
Bioprospecting of Less-Polar Constituents from Endemic Brown Macroalga Fucus virsoides J. Agardh from the Adriatic Sea and Targeted Antioxidant Effects In Vitro and In Vivo (Zebrafish Model)

 ,
,  ,
,  , , ,
, , , 



Abstract
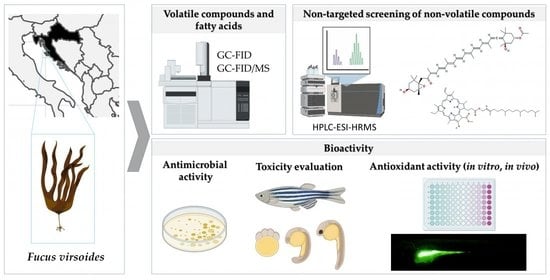
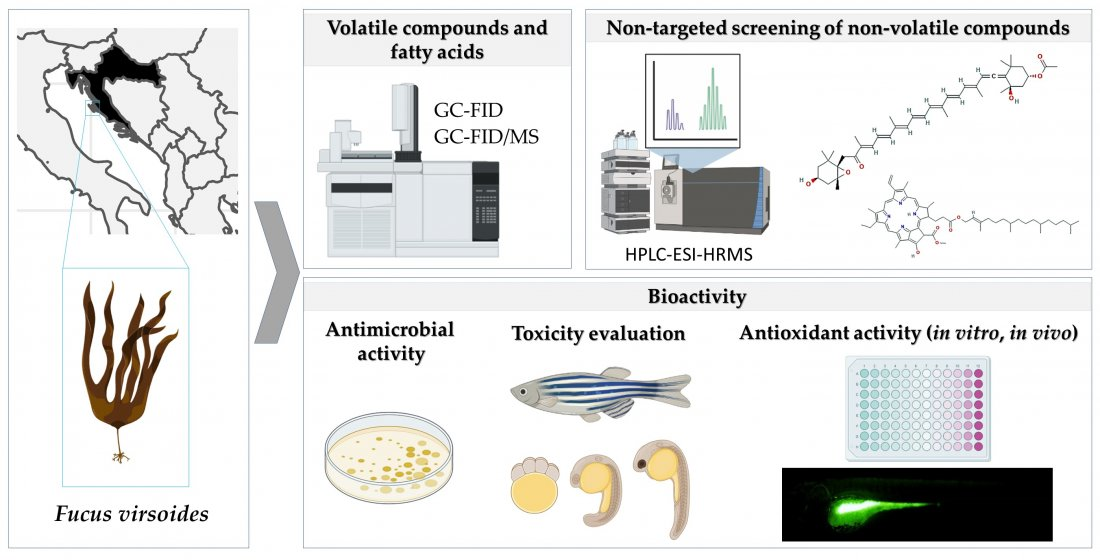
1. Introduction
2. Results and Discussion
2.1. Volatile Organic Compounds (VOCs) Analysis
2.1.1. The Headspace Composition
2.1.2. The Volatile Oil Composition
2.2. Analysis of Fatty Acids
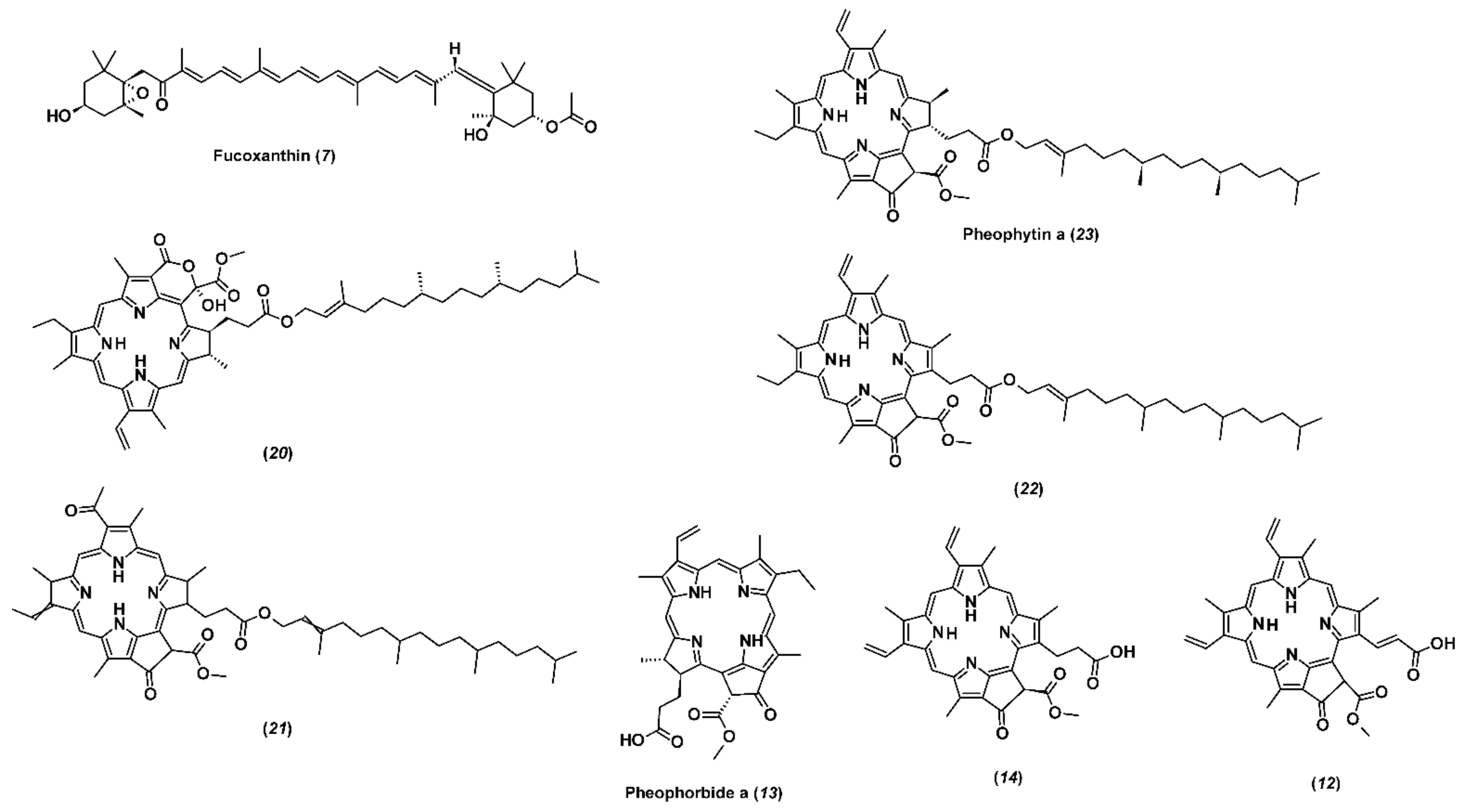
2.3. Non-Targeted Screening of Less Polar Non-Volatile Compounds
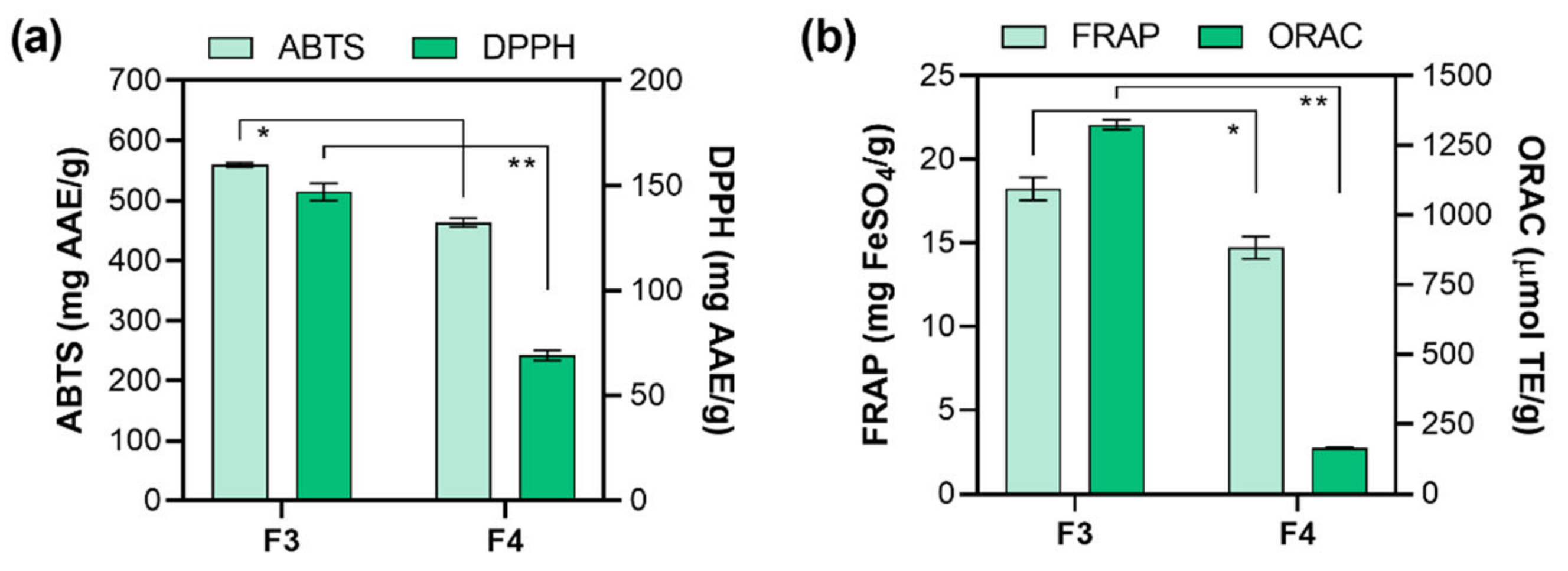
2.4. In Vitro Antioxidant Activity Determination
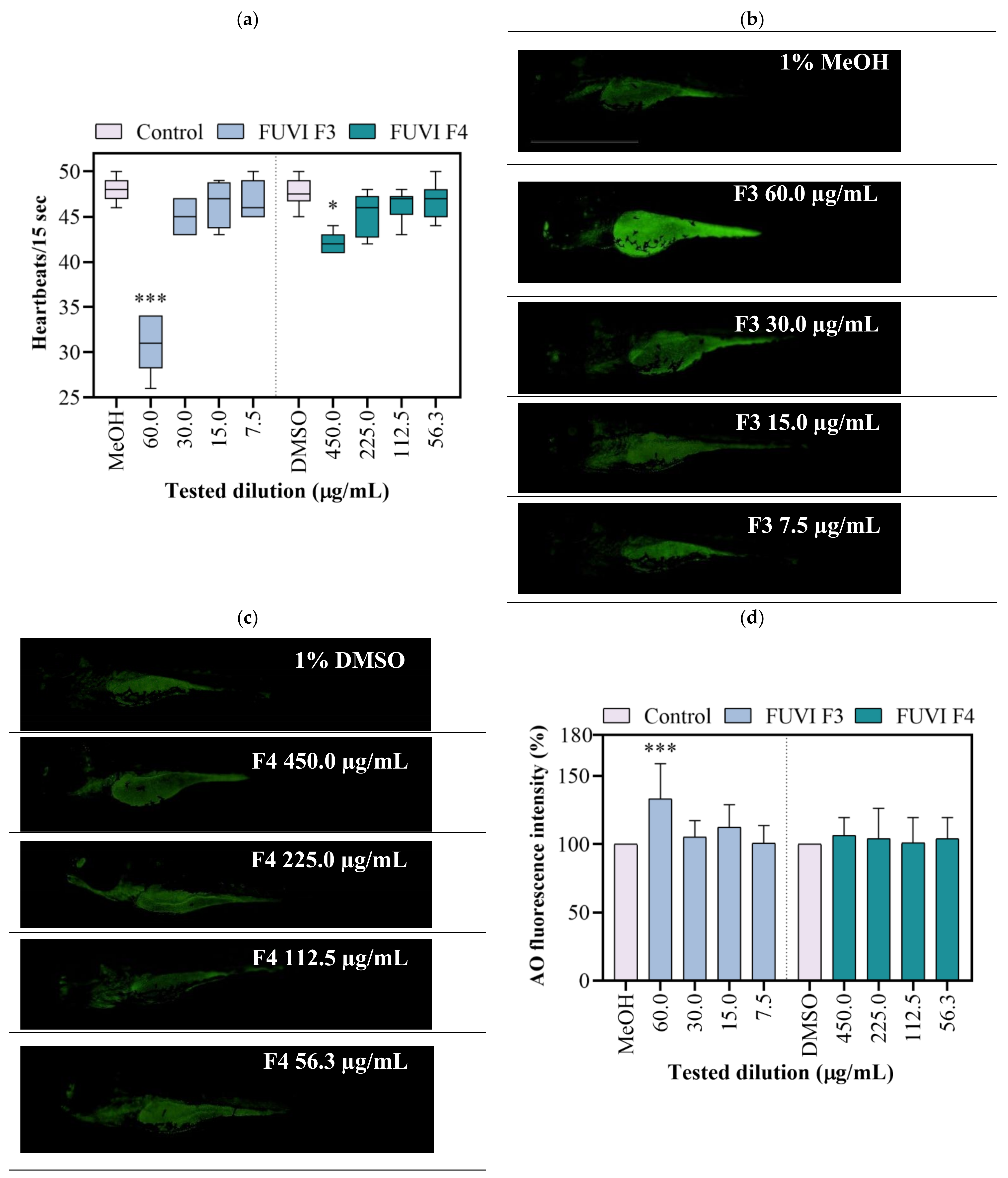
2.5. Zebrafish Embryotoxicity
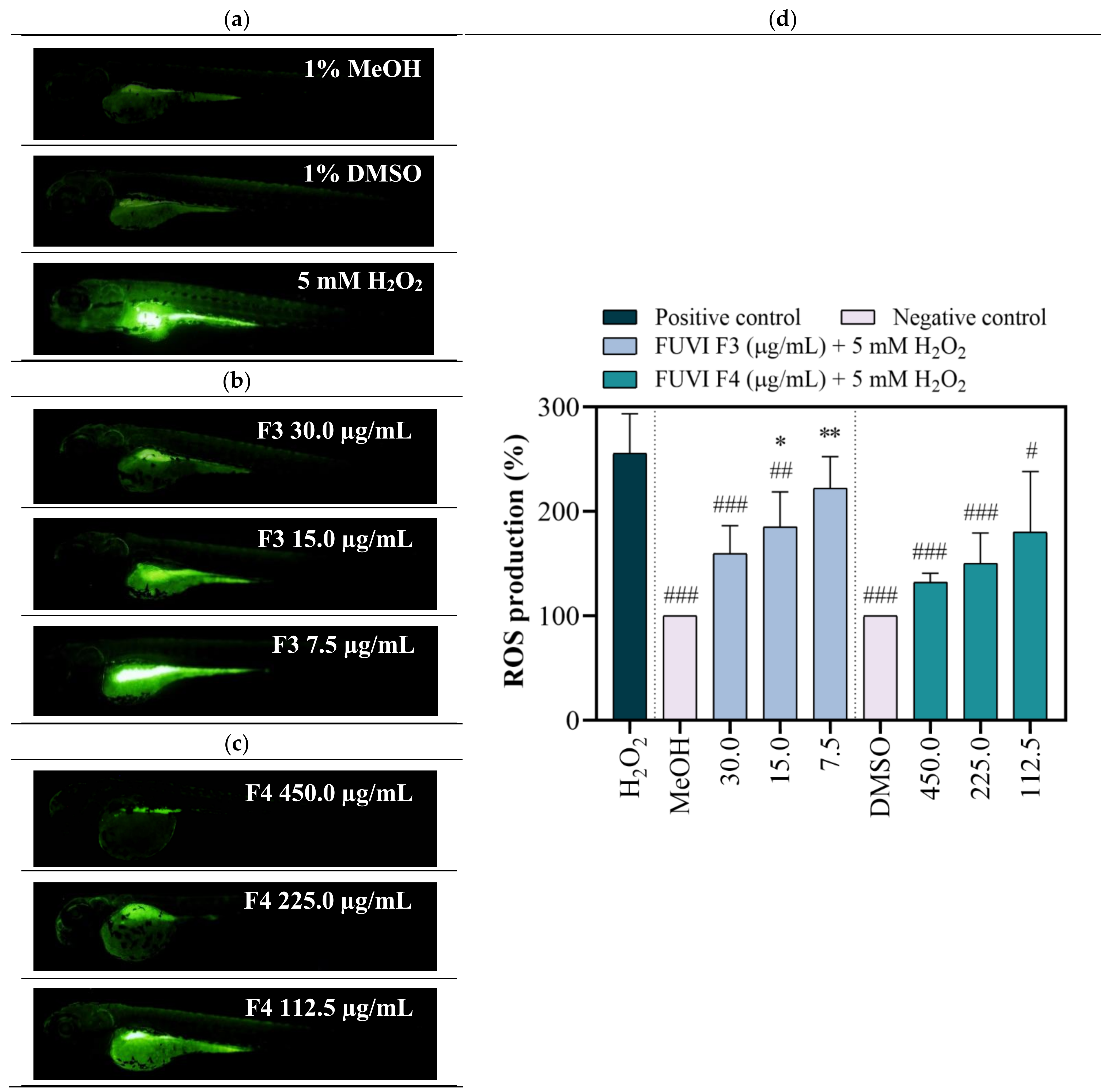
2.6. In Vivo Antioxidant Activity (Zebrafish Model)
2.7. Antimicrobial Activity
3. Materials and Methods
3.1. Chemicals
3.2. The Sample Collection and Preparation Procedure
3.3. Headspace Solid-Phase Microextraction (HS-SPME)
3.4. Hydrodistillation (HD)
3.5. Gas Chromatography and Mass Spectrometry (GC-FID/MS) Analysis
3.6. The Analysis of Fatty Acids by GC-FID
3.7. Fractionation Using Solid-Phase Extraction (SPE)
3.8. Non-Targeted Screening by High Performance Liquid Chromatography-High-Resolution Mass Spectrometry (HPLC-ESI-HRMS)
3.9. In Vitro Antioxidant Activity Determination
3.10. Zebrafish Embryotoxicity Test
3.11. In Vivo Antioxidant Activity Determination by Zebrafish Model
3.12. Testing of Antimicrobial Activity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Andrade, P.B.; Barbosa, M.; Matos, R.P.; Lopes, G.; Vinholes, J.; Mouga, T.; Valentão, P. Valuable compounds in macroalgae extracts. Food Chem. 2013, 138, 1819–1828. [Google Scholar] [CrossRef] [PubMed]
- Kremer, B.P.; Munda, I.M. Ecophysiological Studies of the Adriatic Seaweed, Fucus virsoides. Mar. Ecol. 1982, 3, 75–93. [Google Scholar] [CrossRef]
- Zavodnik, N. Seasonal variations in rate of photosynthetic activity and chemical composition of the littoral seaweeds common to north Adriatic. Part I. Fucus virsoides (DON) J. AG. Bot. Mar. 1973, 16, 155–165. [Google Scholar] [CrossRef]
- Falsone, G.; Cateni, F.; Vrech, E. Triacylglycerols, fucosterol, diacylglyceroglycolipids and fucoxanthin from Fucus virsoides. J. Ag. Z. Nat. 1994, 49, 1297–1304. [Google Scholar] [CrossRef]
- Pelivan, A.; Lutkić, A. Sulphated polysaccharides of brown seaweeds Cystoseira compressa, Fucus virsoides, and Dyctiopteris membranacea. Croat. Chem. Acta 1994, 67, 407–413. [Google Scholar]
- Kapetanovic, R.; Sladic, D.; Popov, S.; Zlatovic, M.; Kljajic, Z.; Gasic, M. Sterol composition of the Adriatic sea algae Ulva lactuca, Codium dichotomum, Cystoseira adriatica and Fucus virsoides. J. Serb. Chem. Soc. 2005, 70, 1395–1400. [Google Scholar] [CrossRef]
- Najdek, M.; Iveša, Lj.; Paliaga, P.; Blažina, M.; Čelig, A. Changes in the fatty acid composition of Fucus virsoides J. Agardh in response to the type of substratum. Acta Adriat. 2014, 55, 19–30. [Google Scholar]
- Grozdanić, N.; Zdunić, G.; Šavikin, K.; Đuričić, I.; Kosanić, M.; Mačić, V.; Matić, I.; Stanojković, T. Seasonal variation in biopharmaceutical activity and fatty acid content of endemic Fucus virsoides algae from Adriatic Sea. Acta Pol. Pharm. Drug Res. 2019, 76, 833–844. [Google Scholar] [CrossRef]
- Youngblood, W.W.; Blumer, M.; Guillard, R.L.; Fiore, F. Saturated and unsaturated hydrocarbons in marine benthic algae. Mar. Biol. 1971, 8, 190–201. [Google Scholar] [CrossRef]
- Youngblood, W.W.; Blumer, M. Alkanes and alkenes in marine benthic algae. Mar. Biol. 1973, 21, 163–172. [Google Scholar] [CrossRef]
- Clark, R.C., Jr.; Blumer, M. Distribution of n-paraffins in marine organisms and sediment. Limnol. Oceanogr. 1967, 12, 79–87. [Google Scholar] [CrossRef]
- Miyashita, K.; Mikami, N.; Hosokawa, M. Chemical and nutritional characteristics of brown seaweed lipids: A review. J. Funct. Foods 2013, 5, 1507–1517. [Google Scholar] [CrossRef]
- Maeda, H.; Hosokawa, M.; Sashima, T.; Miyashita, K. Dietary Combination of fucoxanthin and fish oil attenuates the weight gain of white adipose tissue and decreases blood glucose in obese/diabetic KK-AyMice. J. Agric. Food Chem. 2007, 55, 7701–7706. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Fang, S.; Xiong, Z. Protective effect of polysaccharide from Ligusticum chuanxiong hort against H2O2-induced toxicity in zebrafish embryo. Carbohydr. Polym. 2019, 221, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Jerković, I.; Kranjac, M.; Marijanović, Z.; Roje, M.; Jokić, S. Chemical diversity of headspace and volatile oil composition of two brown algae (Taonia atomaria and Padina pavonica) from the Adriatic Sea. Molecules 2019, 24, 495. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, F.J.; Zamora, R. Formation of phenylacetic acid and benzaldehyde by degradation of phenylalanine in the presence of lipid hydroperoxides: New routes in the amino acid degradation pathways initiated by lipid oxidation products. Food Chem. X 2019, 2, 100037. [Google Scholar] [CrossRef] [PubMed]
- McInnes, A.G.; Walter, J.A.; Wright, J.L.C. Biosynthesis of hydrocarbons by algae: Decarboxylation of stearic acid to n-heptadecane in Anacystis nidulans determined by 13C- and 2H-labeling and 13C nuclear magnetic resonance. Lipids 1980, 15, 609–615. [Google Scholar] [CrossRef]
- Rontani, J.-F.; Volkman, J.K. Phytol degradation products as biogeochemical tracers in aquatic environments. Org. Geochem. 2003, 34, 1–35. [Google Scholar] [CrossRef]
- Rontani, J.-F.; Giral, P.-P.; Baillet, G.; Raphel, D. “Bound” 6,10,14-trimethylpentadecan-2-one: A useful marker for photodegradation of chlorophylls with a phytol ester group in seawater. Org. Geochem. 1992, 18, 139–142. [Google Scholar] [CrossRef]
- Grozdanic, N.; Djuricic, I.; Kosanic, M.; Zdunic, G.; Savikin, K.; Etahiri, S.; Assobhei, O.; Benba, J.; Petovic, S.; Matic, I.Z.; et al. Fucus spiralis extract and fractions: Anticancer and pharmacological potentials. J. B.U. Off. J. Balk. Union Oncol. 2020, 25, 1219–1229. [Google Scholar]
- Van Ginneken, V.J.; Helsper, J.P.; De Visser, W.; Van Keulen, H.; Brandenburg, W.A. Polyunsaturated fatty acids in various macroalgal species from north Atlantic and tropical seas. Lipids Health Dis. 2011, 10, 104–108. [Google Scholar] [CrossRef]
- Francisco, J.; Horta, A.; Pedrosa, R.; Afonso, C.; Cardoso, C.; Bandarra, N.M.; Gil, M.M. Bioaccessibility of antioxidants and fatty acids from Fucus spiralis. Foods 2020, 9, 440. [Google Scholar] [CrossRef]
- Lorenzo, J.M.; Agregán, R.; Munekata, P.E.S.; Franco, D.; Carballo, J.; Şahin, S.; Lacomba, R.; Barba, F.J. Proximate Composition and nutritional value of three macroalgae: Ascophyllum nodosum, Fucus vesiculosus and Bifurcaria bifurcata. Mar. Drugs 2017, 15, 360. [Google Scholar] [CrossRef]
- Kim, M.-K.; Dubacq, J.-P.; Thomas, J.-C.; Giraud, G. Seasonal variations of triacylglycerols and fatty acids in Fucus serratus. Phytochemistry 1996, 43, 49–55. [Google Scholar] [CrossRef]
- Paiva, L.; Lima, E.; Patarra, R.F.; Neto, A.I.; Baptista, J. Edible Azorean macroalgae as source of rich nutrients with impact on human health. Food Chem. 2014, 164, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Khotimchenko, S.V.; Vaskovsky, V.E.; Titlyanova, T.V. Fatty acids of marine algae from the Pacific coast of North California. Bot. Mar. 2002, 45, 17–22. [Google Scholar] [CrossRef]
- Herbreteau, F.; Coiffard, L.J.M.; Derrien, A.; De Roeck-Holtzhauer, Y. The fatty acid composition of five species of macroalgae. Bot. Mar. 1997, 40, 25–28. [Google Scholar] [CrossRef]
- Silva, G.; Pereira, R.B.; Valentão, P.; Andrade, P.B.; Sousa, C. Distinct fatty acid profile of ten brown macroalgae. Rev. Bras. De Farm. 2013, 23, 608–613. [Google Scholar] [CrossRef]
- Ochi, M.; Watanabe, M.; Miura, I.; Taniguchi, M.; Tokoroyama, T. Amijiol, isoamijiol, and 14-deoxyamijiol, three new diterpenoids from the brown seaweed Dictyota linearis. Chem. Lett. 1980, 9, 1229–1232. [Google Scholar] [CrossRef]
- Matsuno, T. Aquatic animal carotenoids. Fish. Sci. 2001, 67, 771–783. [Google Scholar] [CrossRef]
- Dasgupta, C.N. Algae as a source of phycocyanin and other industrially important pigments. In Algae Biorefinery: An Integrated Approach, 1st ed.; Das, D., Ed.; Springer International Publishing: Cham, Switzerland, 2015; pp. 253–276. [Google Scholar]
- Maeda, H.; Fukuda, S.; Izumi, H.; Saga, N. Anti-oxidant and fucoxanthin contents of brown alga ishimozuku (Sphaerotrichia divaricata) from the west coast of Aomori, Japan. Mar. Drugs 2018, 16, 255. [Google Scholar] [CrossRef]
- Méresse, S.; Fodil, M.; Fleury, F.; Chénais, B. Fucoxanthin, a marine-derived carotenoid from brown seaweeds and microalgae: A promising bioactive compound for cancer therapy. Int. J. Mol. Sci. 2020, 21, 9273. [Google Scholar] [CrossRef]
- Osório, C.; Machado, S.; Peixoto, J.; Bessada, S.; Pimentel, F.B.; Alves, R.C.; Oliveira, M.B.P.P. Pigments content (chlorophylls, fucoxanthin and phycobiliproteins) of different commercial dried algae. Separations 2020, 7, 33. [Google Scholar] [CrossRef]
- Haugan, J.A.; Liaaen-Jensen, S. Algal carotenoids 54. Carotenoids of brown algae (Phaeophyceae). Biochem. Syst. Ecol. 1994, 22, 31–41. [Google Scholar] [CrossRef]
- Haugan, J.A.; Liaaen-Jensen, S. Improved isolation procedure for fucoxanthin. Phytochemistry 1989, 28, 2797–2798. [Google Scholar] [CrossRef]
- Zaragozá, M.C.; López, D.; Sáiz, M.P.; Poquet, M.; Pérez, J.; Puig-Parellada, P.; Màrmol, F.; Simonetti, P.; Gardana, C.; Lerat, Y.; et al. Toxicity and antioxidant activity in vitro and in vivo of two Fucus vesiculosus extracts. J. Agric. Food Chem. 2008, 56, 7773–7780. [Google Scholar] [CrossRef] [PubMed]
- Imbs, T.I.; Ermakova, S.P.; Fedoreyev, S.A.; Anastyuk, S.D.; Zvyagintseva, T.N. Isolation of fucoxanthin and highly unsaturated monogalactosyldiacylglycerol from brown alga Fucus evanescens C Agardh and in vitro investigation of their antitumor activity. Mar. Biotechnol. 2013, 15, 606–612. [Google Scholar] [CrossRef] [PubMed]
- Ramus, J.; Lemons, F.; Zimmerman, C. Adaptation of light-harvesting pigments to downwelling light and the consequent photosynthetic performance of the eulittoral rockweeds Ascophyllum nodosum and Fucus vesiculosus. Mar. Biol. 1977, 42, 293–303. [Google Scholar] [CrossRef]
- Terasaki, M.; Hirose, A.; Narayan, B.; Baba, Y.; Kawagoe, C.; Yasui, H.; Saga, N.; Hosokawa, M.; Miyashita, K. Evaluation of recoverable functional lipid components of several brown seaweeds (phaeophyta) from japan with special reference to fucoxanthin and fucosterol contents. J. Phycol. 2009, 45, 974–980. [Google Scholar] [CrossRef]
- Lalegerie, F.; Gager, L.; Stiger-Pouvreau, V.; Connan, S. The stressful life of red and brown seaweeds on the temperate intertidal zone: Effect of abiotic and biotic parameters on the physiology of macroalgae and content variability of particular metabolites. In Advances in Botanical Research; Elsevier BV: Amsterdam, The Netherlands, 2020; pp. 247–287. [Google Scholar]
- Matishov, G.G.; Makarov, M.V. Changes in the pigment composition of Fucus vesiculosus L. and Fucus serratus L. from the Barents Sea during prolonged exposure to the dark. Dokl. Biol. Sci. 2004, 397, 338–339. [Google Scholar] [CrossRef]
- Connan, S. Spectrophotometric assays of major compounds extracted from algae. Adv. Struct. Saf. Stud. 2015, 1308, 75–101. [Google Scholar] [CrossRef]
- Kuczynska, P.; Jemiola-Rzeminska, M.; Strzalka, K. Photosynthetic pigments in diatoms. Mar. Drugs 2015, 13, 5847–5881. [Google Scholar] [CrossRef] [PubMed]
- Indrawati, R.; Sukowijoyo, H.; Indriatmoko; Wijayanti, R.D.E.; Limantara, L. Encapsulation of brown seaweed pigment by freeze drying: Characterization and its stability during storage. Procedia Chem. 2015, 14, 353–360. [Google Scholar] [CrossRef]
- Corsetto, P.A.; Montorfano, G.; Zava, S.; Colombo, I.; Ingadottir, B.; Jonsdottir, R.; Sveinsdottir, K.; Rizzo, A.M. Characterization of antioxidant potential of seaweed extracts for enrichment of convenience food. Antioxidants 2020, 9, 249. [Google Scholar] [CrossRef] [PubMed]
- Everette, J.D.; Bryant, Q.M.; Green, A.M.; Abbey, Y.A.; Wangila, G.W.; Walker, R.B. Thorough study of reactivity of various compound classes toward the Folin−Ciocalteu reagent. J. Agric. Food Chem. 2010, 58, 8139–8144. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Rangel, J.C.; Benavides, J.; Heredia, J.B.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. The Folin–Ciocalteu assay revisited: Improvement of its specificity for total phenolic content determination. Anal. Methods 2013, 5, 5990–5999. [Google Scholar] [CrossRef]
- El-Hamidi, M.; Zaher, F.A.; El-Shami, S.M. Interaction of oilseed pigments and phospholipids in the determination of total phenolic compounds using the Folin-Ciocalteu reagent. Int. J. Pharm. Tech. Res. 2016, 9, 207–214. [Google Scholar]
- Heffernan, N.; Smyth, T.J.; Soler-Villa, A.; Fitzgerald, R.J.; Brunton, N.P. Phenolic content and antioxidant activity of fractions obtained from selected Irish macroalgae species (Laminaria digitata, Fucus serratus, Gracilaria gracilis and Codium fragile). Environ. Boil. Fishes 2015, 27, 519–530. [Google Scholar] [CrossRef]
- O’Sullivan, A.; O’Callaghan, Y.; O’Grady, M.; Queguineur, B.; Hanniffy, D.; Troy, D.; Kerry, J.; O’Brien, N. In vitro and cellular antioxidant activities of seaweed extracts prepared from five brown seaweeds harvested in spring from the west coast of Ireland. Food Chem. 2011, 126, 1064–1070. [Google Scholar] [CrossRef]
- Hermund, D.B.; Heung, S.Y.; Thomsen, B.R.; Akoh, C.C.; Jacobsen, C. Improving oxidative stability of skin-care emulsions with antioxidant extracts from brown alga Fucus vesiculosus. J. Am. Oil Chem. Soc. 2018, 95, 1509–1520. [Google Scholar] [CrossRef]
- Cikoš, A.-M.; Flanjak, I.; Bojanić, K.; Babić, S.; Čižmek, L.; Čož-Rakovac, R.; Jokić, S.; Jerković, I. Bioprospecting of coralline red alga Amphiroa rigida J.V. Lamouroux: Volatiles, fatty acids and pigments. Molecules 2021, 26, 520. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Hansen, C. Antioxidant capacity, phenolic content, and polysaccharide content of Lentinus edodes grown in whey permeate-based submerged culture. J. Food Sci. 2007, 73, M1–M8. [Google Scholar] [CrossRef]
- Goiris, K.; Muylaert, K.; Fraeye, I.; Foubert, I.; De Brabanter, J.; De Cooman, L. Antioxidant potential of microalgae in relation to their phenolic and carotenoid content. Environ. Boil. Fishes 2012, 24, 1477–1486. [Google Scholar] [CrossRef]
- Souza, B.W.S.; Cerqueira, M.A.; Martins, J.T.; Quintas, M.A.C.; Ferreira, A.C.S.; Teixeira, J.A.; Vicente, A.A. Antioxidant potential of two red seaweeds from the Brazilian coasts. J. Agric. Food Chem. 2011, 59, 5589–5594. [Google Scholar] [CrossRef]
- Antolovich, M.; Prenzler, P.D.; Patsalides, E.; McDonald, S.; Robards, K. Methods for testing antioxidant activity. Analyst 2002, 127, 183–198. [Google Scholar] [CrossRef] [PubMed]
- Gan, J.; Feng, Y.; He, Z.; Li, X.; Zhang, H. Correlations between antioxidant activity and alkaloids and phenols of maca (Lepidium meyenii). J. Food Qual. 2017, 2017, 1–10. [Google Scholar] [CrossRef]
- Yalçın, S.; Karakaş, Ö.; Okudan, E.Ş.; Başkan, K.S.; Çekiç, S.D.; Apak, R. HPLC Detection and antioxidant capacity determination of brown, red and green algal pigments in seaweed extracts. J. Chromatogr. Sci. 2021, 59, 325–337. [Google Scholar] [CrossRef] [PubMed]
- Kusmita, L.; Puspitaningrum, I.; Limantara, L. Identification, isolation and antioxidant activity of pheophytin from green tea (Camellia Sinensis (L.) Kuntze). Procedia Chem. 2015, 14, 232–238. [Google Scholar] [CrossRef]
- Di Paolo, C.; Seiler, T.-B.; Keiter, S.; Hu, M.; Muz, M.; Brack, W.; Hollert, H. The value of zebrafish as an integrative model in effect-directed analysis—A review. Environ. Sci. Eur. 2015, 27, 8. [Google Scholar] [CrossRef]
- Jayawardena, T.U.; Wang, L.; Sanjeewa, K.K.A.; Kang, S.I.; Lee, J.-S.; Jeon, Y.-J. Antioxidant potential of sulfated polysaccharides from Padina boryana; Protective effect against oxidative stress in in vitro and in vivo zebrafish model. Mar. Drugs 2020, 18, 212. [Google Scholar] [CrossRef]
- Oh, J.-Y.; Kim, E.-A.; Kang, S.I.; Yang, H.-W.; Ryu, B.; Wang, L.; Lee, J.-S.; Jeon, Y.-J. Protective effects of fucoidan isolated from Celluclast-Assisted Extract of Undaria pinnatifida Sporophylls against AAPH-Induced Oxidative Stress In Vitro and In Vivo zebrafish model. Molecules 2020, 25, 2361. [Google Scholar] [CrossRef]
- Kim, E.-A.; Lee, S.-H.; Ko, C.-I.; Cha, S.-H.; Kang, M.-C.; Kang, S.-M.; Ko, S.-C.; Lee, W.-W.; Ko, J.-Y.; Lee, J.-H.; et al. Protective effect of fucoidan against AAPH-induced oxidative stress in zebrafish model. Carbohydr. Polym. 2014, 102, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Oh, J.Y.; Kim, H.S.; Lee, W.; Cui, Y.; Lee, H.G.; Kim, Y.-T.; Jeon, Y.-J. Protective effect of polysaccharides from Celluclast-assisted extract of Hizikia fusiforme against hydrogen peroxide-induced oxidative stress in vitro in Vero cells and in vivo in zebrafish. Int. J. Biol. Macromol. 2018, 112, 483–489. [Google Scholar] [CrossRef]
- Sachindra, N.M.; Sato, E.; Maeda, H.; Hosokawa, M.; Niwano, Y.; Kohno, M.; Miyashita, K. Radical scavenging and singlet oxygen quenching activity of marine carotenoid fucoxanthin and its metabolites. J. Agric. Food Chem. 2007, 55, 8516–8522. [Google Scholar] [CrossRef] [PubMed]
- Heo, S.-J.; Ko, S.-C.; Kang, S.-M.; Kang, H.-S.; Kim, J.-P.; Kim, S.-H.; Lee, K.-W.; Cho, M.-G.; Jeon, Y.-J. Cytoprotective effect of fucoxanthin isolated from brown algae Sargassum siliquastrum against H2O2-induced cell damage. Eur. Food Res. Technol. 2008, 228, 145–151. [Google Scholar] [CrossRef]
- Kang, M.-C.; Lee, S.-H.; Lee, W.-W.; Kang, N.; Kim, E.-A.; Kim, S.Y.; Lee, D.H.; Kim, D.; Jeon, Y.-J. Protective effect of fucoxanthin isolated from Ishige okamurae against high-glucose induced oxidative stress in human umbilical vein endothelial cells and zebrafish model. J. Funct. Foods 2014, 11, 304–312. [Google Scholar] [CrossRef]
- Alghazwi, M.; Smid, S.; Musgrave, I.; Zhang, W. In vitro studies of the neuroprotective activities of astaxanthin and fucoxanthin against amyloid beta (Aβ1-42) toxicity and aggregation. Neurochem. Int. 2019, 124, 215–224. [Google Scholar] [CrossRef]
- Yu, J.; Lin, J.-J.; Yu, R.; He, S.; Wang, Q.-W.; Cui, W.; Zhang, J.-R. Fucoxanthin prevents H2O2-induced neuronal apoptosis via concurrently activating the PI3-K/Akt cascade and inhibiting the ERK pathway. Food Nutr. Res. 2017, 61, 1304678. [Google Scholar] [CrossRef]
- Lanfer-Marquez, U.M.; Barros, R.M.; Sinnecker, P. Antioxidant activity of chlorophylls and their derivatives. Food Res. Int. 2005, 38, 885–891. [Google Scholar] [CrossRef]
- Rizzo, C.; Genovese, G.; Morabito, M.; Faggio, C.; Pagano, M.; Spanò, A.; Zammuto, V.; Minicante, S.; Manghisi, A.; Cigala, R.; et al. Potential antibacterial activity of marine macroalgae against pathogens relevant for aquaculture and human health. J. Pure Appl. Microbiol. 2017, 11, 1695–1706. [Google Scholar] [CrossRef]
- Jerković, I.; Kranjac, M.; Marijanović, Z.; Šarkanj, B.; Cikoš, A.-M.; Aladić, K.; Pedisić, S.; Jokić, S. Chemical diversity of Codium bursa (Olivi) C. Agardh headspace compounds, volatiles, fatty acids and insight into its antifungal activity. Molecules 2019, 24, 842. [Google Scholar] [CrossRef] [PubMed]
- International Organization for Standardization. Animal and Vegetable Fats and Oils. Gas. Chromatography of Fatty Acid Methyl Esters. Part. 2: Preparation of Methyl Esters of Fatty Acids; EN ISO 12966-2:2017; International Organization for Standardization: Geneva, Switzerland, 2017. [Google Scholar]
- International Organization for Standardization. Animal and Vegetable Fats and Oils. Gas. Chromatography of Fatty Acid Methyl Esters. Part. 4: Determination by Capillary Gas. Chromatography; EN ISO 12966-4:2015; International Organization for Standardization: Geneva, Switzerland, 2015. [Google Scholar]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Brand-Williams, W.; Cuvelier, M.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Ou, B.; Hampsch-Woodill, M.; Flanagan, J.A.; Prior, R.L. High-throughput assay of oxygen radical absorbance capacity (ORAC) using a multichannel liquid handling system coupled with a microplate fluorescence reader in 96-well format. J. Agric. Food Chem. 2002, 50, 4437–4444. [Google Scholar] [CrossRef]
- Babić, S.; Čižmek, L.; Maršavelski, A.; Malev, O.; Pflieger, M.; Strunjak-Perović, I.; Popović, N.T.; Čož-Rakovac, R.; Trebše, P. Utilization of the zebrafish model to unravel the harmful effects of biomass burning during Amazonian wildfires. Sci. Rep. 2021, 11, 1–12. [Google Scholar] [CrossRef]
- OECD. Test. No. 236: Fish. Embryo Acute Toxicity (FET) Test. OECD Guidelines for the Testing of Chemicals, Section 2; OECD Publishing: Paris, France, 2013. [Google Scholar]
- Maes, J.; Verlooy, L.; Buenafe, O.E.; De Witte, P.A.M.; Esguerra, C.V.; Crawford, A.D. Evaluation of 14 organic solvents and carriers for screening applications in zebrafish embryos and larvae. PLoS ONE 2012, 7, e43850. [Google Scholar] [CrossRef]
- Palčić, A.; Babić, S.; Maršavelski, A.; Galić, M.; Popović, N.T.; Perović, I.S.; Čož-Rakovac, R.; Bronić, J.; Valtchev, V. Nanosized zeolite beta—Determining the safety of usage by zebrafish Danio rerio embryos. Microporous Mesoporous Mater. 2020, 299, 110103. [Google Scholar] [CrossRef]
- Eruslanov, E.; Kusmartsev, S. Identification of ROS using oxidized DCFDA and flow-cytometry. Methods Mol. Biol. 2010, 594, 57–72. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, 10th ed.; M07-A10; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2015. [Google Scholar]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi, 3rd ed.; M38; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compound | RI | A Av ± SD | B Av ± SD | C Av ± SD | D Av ± SD |
|---|---|---|---|---|---|---|
| 1. | Hexanal S | <900 | - | 1.14 ± 0.03 | - | 0.72 ± 0.01 |
| 2. | 3-Methylbutanoic acid S | <900 | - | 0.25 ± 0.02 | - | - |
| 3. | Heptanal S | 901 | - | 1.41 ± 0.02 | - | 0.60 ± 0.02 |
| 4. | (E)-Hept-2-enal | 962 | - | 0.35 ± 0.01 | - | - |
| 5. | Benzaldehyde S | 968 | - | 1.43 ± 0.04 | - | 1.18 ± 0.03 |
| 6. | Oct-1-en-3-one S | 983 | - | 0.18 ± 0.01 | - | 0.14 ± 0.01 |
| 7. | Oct-1-en-3-ol S | 983 | - | 0.87 ± 0.02 | - | 0.52 ± 0.01 |
| 8. | 6-Methylhept-5-en-2-one S | 988 | - | 0.47 ± 0.02 | - | - |
| 9. | Octanal S | 1003 | - | 0.56 ± 0.01 | - | - |
| 10. | Benzyl alcohol S | 1044 | - | 19.67 ± 0.42 | - | 15.75 ± 0.51 |
| 11. | Nonanal S | 1106 | - | 3.36 ± 0.05 | - | 1.98 ± 0.02 |
| 12. | Tridecane S | 1300 | - | 0.89 ± 0.01 | - | 1.66 ± 0.02 |
| 13. | Pentadec-1-ene S | 1493 | 17.98 ± 0.82 | - | 11.00 ± 0.50 | - |
| 14. | Pentadecane S | 1500 | 61.90 ± 1.51 | 60.27 ± 2.00 | 71.55 ± 1.98 | 71.43 ± 1.84 |
| 15. | Tridecanal S | 1511 | - | 1.13 ± 0.02 | 5.45 ± 0.10 | 1.03 ± 0.03 |
| 16. | (E)-Pentadec-7-ene | 1517 | 9.12 ± 0.80 | - | - | - |
| No. | Compound | RI | E Av ± SD | F Av ± SD |
|---|---|---|---|---|
| 1. | (E)-Hex-2-enal S | <900 | - | 1.13 ± 0.03 |
| 2. | Heptanal S | 901 | - | 0.86 ± 0.05 |
| 3. | (Z)-Hept-2-enal S | 961 | 0.32 ± 0.01 | 0.42 ± 0.02 |
| 4. | Benzaldehyde S | 968 | 0.09 ± 0.01 | 0.69 ± 0.02 |
| 5. | Oct-1-en-3-ol S | 983 | - | 0.73 ± 0.03 |
| 6. | Octan-2,3-dione S | 985 | - | 0.24 ± 0.01 |
| 7. | 2-Pentylfuran S | 993 | 0.15 ± 0.01 | 0.70 ± 0.02 |
| 8. | (E,E)-Hepta-2,4-dienal S | 999 | - | 0.28 ± 0.01 |
| 9. | Octanal S | 1003 | - | 0.23 ± 0.01 |
| 10. | (E,Z)-Hepta-2,4-dienal | 1007 | - | 0.13 ± 0.01 |
| 11. | (Z,Z)-Hepta-2,4-dienal S | 1015 | - | 0.63 ± 0.05 |
| 12. | 2,2,6-Trimethylcyclohexanone S | 1040 | 0.01 ± 0.01 | 0.22 ± 0.03 |
| 13. | Benzyl alcohol S | 1044 | 0.62 ± 0.05 | 0.67 ± 0.05 |
| 14. | Phenylacetaldehyde S | 1050 | 0.54 ± 0.03 | 0.87 ± 0.02 |
| 15. | (E)-Oct-2-enal S | 1062 | - | 0.45 ± 0.02 |
| 16. | (E)-Oct-2-en-1-ol S | 1073 | - | 0.32 ± 0.02 |
| 17. | (E,E)-Octa-3,5-dien-2-one | 1074 | - | 0.44 ± 0.08 |
| 18. | Octan-1-ol S | 1076 | - | 0.18 ± 0.01 |
| 19. | Nonan-2-one S | 1094 | - | 0.95 ± 0.03 |
| 20. | (E,Z)-Octa-3,5-dien-2-one | 1095 | - | 0.20 ± 0.01 |
| 21. | Nonanal S | 1106 | 0.52 ± 0.02 | 3.48 ± 0.08 |
| 22. | 2,6-Dimethylcyclohexanol S | 1111 | - | 0.74 ± 0.02 |
| 23. | 4-Ketoisophorone S | 1148 | 0.89 ± 0.03 | 1.36 ± 0.05 |
| 24. | (E,E)-Nona-2,6-dienal S | 1157 | - | 0.09 ± 0.01 |
| 25. | 5-Methylundecane | 1159 | - | 0.14 ± 0.01 |
| 26. | (E)-Non-2-enal S | 1163 | - | 0.26 ± 0.01 |
| 27. | Dodec-1-ene S | 1193 | 0.03 ± 0.01 | 0.20 ± 0.03 |
| 28. | Decanal S | 1207 | - | 0.27 ± 0.02 |
| 29. | β-Cyclocitral S | 1224 | 0.15 ± 0.01 | 0.75 ± 0.04 |
| 30. | Benzothiazole S | 1228 | 0.14 ± 0.01 | 0.27 ± 0.01 |
| 31. | Benzenepropanenitrile | 1245 | - | 0.45 ± 0.03 |
| 32. | β-Homocyclocitral S | 1260 | - | 0.31 ± 0.02 |
| 33. | (E)-Dec-2-enal S | 1265 | 3.48 ± 0.05 | 1.34 ± 0.07 |
| 34. | (E,Z)-Deca-2,4-dienal S | 1295 | 0.32 ± 0.01 | 0.51 ± 0.02 |
| 35. | 1H-Indole S | 1301 | 0.15 ± 0.01 | 0.01 ± 0.01 |
| 36. | Undecanal S | 1308 | - | 0.64 ± 0.03 |
| 37. | (E,E)-Deca-2,4-dienal S | 1320 | 0.66 ± 0.01 | 2.23 ± 0.04 |
| 38. | Tetradec-1-ene S | 1393 | 0.12 ± 0.01 | - |
| 39. | Dodecanal S | 1410 | - | 3.16 ± 0.01 |
| 40. | α-Ionone S | 1430 | - | 0.79 ± 0.01 |
| 41. | (E)-Geranylacetone S | 1455 | - | 0.50 ± 0.02 |
| 42. | Dodecan-1-ol S | 1480 | - | 1.47 ± 0.05 |
| 43. | β-Ionone S | 0.77 ± 0.04 | 5.80 ± 0.06 | |
| 44. | Pentadec-1-ene S | 1493 | 19.32 ± 1.21 | 1.54 ± 0.02 |
| 45. | Pentadecane S | 1500 | 5.87 ± 0.04 | 3.28 ± 0.03 |
| 46. | Tridecanal S | 1511 | 2.67 ± 0.01 | 18.51 ± 1.10 |
| 47. | (E)-Pentadec-7-ene | 1517 | 8.35 ± 0.12 | 3.16 ± 0.08 |
| 48. | Tridecan-1-ol S | 1581 | - | 0.59 ± 0.02 |
| 49. | Tetradecanal S | 1614 | 0.51 ± 0.01 | 1.01 ± 0.05 |
| 50. | (E)-Heptadec-8-ene | 1679 | 0.61 ± 0.01 | - |
| 51. | γ-Dodecalactone S | 1681 | 3.31 ± 0.08 | 2.20 ± 0.21 |
| 52. | (Z)-Heptadec-3-ene | 1688 | 6.07 ± 0.51 | 1.23 ± 0.11 |
| 53. | Heptadec-1-ene S | 1694 | 5.05 ± 0.51 | 0.91 ± 0.02 |
| 54. | Heptadecane S | 1700 | 0.95 ± 0.01 | - |
| 55. | Pentadecanal S | 1716 | 1.34 ± 0.22 | 4.09 ± 0.11 |
| 56. | Tetradecanoic acid S | 1778 | 3.07 ± 0.03 | 4.77 ± 0.11 |
| 57. | Hexahydrofarnesyl acetone S | 1848 | 2.23 ± 0.11 | 1.59 ± 0.10 |
| 58. | Hexadecanoic acid S | 1973 | - | 2.69 ± 0.08 |
| 59. | Hexadecanal S | 1917 | 1.31 ± 0.04 | - |
| 60. | Dibutyl phthalate S | 1961 | 0.33 ± 0.05 | 0.20 ± 0.01 |
| 61. | (Z,Z,Z)-Octadeca-9,12,15-trien-1-ol (Linolenic alcohol) | 2042 | 12.89 ± 1.01 | 5.08 ± 0.21 |
| 62. | Phytol S | 2115 | 7.19 ± 0.82 | 4.06 ± 0.11 |
| No. | Fatty Acid | Av ± SD (%) | |
|---|---|---|---|
| 1. | C12:0 | Lauric acid S | 0.06 ± 0.01 |
| 2. | C14:0 | Myristic acid S | 10.51 ± 0.07 |
| 3. | C15:0 | Pentadecyclic acid S | 0.18 ± 0.00 |
| 4. | C16:0 | Palmitic acid S | 8.10 ± 0.07 |
| 5. | C18:0 | Stearic acid S | 1.35 ± 0.02 |
| 6. | C20:0 | Arachidic acid S | 0.43 ± 0.11 |
| 7. | C24:0 | Lignoceric acid S | 1.42 ± 0.33 |
| Total saturated fatty acids (SFA) | 22.03 | ||
| 8. | C14:1 | Myristoleic acid S | 0.20 ± 0.00 |
| 9. | C16:1 | Palmitoleic acid S | 0.99 ± 0.07 |
| 10. | C18:1n9t + C18:1n9c | cis-Oleic acid+trans-Oleic acid S | 42.28 ± 0.24 |
| 11. | C20:1n9 | Eicosenoic acid S | 0.65 ± 0.10 |
| 12. | C22:1n9 | Erucic acid S | 0.32 ± 0.01 |
| Total monounsaturated fatty acids (MUFA) | 44.43 | ||
| 13. | C18:2n6c | cis-Linoleic acid S | 6.51 ± 0.07 |
| 14. | C18:3n3 | α-linolenic acid S | 2.13 ± 0.93 |
| 15. | C18:3n6 | γ-Linolenic acid S | 0.42 ± 0.02 |
| 16. | C20:2n6 | Eicosadienoic acid S | 1.51 ± 0.08 |
| 17. | C20:3n3 | Eicosatrienoic acid S | 0.26 ± 0.06 |
| 18. | C20:3n6 (DGLA) | Dihomo-γ-linolenic acid S | 2.96 ± 0.03 |
| 19. | C20:4n6 | Arachidonic acid S | 15.00 ± 0.05 |
| 20. | C20:5n3 (EPA) | Eicosapentaenoic acid S | 3.71 ± 0.02 |
| 21. | C22:2 | Docosadienoic acid S | 1.24 ± 0.04 |
| Total polyunsaturated fatty acids (PUFA) | 33.71 | ||
| Total ω3 fatty acids | 6.09 | ||
| Total ω6 fatty acids | 27.62 |
| No. | Compound | Rt (min) | Elemental Composition | m/z (Δ, ppm) | Peak Area (Arbitrary Units) | |
|---|---|---|---|---|---|---|
| F3 | F4 | |||||
| 1. | 1,3-Dihydroxy-2-propanyl 5,8,11,14-icosatetraenoate | 13.70 | C23H38O4 | 379.2844 (0.3) | 2045 | 26 |
| 2. | 2,3-Dihydroxypropyl palmitate | 14.05 | C19H38O4 | 331.2843 (0.0) | 13,300 | 19,400 |
| 3. | 2,3-Dihydroxypropyl 9-octadecenoate | 14.34 | C21H40O4 | 357.2998 (−0.4) | 9240 | 330 |
| 4. | 2,3-Dihydroxypropyl stearate | 15.14 | C21H42O4 | 359.3154 (−0.5) | 27,100 | 17,900 |
| 5. | Isoamijiol oxidation product * | 14.55 | C20H30O2 | 303.2312 (−2.2) | 1100 | 610 |
| 6. | (3aR,4aR,6S,8aR)-1-Isopropyl-3a,8a-dimethyl-5-methylene-2,3a,4,5,6,7,8,8a,9,10-decahydrobenzo[f]azulene-4a,6(3H)-diol (Isoamijiol) | 14.54 | C20H32O2 | 305.2469 (−2.0) | 1800 | 60 |
| 7. | Fucoxanthin | 15.50 | C42H58O6 | 659.4299 (−1.1) | 1340 | 10 |
| 8. | 2-Hydroxypropyl palmitate | 15.55 | C19H38O3 | 315.2884 (−3.1) | 197 | 788 |
| 9. | 2-Hydroxypropyl stearate | 16.55 | C21H42O3 | 343.3200 (−2.0) | 482 | 1690 |
| 10. | (3β,6α)-14-Methylergosta-8,24(28)-diene-3,6-diol (few isomers) ** | 16.25 | C29H48O2 | 429.3723 (−0.9) | 220 | 18 |
| 11. | 13-Docosenamide | 16.67 | C22H43NO | 338.3412 (−1.6) | 7550 | 3650 |
| 12. | (2E)-3-[21-(Methoxycarbonyl)-4,8,13,18-tetramethyl-20-oxo-9,14-divinyl-3,4-didehydro-3-24,25-dihydrophorbinyl]acrylic acid | 16.71 | C35H30N4O5 | 587.2273 (−2.7) | 1820 | 64 |
| 13. | Pheophorbide a | 16.73 | C35H36N4O5 | 593.2741 (−2.9) | 1430 | 106 |
| 14. | 3-[(21R)-21-(Methoxycarbonyl)-4,8,13,18-tetramethyl-20-oxo-9,14-divinyl-3,4-didehydro-3--24,25-dihydrophorbinyl]propanoic acid | 16.77 | C35H32N4O5 | 589.2422 (−4.0) | 1240 | 62 |
| 15. | (3β)-3-Hydroxystigmast-5-en-7-one | 18.22 | C29H48O2 | 429.3720 (−1.6) | 6380 | 205 |
| 16. | (3β,20R,22E,24S)-Stigmasta-5,22-dien-3-ol (β-Stigmasterol) | 18.39 | C29H48O | 395.3664 *** (−2.1) | 1950 | 28,670 |
| 17. | (2S)-1-Hydroxy-3-(tetradecanoyloxy)-2-propanyl (9Z)-9-octadecenoate | 20.29 | C35H66O5 | 567.4972 (−1.9) | 849 | 2410 |
| 18. | 3-Hydroxy-1,2-propanediyl bis(9-octadecenoate) | 21.10 | C39H72O5 | 621.5435 (−2.8) | 66 | 1270 |
| 19. | 3-Hydroxy-2-(palmitoyloxy)propyl stearate | 21.60 | C37H72O5 | 597.5433 (−3.3) | 229 | 309 |
| 20. | Methyl (3R,10Z,14Z,20Z,22S,23S)-12-ethyl-3-hydroxy-13,18,22,27-tetramethyl-5-oxo-23-(3-oxo-3-{[(2E,7R,11R)-3,7,11,15-tetramethyl-2-hexadecen-1-yl]oxy}propyl)-17-vinyl-4-oxa-8,24,25,26-tetraazahexacycl; o[19.2.1.16,9.111,14.116,19.02,7]heptacosa-1(24),2(7),6(27),8,10,12,14,16,18,20-decaene-3-carboxylate | 21.56 | C55H74N4O7 | 903.5610 (−2.2) | - | 1970 |
| 21. | 3-Phorbinepropanoic acid, 9-acetyl-14-ethylidene-13,14-dihydro-21-(methoxycarbonyl)-4,8,13,18-tetramethyl-20-oxo-, 3,7,11,15-tetramethyl-2-hexadecen-1-yl ester | 22.48 | C55H74N4O6 | 887.5661 (−2.3) | 17 | 2960 |
| 22. | 3-Phorbinepropanoic acid, 3,4-didehydro-9-ethenyl-14-ethyl-24,25-dihydro-21-(methoxycarbonyl)-4,8,13,18-tetramethyl-20-oxo-, (2E)-3,7,11,15-tetramethyl-2-hexadecen-1-yl ester | 22.53 | C55H72N4O5 | 869.5550 (−2.9) | - | 300 |
| 23. | Pheophytin a | 22.75 | C55H74N4O5 | 871.5711 (−2.4) | 288 | 20,600 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jerković, I.; Cikoš, A.-M.; Babić, S.; Čižmek, L.; Bojanić, K.; Aladić, K.; Ul’yanovskii, N.V.; Kosyakov, D.S.; Lebedev, A.T.; Čož-Rakovac, R.; et al. Bioprospecting of Less-Polar Constituents from Endemic Brown Macroalga Fucus virsoides J. Agardh from the Adriatic Sea and Targeted Antioxidant Effects In Vitro and In Vivo (Zebrafish Model). Mar. Drugs 2021, 19, 235. https://doi.org/10.3390/md19050235
Jerković I, Cikoš A-M, Babić S, Čižmek L, Bojanić K, Aladić K, Ul’yanovskii NV, Kosyakov DS, Lebedev AT, Čož-Rakovac R, et al. Bioprospecting of Less-Polar Constituents from Endemic Brown Macroalga Fucus virsoides J. Agardh from the Adriatic Sea and Targeted Antioxidant Effects In Vitro and In Vivo (Zebrafish Model). Marine Drugs. 2021; 19(5):235. https://doi.org/10.3390/md19050235
Chicago/Turabian StyleJerković, Igor, Ana-Marija Cikoš, Sanja Babić, Lara Čižmek, Krunoslav Bojanić, Krunoslav Aladić, Nikolay V. Ul’yanovskii, Dmitry S. Kosyakov, Albert T. Lebedev, Rozelindra Čož-Rakovac, and et al. 2021. "Bioprospecting of Less-Polar Constituents from Endemic Brown Macroalga Fucus virsoides J. Agardh from the Adriatic Sea and Targeted Antioxidant Effects In Vitro and In Vivo (Zebrafish Model)" Marine Drugs 19, no. 5: 235. https://doi.org/10.3390/md19050235
APA StyleJerković, I., Cikoš, A.-M., Babić, S., Čižmek, L., Bojanić, K., Aladić, K., Ul’yanovskii, N. V., Kosyakov, D. S., Lebedev, A. T., Čož-Rakovac, R., Trebše, P., & Jokić, S. (2021). Bioprospecting of Less-Polar Constituents from Endemic Brown Macroalga Fucus virsoides J. Agardh from the Adriatic Sea and Targeted Antioxidant Effects In Vitro and In Vivo (Zebrafish Model). Marine Drugs, 19(5), 235. https://doi.org/10.3390/md19050235