
Biochemical Properties and Anti-Biofilm Activity of Chitosan-Immobilized Papain

 ,
,  and
and
Abstract
1. Introduction
2. Results
2.1. Immobilization of Papain on Chitosan Matrixes
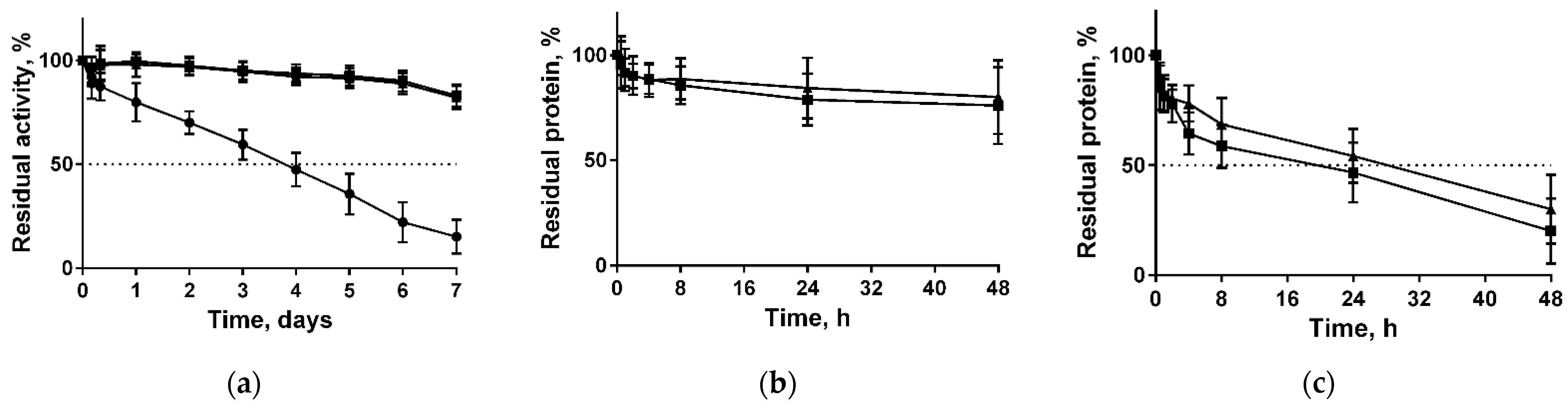
2.2. Catalytic Properties of the Native and Immobilized Papain
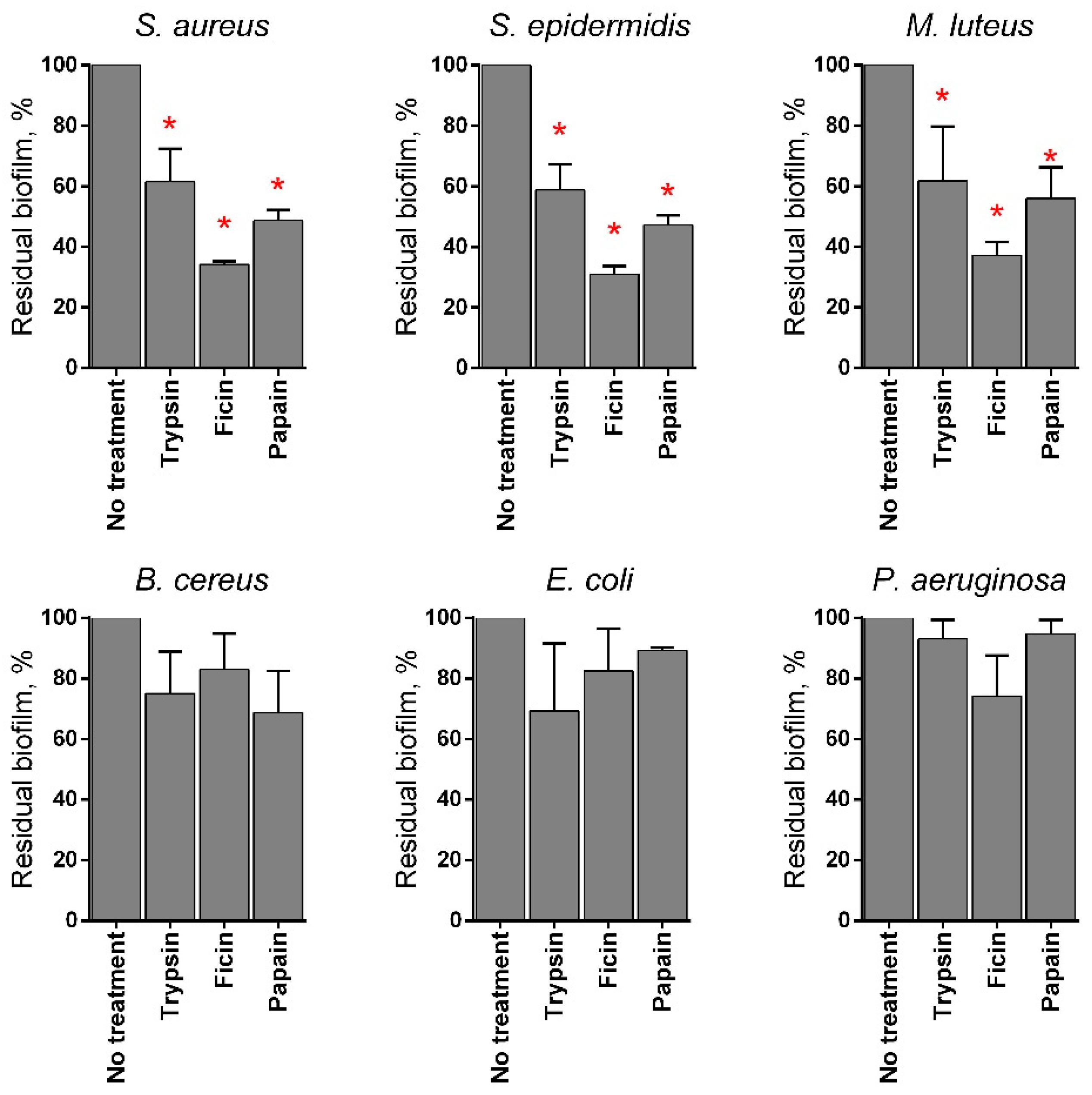
2.3. Anti-Biofilm Properties of Immobilized Papain
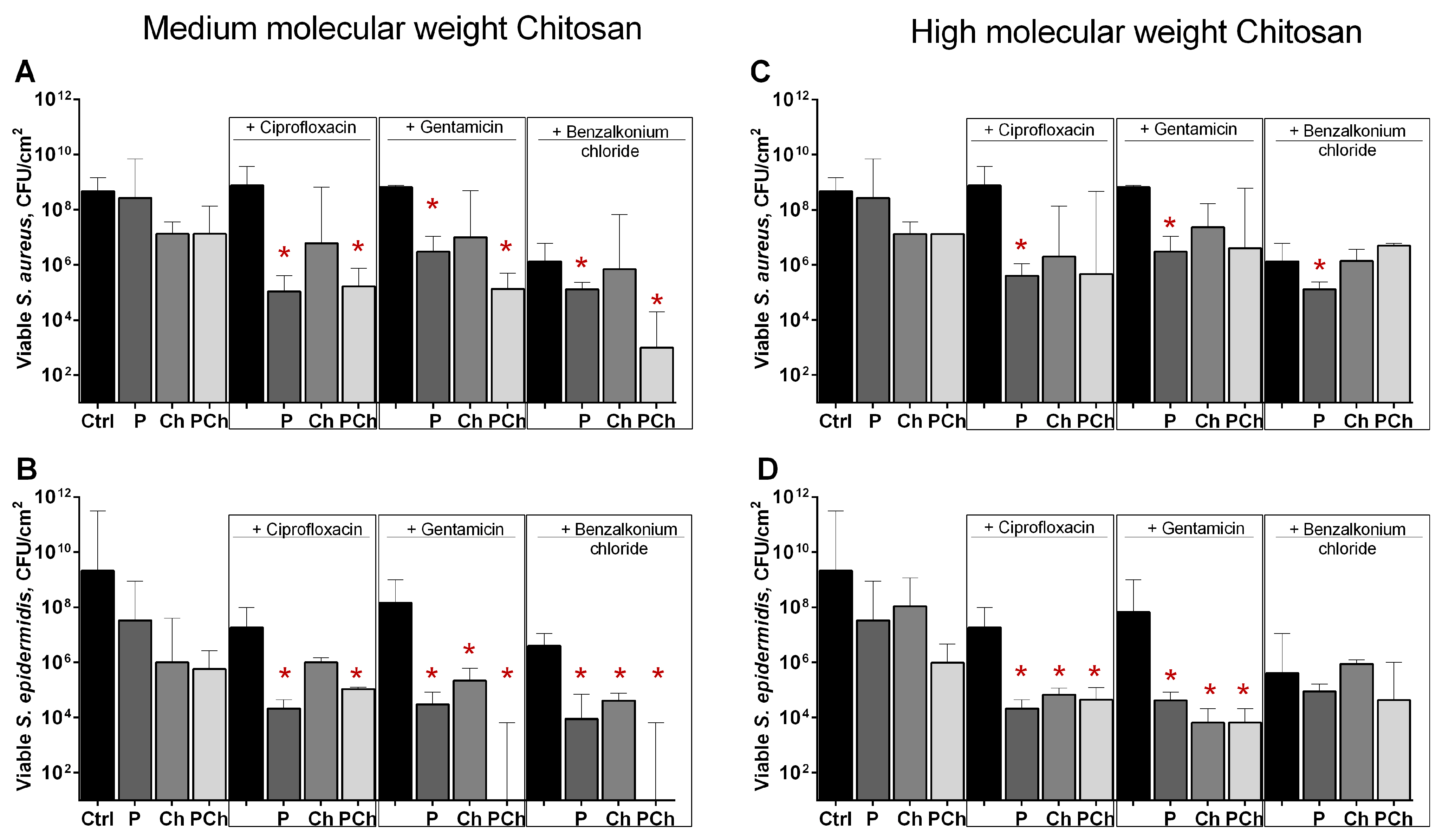
2.4. Increasing the Efficiency of Antimicrobials Against Staphylococcal Biofilms by Soluble and Citosan-Immobilized Papain
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals
5.2. Papain Immobilization on Chitosans
5.3. Proteolytic Activity Measurements
5.4. Kinetic Properties
5.5. Bacterial Strains and Growth Conditions
5.6. Determination of Minimum Inhibitory Concentration (MIC)
5.7. Biofilm Assays
5.8. Statistical Analysis
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Haesaerts, S.; Buitrago, J.A.R.; Loris, R.; Baeyens-Volant, D.; Azarkan, M. Crystallization and preliminary X-ray analysis of four cysteine proteases from Ficus carica latex. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2015, 71, 459–465. [Google Scholar] [CrossRef]
- Zare, H.; Moosavi-Movahedi, A.A.; Salami, M.; Mirzaei, M.; Saboury, A.A.; Sheibani, N. Purification and autolysis of the ficin isoforms from fig (Ficus carica cv. Sabz) latex. Phytochemistry 2013, 87, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Lim, T.K. Edible Medicinal and Non-Medicinal Plants: Volume 1, Fruits; Springer: Berlin/Heidelberg, Germany, 2012; pp. 1–835. [Google Scholar] [CrossRef]
- Mohr, T.; Desser, L. Plant proteolytic enzyme papain abrogates angiogenic activation of human umbilical vein endothelial cells (HUVEC) in vitro. BMC Complementary Altern. Med. 2013, 13. [Google Scholar] [CrossRef]
- Vootukuri, R.S.; Phillpot, M.P.; Trigiante, G. Fluorimetric ex vivo quantification of protease debriding efficacy on natural substrate. Wound Repair Regen. 2020, 28, 844–847. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, N.F.; Cunha, A.P.; Ricardo, N.; Freire, R.S.; Vieira, L.D.P.; Santa Brigida, A.I.; Borges, M.D.; Rosa, M.D.; Vieira, R.S.; Andrade, F.K. Papain immobilization on heterofunctional membrane bacterial cellulose as a potential strategy for the debridement of skin wounds. Int. J. Biol. Macromol. 2020, 165, 3065–3077. [Google Scholar] [CrossRef] [PubMed]
- Tadikonda, A.; Pentapati, K.C.; Urala, A.S.; Acharya, S. Anti-plaque and anti-gingivitis effect of Papain, Bromelain, Miswak and Neem containing dentifrice: A randomized controlled trial. J. Clin. Exp. Dent. 2017, 9, e649–e653. [Google Scholar] [CrossRef] [PubMed]
- Mugita, N.; Nambu, T.; Takahashi, K.; Wang, P.L.; Komasa, Y. Proteases, actinidin, papain and trypsin reduce oral biofilm on the tongue in elderly subjects and in vitro. Arch. Oral Biol. 2017, 82, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, S.H.; Mohamed, M.S.; Khalil, M.S.; Mohamed, W.S.; Mabrouk, M.I. Antibiofilm activity of papain enzyme against pathogenic Klebsiella pneumoniae. J. Appl. Pharm. Sci. 2018, 6, 163–168. [Google Scholar] [CrossRef]
- Song, Y.J.; Yu, H.H.; Kim, Y.J.; Lee, N.K.; Paik, H.D. The use of papain for the removal of biofilms formed by pathogenic Staphylococcus aureus and Campylobacter jejuni. LWT Food Sci. Technol. 2020, 127. [Google Scholar] [CrossRef]
- Atacan, K.; Ozacar, M. Investigation of antibacterial properties of novel papain immobilized on tannic acid modified Ag/CuFe2O4 magnetic nanoparticles. Int. J. Biol. Macromol. 2018, 109, 720–731. [Google Scholar] [CrossRef] [PubMed]
- Trizna, E.; Baydamshina, D.; Kholyavka, M.; Sharafutdinov, I.; Hairutdinova, A.; Khafizova, F.; Zakirova, E.; Hafizov, R.; Kayumov, A. Soluble and immobilized papain and trypsin as destroyers of bacterial biofilms. Genes Cells 2016, 10, 106–112. [Google Scholar]
- Garcia-Galan, C.; Berenguer-Murcia, A.; Fernandez-Lafuente, R.; Rodrigues, R.C. Potential of Different Enzyme Immobilization Strategies to Improve Enzyme Performance. Adv. Synth. Catal. 2011, 353, 2885–2904. [Google Scholar] [CrossRef]
- Hanefeld, U.; Cao, L.Q.; Magner, E. Enzyme immobilisation: Fundamentals and application. Chem. Soc. Rev. 2013, 42, 6211–6212. [Google Scholar] [CrossRef]
- Homaei, A.; Samari, F. Investigation of activity and stability of papain by adsorption on multi-wall carbon nanotubes. Int. J. Biol. Macromol. 2017, 105, 1630–1635. [Google Scholar] [CrossRef] [PubMed]
- Bolivar, J.M.; Mateo, C.; Rocha-Martin, J.; Cava, F.; Berenguer, J.; Fernandez-Lafuente, R.; Guisana, J.M. The adsorption of multimeric enzymes on very lowly activated supports involves more enzyme subunits: Stabilization of a glutamate dehydrogenase from Thermus thermophilus by immobilization on heterofunctional supports. Enzym. Microb. Technol. 2009, 44, 139–144. [Google Scholar] [CrossRef]
- Min, K.; Yoo, Y.J. Recent progress in nanobiocatalysis for enzyme immobilization and its application. Biotechnol. Bioprocess. Eng. 2014, 19, 553–567. [Google Scholar] [CrossRef]
- Rodrigues, R.C.; Ortiz, C.; Berenguer-Murcia, A.; Torres, R.; Fernandez-Lafuente, R. Modifying enzyme activity and selectivity by immobilization. Chem. Soc. Rev. 2013, 42, 6290–6307. [Google Scholar] [CrossRef] [PubMed]
- Cipolatti, E.P.; Valerio, A.; Henriques, R.O.; Moritz, D.E.; Ninow, J.L.; Freire, D.M.G.; Manoel, E.A.; Fernandez-Lafuente, R.; de Oliveira, D. Nanomaterials for biocatalyst immobilization—State of the art and future trends. RSC Adv. 2016, 6, 104675–104692. [Google Scholar] [CrossRef]
- Gobom, J.; Nordhoff, E.; Ekman, R.; Roepstorff, P. Rapid micro-scale proteolysis of proteins for MALDI-MS peptide mapping using immobilized trypsin. Int. J. Mass Spectrom. 1997, 169, 153–163. [Google Scholar] [CrossRef]
- He, Z.Y.; Nie, H.L.; Zhou, Y.T.; Zhu, L.M. Analysis of papain adsorption on nylon-based immobilized copper ion affinity membrane. J. Biotechnol. 2008, 136, S307. [Google Scholar] [CrossRef]
- Zhang, H.T.; Nie, H.L.; Li, S.B.; Xue, Y.; Zhu, L.M. Electrospun nylon nanofiber as affinity membrane for papain adsorption. J. Biotechnol. 2008, 136, S416. [Google Scholar] [CrossRef]
- Xue, Y.; Nie, H.L.; Zhu, L.M.; Li, S.B.; Zhang, H.T. Immobilization of Modified Papain with Anhydride Groups on Activated Cotton Fabric. Appl. Biochem. Biotechnol. 2010, 160, 109–121. [Google Scholar] [CrossRef]
- Alpay, P.; Uygun, D.A. Usage of immobilized papain for enzymatic hydrolysis of proteins. J. Mol. Catal. B Enzym. 2015, 111, 56–63. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Lu, Y.H.; Ma, C.H.; Tao, W.W.; Zhu, J.J.; Zhang, X. A novel elastic liposome for skin delivery of papain and its application on hypertrophic scar. Biomed. Pharmacother. 2017, 87, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Homaei, A.A.; Sajedi, R.H.; Sariri, R.; Seyfzadeh, S.; Stevanato, R. Cysteine enhances activity and stability of immobilized papain. Amino Acids 2010, 38, 937–942. [Google Scholar] [CrossRef] [PubMed]
- Li, F.Y.; Xing, Y.J.; Ding, X. Immobilization of papain on cotton fabric by sol-gel method. Enzym. Microb. Technol. 2007, 40, 1692–1697. [Google Scholar] [CrossRef]
- Manosroi, A.; Chankhampan, C.; Manosroi, W.; Manosroi, J. Toxicity Reduction and MMP-2 Stimulation of Papain and Bromelain Loaded in Elastic Niosomes. J. Biomed. Nanotechnol. 2012, 8, 720–729. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Cortez, I.E.; Romero-Garcia, J.; Gonzalez-Gonzalez, V.; Garcia-Gutierrez, D.I.; Garza-Navarro, M.A.; Cruz-Silva, R. Encapsulation and immobilization of papain in electrospun nanofibrous membranes of PVA cross-linked with glutaraldehyde vapor. Mater. Sci. Eng. C Mater. Biol. Appl. 2015, 52, 306–314. [Google Scholar] [CrossRef]
- Muller, C.; Perera, G.; Konig, V.; Bernkop-Schnurch, A. Development and in vivo evaluation of papain-functionalized nanoparticles. Eur. J. Pharm. Biopharm. 2014, 87, 125–131. [Google Scholar] [CrossRef]
- Pinto, C.; Lopes, P.S.; Sarruf, F.D.; Polakiewicz, B.; Kaneko, T.M.; Baby, A.R.; Velasco, M.V.R. Comparative study of the stability of free and modified papain incorporated in topical formulations. Braz. J. Pharm. Sci. 2011, 47, 751–760. [Google Scholar] [CrossRef]
- Sangeetha, K.; Abraham, T.E. Chemical modification of papain for use in alkaline medium. J. Mol. Catal. B Enzym. 2006, 38, 171–177. [Google Scholar] [CrossRef]
- Sim, Y.C.; Lee, S.G.; Lee, D.C.; Kang, B.Y.; Park, K.M.; Lee, J.Y.; Kim, M.S.; Chang, I.S.; Rhee, J.S. Stabilization of papain and lysozyme for application to cosmetic products. Biotechnol. Lett. 2000, 22, 137–140. [Google Scholar] [CrossRef]
- Chatla, S.; Sivareddy, C.; Basaveswarao, M.; Harshita, E.; Yashwanthi, P. Anti-bacterial, anti-fungal and analgesic activity of papain conjugated quercetin. Eur. J. Biomed. 2017, 4, 280–285. [Google Scholar]
- Adriano, W.S.; Filho, E.H.C.; Silva, J.A.; Giordano, R.L.C.; Goncalves, L.R.B. Stabilization of penicillin G acylase by immobilization on glutaraldehyde-activated chitosan. Braz. J. Chem. Eng. 2005, 22, 529–538. [Google Scholar] [CrossRef]
- Manrich, A.; Galvao, C.M.A.; Jesus, C.D.F.; Giordano, R.C.; Giordano, R.L.C. Immobilization of trypsin on chitosan gels: Use of different activation protocols and comparison with other supports. Int. J. Biol. Macromol. 2008, 43, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Mei, S.; Han, P.P.; Wu, H.; Shi, J.F.; Tang, L.; Jiang, Z.G. One-pot fabrication of chitin-shellac composite microspheres for efficient enzyme immobilization. J. Biotechnol. 2018, 266, 1–8. [Google Scholar] [CrossRef]
- Silva, J.A.; Macedo, G.P.; Rodrigues, D.S.; Giordano, R.L.C.; Goncalves, L.R.B. Immobilization of Candida antarctica lipase B by covalent attachment on chitosan-based hydrogels using different support activation strategies. Biochem. Eng. J. 2012, 60, 16–24. [Google Scholar] [CrossRef]
- Elchinger, P.H.; Delattre, C.; Faure, S.; Roy, O.; Badel, S.; Bernardi, T.; Taillefumier, C.; Michaud, P. Immobilization of proteases on chitosan for the development of films with anti-biofilm properties. Int. J. Biol. Macromol. 2015, 72, 1063–1068. [Google Scholar] [CrossRef]
- Khor, E.; Lim, L.Y. Implantable applications of chitin and chitosan. Biomaterials 2003, 24, 2339–2349. [Google Scholar] [CrossRef]
- Krajewska, B. Application of chitin- and chitosan-based materials for enzyme immobilizations: A review. Enzym. Microb. Technol. 2004, 35, 126–139. [Google Scholar] [CrossRef]
- Ozturk, E.; Agalar, C.; Kececi, K.; Denkbas, E.B. Preparation and characterization of ciprofloxacin-loaded alginate/chitosan sponge as a wound dressing material. J. Appl. Polym. Sci. 2006, 101, 1602–1609. [Google Scholar] [CrossRef]
- Urrutia, P.; Bernal, C.; Escobar, S.; Santa, C.; Mesa, M.; Wilson, L.; Illanes, A. Influence of Chitosan Derivatization on Its Physicochemical Characteristics and Its Use as Enzyme Support. J. Appl. Polym. Sci. 2014, 131. [Google Scholar] [CrossRef]
- Kumar, M.; Muzzarelli, R.A.A.; Muzzarelli, C.; Sashiwa, H.; Domb, A.J. Chitosan chemistry and pharmaceutical perspectives. Chem. Rev. 2004, 104, 6017–6084. [Google Scholar] [CrossRef] [PubMed]
- Li, H.B.; Du, Y.M.; Xu, Y.M. Adsorption and complexation of chitosan wet-end additives in papermaking systems. J. Appl. Polym. Sci. 2004, 91, 2642–2648. [Google Scholar] [CrossRef]
- Muzzarelli, R.A.A.; Muzzarelli, C. Chitosan chemistry: Relevance to the biomedical sciences. Polysacch. 1 Struct. Charact. Use 2005, 186, 151–209. [Google Scholar] [CrossRef]
- Nahar, S.; Mizan, M.F.R.; Ha, A.J.W.; Ha, S.D. Advances and Future Prospects of Enzyme-Based Biofilm Prevention Approaches in the Food Industry. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1484–1502. [Google Scholar] [CrossRef] [PubMed]
- Thallinger, B.; Prasetyo, E.N.; Nyanhongo, G.S.; Guebitz, G.M. Antimicrobial enzymes: An emerging strategy to fight microbes and microbial biofilms. Biotechnol. J. 2013, 8, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Baidamshina, D.R.; Koroleva, V.A.; Trizna, E.Y.; Pankova, S.M.; Agafonova, M.N.; Chirkova, M.N.; Vasileva, O.S.; Akhmetov, N.; Shubina, V.V.; Porfiryev, A.G.; et al. Anti-biofilm and wound-healing activity of chitosan-immobilized Ficin. Int. J. Biol. Macromol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Niazi, S.A.; Al-Ali, W.M.; Patel, S.; Foschi, F.; Mannocci, F. Synergistic effect of 2% chlorhexidine combined with proteolytic enzymes on biofilm disruption and killing. Int. Endod. J. 2015, 48, 1157–1167. [Google Scholar] [CrossRef] [PubMed]
- Baidamshina, D.R.; Trizna, E.Y.; Holyavka, M.G.; Bogachev, M.I.; Artyukhov, V.G.; Akhatova, F.S.; Rozhina, E.V.; Fakhrullin, R.F.; Kayumov, A.R. Targeting microbial biofilms using Ficin, a nonspecific plant protease. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Lucas, J.; Castaneda, D.; Hormigo, D. New trends for a classical enzyme: Papain, a biotechnological success story in the food industry. Trends Food Sci. Technol. 2017, 68, 91–101. [Google Scholar] [CrossRef]
- Medeiros. Study of the effectiveness of papain in wound healing and specific approach to its application in patients with venous ulcers: A systematic review. Adv. Plast. Reconstr. Surg. 2018, 2, 183–187. [Google Scholar]
- Azevedo, F.F.; Santanna, L.P.; Bobbo, V.C.; Libert, E.A.; Araujo, E.P.; Saad, M.A.; Lima, M.H.M. Evaluating the Effect of 3% Papain Gel Application in Cutaneous Wound Healing in Mice. Wounds A Compend. Clin. Res. Pract. 2017, 29, 96–101. [Google Scholar]
- Porsani, M.Y.H.; Carvalho, L.A.R.; Pereira, C.S.; Paludetti, M.; Zangeronimo, M.G.; Pereira, L.J. The use of papain gel cream and sunflower oil in promoting healing in a wound in dogs: Three case reports. Arq. Bras. De Med. Vet. E Zootec. 2016, 68, 1201–1206. [Google Scholar] [CrossRef]
- Ribeiro, A.P.L.; de Oliveira, B.; Soares, M.F.; Barreto, B.M.F.; Futuro, D.O.; de Castilho, S.R. Effectiveness of 2% and 4% papain gels in the healing of venous ulcers. Rev. Da Esc. De Enferm. Da USP 2015, 49, 394–400. [Google Scholar] [CrossRef]
- Dai, T.H.; Tanaka, M.; Huang, Y.Y.; Hamblin, M.R. Chitosan preparations for wounds and burns: Antimicrobial and wound-healing effects. Expert Rev. Anti-Infect. Ther. 2011, 9, 857–879. [Google Scholar] [CrossRef]
- Younes, I.; Rinaudo, M. Chitin and Chitosan Preparation from Marine Sources. Structure, Properties and Applications. Mar. Drugs 2015, 13, 1133–1174. [Google Scholar] [CrossRef]
- Bracco, L.F.; Levin, G.J.; Urtasun, N.; del Canizo, A.A.N.; Wolman, F.J.; Miranda, M.V.; Cascone, O. Covalent immobilization of soybean seed hull urease on chitosan mini-spheres and the impact on their properties. Biocatal. Agric. Biotechnol. 2019, 18. [Google Scholar] [CrossRef]
- Klein, M.P.; Hackenhaar, C.R.; Lorenzoni, A.S.G.; Rodrigues, R.C.; Costa, T.M.H.; Ninow, J.L.; Hertz, P.F. Chitosan crosslinked with genipin as support matrix for application in food process: Support characterization and beta-D-galactosidase immobilization. Carbohydr. Polym. 2016, 137, 184–190. [Google Scholar] [CrossRef]
- Ma, H.F.; Meng, G.; Cui, B.K.; Si, J.; Dai, Y.C. Chitosan crosslinked with genipin as supporting matrix for biodegradation of synthetic dyes: Laccase immobilization and characterization. Chem. Eng. Res. Des. 2018, 132, 664–676. [Google Scholar] [CrossRef]
- Fini, A. The Role of Chitosan in Drug Delivery. Am. J. Drug Deliv. 2003, 1, 43–59. [Google Scholar] [CrossRef]
- Johnson, K.A. Role of induced fit in enzyme specificity: A molecular forward/reverse switch. J. Biol. Chem. 2008, 283, 26297–26301. [Google Scholar] [CrossRef] [PubMed]
- Michel, D. Conformational selection or induced fit? New insights from old principles. Biochimie 2016, 128, 48–54. [Google Scholar] [CrossRef]
- dos Santos, J.C.S.; Barbosa, O.; Ortiz, C.; Berenguer-Murcia, A.; Rodrigues, R.C.; Fernandez-Lafuente, R. Importance of the Support Properties for Immobilization or Purification of Enzymes. Chemcatchem 2015, 7, 2413–2432. [Google Scholar] [CrossRef]
- Mateo, C.; Grazu, V.; Pessela, B.C.C.; Montes, T.; Palomo, J.M.; Torres, R.; Lopez-Gallego, F.; Fernandez-Lafuente, R.; Guisan, J.M. Advances in the design of new epoxy supports for enzyme immobilization-stabilization. Biochem. Soc. Trans. 2007, 35, 1593–1601. [Google Scholar] [CrossRef] [PubMed]
- Akila, M.; Sushama, A.; Kumaresan, R. Study on in vitro cytotoxicity of papain against liver cancer cell line Hep G2. Int. J. Pharm. Pharm. Sci. 2014, 6, 160–161. [Google Scholar]
- Chandran, S.P.; Nachinmuthu, K.P.; Natarajan, S.B.; Inamdar, M.G.; Shahimi, M. Papain Loaded Solid Lipid Nanoparticles for Colorectal Cancer Therapy. Curr. Cancer Ther. Rev. 2018, 14, 75–87. [Google Scholar] [CrossRef]
- Chankhampan, C.; Manosroi, J.; Yamamoto, H.; Tahara, K.; Manosroi, W.; Kawashima, Y.; Manosroi, A. Chemical stability enhancement and cytotoxicity reduction of papain loaded in PLGA nanospheres. J. Exp. Nanosci. 2014, 9, 138–151. [Google Scholar] [CrossRef]
- Silva, Z.S.; Huang, Y.Y.; de Freitas, L.F.; Franca, C.M.; Botta, S.B.; Ana, P.A.; Mesquita-Ferrari, R.A.; Fernandes, K.P.S.; Deana, A.; Leal, C.R.L.; et al. Papain gel containing methylene blue for simultaneous caries removal and antimicrobial photoinactivation against Streptococcus mutans biofilms. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef]
- Kirker, K.R.; James, G.A. In vitro studies evaluating the effects of biofilms on wound-healing cells: A review. Apmis 2017, 125, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Atshan, S.S.; Shamsudin, M.N.; Sekawi, Z.; Lung, L.T.T.; Barantalab, F.; Liew, Y.K.; Alreshidi, M.A.; Abduljaleel, S.A.; Hamat, R.A. Comparative proteomic analysis of extracellular proteins expressed by various clonal types of Staphylococcus aureus and during planktonic growth and biofilm development. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef]
- Lewis, K. Riddle of biofilm resistance. Antimicrob. Agents Chemother. 2001, 45, 999–1007. [Google Scholar] [CrossRef]
- Cosgrove, S.E.; Kaye, K.S.; Eliopoulous, G.M.; Carmeli, Y. Health and economic outcomes of the emergence of third-generation cephalosporin resistance in Enterobacter species. Arch. Intern. Med. 2002, 162, 185–190. [Google Scholar] [CrossRef]
- Sanchez-Vizuete, P.; Orgaz, B.; Aymerich, S.; Le Coq, D.; Briandet, R. Pathogens protection against the action of disinfectants in multispecies biofilms. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef]
- Raafat, D.; Otto, M.; Reppschlager, K.; Iqbal, J.; Holtfreter, S. Fighting Staphylococcus aureus Biofilms with Monoclonal Antibodies. Trends Microbiol. 2019, 27, 303–322. [Google Scholar] [CrossRef]
- Adamus-Bialek, W.; Vollmerhausen, T.L.; Janik, K. Hydrogen peroxide stimulates uropathogenic Escherichia coli strains to cellulose production. Microb. Pathog. 2019, 126, 287–291. [Google Scholar] [CrossRef]
- Izano, E.A.; Amarante, M.A.; Kher, W.B.; Kaplan, J.B. Differential roles of poly-N-acetylglucosamine surface polysaccharide and extracellular DNA in Staphylococcus aureus and Staphylococcus epidermidis biofilms. Appl. Environ. Microbiol. 2008, 74, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Ray, P.; Das, A.; Sharma, M. Penetration of antibiotics through Staphylococcus aureus and Staphylococcus epidermidis biofilms. J. Antimicrob. Chemother. 2010, 65, 1955–1958. [Google Scholar] [CrossRef] [PubMed]
- Holyavka, M.G.; Kayumov, A.R.; Baydamshina, D.R.; Koroleva, V.A.; Trizna, E.Y.; Trushin, M.V.; Artyukhov, V.G. Efficient fructose production from plant extracts by immobilized inulinases from Kluyveromyces marxianus and Helianthus tuberosus. Int. J. Biol. Macromol. 2018, 115, 829–834. [Google Scholar] [CrossRef] [PubMed]
- Holyavka, M.; Pankova, S.; Koroleva, V.; Vyshkvorkina, Y.; Lukin, A.; Kondratyev, M.; Artyukhov, V. Influence of UV radiation on molecular structure and catalytic activity of free and immobilized bromelain, ficin and papain. J. Photochem. Photobiol. B Biol. 2019, 201. [Google Scholar] [CrossRef] [PubMed]
- Sabirova, A.R.; Rudakova, N.L.; Balaban, N.P.; Ilyinskaya, O.N.; Demidyuk, I.V.; Kostrov, S.V.; Rudenskaya, G.N.; Sharipova, M.R. A novel secreted metzincin metalloproteinase from Bacillus intermedius. FEBS Lett. 2010, 584, 4419–4425. [Google Scholar] [CrossRef]
- Kayumov, A.R.; Khakimullina, E.N.; Sharafutdinov, I.S.; Trizna, E.Y.; Latypova, L.Z.; Lien, H.T.; Margulis, A.B.; Bogachev, M.I.; Kurbangalieva, A.R. Inhibition of biofilm formation in Bacillus subtilis by new halogenated furanones. J. Antibiot. 2015, 68, 297–301. [Google Scholar] [CrossRef]
- Trizna, E.Y.; Khakimullina, E.N.; Latypova, L.Z.; Kurbangalieva, A.R.; Sharafutdinov, I.S.; Evtyugin, V.G.; Babynin, E.V.; Bogachev, M.I.; Kayumov, A.R. Thio Derivatives of 2(5H)-Furanone As Inhibitors against Bacillus subtilis Biofilms. Acta Nat. 2015, 7, 102–107. [Google Scholar] [CrossRef]
- Leclercq, R.; Canton, R.; Brown, D.F.J.; Giske, C.G.; Heisig, P.; MacGowan, A.P.; Mouton, J.W.; Nordmann, P.; Rodloff, A.C.; Rossolini, G.M.; et al. EUCAST expert rules in antimicrobial susceptibility testing. Clin. Microbiol. Infect. 2013, 19, 141–160. [Google Scholar] [CrossRef]
- O’Toole, G.A.; Kolter, R. Initiation of biofilm formation in Pseudomonas fluorescens WCS365 proceeds via multiple, convergent signalling pathways: A genetic analysis. Mol. Microbiol. 1998, 28, 449–461. [Google Scholar] [CrossRef]
- Herigstad, B.; Hamilton, M.; Heersink, J. How to optimize the drop plate method for enumerating bacteria. J. Microbiol. Methods 2001, 44, 121–129. [Google Scholar] [CrossRef]
- Sharafutdinov, I.; Pavlova, A.; Khabibrakhmanova, A.; Kurbangalieva, A.; Kayumov, A. The antimicrobial effect of the 5-((-)bornyloxy)-2(5H)-furanone derivative on grampositive bacteria. FEBS J. 2017, 284, 143. [Google Scholar]
- Bogachev, M.I.; Volkov, V.Y.; Markelov, O.A.; Trizna, E.Y.; Baydamshina, D.R.; Melnikov, V.; Murtazina, R.R.; Zelenikhin, P.V.; Sharafutdinov, I.S.; Kayumov, A.R. Fast and simple tool for the quantification of biofilm-embedded cells sub-populations from fluorescent microscopic images. PLoS ONE 2018, 13. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Buffer System | pH | Total Protein, mg g Chitosan−1 | Protein, mg mL−1 | Activity, U mL−1 | Specific Activity, U mg Protein−1 | |||
|---|---|---|---|---|---|---|---|---|
| 0.2 M Sodium acetate | 4.0 | 2.6 ± 0.50 | (13.2%) | 0.13 ± 0.02 | 42 ± 4.1 | (44.6%) | 320 ± 30.8 | (33.8%) |
| 4.5 | 3.3 ± 0.38 | (16.4%) | 0.16 ± 0.02 | 79 ± 5.4 | (83.7%) | 484 ± 33.0 | (51.0%) | |
| 5.0 | 3.9 ± 0.37 | (19.4%) | 0.19 ± 0.02 | 82 ± 6.0 | (87.0%) | 425 ± 30.8 | (44.9%) | |
| 5.8 | 5.4 ± 0.84 | (26.8%) | 0.27 ± 0.04 | 90 ± 10.1 | (95.4%) | 337 ±37.7 | (35.6%) | |
| 0.1 M di-potassium hydrogen phosphate | 5.8 | 5.4 ± 0.51 | (26.9%) | 0.27 ± 0.03 | 22 ± 1.8 | (23.5%) | 83 ± 6.8 | (8.7%) |
| 6.0 | 5.1 ± 0.40 | (25.8%) | 0.26 ± 0.02 | 18 ± 0.4 | (20.0%) | 73 ± 1.80 | (7.8%) | |
| 6.5 | 5.2 ± 0.22 | (26.0%) | 0.26 ± 0.01 | 23 ± 0.4 | (24.6%) | 89 ± 1.5 | (9.4%) | |
| 7.0 | 4.6 ± 0.45 | (22.8%) | 0.23 ± 0.02 | 18 ± 0.8 | (19.6%) | 81 ± 3.7 | (8.6%) | |
| 7.5 | 4.9 ± 0.28 | (24.5%) | 0.24 ± 0.01 | 17 ± 0.4 | (18.6%) | 72 ± 1.8 | (7.6%) | |
| 8.0 | 4.6 ± 0.28 | (22.8%) | 0.23 ± 0.01 | 19 ± 1.0 | (20.5%) | 85± 4.3 | (9.0%) | |
| 0.05 M Tri-sodium borate, 0.1 M KCl | 8.0 | 7.6 ± 0.76 | (38.0%) | 0.38 ± 0.04 | 38 ± 4.6 | (40.6%) | 101 ± 8.8 | (10.7%) |
| 8.5 | 7.2 ± 1.04 | (36.1%) | 0.36 ± 0.05 | 39 ± 2.8 | (41.4%) | 109 ± 6.7 | (11.5%) | |
| 9.0 | 7.3 ± 0.79 | (36.7%) | 0.37 ± 0.04 | 39 ± 1.0 | (41.7%) | 108 ± 2.6 | (11.4%) | |
| 9.5 | 7.4 ± 0.89 | (36.9%) | 0.37 ± 0.04 | 39 ± 0.4 | (41.2%) | 106 ± 1.1 | (11.1%) | |
| 10.0 | 6.6 ± 0.41 | (33.2%) | 0.33 ± 0.02 | 41 ± 6.6 | (43.0%) | 123 ± 19.5 | (12.9%) | |
| 0.05 M Tris-glycine | 8.5 | 5.9 ± 0.24 | (29.5%) | 0.29 ± 0.01 | 82 ± 5.1 | (86.7%) | 275 ± 17.1 | (29.0%) |
| 9.0 | 5.5 ± 0.17 | (27.7%) | 0.28 ± 0.01 | 88 ± 6.5 | (92.7%) | 317 ± 23.6 | (33.4%) | |
| 0.05 M Glycine | 8.6 | 5.8 ± 0.31 | (28.8%) | 0.29 ± 0.01 | 91 ± 2.0 | (96.3%) | 317 ± 6.2 | (33.4%) |
| 9.0 | 5.9 ± 0.22 | (29.6%) | 0.30 ± 0.01 | 94 ± 2.3 | (99.3%) | 318 ± 24.9 | (33.5%) | |
| 9.5 | 5.9 ± 0.20 | (29.5%) | 0.29 ± 0.01 | 89 ± 2.7 | (94.4%) | 303 ± 7.9 | (32.0%) | |
| 10.0 | 5.8 ± 0.46 | (29.3%) | 0.29 ± 0.02 | 88 ± 3.8 | (92.6%) | 300 ± 12.9 | (31.6%) | |
| 10.5 | 5.7 ± 0.41 | (28.5%) | 0.28 ± 0.02 | 86 ± 1.9 | (90.3%) | 300 ± 5.8 | (31.7%) | |
| Buffer System | pH | Total Protein, mg g Chitosan−1 | Protein, mg mL−1 | Activity, U mL−1 | Specific Activity, U mg Protein−1 | |||
|---|---|---|---|---|---|---|---|---|
| 0.2 M Sodium acetate | 4.0 | 3.4 ± 0.63 | (16.8%) | 0.17 ± 0.03 | 46 ± 8.8 | (48.0%) | 271 ± 52.3 | (28.6%) |
| 4.5 | 4.7 ± 0.33 | (21.3%) | 0.21 ± 0.02 | 50 ± 8.4 | (52.8%) | 235 ± 39.5 | (24.8%) | |
| 5.0 | 6.9 ± 0.69 | (34.4%) | 0.34 ± 0.04 | 88 ± 4.6 | (93.4%) | 257 ± 13.3 | (27.1%) | |
| 5.8 | 7.1 ± 0.89 | (35.7%) | 0.36 ±0.05 | 78 ± 8.4 | (82.0%) | 218 ± 23.7 | (23.0%) | |
| 0.1 M di-potassium hydrogen phosphate | 5.8 | 5.7 ± 0.64 | (28.3%) | 0.28 ±0.03 | 26 ± 2.4 | (27.7%) | 92 ± 5.3 | (9.8%) |
| 6.0 | 5.3 ± 0.54 | (26.5%) | 0.26 ± 0.03 | 27 ± 2.0 | (28.5%) | 102 ± 7.6 | (10.7%) | |
| 6.5 | 5.0 ± 0.33 | (25.4%) | 0.25 ± 0.01 | 27 ± 2.1 | (28.6%) | 107 ± 6.7 | (11.3%) | |
| 7.0 | 4.7 ± 0.74 | (23.4%) | 0.23 ± 0.02 | 25 ± 1.8 | (26.9%) | 109 ± 5.6 | (11.6%) | |
| 7.5 | 4.3 ± 0.47 | (21.4%) | 0.21 ± 0.02 | 25 ± 1.4 | (26.4%) | 117 ± 8.2 | (12.3%) | |
| 8.0 | 4.2 ± 0.35 | (21.1%) | 0.21 ± 0.01 | 24 ± 1.3 | (25.7%) | 114 ± 10.0 | (12.0%) | |
| 0.05 M Tri-sodium borate, 0.1 M KCl | 8.0 | 8.2 ± 0.68 | (41.3%) | 0.41 ± 0.03 | 94 ± 5.1 | (99.6%) | 229 ± 8.3 | (24.1%) |
| 8.5 | 7.9 ± 1.19 | (39.7%) | 0.40 ± 0.06 | 93 ± 6.4 | (98.7%) | 235 ± 16.1 | (24.8%) | |
| 9.0 | 7.5 ± 0.73 | (37.6%) | 0.38 ± 0.04 | 93 ± 3.8 | (98.8%) | 249 ± 10.2 | (26.3%) | |
| 9.5 | 6.2 ± 1.91 | (30.1%) | 0.31 ± 0.09 | 94 ± 3.2 | (99.6%) | 305 ± 10.4 | (21.2%) | |
| 10.0 | 5.9 ± 1.08 | (29.4%) | 0.29 ± 0.05 | 94 ± 5.3 | (99.1%) | 319 ± 10.1 | (33.7%) | |
| 0.05 M Tris-glycine | 8.5 | 8.4 ± 1.5 | (42.2%) | 0.42 ± 0.08 | 92 ± 6.9 | (97.4%) | 219 ± 16.3 | (23.1%) |
| 9.0 | 7.2 ± 0.8 | (36.2%) | 0.36 ± 0.04 | 91 ± 5.2 | (96.6%) | 253 ± 14.3 | (26.7%) | |
| 0.05 M Glycine | 8.6 | 7.0 ± 0.6 | (35.1%) | 0.35 ± 0.03 | 86 ± 5.2 | (91.1%) | 246 ± 10.4 | (26.0%) |
| 9.0 | 7.2 ± 1.04 | (36.0%) | 0.36 ± 0.05 | 94 ± 7.9 | (99.8%) | 263 ± 15.3 | (27.7%) | |
| 9.5 | 8.2 ± 0.47 | (40.9%) | 0.41 ± 0.02 | 94 ± 4.2 | (99.5%) | 231 ± 9.6 | (24.3%) | |
| 10.0 | 8.1 ± 0.9 | (40.7%) | 0.41 ± 0.05 | 94 ± 5.7 | (99.0%) | 230 ± 11.7 | (24.3%) | |
| 10.5 | 7.9 ± 1.2 | (39.7%) | 0.39 ± 0.06 | 93 ± 9.7 | (98.3%) | 235 ± 24.4 | (24.8%) | |
| Sample | Km, µM | Vmax, µM mg−1 min−1 | kcat, min−1 | Vmax/Km | kcat/Km |
|---|---|---|---|---|---|
| Soluble papain | 22.3 ± 3.5 | 1253 ± 64 | 57.8 ± 2.9 | 56.2 | 2.59 |
| Medium molecular weight chitosan immobilized papain | 24.7 ± 5.4 | 432 ± 32 | 6.8 ± 0.5 | 17.3 | 0.28 |
| High molecular weight chitosan immobilized papain | 23.1 ± 4.3 | 349 ± 21 | 4.5 ± 0.3 | 15.2 | 0.19 |
| S. aureus | S. epidermidis | |||
|---|---|---|---|---|
| MIC, µg mL−1 | MBC, µg mL−1 | MIC, µg mL−1 | MBC, µg mL−1 | |
| Gentamycin | 4 | 16 | 1 | 4 |
| Ciprofloxacin | 2 | 32 | 0.5 | 4 |
| Benzalkonium chloride | 1 | 16 | 1 | 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baidamshina, D.R.; Koroleva, V.A.; Olshannikova, S.S.; Trizna, E.Y.; Bogachev, M.I.; Artyukhov, V.G.; Holyavka, M.G.; Kayumov, A.R. Biochemical Properties and Anti-Biofilm Activity of Chitosan-Immobilized Papain. Mar. Drugs 2021, 19, 197. https://doi.org/10.3390/md19040197
Baidamshina DR, Koroleva VA, Olshannikova SS, Trizna EY, Bogachev MI, Artyukhov VG, Holyavka MG, Kayumov AR. Biochemical Properties and Anti-Biofilm Activity of Chitosan-Immobilized Papain. Marine Drugs. 2021; 19(4):197. https://doi.org/10.3390/md19040197
Chicago/Turabian StyleBaidamshina, Diana R., Victoria A. Koroleva, Svetlana S. Olshannikova, Elena Yu. Trizna, Mikhail I. Bogachev, Valeriy G. Artyukhov, Marina G. Holyavka, and Airat R. Kayumov. 2021. "Biochemical Properties and Anti-Biofilm Activity of Chitosan-Immobilized Papain" Marine Drugs 19, no. 4: 197. https://doi.org/10.3390/md19040197
APA StyleBaidamshina, D. R., Koroleva, V. A., Olshannikova, S. S., Trizna, E. Y., Bogachev, M. I., Artyukhov, V. G., Holyavka, M. G., & Kayumov, A. R. (2021). Biochemical Properties and Anti-Biofilm Activity of Chitosan-Immobilized Papain. Marine Drugs, 19(4), 197. https://doi.org/10.3390/md19040197