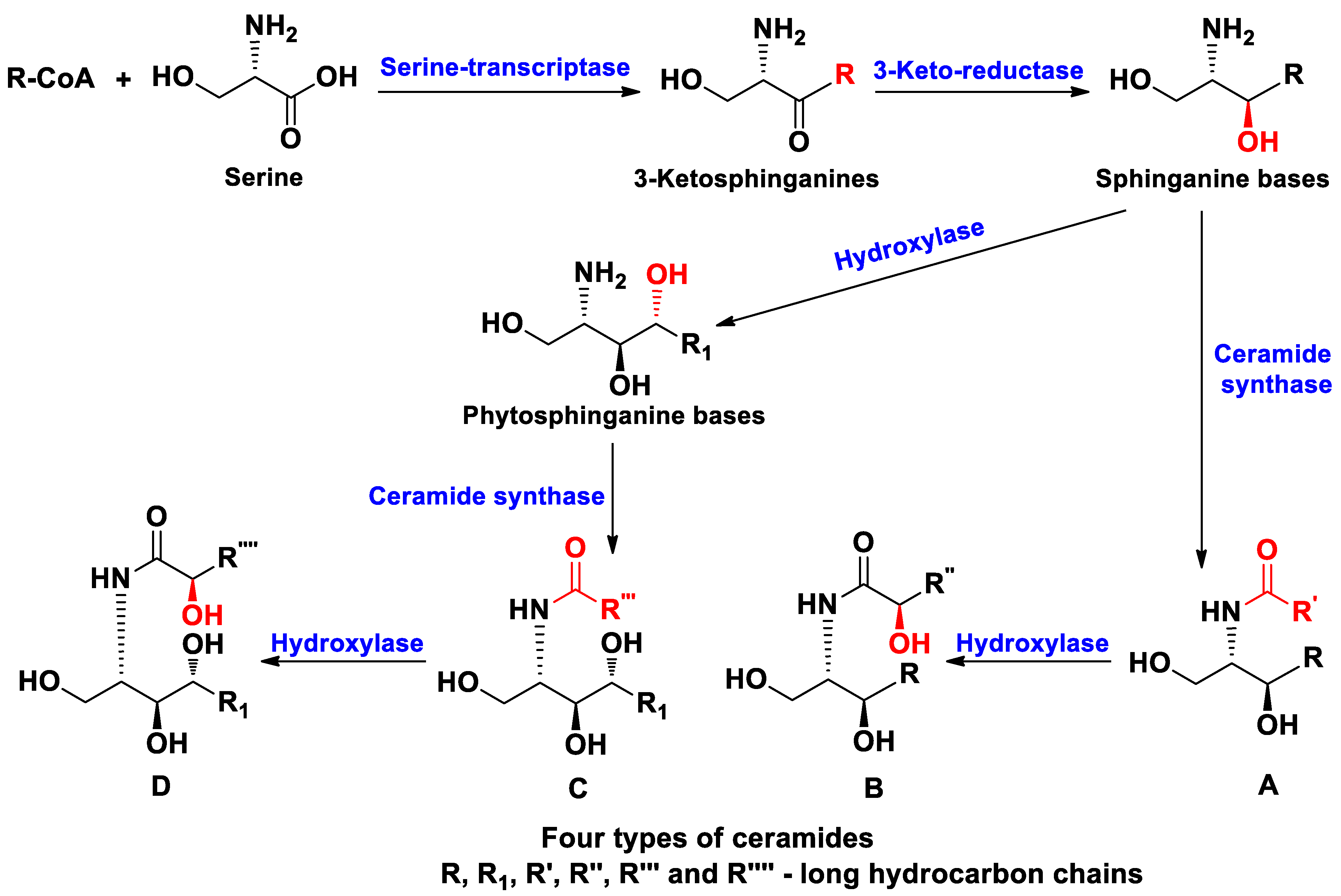
1. Introduction
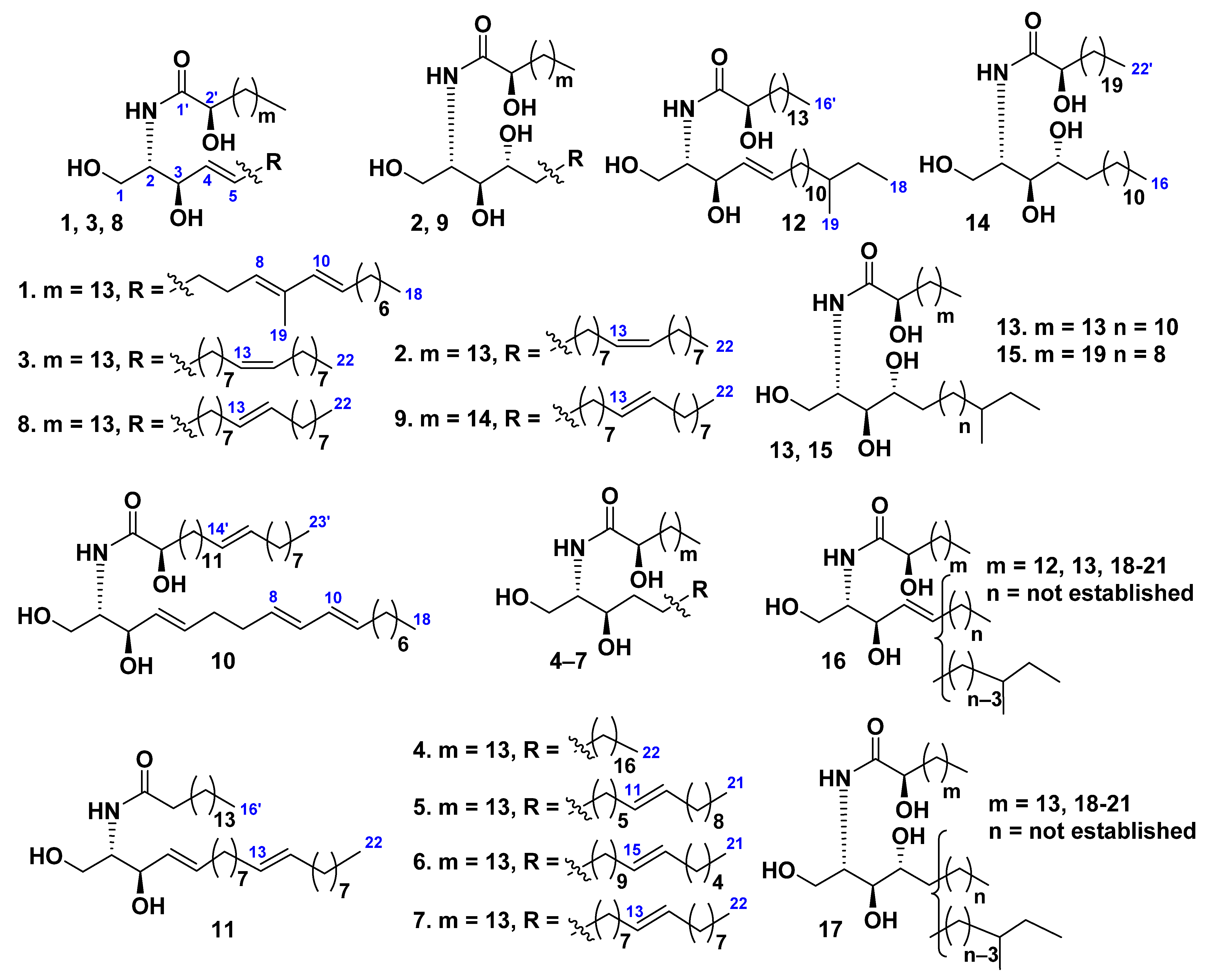
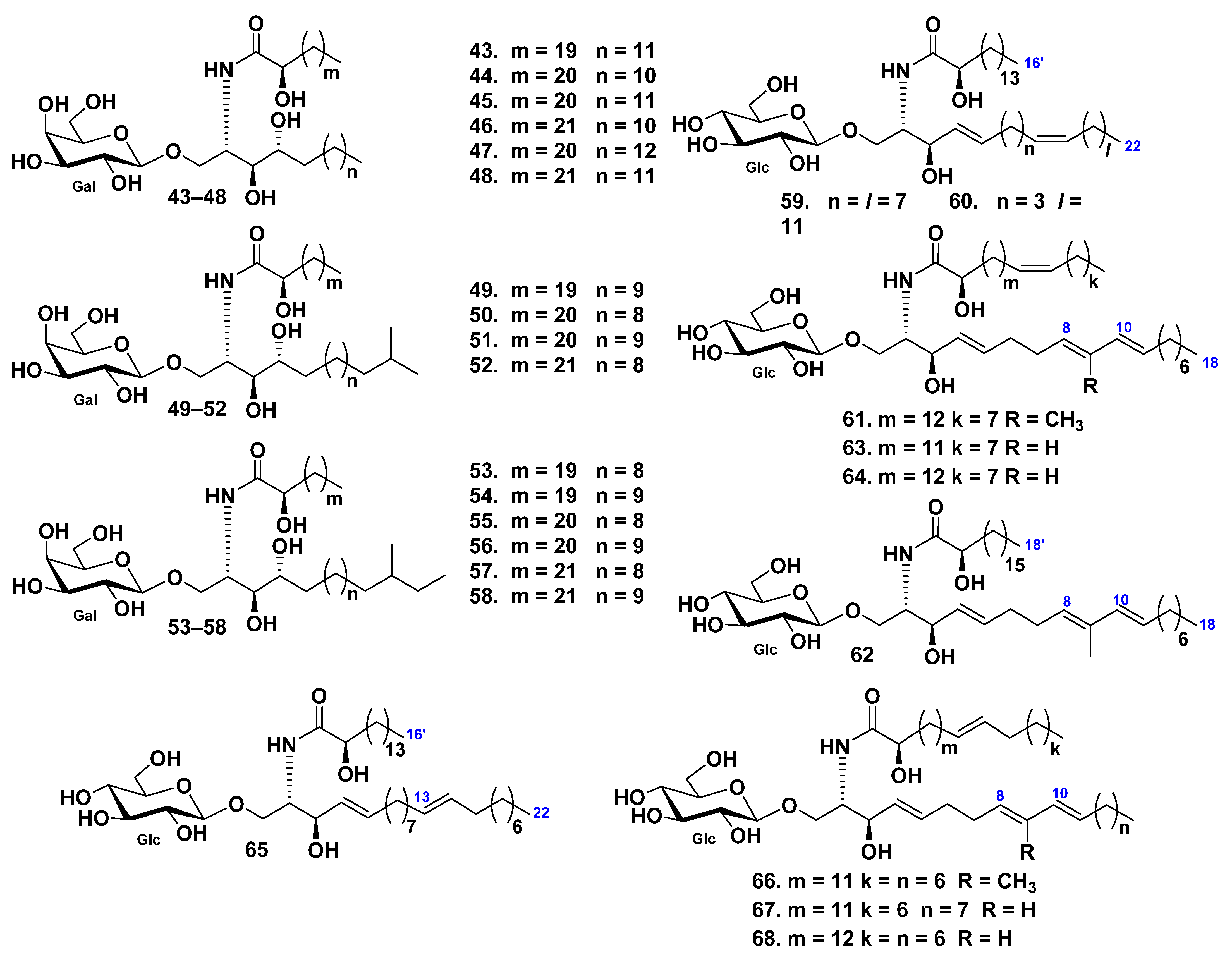
2. Ceramides
Class Asteroidea
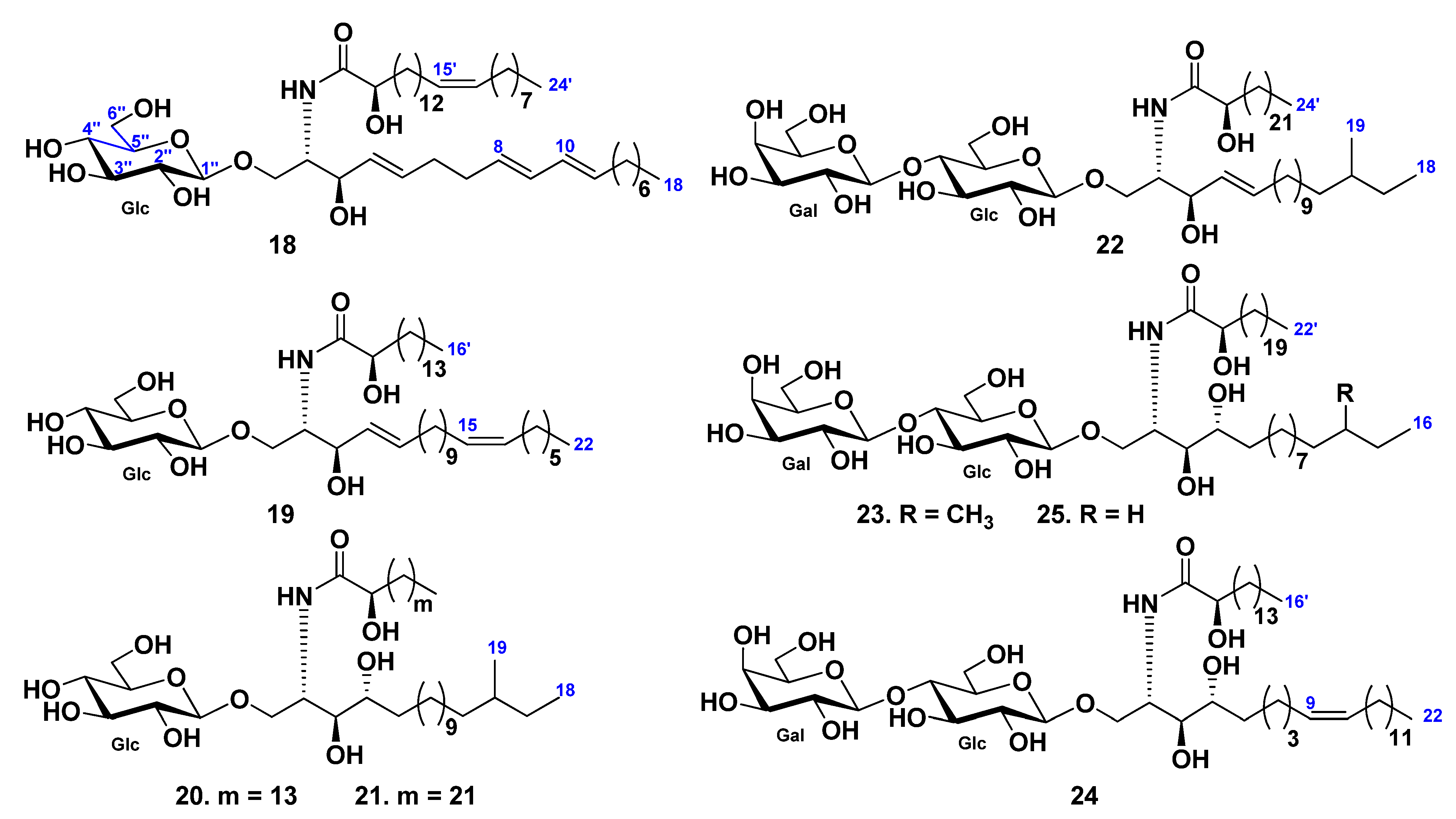
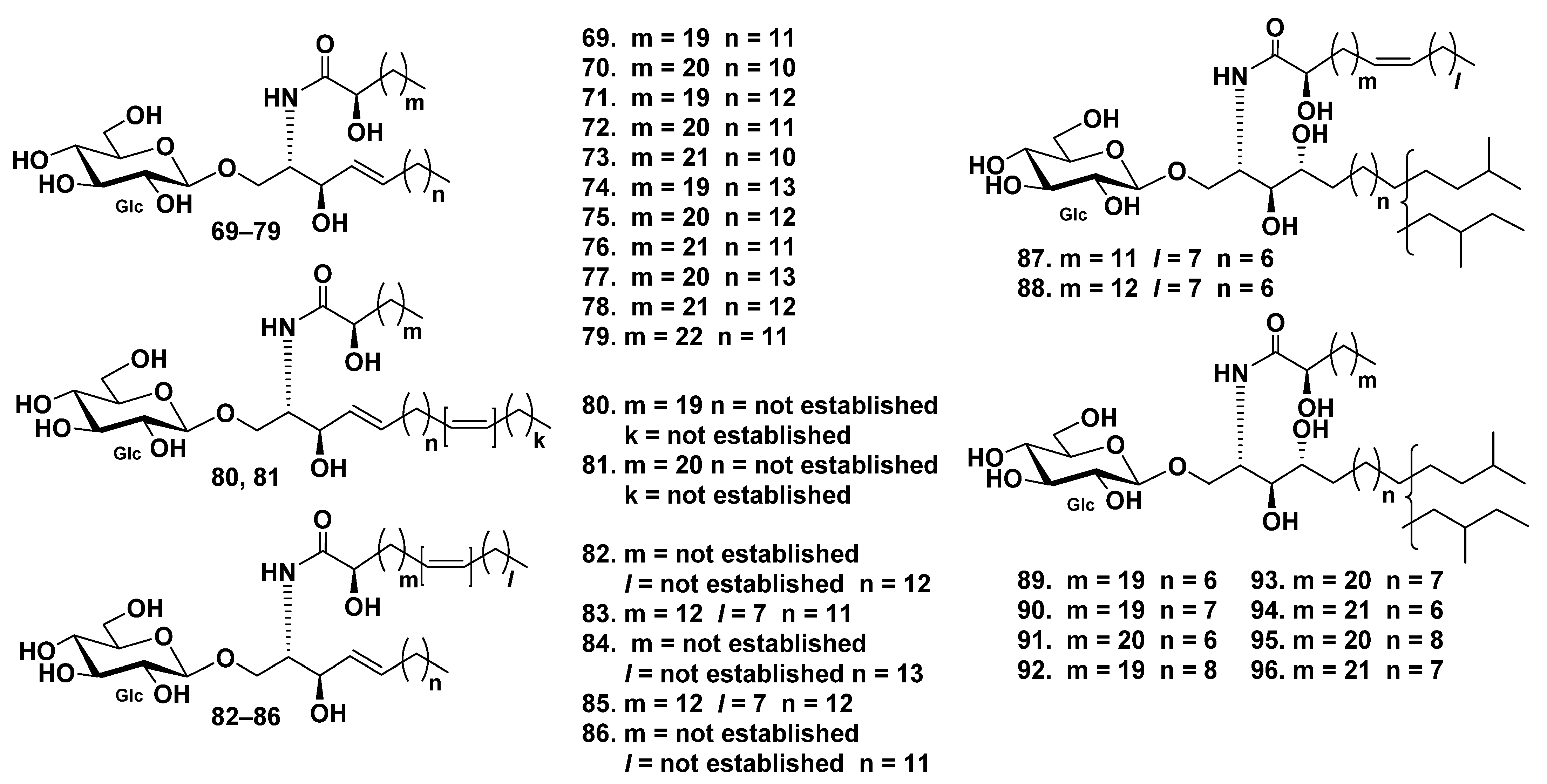
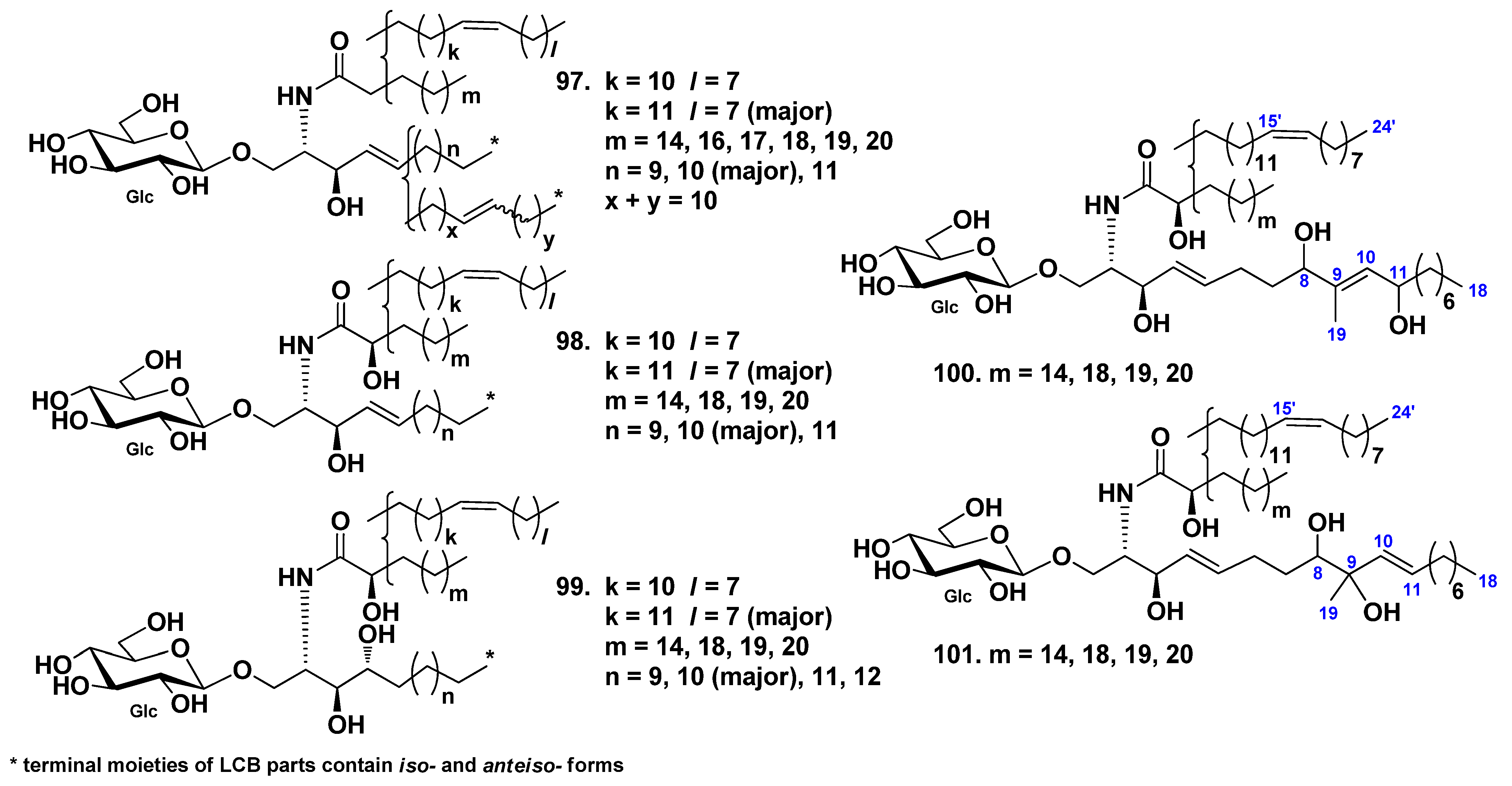
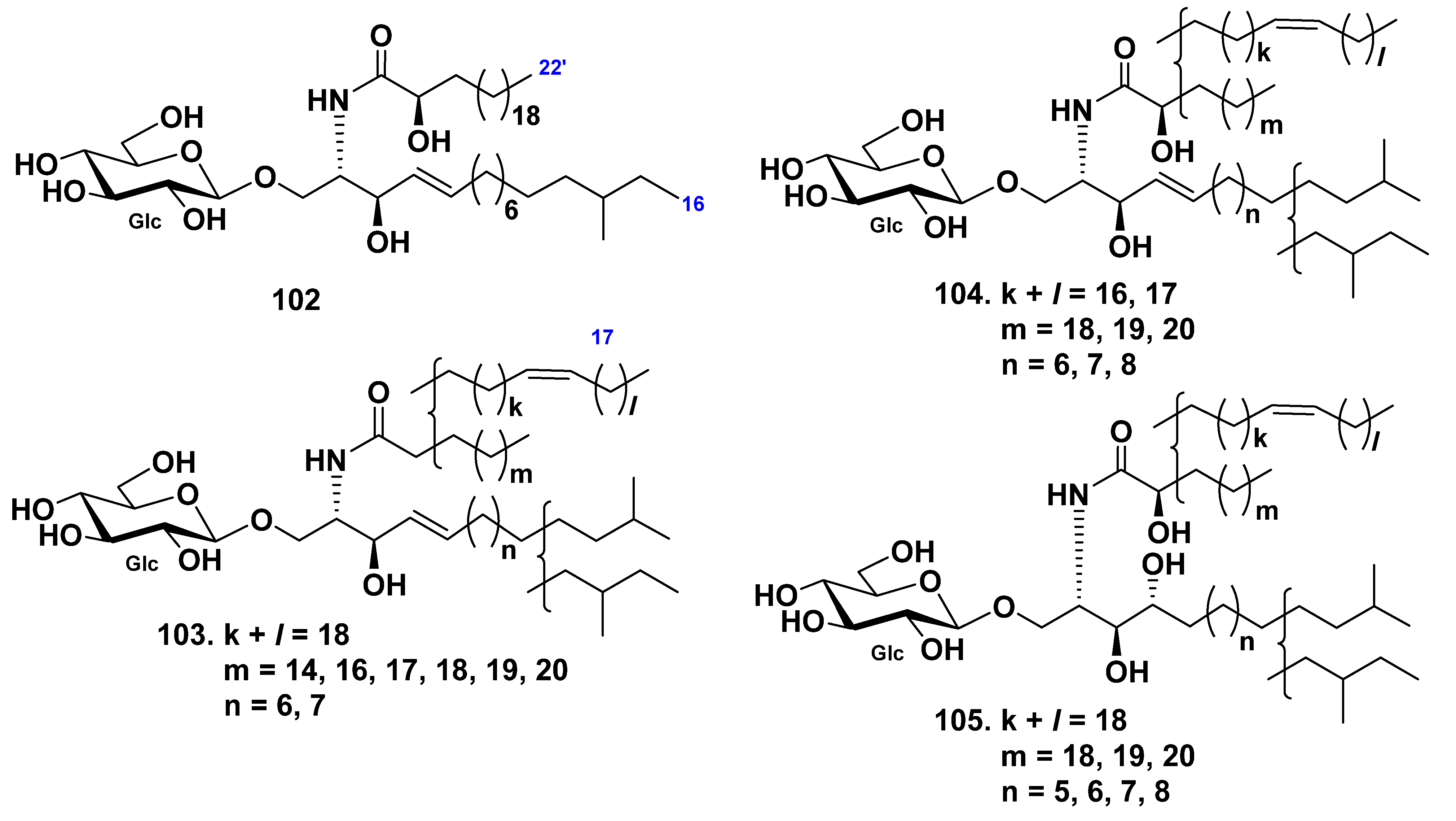
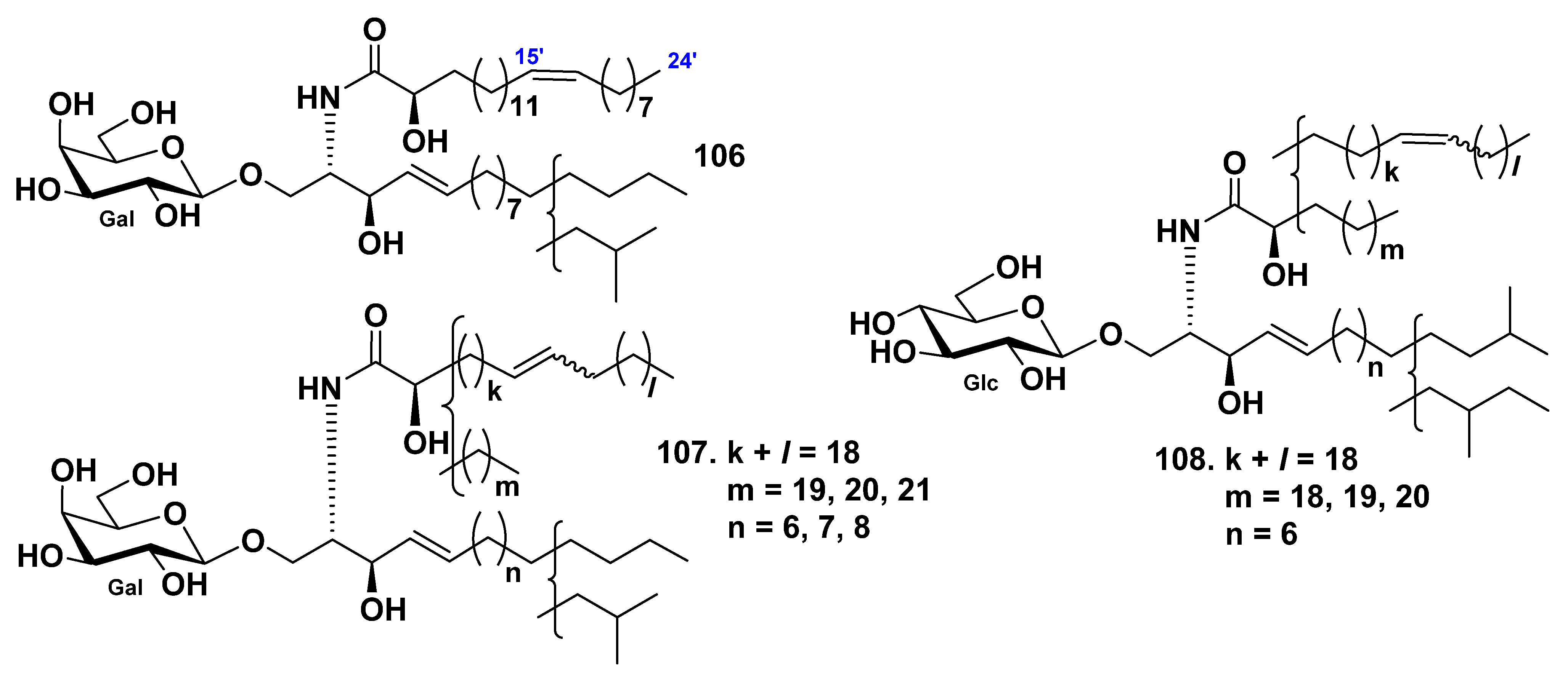
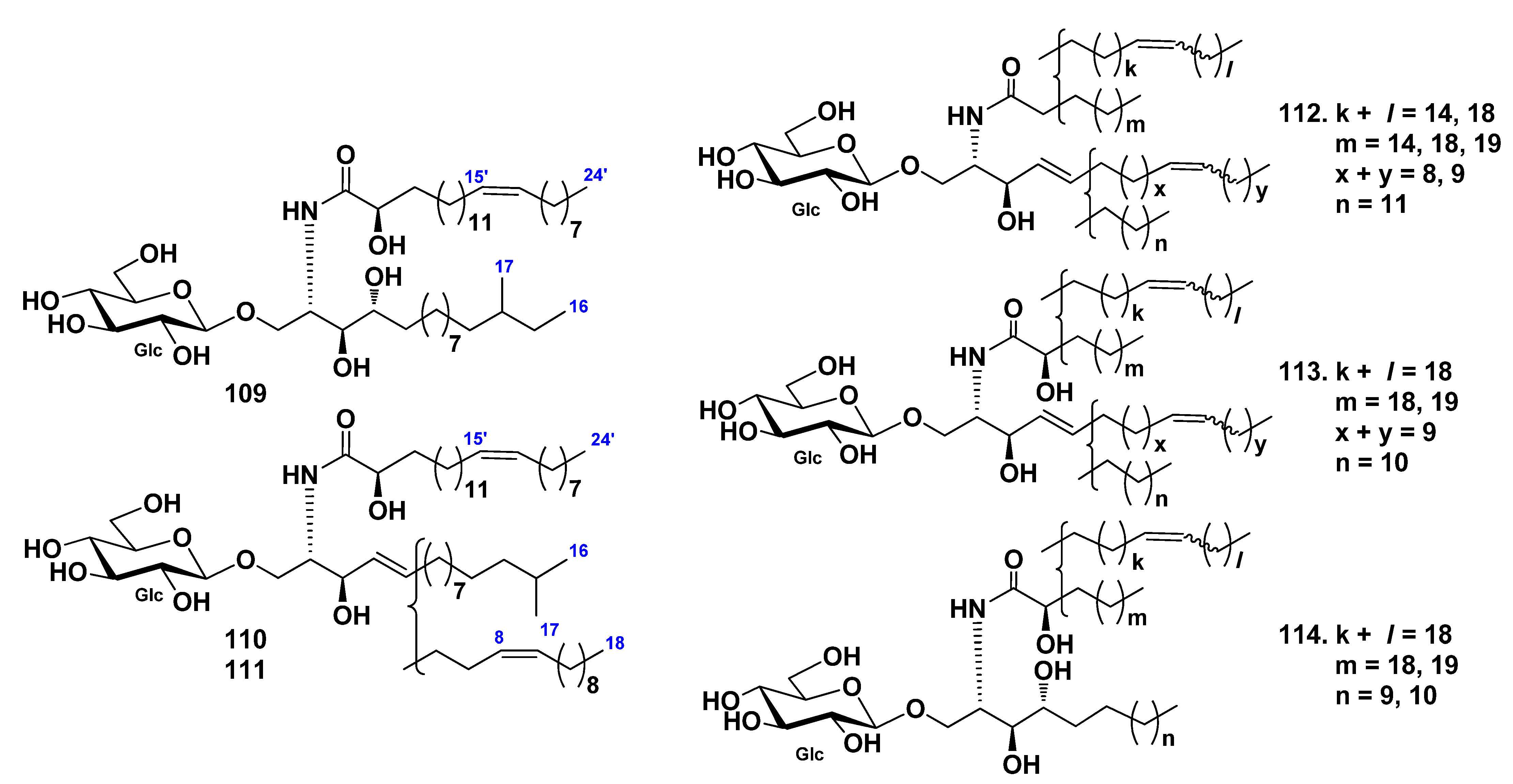
3. Cerebrosides
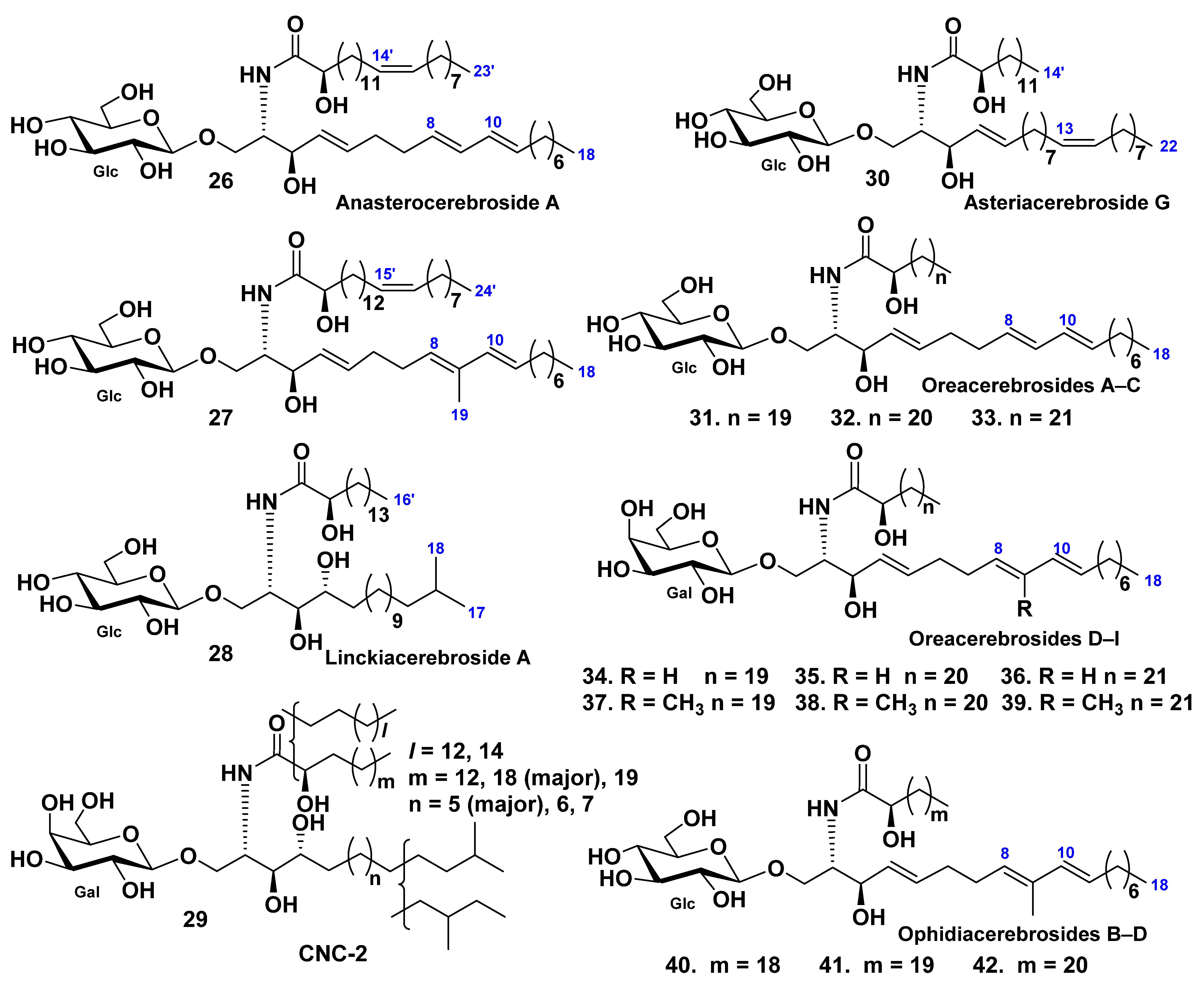
3.1. Class Asteroidea
3.2. Class Holothuroidea
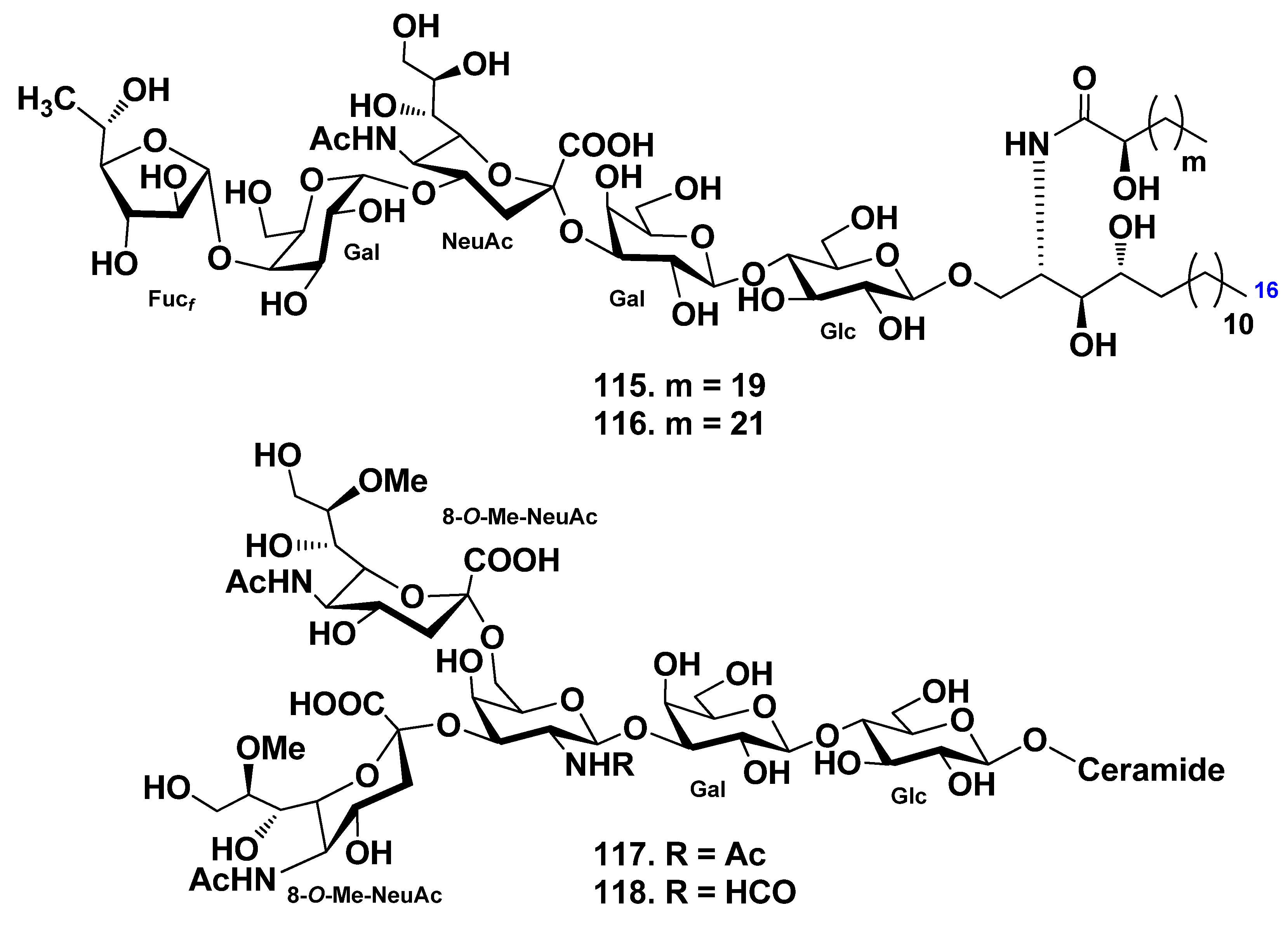
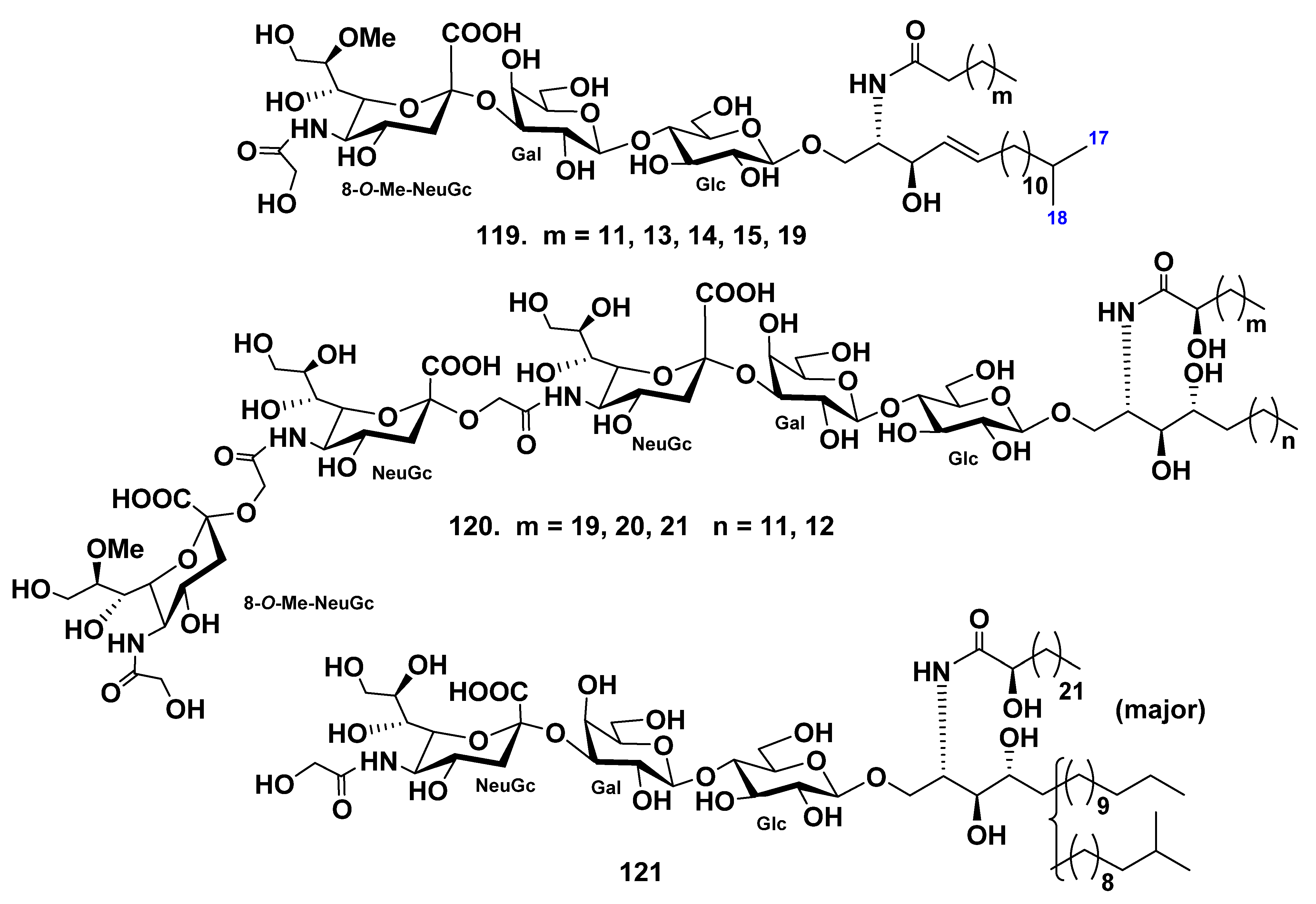
4. Gangliosides
4.1. Class Asteroidea
4.2. Class Holothuroidea
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Stonik, V.A.; Kalinin, V.I.; Avilov, S.A. Toxins from sea cucumbers (holothuroids): Chemical structures, properties, taxonomic distribution, biosynthesis and evolution. J. Nat. Toxins 1999, 8, 235–248. [Google Scholar] [PubMed]
- Stonik, V.A. Marine polar steroids. Russ. Chem. Rev. 2001, 70, 673–715. [Google Scholar] [CrossRef]
- Stonik, V.A.; Ivanchina, N.V.; Kicha, A.A. New polar steroids from starfish. Nat. Prod. Commun. 2008, 3, 1587–1610. [Google Scholar] [CrossRef]
- Ivanchina, N.V.; Kicha, A.A.; Stonik, V.A. Steroid glycosides from marine organisms. Steroids 2011, 76, 425–454. [Google Scholar] [CrossRef]
- Ivanchina, N.V.; Kicha, A.A.; Malyarenko, T.V.; Stonik, V.A. Advances in Natural Products Discovery; Gomes, A.R., Rocha-Santos, T., Duarte, A., Eds.; Nova Science Publishers: New York, NY, USA, 2017; Volume 6, pp. 191–224. [Google Scholar]
- Stonik, V.A.; Kicha, A.A.; Malyarenko, T.V.; Ivanchina, N.V. Asterosaponins: Structures, taxonomic distribution, biogenesis and biological activities. Mar. Drugs 2020, 18, 584. [Google Scholar] [CrossRef] [PubMed]
- Fattorusso, E.; Mangoni, A. Marine glycolipids. Prog. Chem. Org. Nat. Prod. 1997, 72, 215–301. [Google Scholar]
- Popov, R.S.; Ivanchina, N.V.; Kicha, A.A.; Malyarenko, T.V.; Dmitrenok, P.S.; Stonik, V.A. Metabolite profiling of polar steroid constituents in the Far Eastern starfish Aphelasterias japonica using LC–ESI MS/MS. Metabolomics 2014, 10, 1152–1168. [Google Scholar] [CrossRef]
- Popov, R.S.; Ivanchina, N.V.; Kicha, A.A.; Malyarenko, T.V.; Dmitrenok, P.S.; Stonik, V.A. LC-ESI MS/MS profiling of polar steroid metabolites of the Far Eastern starfish Patiria (=Asterina) pectinifera. Metabolomics 2016, 12, 21. [Google Scholar] [CrossRef]
- Popov, R.S.; Ivanchina, N.V.; Kicha, A.A.; Malyarenko, T.V.; Grebnev, B.B.; Dmitrenok, P.S.; Stonik, V.A. LC-MS-based metabolome analysis on steroid metabolites from the starfish Patiria (=Asterina) pectinifera in conditions of active feeding and stresses. Metabolomics 2016, 12, 106. [Google Scholar] [CrossRef]
- Popov, R.S.; Ivanchina, N.V.; Kicha, A.A.; Malyarenko, T.V.; Dmitrenok, P.S. Structural characterization of polar steroid compounds of the Far Eastern starfish Lethasterias fusca by nanoflow liquid chromatography coupled to quadrupole time-of-flight tandem mass spectrometry. J. Am. Soc. Mass Spectrom. 2019, 30, 743–764. [Google Scholar] [CrossRef]
- Popov, R.S.; Ivanchina, N.V.; Kicha, A.A.; Malyarenko, T.V.; Grebnev, B.B.; Stonik, V.A.; Dmitrenok, P.S. The distribution of asterosaponins, polyhydroxysteroids and related glycosides in different body components of the Far Eastern starfish Lethasterias fusca. Mar. Drugs 2019, 17, 523. [Google Scholar] [CrossRef]
- Popov, R.S.; Ivanchina, N.V.; Silchenko, A.S.; Avilov, S.A.; Kalinin, V.I.; Dolmatov, I.Y.; Stonik, V.A.; Dmitrenok, P.S. Metabolite profiling of triterpene glycosides of the Far Eastern sea cucumber Eupentacta fraudatrix and their distribution in various body components using LC-ESI QTOF-MS. Mar. Drugs 2017, 15, 302. [Google Scholar] [CrossRef]
- Degroote, S.; Wolthoorn, J.; van Meer, G. The cell biology of glycosphingolipids. Semin. Cell Dev. Biol. 2004, 15, 375–387. [Google Scholar] [CrossRef]
- Hakomori, S.; Igarashi, Y. Functional role of glycosphingolipids in cell recognition and signaling. J. Biochem. 1995, 118, 1091–1103. [Google Scholar] [CrossRef] [PubMed]
- Fahy, E.; Subramaniam, S.; Brown, H.A.; Glass, C.K.; Merrill, A.H., Jr.; Murphy, R.C.; Raetz, C.R.H.; Russell, D.W.; Seyama, Y.; Shaw, W.; et al. A comprehensive classification system for lipids. J. Lipid Res. 2005, 46, 839–861. [Google Scholar] [CrossRef]
- Chebane, K.; Guyot, M. Occurrence of erythro-docosasphinga-4,8-dienine, as an ester, in Anemonia sulcata. Tetrahedron Lett. 1986, 27, 1495–1496. [Google Scholar] [CrossRef]
- Costantino, V.; Fattorusso, E.; Imperatore, C.; Mangoni, A. Glycolipids from sponges. 13.1 Clarhamnoside, the first rhamnosylated α-galactosylceramide from Agelas clathrodes. Improving spectral strategies for glycoconjugate structure determination. J. Org. Chem. 2004, 69, 1174–1179. [Google Scholar] [CrossRef] [PubMed]
- Mansoor, T.A.; Shinde, P.B.; Luo, X.; Hong, J.; Lee, C.O.; Sim, X.; Son, B.W.; Jung, J.H. Renierosides, cerebrosides from a marine sponge Haliclona (Reniera) sp. J. Nat. Prod. 2007, 70, 1481–1486. [Google Scholar] [CrossRef] [PubMed]
- Costantino, V.; Fattorusso, E.; Imperatore, C.; Mangoni, A.; Freigang, S.; Teyton, L. Corrugoside: A new immunostimulatory α-galactoglycosphingolipid from the marine sponge Axinella corrugata. Bioorg. Med. Chem. 2008, 16, 2077–2085. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.Y.; Wen, Z.H.; Chiou, S.F.; Tsai, C.W.; Wang, S.K.; Hsu, C.H.; Dai, C.F.; Chiang, M.Y.; Wang, W.H.; Duh, C.Y. Ceramide and cerebrosides from the octocoral Sarcophyton ehrenbergi. J. Nat. Prod. 2009, 72, 465–468. [Google Scholar] [CrossRef]
- Durán, R.; Zubia, E.; Ortega, M.J.; Naranjo, S.; Salvá, J. Phallusides, new glucosphingolipids from the ascidian Phallusia fumigata. Tetrahedron 1998, 54, 14597–14602. [Google Scholar] [CrossRef]
- Ishii, T.; Okino, T.; Mino, Y. A ceramide and cerebroside from the starfish Asterias amurensis Lütken and their plant-growth promotion activities. J. Nat. Prod. 2006, 69, 1080–1082. [Google Scholar] [CrossRef]
- Duan, J.; Sugawara, T.; Sakai, S.; Aida, K.; Hirata, T. Oral glucosylceramide reduces 2,4-dinitrofluorobenzene induced inflammatory response in mice by reducing TNF-alpha levels and leukocyte infiltration. Lipids 2011, 46, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.; Sugawara, T.; Hirose, M.; Aida, K.; Sakai, S.; Fujii, A.; Hirata, T. Dietary sphingolipids improve skin barrier functions via the upregulation of ceramide synthases in the epidermis. Exp. Dermatol. 2012, 21, 448–452. [Google Scholar] [CrossRef]
- Dillehay, D.L.; Webb, S.K.; Schmelz, E.M.; Merrill, A.H., Jr. Dietary sphingomyelin inhibits 1,2-dimethylhydrazine-induced colon cancer in CF1 mice. J. Nutr. 1994, 124, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Carmeliet, P. Angiogenesis in health and disease. Nat. Med. 2003, 9, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Muralidhar, P.; Radhika, P.; Krishna, N.; Venkata Rao, D. Bheemasankara Rao, Ch. Sphingolipids from marine organisms: A review. Nat. Prod. Sci. 2003, 9, 117–142. [Google Scholar]
- Higuchi, R.; Inagaki, M.; Yamada, K.; Miyamoto, T. Biologically active gangliosides from echinoderms. J. Nat. Med. 2007, 61, 367–370. [Google Scholar] [CrossRef]
- Yamada, K. Chemo-pharmaceutical studies on the glycosphingolipid constituents from Echinoderm, sea cucumbers, as the medicinal materials. Yakugaku Zasshi 2002, 122, 1133–1143. [Google Scholar] [CrossRef]
- Inagaki, M. Structure and biological activity of glycosphingolipids from starfish and feather star. Yakugaku Zasshi 2008, 128, 1187–1194. [Google Scholar] [CrossRef][Green Version]
- Sperling, P.; Heinz, E. Plant sphingolipids: Structural diversity, biosynthesis, first genes and functions. Biochem. Biophys. Acta 2003, 1632, 1–15. [Google Scholar] [CrossRef]
- Rho, J.-R.; Kim, Y.H. Isolation and structure determination of three new ceramides from the starfish Distolasterias nipon. Bull. Korean Chem. Soc. 2005, 26, 1457–1460. [Google Scholar] [CrossRef]
- Yoo, J.S.; Park, T.; Bang, G.; Lee, C.; Rho, J.R.; Kim, Y.H. High-energy collision-induced dissociation of [M+Na]+ ions desorbed by fast atom bombardment of ceramides isolated from the starfish Distolasterias nipon. J. Mass Spectrom. 2013, 48, 164–171. [Google Scholar] [CrossRef]
- Inagaki, M.; Ikeda, Y.; Kawatake, S.; Nakamura, K.; Tanaka, M.; Misawa, E.; Yamada, M.; Higuchi, R. Isolation and structure of four new ceramides from the starfish Luidia maculata. Chem. Pharm. Bull. 2006, 54, 1647–1649. [Google Scholar] [CrossRef]
- Bibel, D.J.; Aly, R.; Shinefield, H.R. Antimicrobial activity of sphingosines. J. Investig. Dermatol. 1992, 98, 269–273. [Google Scholar] [CrossRef]
- Fischer, C.L.; Drake, D.R.; Dawson, D.V.; Blanchette, D.R.; Brogden, K.A.; Wertz, P.W. Antibacterial activity of sphingoid bases and fatty acids against gram-positive and gram-negative bacteria. Antimicrob. Agents Chemother. 2012, 56, 1157–1161. [Google Scholar] [CrossRef] [PubMed]
- Murshid, S.S.A.; Badr, J.M.; Youssef, D.T.A. Penicillosides A and B: New cerebrosides from the marine-derived fungus Penicillum species. Rev. Bras. Farmacognosia. 2016, 26, 29–33. [Google Scholar] [CrossRef]
- De Vivar, M.E.D.; Seldes, A.M.; Maier, M.S. Two novel glucosylceramides from gonads and body walls of the Patagonian starfish Allostichaster inaequalis. Lipids 2002, 37, 597–603. [Google Scholar] [CrossRef]
- Maier, M.S.; Kuriss, A.; Seldes, A.M. Isolation and structure of glucosylceramides from the starfish Cosmasterias lurida. Lipids 1998, 33, 825–827. [Google Scholar] [CrossRef]
- Kawatake, S.; Nakamura, K.; Inagaki, M.; Higuchi, R. Isolation and structure determination of six glucocerebrosides from the starfish Luidia maculata. Chem. Pharm. Bull. 2002, 50, 1091–1096. [Google Scholar] [CrossRef]
- Inagaki, M.; Nakamura, K.; Kawatake, S.; Higuchi, R. Isolation and structural determination of four new ceramide lactosides from the Starfish Luidia maculata. Eur. J. Org. Chem. 2003, 325–331. [Google Scholar] [CrossRef]
- Chludil, H.D.; Seldes, A.M.; Maier, M.S. Anasterocerebroside A, a new glucosylceramide from the Patagonian starfish Anasterias minuta. Z. Naturforsch. C 2003, 58, 433–440. [Google Scholar] [CrossRef]
- Maruta, T.; Saito, T.; Inagaki, M.; Shibata, O.; Higuchi, R. Biologically active glycosides from Asteroidea, 41. Isolation and structure determination of glucocerebrosides from the starfish Linckia laevigata. Chem. Pharm. Bull. 2005, 53, 1255–1258. [Google Scholar] [CrossRef] [PubMed]
- Inagaki, M.; Nakata, T.; Higuchi, R. Isolation and structure of a galactocerebroside molecular species from the starfish Culcita novaeguineae. Chem. Pharm. Bull. 2006, 54, 260–261. [Google Scholar] [CrossRef] [PubMed]
- Costantino, V.; de Rosa, C.; Fattorusso, E.; Imperatore, C.; Mangoni, A.; Irace, C.; Maffettone, C.; Capasso, D.; Malorni, L.; Palumbo, R.; et al. Oreacerebrosides: Bioactive cerebrosides with a triunsaturated sphingoid base from the sea star Oreaster reticulatus. Eur. J. Org. Chem. 2007, 5277–5283. [Google Scholar] [CrossRef]
- Farokhi, F.; Wielgosz-Collin, G.; Clement, M.; Kornprobst, J.-M.; Barnathan, G. Cytotoxicity on human cancer cells of ophidiacerebrosides isolated from the African starfish Narcissia canariensis. Mar. Drugs 2010, 8, 2988–2998. [Google Scholar] [CrossRef]
- Jin, W.; Rinehart, K.L.; Jares-Erijman, E.A. Ophidiacerebrosides: Cytotoxic glycosphingolipids containing a novel sphingosine from a sea star. J. Org. Chem. 1994, 59, 144–147. [Google Scholar] [CrossRef]
- Pan, K.; Inagaki, M.; Ohno, N.; Tanaka, C.; Higuchi, R.; Miyamoto, T. Identification of sixteen new galactocerebrosides from the starfish Protoreaster nodosus. Chem. Pharm. Bull. 2010, 58, 470–474. [Google Scholar] [CrossRef]
- Park, T.; Park, Y.S.; Rho, J.-R.; Kim, Y.H. Structural determination of cerebrosides isolated from Asterias amurensis starfish eggs using high-energy collision-induced dissociation of sodium-adducted molecules. Rapid Commun. Mass Spectrom. 2011, 25, 572–578. [Google Scholar] [CrossRef]
- Mikami, D.; Sakai, S.; Sasaki, S.; Igarashi, Y. Effects of Asterias amurensis-derived sphingoid bases on the de novo ceramide synthesis in cultured normal human epidermal keratinocytes. J. Oleo Sci. 2016, 65, 671–680. [Google Scholar] [CrossRef]
- Hue, N.; Montagnac, A.; Paїs, M.; Serani, L.; Laprѐvote, O. Structural elucidation of eighteen cerebrosides from Holothuria coronopertusa in a complex mixture by high-energy collision-induced dissociation of [M + Li]+ ions. Eur. J. Mass Spectrom. 2001, 7, 409–417. [Google Scholar] [CrossRef]
- Yamada, K.; Sasaki, K.; Harada, Y.; Isobe, R.; Higuchi, R. Constituents of Holothuroidea. 12. Isolation and structure of glucocerebrosides from the sea cucumber Holothuria pervicax. Chem. Pharm. Bull. 2002, 50, 1467–1470. [Google Scholar] [CrossRef][Green Version]
- Kisa, F.; Yamada, K.; Kaneko, M.; Inagaki, M.; Higuchi, R. Constituents of Holothuroidea, 14. Isolation and structure of new glucocerebroside molecular species from the sea cucumber Stichopus japonicus. Chem. Pharm. Bull. 2005, 53, 382–386. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Duan, J.; Ishida, M.; Aida, K.; Tsuduki, T.; Zhang, J.; Manabe, Y.; Hirata, T.; Sugawara, T. Dietary cerebroside from sea cucumber (Stichopus japonicus): Absorption and effects on skin barrier and caecal short-chain fatty acids. J. Agric. Food Chem. 2016, 64, 7014–7021. [Google Scholar] [CrossRef]
- Yamada, K.; Wada, N.; Onaka, H.; Matsubara, R.; Isobe, R.; Inagaki, M.; Higuchi, R. Constituents of Holothuroidea. 15. Isolation of ante-iso type regioisomer on long chain base moiety of glucocerebroside from the sea cucumber Holothuria leucospilota. Chem. Pharm. Bull. 2005, 53, 788–791. [Google Scholar] [CrossRef]
- Sugawara, T.; Zaima, N.; Yamamoto, A.; Sakai, S.; Noguchi, R.; Hirata, T. Isolation of sphingoid bases of sea cucumber cerebrosides and their cytotoxicity against human colon cancer cells. Biosci. Biotechnol. Biochem. 2006, 70, 2906–2912. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, Y.; Inagaki, M.; Yamada, K.; Zhang, X.W.; Zhang, B.; Miyamoto, T.; Higuchi, R. Isolation and structure of a galactocerebroside from the sea cucumber Bohadschia argus. Chem. Pharm. Bull. 2009, 57, 315–317. [Google Scholar] [CrossRef]
- Xu, J.; Wang, Y.M.; Feng, T.Y.; Zhang, B.; Sugawara, T.; Xue, C.H. Isolation and anti-fatty liver activity of a novel cerebroside from the sea cucumber Acaudina molpadioides. Biosci. Biotechnol. Biochem. 2011, 75, 1466–1471. [Google Scholar] [CrossRef]
- Xu, J.; Duan, J.; Xue, C.; Feng, T.; Dong, P.; Sugawara, T.; Hirata, T. Analysis and comparison of glucocerebroside species from three edible sea cucumbers using liquid chromatography–ion trap–time-of-flight mass spectrometry. J. Agric. Food Chem. 2011, 59, 12246–12253. [Google Scholar] [CrossRef]
- Du, L.; Xu, J.; Xue, Y.; Takahashi, K.; Xue, C.H.; Wang, J.F.; Wang, Y.M. Cerebrosides from sea cucumber ameliorates cancer-associated cachexia in mice by attenuating adipose atrophy. J. Funct. Foods 2015, 17, 352–363. [Google Scholar] [CrossRef]
- Liu, X.; Xu, J.; Xue, Y.; Gao, Z.; Li, Z.; Leng, K.; Wang, J.; Xue, C.; Wang, Y. Sea cucumber cerebrosides and long-chain bases from Acaudina molpadioides protect against high fat diet-induced metabolic disorders in mice. Food Funct. 2015, 6, 3428–3536. [Google Scholar] [CrossRef]
- La, M.-P.; Shao, J.-J.; Jiao, J.; Yi, Y.-H. Three cerebrosides from the sea cucumber Cucumaria frondosa. Chin. J. Nat. Med. 2012, 10, 105–109. [Google Scholar] [CrossRef]
- Xu, J.; Guo, S.; Du, L.; Wang, Y.M.; Sugawara, T.; Hirata, T.; Xue, C.H. Isolation of cytotoxic glucocerebrosides and long-chain bases from sea cucumber Cucumaria frondosa using high speed counter-current chromatography. J. Oleo Sci. 2013, 62, 133–142. [Google Scholar] [CrossRef][Green Version]
- Xu, H.; Wang, F.; Wang, J.; Xu, J.; Wang, Y.; Xue, C. The WNT/β-catenin pathway is involved in the anti-adipogenic activity of cerebrosides from the sea cucumber Cucumaria frondosa. Food Funct. 2015, 6, 2396–2404. [Google Scholar] [CrossRef]
- Smirnova, G.P. Gangliosides from the starfish Evasterias echinosoma: Identification of a disialoganglioside containing 8-O-methyl-N-acetylneuraminic acid and N-formylgalactosamine. Russ. Chem. Bull. 2000, 49, 159–164. [Google Scholar] [CrossRef]
- Smirnova, G.P. Structure of gangliosides from gonads of the starfish Evasterias retifera. Russ. Chem. Bull. 2003, 52, 2270–2275. [Google Scholar] [CrossRef]
- Kawatake, S.; Inagaki, M.; Isobe, R.; Miyamoto, T.; Higuchi, R. Isolation and structure of monomethylated GM3-type ganglioside molecular species from the starfish Luidia maculata. Chem. Pharm. Bull. 2002, 50, 1386–1389. [Google Scholar] [CrossRef][Green Version]
- Kawatake, S.; Inagaki, M.; Isobe, R.; Miyamoto, T.; Higuchi, R. Isolation and structure of a GD3-type ganglioside molecular species possessing neuritogenic activity from the starfish Luidia maculata. Chem. Pharm. Bull. 2004, 52, 1002–1004. [Google Scholar] [CrossRef][Green Version]
- Miyamoto, T.; Yamamoto, A.; Wakabayashi, M.; Nagaregawa, Y.; Inagaki, M.; Higuchi, R.; Iha, M.; Teruya, K. Biologically active glycosides from Asteroidea. 40. Two new gangliosides, acanthagangliosides I and J from the starfish Acanthaster planci. Eur. J. Org. Chem. 2000, 2295–2301. [Google Scholar] [CrossRef]
- Kawano, Y.; Higuchi, R.; Komori, T. Biologically active glycosides from Asteroidea. XIX. Glycosphingolipids from the starfish Acanthaster planci. 4. Isolation and structure of five new gangliosides. Liebigs Ann. Chem. 1990, 43–50. [Google Scholar] [CrossRef]
- Miyamoto, T.; Inagaki, M.; Isobe, R.; Tanaka, Y.; Higuchi, R.; Iha, M.; Teruya, K. Biologically active glycosides from Asteroidea. 36. Re-examination of the structure of acanthaganglioside C, and the identification of three minor acanthagangliosides F, G and H. Liebigs Ann. Chem. 1997, 931–936. [Google Scholar] [CrossRef]
- Hanashima, S.; Sato, K.I.; Naito, Y.; Takematsu, H.; Kozutsumi, Y.; Ito, Y.; Yamaguchi, Y. Synthesis and binding analysis of unique AG2 pentasaccharide to human Siglec-2 using NMR techniques. Bioorg. Med. Chem. 2010, 18, 3720–3725. [Google Scholar] [CrossRef]
- Higuchi, R.; Inoue, S.; Inagaki, K.; Sakai, M.; Miyamoto, T.; Komori, T.; Inagaki, M.; Isobe, R. Biologically active glycosides from Asteroidea. 42. Isolation and structure of a new biologically active ganglioside molecular species from the starfish Asterina pectinifera. Chem. Pharm. Bull. 2006, 54, 287–291. [Google Scholar] [CrossRef]
- Smirnova, G.P. Hematoside with 8-O-methyi-N-glycolylneuraminic acid from the starfish Linckia laevigata. Russ. Chem. Bull. 2000, 49, 165–168. [Google Scholar] [CrossRef]
- Inagaki, M.; Miyamoto, T.; Isobe, R.; Higuchi, R. Biologically active glycosides from Asteroidea. 43. Isolation and structure of a new neuritogenic-active ganglioside molecular species from the starfish Linckia laevigata. Chem. Pharm. Bull. 2005, 53, 1551–1554. [Google Scholar] [CrossRef] [PubMed]
- Inagaki, M.; Saito, T.; Miyamoto, T.; Higuchi, R. Isolation and structure of hematoside-type ganglioside from the starfish Linckia laevigata. Chem. Pharm. Bull. 2009, 57, 204–206. [Google Scholar] [CrossRef]
- Pan, K.; Tanaka, C.; Inagaki, M.; Higuchi, R.; Miyamoto, T. Isolation and structure elucidation of GM4-type gangliosides from the Okinawan starfish Protoreaster nodosus. Mar. Drugs 2012, 10, 2467–2480. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Harada, Y.; Miyamoto, T.; Isobe, R.; Higuchi, R. Constituents of Holothuroidea. 9. Isolation and structure of a new ganglioside molecular species from the sea cucumber Holothuria pervicax. Chem. Pharm. Bull. 2000, 48, 157–159. [Google Scholar] [CrossRef]
- Yamada, K.; Harada, Y.; Nagaregawa, Y.; Miyamoto, T.; Isobe, R.; Higuchi, R. Constituents of Holothuroidea. 7. Isolation and structure of biologically active gangliosides from the sea cucumber Holothuria pervicax. Eur. J. Org. Chem. 1998, 2519–2525. [Google Scholar] [CrossRef]
- Yamada, K.; Matsubara, R.; Kaneko, M.; Miyamoto, T.; Higuchi, R. Constituents of Holothuroidea. 10. Isolation and structure of a biologically active ganglioside molecular species from the sea cucumber Holothuria leucospilota. Chem. Pharm. Bull. 2001, 49, 447–452. [Google Scholar] [CrossRef]
- Kaneko, M.; Kisa, F.; Yamada, K.; Miyamoto, T.; Higuchi, R. Structure of a new neuritogenic-active ganglioside from the sea cucumber Stichopus japonicus. Eur. J. Org. Chem. 2003, 1004–1008. [Google Scholar] [CrossRef]
- Kaneko, M.; Kisa, F.; Yamada, K.; Miyamoto, T.; Higuchi, R. Constituents of Holothuroidea. 8-Structure of neuritogenic active ganglioside from the sea cucumber Stichopus japonicus. Eur. J. Org. Chem. 1999, 3171–3174. [Google Scholar] [CrossRef]
- Yamada, K.; Hamada, A.; Kisa, F.; Miyamoto, T.; Higuchi, R. Constituents of Holothuroidea. 13. Structure of neuritogenic active ganglioside molecular species from the sea cucumber Stichopus chloronotus. Chem. Pharm. Bull. 2003, 51, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Kisa, F.; Yamada, K.; Miyamoto, T.; Inagaki, M.; Higuchi, R. Constituents of Holothuroidea. 17. Isolation and structure of biologically active monosialo-gangliosides from the sea cucumber Cucumaria echinata. Chem. Pharm. Bull. 2006, 54, 982–987. [Google Scholar] [CrossRef]
- Kisa, F.; Yamada, K.; Miyamoto, T.; Inagaki, M.; Higuchi, R. Constituents of Holothuroidea. 18. Isolation and structure of biologically active disialo- and trisialo-gangliosides from the sea cucumber Cucumaria echinata. Chem. Pharm. Bull. 2006, 54, 1293–1298. [Google Scholar] [CrossRef] [PubMed]
- Traving, C.; Schauer, R. Structure, function and metabolism of sialic acids. Cell. Mol. Life Sci. 1998, 54, 1330–1349. [Google Scholar] [CrossRef]
- Kolter, T. Ganglioside biochemistry. ISRN Biochem. 2012, 2012, 506160. [Google Scholar] [CrossRef]
- Sipione, S.; Monyror, J.; Galleguillos, D.; Steinberg, N.; Kadam, V. Gangliosides in the brain: Physiology, pathophysiology and therapeutic applications. Front. Neurosci. 2020, 14, 572965. [Google Scholar] [CrossRef]
- Schengrund, C.-L. Gangliosides and neuroblastomas. Int. J. Mol. Sci. 2020, 21, 5313. [Google Scholar] [CrossRef] [PubMed]
- Schömel, N.; Geisslingera, G.; Wegnera, M.-S. Influence of glycosphingolipids on cancer cell energy metabolism. Prog. Lipid Res. 2020, 79, 101050. [Google Scholar] [CrossRef]
- Yamada, K.; Hara, E.; Miyamoto, T.; Higuchi, R.; Isobe, R.; Honda, S. Constituents of Holothuroidea, 6-Isolation and structure of biologically active glycosphingolipids from the sea cucumber Cucumaria echinata. Eur. J. Org. Chem. 1998, 371–378. [Google Scholar] [CrossRef]
- Kisa, F.; Yamada, K.; Miyamoto, T.; Inagaki, M.; Higuchi, R. Determination of the absolute configuration of sialic acids in gangliosides from the sea cucumber Cucumaria echinata. Chem. Pharm. Bull. 2007, 55, 1051–1052. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kicha, A.A.; Kalinovsky, A.I.; Malyarenko, T.V.; Ivanchina, N.V.; Dmitrenok, P.S.; Menchinskaya, E.S.; Yurchenko, E.A.; Pislyagin, E.A.; Aminin, D.L.; Huong, T.T.; et al. Cyclic steroid glycosides from the starfish Echinaster luzonicus: Structures and immunomodulatory activities. J. Nat. Prod. 2015, 78, 1397–1405. [Google Scholar] [CrossRef]
- Malyarenko, T.V.; Kharchenko, S.D.; Kicha, A.A.; Ivanchina, N.V.; Dmitrenok, P.S.; Chingizova, E.A.; Pislyagin, E.A.; Evtushenko, E.V.; Antokhina, T.I.; Minh, C.V.; et al. Anthenosides L‒U, steroidal glycosides with unusual structural features from the starfish Anthenea aspera. J. Nat. Prod. 2016, 79, 3047–3056. [Google Scholar] [CrossRef]
- Mondol, M.A.M.; Shin, H.J.; Rahman, M.A.; Islam, M.T. Sea cucumber glycosides: Chemical structures, producing species and important biological properties. Mar. Drugs 2017, 15, 317. [Google Scholar] [CrossRef]





















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Order | Family | Scientific Name | Compounds | Type of Biological Activity | Ref. | ||
|---|---|---|---|---|---|---|---|
| Ceramides | Cerebrosides | Gangliosides | |||||
| Class Asteroidea | |||||||
| Forcipulatida | Asteriidae | Allostichaster inaequalis | 18, 19 | [39] | |||
| Anasterias minuta | 26, 27 | [43] | |||||
| Asterias amurensis | 2 | 30, 59–64 | Stimulating root growth Brassica campestris (2, 30); | [23,50] | |||
| Distolasterias nipon | 1–11 | 65–68 | [33,34] | ||||
| Evasterias echinosoma | 117, 118 | [66] | |||||
| Evasterias retifera | 125, 126 | [67] | |||||
| Cosmasterias lurida | 27 | [40] | |||||
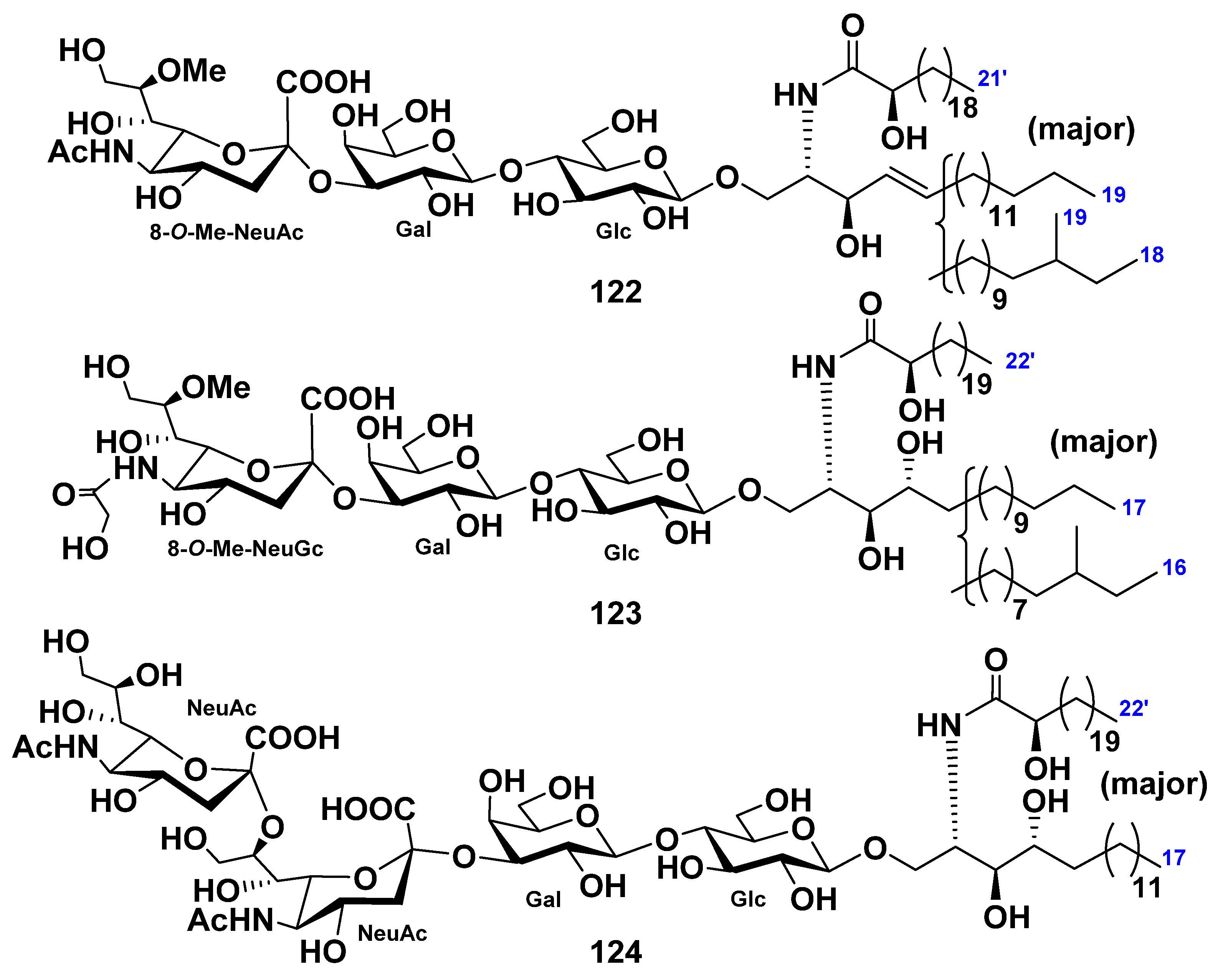
| Paxillosida | Luidiidae | Luidia maculata | 12–17 | 20–25 | 122–124 | Neuritogenic activity toward the rat pheochromocytoma PC12 cells in the presence of NGF (124) | [35,41,42,68,69] |
| Valvatida | Acanthaster-idae | Acanthaster planci | 115, 116 | Binding epitope of AG2 pentasaccharide to human Siglec-2 | [70,71,72,73] | ||
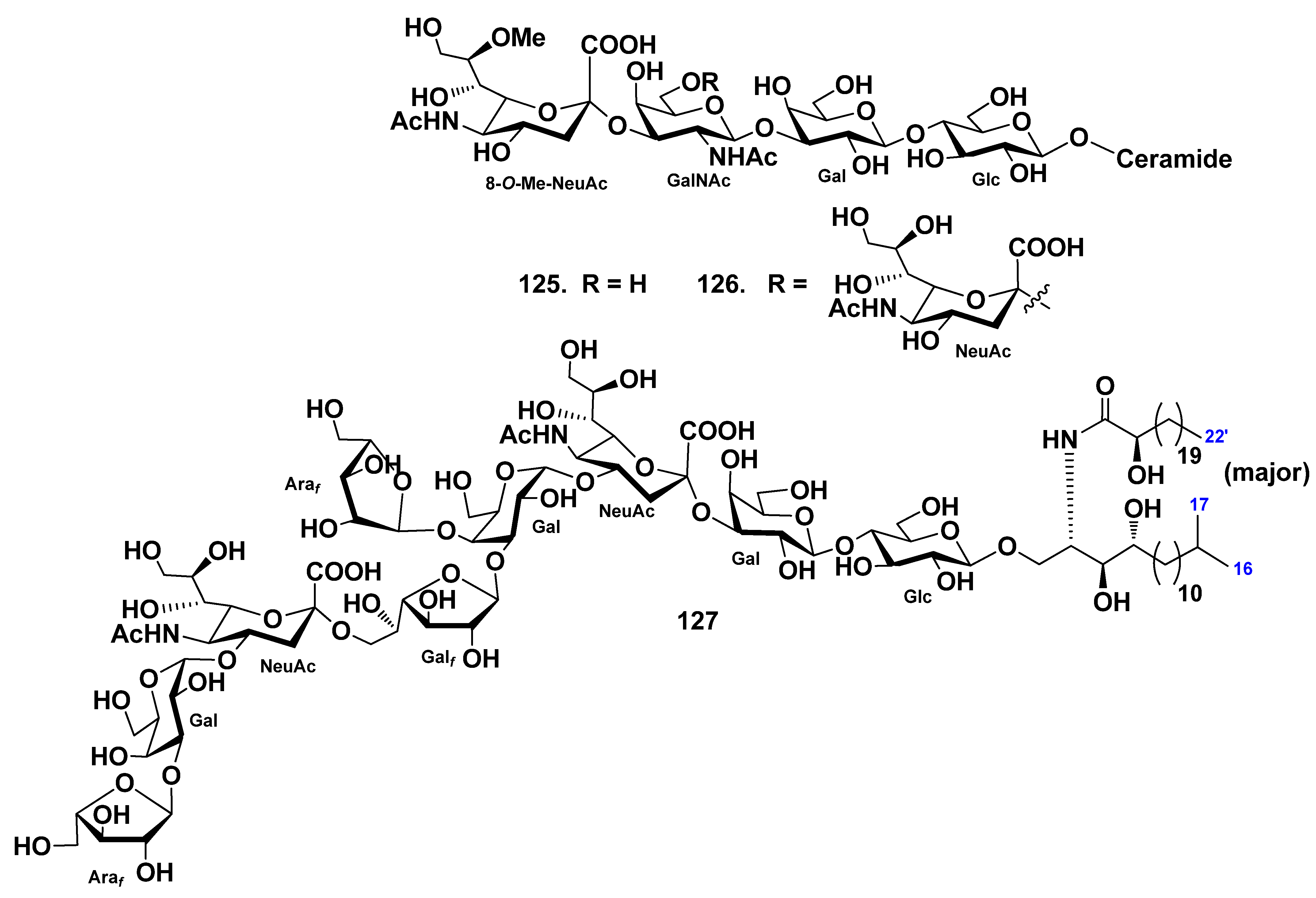
| Asterinidae | Patiria (=Asterina) pectinifera | 127 | Neuritogenic activity toward the rat pheochromocytoma PC12 cells in the presence of NGF | [74] | |||
| Ophidiasteridae | Linckia laevigata | 28 | 119–121 | Neuritogenic activity toward the rat pheochromocytoma PC12 cells in the presence of NGF (120) | [44,75,76,77] | ||
| Oreasteridae | Culcita novaeguineae | 29 | [45] | ||||
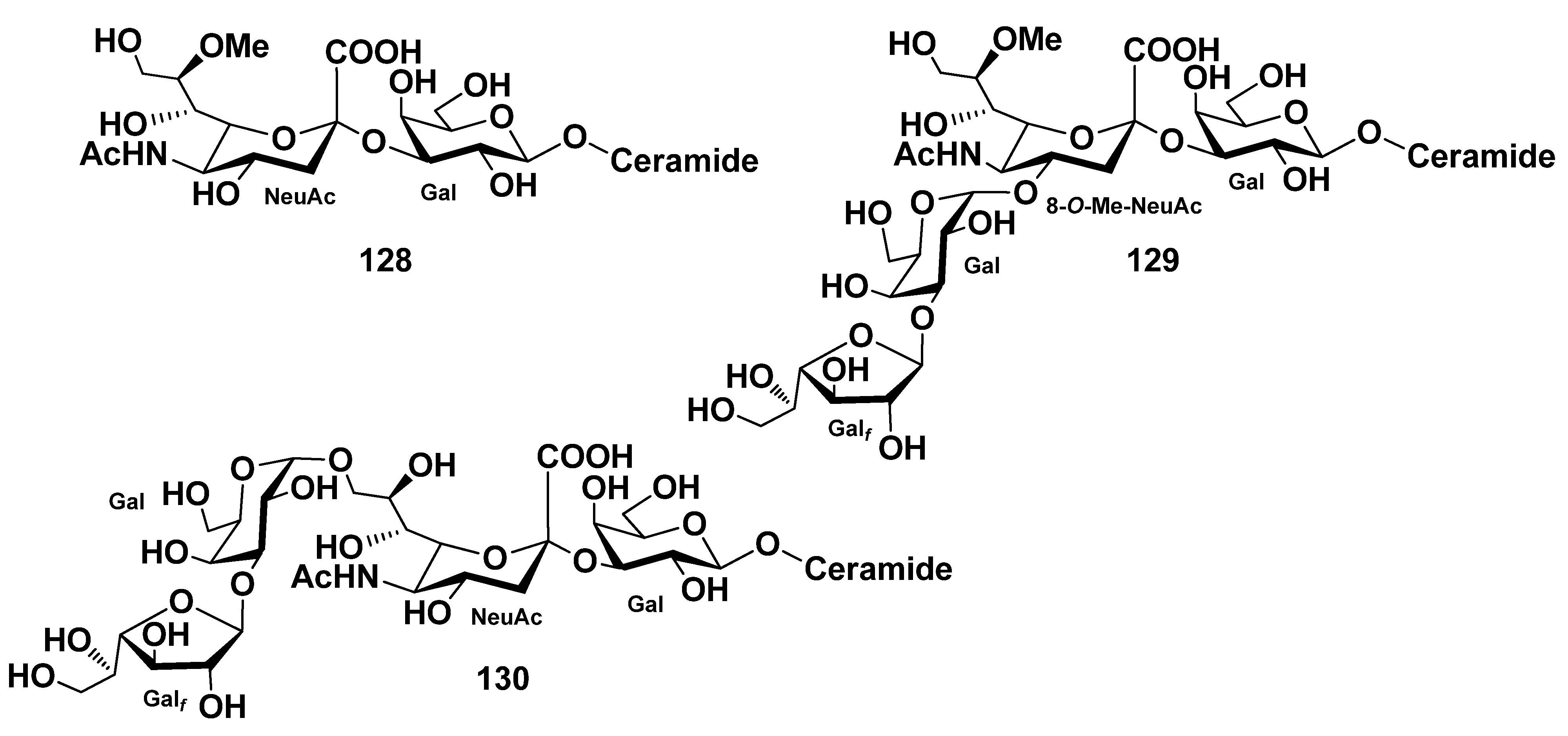
| Protoreaster nodosus | 43–58 | 128–130 | [49,78] | ||||
| Oreaster reticulatus | 31–39 | (1) Mildly cytotoxic activity on the rat glioma C6 cells (31–39); (2) exertion of proangiogenic activity and increase of VEGF-induced human endothelial cell proliferation (39). | [46] | ||||
| Narcissia canariensis | 40–42 | Cytotoxic activity against KB cells (40) | [47] | ||||
| Class Holothuroidea | |||||||
| Holothuriida | Holothuri-idae | Holothuria coronopertusa | 69–86 | [52] | |||
| Holothuria pervicax | 87–96 | 131 | Neuritogenic activity toward the rat pheochromocytoma PC12 cell line (131) | [53,79,80] | |||
| Holothuria leucospilota | 102–105 | 132–134 | Neuritogenic activity toward the rat pheochromocytoma PC12 cell line (132–134) | [56,81] | |||
| Bohadschia argus | 106, 107 | [58] | |||||
| Synallactida | Stichopodi-dae | Stichopus japonicus | 97–101 | 135 | (1) Absorption of cerebrosides in vivo and improving skin barrier functions (97–101); (2) neuritogenic activity toward the rat pheochromocytoma PC12 cells in the presence of NGF (135). | [54,55,82,83] | |
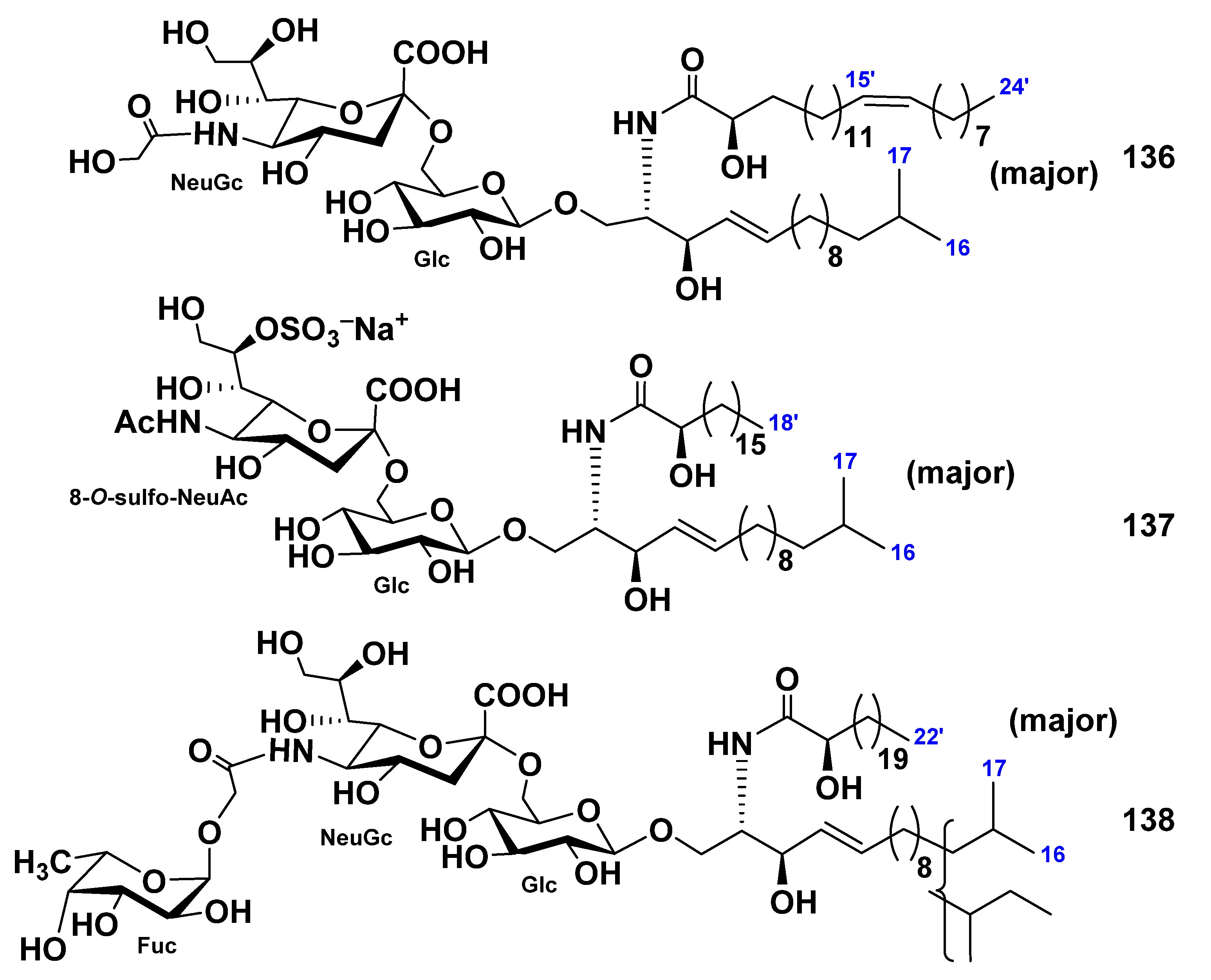
| Stichopus chloronotus | 136–138 | Neuritogenic activity toward the rat pheochromocytoma PC12 cells in the presence of NGF | [84] | ||||
| Molpadida | Caudinidae | Acaudina molpadioides | 108 | (1) Anti-fatty liver activity of 108 in the rats with fatty liver induced by orotic acid; (2) alleviating adipose atrophy in the cancer-associated cachexia mice; (3) effects of cerebrosides on the obesity-related metabolic disorders in mice. | [59,61,62] | ||
| Dendrochi-rotida | Cucu-mariidae | Cucumaria frondosa | 109–114 | (1) In vitro cytotoxic activity against Caco-2 colon cancer cells (112–114); (2) in vitro antiadipogenic activity of cerebrosides. | [63,64,65] | ||
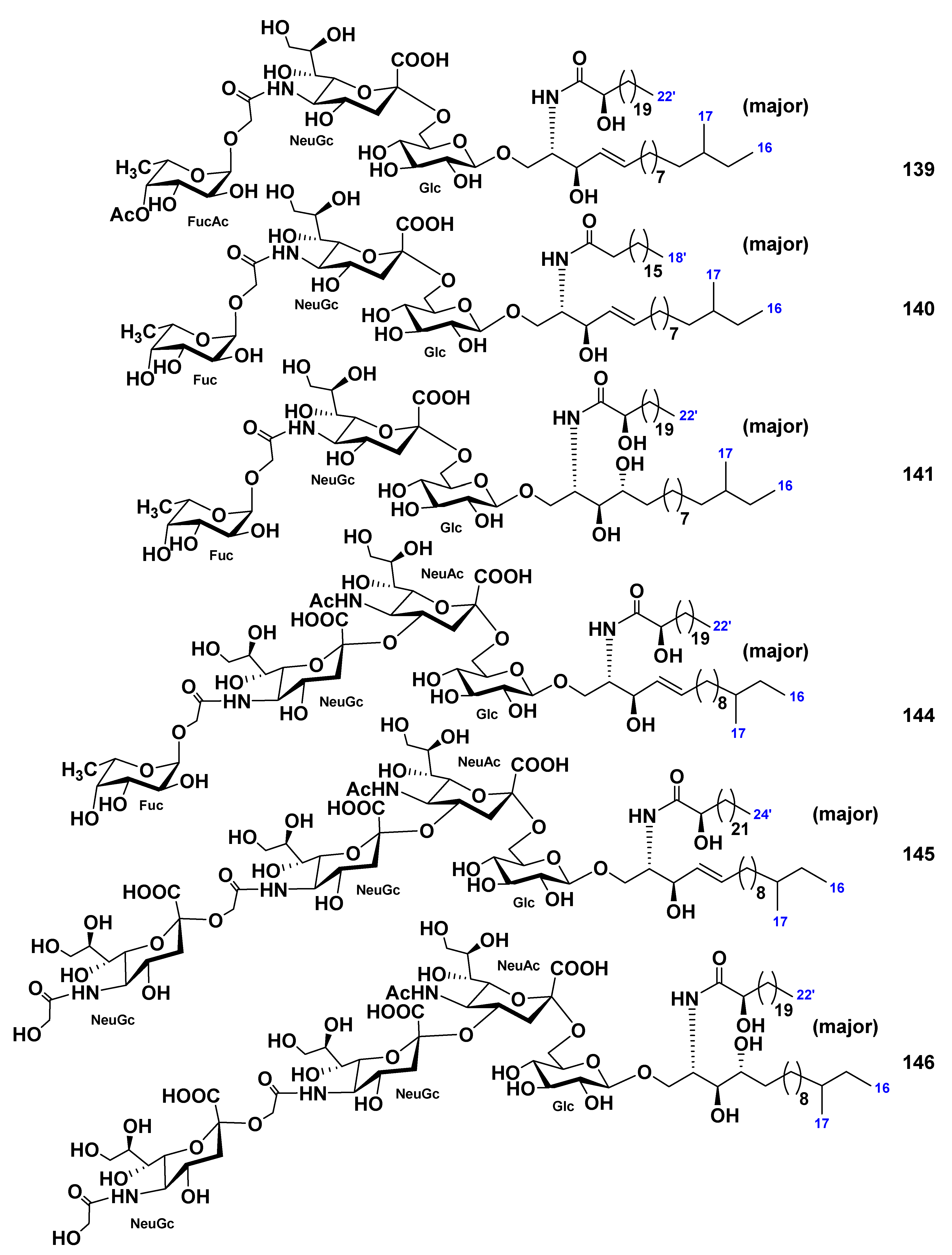
| Cucumaria echinata | 139–141, 144–146 142, 143 | Neuritogenic activity toward the rat pheochromocytoma PC12 cells in the presence of NGF | [85,86] | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malyarenko, T.V.; Kicha, A.A.; Stonik, V.A.; Ivanchina, N.V. Sphingolipids of Asteroidea and Holothuroidea: Structures and Biological Activities. Mar. Drugs 2021, 19, 330. https://doi.org/10.3390/md19060330
Malyarenko TV, Kicha AA, Stonik VA, Ivanchina NV. Sphingolipids of Asteroidea and Holothuroidea: Structures and Biological Activities. Marine Drugs. 2021; 19(6):330. https://doi.org/10.3390/md19060330
Chicago/Turabian StyleMalyarenko, Timofey V., Alla A. Kicha, Valentin A. Stonik, and Natalia V. Ivanchina. 2021. "Sphingolipids of Asteroidea and Holothuroidea: Structures and Biological Activities" Marine Drugs 19, no. 6: 330. https://doi.org/10.3390/md19060330
APA StyleMalyarenko, T. V., Kicha, A. A., Stonik, V. A., & Ivanchina, N. V. (2021). Sphingolipids of Asteroidea and Holothuroidea: Structures and Biological Activities. Marine Drugs, 19(6), 330. https://doi.org/10.3390/md19060330