
A Metataxonomic Approach Reveals Diversified Bacterial Communities in Antarctic Sponges





 ,
,  ,
,  ,
,  ,
, 


Abstract
1. Introduction
2. Results
2.1. Species Identification
2.1.1. Morphological Analysis
2.1.2. Molecular Analysis
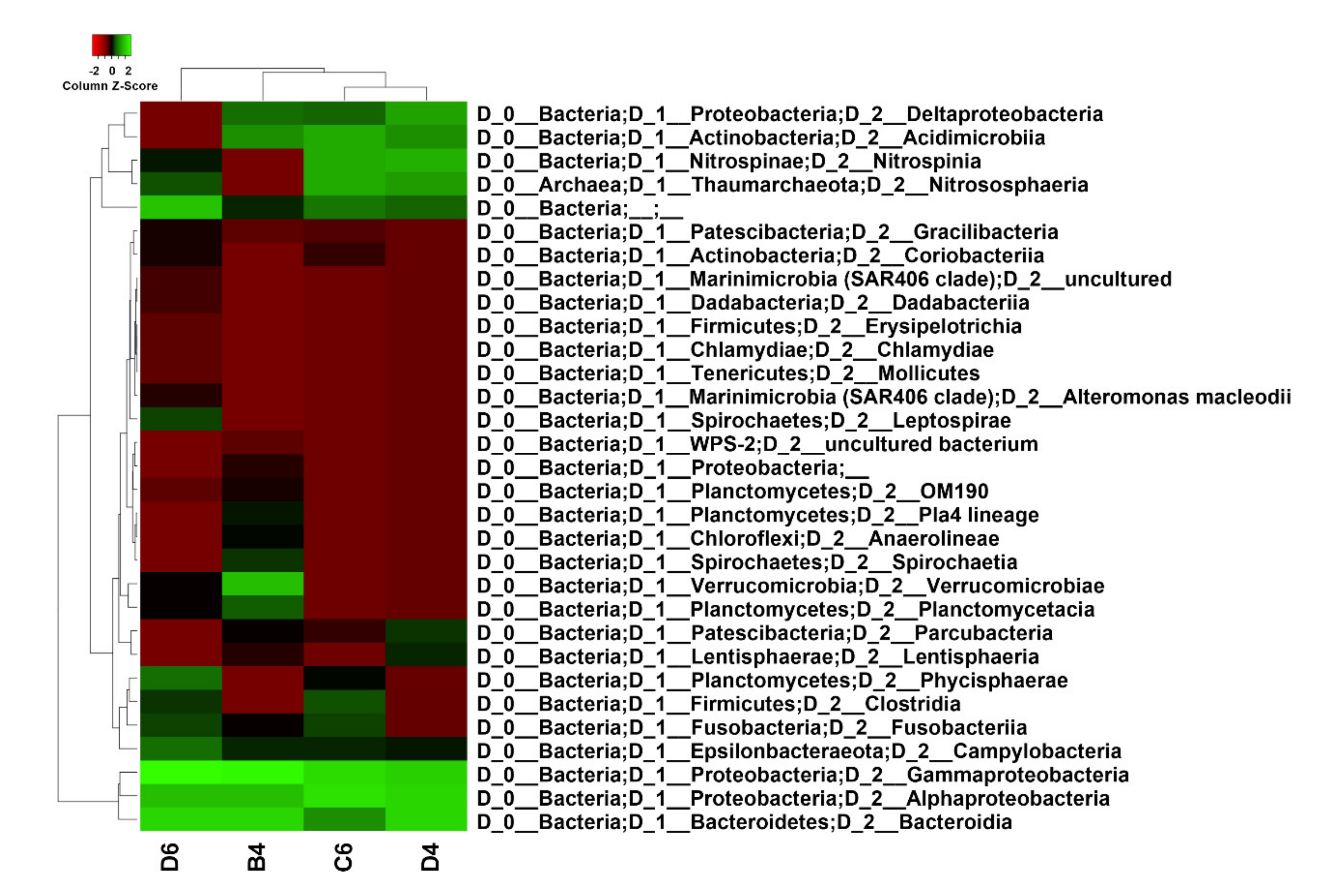
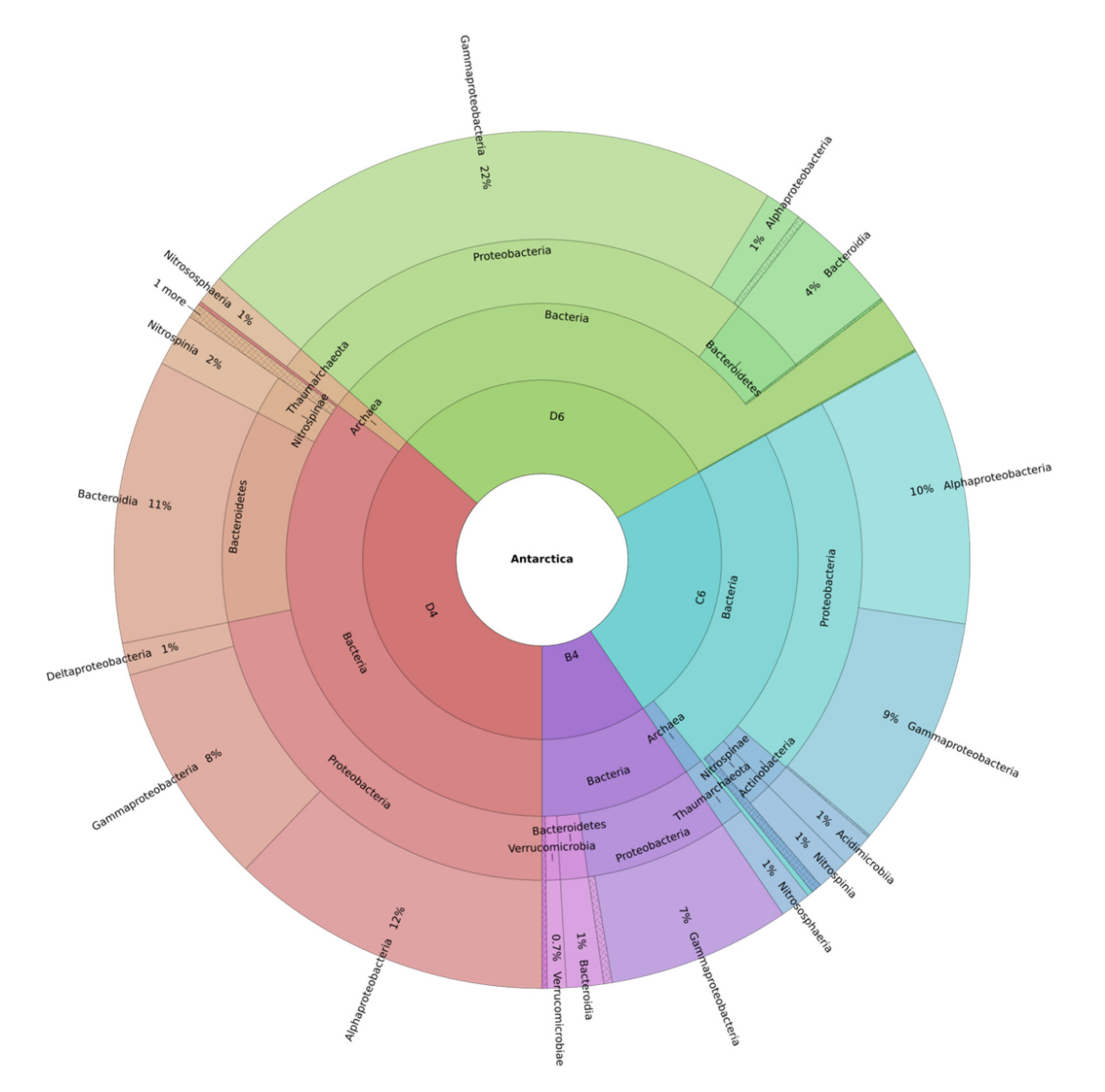
2.2. Metataxonomic Data Analysis
3. Discussion
4. Materials and Methods
4.1. Sponge Collection
4.2. Morphological Analysis of Spicules
4.3. DNA Extraction and PCR Amplification
4.4. Metagenomic DNA Extraction and Illumina MiSeq Sequencing
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Peck, L.S. Antarctic marine biodiversity: Adaptations, environments and responses to change. In Oceanography and Marine Biology: An Annual Review; Hawkins, S.J., Evans, A.J., Dale, A.C., Firth, L.B., Smith, I.P., Eds.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2018; Volume 56, pp. 2–133. ISBN 9781138318625. [Google Scholar]
- Arnaud, P. Contribution à la bionomie marine benthique des régions antarctiques et subantarctiques. In Téthys; Station Marine d’Endoume, Centre d’Océanographie de Marseille: Marseille, France, 1974; Volume 6, pp. 465–656. [Google Scholar]
- Konecki, J.T.; Targett, T.E. Eggs and larvae of Nototheniops larseni from the spongocoel of a hexactinellid sponge near Hugo island, Antarctic Peninsula. Polar Biol. 1989, 10, 197–198. [Google Scholar] [CrossRef]
- Pearse, J.S.; Mcclintock, J.B.; Bosch, I. Reproduction of Antarctic benthic marine invertebrates: Tempos, modes, and timing. Integr. Comp. Biol. 1991, 31, 65–80. [Google Scholar] [CrossRef]
- Dayton, P.K. Polar benthos. In Polar oceanography-Part B-Chemistry, Biology and Geology; Smith, W.O., Ed.; Academic Press: London, UK, 1990; pp. 631–685. ISBN 9780080925950. [Google Scholar]
- Dayton, P.K.; Mordida, B.J.; Bacon, F. Polar marine communities. Am. Zool. 1994, 34, 90–99. [Google Scholar] [CrossRef]
- Thompson, M.R.A.; Crame, J.A.; Thompson, J.W. Geological Evolution of Antarctica; Cambridge University Press: Cambridge, UK, 1991; ISBN 9780521372664. [Google Scholar]
- Tingey, R. The Geology of Antarctica; Oxford University Press: Oxford, UK, 1991; ISBN 9780198544678. [Google Scholar]
- Clarke, A. Is there a latitudinal diversity cline in the sea? Trends Ecol. Evol. 1992, 7, 286–287. [Google Scholar] [CrossRef]
- Crame, J.; Clarke, A. The historical component of marine taxonomic diversity gradients. In Marine Biodiversity: Patterns and Processes; Ormond, R.F.G., Gage, J.D., Angel, M.V., Eds.; Cambridge University Press: Cambridge, UK, 1997; pp. 258–273. [Google Scholar]
- Gray, J.S. Antarctic marine benthic biodiversity in a world-wide latitudinal context. Polar Biol. 2001, 24, 633–641. [Google Scholar] [CrossRef]
- McClintock, J.B.; Amsler, C.D.; Baker, B.J.; Van Soest, R.W.M. Ecology of antarctic marine sponges: An overview. Integr. Comp. Biol. 2005, 45, 359–368. [Google Scholar] [CrossRef]
- Janussen, D.; Downey, R. V Porifera. In Biogeographic Atlas of the Southern Ocean; De Broyer, C., Koubbi, P., Eds.; SCAR: Cambridge, UK, 2014; pp. 1–10. [Google Scholar]
- Mangano, S.; Michaud, L.; Caruso, C.; Brilli, M.; Bruni, V.; Fani, R.; Lo Giudice, A. Antagonistic interactions between psychrotrophic cultivable bacteria isolated from Antarctic sponges: A preliminary analysis. Res. Microbiol. 2009, 160, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Hentschel, U.; Piel, J.; Degnan, S.M.; Taylor, M.W. Genomic insights into the marine sponge microbiome. Nat. Rev. Microbiol. 2012, 10, 641–654. [Google Scholar] [CrossRef]
- Bayer, K.; Kamke, J.; Hentschel, U. Quantification of bacterial and archaeal symbionts in high and low microbial abundance sponges using real-time PCR. FEMS Microbiol. Ecol. 2014, 89, 679–690. [Google Scholar] [CrossRef]
- Lee, Y.K.; Lee, J.H.; Lee, H.K. Microbial symbiosis in marine sponges. J. Microbiol. 2001, 39, 254–264. [Google Scholar]
- Thoms, C.; Horn, M.; Wagner, M.; Hentschel, U.; Proksch, P. Monitoring microbial diversity and natural product profiles of the sponge Aplysina cavernicola following transplantation. Mar. Biol. 2003, 142, 685–692. [Google Scholar] [CrossRef]
- Hentschel, U.; Usher, K.M.; Taylor, M.W. Marine sponges as microbial fermenters. FEMS Microbiol. Ecol. 2006, 55, 167–177. [Google Scholar] [CrossRef]
- Calcinai, B.; Pansini, M. Four new demosponge species from Terra Nova Bay (Ross Sea, Antarctica). Zoosystema 2000, 22, 369–381. [Google Scholar]
- Ríos, P.; Cristobo, F.J.; Urgorri, V. Poecilosclerida (Porifera, Demospongiae) collected by the Spanish Antarctic expedition BENTART-Cah. Biol. Mar. 2004, 45, 97–119. [Google Scholar]
- Dayton, P.K.; Robilliard, G.A.; Paine, R.T.; Dayton, L.B. Biological accommodation in the benthic community at McMurdo Sound, Antarctica. Ecol. Monogr. 1974, 44, 105–128. [Google Scholar] [CrossRef]
- Cárdenas, C.A.; Font, A.; Steinert, G.; Rondon, R.; González-Aravena, M. Temporal stability of bacterial communities in Antarctic sponges. Front. Microbiol. 2019, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lo Giudice, A.; Rizzo, C. Bacteria associated with marine benthic invertebrates from polar environments: Unexplored frontiers for biodiscovery? Diversity 2018, 10, 80. [Google Scholar] [CrossRef]
- Lo Giudice, A.; Azzaro, M.; Schiaparelli, S. Microbial symbionts of Antarctic marine benthic invertebrates. In The Ecological Role of Micro-Organisms in the Antarctic Environment; Castro-Sowinski, S., Ed.; Springer International Publishing: New York, NY, USA, 2019; pp. 277–296. [Google Scholar]
- Steinert, G.; Wemheuer, B.; Janussen, D.; Erpenbeck, D.; Daniel, R.; Simon, M.; Brinkhoff, T.; Schupp, P.J. Prokaryotic diversity and community patterns in antarctic continental shelf sponges. Front. Mar. Sci. 2019, 6, 1–15. [Google Scholar] [CrossRef]
- Savoca, S.; Lo Giudice, A.; Papale, M.; Mangano, S.; Caruso, C.; Spanò, N.; Michaud, L.; Rizzo, C. Antarctic sponges from the Terra Nova Bay (Ross Sea) host a diversified bacterial community. Sci. Rep. 2019, 9, 16135. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Pino, M.; Cristi, A.; Gillooly, J.F.; Trefault, N. Characterizing the microbiomes of Antarctic sponges: A functional metagenomic approach. Sci. Rep. 2020, 10, 645. [Google Scholar] [CrossRef]
- Papale, M.; Rizzo, C.; Fani, R.; Bertolino, M.; Costa, G.; Paytuví-Gallart, A.; Schiaparelli, S.; Michaud, L.; Azzaro, M.; Lo Giudice, A. Exploring the diversity and metabolic profiles of bacterial communities associated with Antarctic sponges (Terra Nova Bay, Ross Sea). Front. Ecol. Evol. 2020, 8, 1–16. [Google Scholar] [CrossRef]
- Sacristán-Soriano, O.; Pérez Criado, N.; Avila, C. Host species determines symbiotic community composition in Antarctic sponges (Porifera: Demospongiae). Front. Mar. Sci. 2020, 7, 1–36. [Google Scholar] [CrossRef]
- Rodríguez-Marconi, S.; De La Iglesia, R.; Díez, B.; Fonseca, C.A.; Hajdu, E.; Trefault, N.; Webster, N. Characterization of bacterial, archaeal and eukaryote symbionts from antarctic sponges reveals a high diversity at a three-domain level and a particular signature for this ecosystem. PLoS ONE 2015, 10, 1–19. [Google Scholar] [CrossRef]
- Webster, N.S.; Negri, A.P.; Munro, M.M.H.G.; Battershill, C.N. Diverse microbial communities inhabit Antarctic sponges. Environ. Microbiol. 2004, 6, 288–300. [Google Scholar] [CrossRef] [PubMed]
- Mangano, S.; Michaud, L.; Caruso, C.; Lo Giudice, A. Metal and antibiotic resistance in psychrotrophic bacteria associated with the Antarctic sponge Hemigellius pilosus (Kirkpatrick, 1907). Polar Biol. 2014, 37, 227–235. [Google Scholar] [CrossRef]
- Mangano, S.; Caruso, C.; Michaud, L.; Lo Giudice, A. First evidence of quorum sensing activity in bacteria associated with Antarctic sponges. Polar Biol. 2018, 41, 1435–1445. [Google Scholar] [CrossRef]
- Caruso, C.; Rizzo, C.; Mangano, S.; Poli, A.; di Donato, P.; Finore, I.; Nicolaus, B.; di Marco, G.; Michaud, L.; Lo Giudice, A. Production and biotechnological potential of extracellular polymeric substances from sponge-associated Antarctic bacteria. Appl. Environ. Microbiol. 2018, 84, e01624-17. [Google Scholar] [CrossRef]
- Brinkmann, C.M.; Marker, A.; Kurtböke, D.I. An overview on marine sponge-symbiotic bacteria as unexhausted sources for natural product discovery. Diversity 2017, 9, 40. [Google Scholar] [CrossRef]
- Taylor, M.W.; Radax, R.; Stegar, D.; Wagner, M. Sponge-associated microorganisms: Evolution, evology, and biotechnological potential. Microbiol. Mol. Biol. Rev. 2007, 71, 295–347. [Google Scholar] [CrossRef] [PubMed]
- Webster, N.S.; Taylor, M.W. Marine sponges and their microbial symbionts: Love and other relationships. Environ. Microbiol. 2012, 14, 335–346. [Google Scholar] [CrossRef]
- Thacker, R.W.; Freeman, C.J. Sponge-microbe symbioses. Recent advances and new directions. In Advances in Marine Biology; Academic Press: Cambridge, MA, USA, 2012; Volume 62, pp. 57–111. [Google Scholar]
- Björk, J.R.; Díez-Vives, C.; Coma, R.; Ribes, M.; Montoya, J.M. Specificity and temporal dynamics of complex bacteria–sponge symbiotic interactions. Ecology 2013, 94, 2781–2791. [Google Scholar] [CrossRef]
- Hughes, K.A.; Pertierra, L.R.; Walton, D.W.H. Area protection in Antarctica: How can conservation and scientific research goals be managed compatibly? Environ. Sci. Policy 2013, 31, 120–132. [Google Scholar] [CrossRef]
- Kennedy, J.; Codling, C.E.; Jones, B.V.; Dobson, A.D.W.; Marchesi, J.R. Diversity of microbes associated with the marine sponge, Haliclona simulans, isolated from Irish waters and identification of polyketide synthase genes from the sponge metagenome. Environ. Microbiol. 2008, 10, 1888–1902. [Google Scholar] [CrossRef] [PubMed]
- Dupont, S.; Carré-Mlouka, A.; Descarrega, F.; Ereskovsky, A.; Longeon, A.; Mouray, E.; Florent, I.; Bourguet-Kondracki, M.L. Diversity and biological activities of the bacterial community associated with the marine sponge Phorbas tenacior (Porifera, Demospongiae). Lett. Appl. Microbiol. 2014, 58, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Karimi, E.; Ramos, M.; Gonçalves, J.M.S.; Xavier, J.R.; Reis, M.P.; Costa, R. Comparative metagenomics reveals the distinctive adaptive features of the Spongia officinalis endosymbiotic consortium. Front. Microbiol. 2017, 8, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Slaby, B.M.; Hackl, T.; Horn, H.; Bayer, K.; Hentschel, U. Metagenomic binning of a marine sponge microbiome reveals unity in defense but metabolic specialization. ISME J. 2017, 11, 2465–2478. [Google Scholar] [CrossRef]
- Najafi, A.; Moradinasab, M.; Nabipour, I. First record of microbiomes of sponges collected from the Persian Gulf, using tag pyrosequencing. Front. Microbiol. 2018, 9, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Storey, M.A.; Andreassend, S.K.; Bracegirdle, J.; Brown, A.; Keyzers, R.A.; Ackerley, D.F.; Northcote, P.T.; Owen, J.G. Metagenomic exploration of the marine sponge Mycale hentscheli uncovers multiple polyketide-producing bacterial symbionts. MBio 2020, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Zhang, W.; Ding, W.; Wang, M.; Fan, S.; Yang, B.; McMinn, A.; Wang, M.; Xie, B.B.; Qin, Q.L.; et al. Structure and function of the Arctic and Antarctic marine microbiota as revealed by metagenomics. Microbiome 2020, 8, 47. [Google Scholar] [CrossRef] [PubMed]
- Stevens, H.; Stübner, M.; Simon, M.; Brinkhoff, T. Phylogeny of Proteobacteria and Bacteroidetes from oxic habitats of a tidal flat ecosystem. FEMS Microbiol. Ecol. 2005, 54, 351–365. [Google Scholar] [CrossRef] [PubMed]
- Bibi, F.; Alvi, S.A.; Al-Sofyani, A.; Naseer, M.I.; Yasir, M.; Azhar, E.I. Pyrosequencing reveals sponge specific bacterial communities in marine sponges of Red Sea, Saudi Arabia. Saudi J. Biol. Sci. 2020, 27, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Joye, S.B.; Kleindienst, S.; Gilbert, J.A.; Handley, K.M.; Weisenhorn, P.; Overholt, W.A.; Kostka, J.E. Responses of microbial communities to hydrocarbon exposures. Oceanography 2016, 29, 136–149. [Google Scholar] [CrossRef]
- McGenity, T.J. Taxonomy, Genomics and Ecophysiology of Hydrocarbon-Degrading Microbes; Springer International Publishing: New York, NY, USA, 2019; ISBN 9783319600536. [Google Scholar]
- Hamdan, L.J.; Coffin, R.B.; Sikaroodi, M.; Greinert, J.; Treude, T.; Gillevet, P.M. Ocean currents shape the microbiome of Arctic marine sediments. ISME J. 2013, 7, 685–696. [Google Scholar] [CrossRef]
- Williams, T.J.; Cavicchioli, R. Marine metaproteomics: Deciphering the microbial metabolic food web. Trends Microbiol. 2014, 22, 248–260. [Google Scholar] [CrossRef]
- Shivaji, S.; Reddy, G.S.N.; Chattopadhyay, M.K. Bacterial biodiversity, cold adaptation and biotechnological importance of bacteria occurring in Antarctica. Proc. Ind. Natl. Sci. Acad. USA 2017, 83, 327–352. [Google Scholar] [CrossRef]
- Picazo, A.; Rochera, C.; Villaescusa, J.A.; Miralles-Lorenzo, J.; Velázquez, D.; Quesada, A.; Camacho, A. Bacterioplankton community composition along environmental gradients in lakes from Byers peninsula (Maritime Antarctica) as determined by next-generation sequencing. Front. Microbiol. 2019, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Buijs, Y.; Bech, P.K.; Vazquez-Albacete, D.; Bentzon-Tilia, M.; Sonnenschein, E.C.; Gram, L.; Zhang, S. Da Marine Proteobacteria as a source of natural products: Advances in molecular tools and strategies. Nat. Prod. Rep. 2019, 36, 1333–1350. [Google Scholar] [CrossRef]
- Reen, F.; Gutiérrez-Barranquero, J.; Dobson, A.; Adams, C.; O’Gara, F. Emerging Concepts Promising New Horizons for Marine Biodiscovery and Synthetic Biology. Mar. Drugs 2015, 13, 2924–2954. [Google Scholar] [CrossRef] [PubMed]
- Bode, H.B.; Bethe, B.; Höfs, R.; Zeeck, A. Big effects from small changes: Possible ways to explore nature’s chemical diversity. ChemBioChem 2002, 3, 619–627. [Google Scholar] [CrossRef]
- Santos, O.C.S.; Pontes, P.V.M.L.; Santos, J.F.M.; Muricy, G.; Giambiagi-deMarval, M.; Laport, M.S. Isolation, characterization and phylogeny of sponge-associated bacteria with antimicrobial activities from Brazil. Res. Microbiol. 2010, 161, 604–612. [Google Scholar] [CrossRef]
- Lee, L.H.; Cheah, Y.K.; Nurul Syakima, A.M.; Shiran, M.S.; Tang, Y.L.; Lin, H.P.; Hong, K. Analysis of Antarctic protobacteria by PCR fingerprinting and screening for antimicrobial secondary metabolites. Genet. Mol. Res. 2012, 11, 1627–1641. [Google Scholar] [CrossRef] [PubMed]
- Desriac, F.; Jégou, C.; Balnois, E.; Brillet, B.; Le Chevalier, P.; Fleury, Y. Antimicrobial peptides from marine proteobacteria. Mar. Drugs 2013, 11, 3632–3660. [Google Scholar] [CrossRef] [PubMed]
- Bibi, F.; Yasir, M.; Al-Sofyani, A.; Naseer, M.I.; Azhar, E.I. Antimicrobial activity of bacteria from marine sponge Suberea mollis and bioactive metabolites of Vibrio sp. EASaudi J. Biol. Sci. 2020, 27, 1139–1147. [Google Scholar] [CrossRef] [PubMed]
- Haber, M.; Ilan, M. Diversity and antibacterial activity of bacteria cultured from Mediterranean Axinella spp. sponges. J. Appl. Microbiol. 2014, 116, 519–532. [Google Scholar] [CrossRef]
- Rua, C.P.J.; Trindade-Silva, A.E.; Appolinario, L.R.; Venas, T.M.; Garcia, G.D.; Carvalho, L.S.; Lima, A.; Kruger, R.; Pereira, R.C.; Berlinck, R.G.S.; et al. Diversity and antimicrobial potential of culturable heterotrophic bacteria associated with the endemic marine sponge Arenosclera Brasiliensis. PeerJ 2014, 2014, 1–14. [Google Scholar] [CrossRef]
- Lin, J.; Meng, Y.; Shi, Y.; Lin, X. Complete genome sequences of Colwellia sp. Arc7-635, a denitrifying bacterium isolated from Arctic seawater. Curr. Microbiol. 2019, 76, 1061–1065. [Google Scholar] [CrossRef]
- Casillo, A.; Parrilli, E.; Sannino, F.; Mitchell, D.E.; Gibson, M.I.; Marino, G.; Lanzetta, R.; Parrilli, M.; Cosconati, S.; Novellino, E.; et al. Structure-activity relationship of the exopolysaccharide from a psychrophilic bacterium: A strategy for cryoprotection. Carbohydr. Polym. 2017, 156, 364–371. [Google Scholar] [CrossRef]
- Wang, Z.J.; Zhou, H.; Zhong, G.; Huo, L.; Tang, Y.J.; Zhang, Y.; Bian, X. Genome mining and biosynthesis of primary amine-acylated desferrioxamines in a marine gliding bacterium. Org. Lett. 2020, 22, 939–943. [Google Scholar] [CrossRef]
- Choi, T.H.; Lee, H.K.; Lee, K.; Cho, J.C. Ulvibacter antarcticus sp. nov., isolated from Antarctic coastal seawater. Int. J. Syst. Evol. Microbiol. 2007, 57, 2922–2925. [Google Scholar] [CrossRef] [PubMed]
- McBride, M.J. The Family Flavobacteriaceae. In The Prokaryotes: Other Major Lineages of Bacteria and The Archaea; Rosenberg, E., De Long, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer-Verlag: Berlin/Heidelberg, Germany, 2014; pp. 1–1028. [Google Scholar]
- Enisoglu-Atalay, V.; Atasever-Arslan, B.; Yaman, B.; Cebecioglu, R.; Kul, A.; Ozilhan, S.; Ozen, F.; Cata, T. Chemical and molecular characterization of metabolites from Flavobacterium sp. PLoS ONE 2018, 13, e0205817. [Google Scholar] [CrossRef]
- Scheuermayer, M.; Gulder, T.A.M.; Bringmann, G.; Hentschel, U. Rubritalea marina gen. nov., sp. nov., a marine representative of the phylum “Verrucomicrobia”, isolated from a sponge (Porifera). Int. J. Syst. Evol. Microbiol. 2006, 56, 2119–2124. [Google Scholar] [CrossRef]
- Kasai, H.; Katsuta, A.; Sekiguchi, H.; Matsuda, S.; Adachi, K.; Shindo, K.; Yoon, J.; Yokota, A.; Shizuri, Y. Rubritalea squalenifaciens sp. nov., a squalene-producing marine bacterium belonging to subdivision 1 of the phylum “Verrucomicrobia”. Int. J. Syst. Evol. Microbiol. 2007, 57, 1630–1634. [Google Scholar] [CrossRef]
- Yoon, J.; Matsuo, Y.; Matsuda, S.; Adachi, K.; Kasai, H.; Yokota, A. Rubritalea spongiae sp. nov. and Rubritalea tangerina sp. nov., two carotenoid- and squalene-producing marine bacteria of the family Verrucomicrobiaceae within the phylum “Verrucomicrobia”, isolated from marine animals. Int. J. Syst. Evol. Microbiol. 2007, 57, 2337–2343. [Google Scholar] [CrossRef]
- Cuc, N.T.K.; Dat, T.T.H.; Hong, T.T.; Cuong, P.V. Phylogenetic diversity of microorganisms associated with three marine sponges from Mien Trung Sea of Vietnam. J. Sci. Technol. 2017, 55, 168–177. [Google Scholar] [CrossRef]
- Kaluzhnaya, O.; Itskovich, V. Diversity of potential producers of bioactive metabolites having polyketide nature in the Baikal sponge community of Rezinkovia echinata. Limnol. Freshw. Biol. 2020, 2020, 423–428. [Google Scholar] [CrossRef]
- Sizikov, S.; Burgsdorf, I.; Handley, K.M.; Lahyani, M.; Haber, M.; Steindler, L. Characterization of sponge-associated Verrucomicrobia: Microcompartment-based sugar utilization and enhanced toxin–antitoxin modules as features of host-associated Opitutales. Environ. Microbiol. 2020, 22, 4669–4688. [Google Scholar] [CrossRef] [PubMed]
- Wagner, M.; Horn, M. The Planctomycetes, Verrucomicrobia, Chlamydiae and sister phyla comprise a superphylum with biotechnological and medical relevance. Curr. Opin. Biotechnol. 2006, 17, 241–249. [Google Scholar] [CrossRef] [PubMed]
- DeVos, D.P.; Ward, N.L. Mind the PVCs. Environ. Microbiol. 2014, 16, 1217–1221. [Google Scholar] [CrossRef]
- Spring, S.; Bunk, B.; Spröer, C.; Schumann, P.; Rohde, M.; Tindall, B.J.; Klenk, H.P. Characterization of the first cultured representative of Verrucomicrobia subdivision 5 indicates the proposal of a novel phylum. ISME J. 2016, 10, 2801–2816. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.; Matsuo, Y.; Matsuda, S.; Adachi, K.; Kasai, H.; Yokota, A. Rubritalea sabuli sp. nov., a carotenoid- and squalene-producing member of the family Verrucomicrobiaceae, isolated from marine sediment. Int. J. Syst. Evol. Microbiol. 2008, 58, 992–997. [Google Scholar] [CrossRef]
- Yoon, J.; Matsuda, S.; Adachi, K.; Kasai, H.; Yokota, A. Rubritalea halochordaticola sp. nov., a carotenoid producing verrucomicrobial species isolated from a marine chordate. Int. J. Syst. Evol. Microbiol. 2011, 61, 1515–1520. [Google Scholar] [CrossRef] [PubMed]
- Shindo, K.; Asagi, E.; Sano, A.; Hotta, E.; Minemura, N.; Mikami, K.; Tamesada, E.; Misawa, N.; Maoka, T. Diapolycopenedioic acid xylosyl esters A, B, and C, novel antioxidative glyco-C30-carotenoic acids produced by a new marine bacterium Rubritalea Squalenifaciens. J. Antibiot. 2008, 61, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Shindo, K.; Misawa, N. New and rare carotenoids isolated from marine bacteria and their antioxidant activities. Mar. Drugs 2014, 12, 1690–1698. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, A.; Naughton, L.M.; Dobson, A.D.W.; Rai, D.K. High-performance liquid chromatography/electrospray ionisation mass spectrometric characterisation of metabolites produced by Pseudovibrio sp. W64, a marine sponge derived bacterium isolated from Irish waters. Rapid Commun. Mass Spectrom. 2018, 32, 1737–1745. [Google Scholar] [CrossRef]
- Romano, S. Ecology and biotechnological potential of bacteria belonging to the genus Pseudovibrio. Appl. Environ. Microbiol. 2018, 84, e02516-17. [Google Scholar] [CrossRef]
- Brinkhoff, T.; Bach, G.; Heidorn, T.; Liang, L.; Schlingloff, A.; Simon, M. Antibiotic production by a Roseobacter clade-affiliated species from the German Wadden Sea and its antagonistic effects on indigenous isolates. Appl. Environ. Microbiol. 2004, 70, 2560–2565. [Google Scholar] [CrossRef] [PubMed]
- Bruhn, J.B.; Gram, L.; Belas, R. Production of antibacterial compounds and biofilm formation by Roseobacter species are influenced by culture conditions. Appl. Environ. Microbiol. 2007, 73, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Cerqueda-García, D.; García-Maldonado, J.Q.; Aguirre-Macedo, L.; García-Cruz, U. A succession of marine bacterial communities in batch reactor experiments during the degradation of five different petroleum types. Mar. Pollut. Bull. 2020, 150, 110775. [Google Scholar] [CrossRef]
- Peng, C.; Tang, Y.; Yang, H.; He, Y.; Liu, Y.; Liu, D.; Qian, Y.; Lu, L. Time- and compound-dependent microbial community compositions and oil hydrocarbon degrading activities in seawater near the Chinese Zhoushan Archipelago. Mar. Pollut. Bull. 2020, 152, 110907. [Google Scholar] [CrossRef] [PubMed]
- Andreo-Vidal, A.; Sanchez-Amat, A.; Campillo-Brocal, J.C. The Pseudoalteromonas luteoviolacea L-amino acid oxidase with antimicrobial activity is a flavoenzyme. Mar. Drugs 2018, 16, 499. [Google Scholar] [CrossRef]
- Montalvo, N.F.; Mohamed, N.M.; Enticknap, J.J.; Hill, R.T. Novel actinobacteria from marine sponges. Antonie Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2005, 87, 29–36. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, W.; Jin, Y.; Jin, M.; Yu, X. A comparative study on the phylogenetic diversity of culturable actinobacteria isolated from five marine sponge species. Antonie van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2008, 93, 241–248. [Google Scholar] [CrossRef]
- Xin, Y.; Huang, J.; Deng, M.; Zhang, W. Culture-independent nested PCR method reveals high diversity of actinobacteria associated with the marine sponges Hymeniacidon perleve and Sponge sp. Antonie Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2008, 94, 533–542. [Google Scholar] [CrossRef]
- Selvin, J.; Gandhimathi, R.; Kiran, G.S.; Priya, S.S.; Ravji, T.R.; Hema, T.A. Culturable heterotrophic bacteria from the marine sponge Dendrilla nigra: Isolation and phylogenetic diversity of actinobacteria. Helgol. Mar. Res. 2009, 63, 239–247. [Google Scholar] [CrossRef]
- Sun, W.; Dai, S.; Jiang, S.; Wang, G.; Liu, G.; Wu, H.; Li, X. Culture-dependent and culture-independent diversity of Actinobacteria associated with the marine sponge Hymeniacidon perleve from the South China Sea. Antonie Leeuwenhoek, Int. J. Gen. Mol. Microbiol. 2010, 98, 65–75. [Google Scholar] [CrossRef]
- Li, L.; Zhang, D.; Tang, W.Z.; Lin, H.W.; Lu, Y.H. Geodermatophilus marinus sp. nov., isolated from the marine sponge Leucetta chagosensis. Int. J. Syst. Evol. Microbiol. 2019, 69, 2966–2971. [Google Scholar] [CrossRef]
- Dita, S.F.; Budiarti, S.R.I.; Lestari, Y. Sponge-associated Actinobacteria: Morphological character and antibacterial activity against pathogenic bacteria. J. Sumberd. Hayati 2017, 3, 21–26. [Google Scholar]
- Xu, D.; Han, L.; Li, C.; Cao, Q.; Zhu, D.; Barrett, N.H.; Harmody, D.; Chen, J.; Zhu, H.; McCarthy, P.J.; et al. Bioprospecting deep-sea actinobacteria for novel anti-infective natural products. Front. Microbiol. 2018, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kasanah, N.; Triyanto, T. Bioactivities of halometabolites from marine Actinobacteria. Biomolecules 2019, 9, 225. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Wu, S.; Zhang, R.; Wang, D.; Chen, J.; Zhao, J. Diversity and antimicrobial potential of Actinobacteria isolated from diverse marine sponges along the Beibu Gulf of the South China Sea. FEMS Microbiol. Ecol. 2019, 95, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Phuong, T.V.; Thi, N.; Dung, P.; Diep, C.N. Diversified collection of novel sponge-associated actinobacteria of Ha Tien sea, Kien Giang province, Vietnam. Int. J. Innov. Eng. Technol. 2019, 14, 33–41. [Google Scholar]
- Puttaswamygowda, G.H.; Olakkaran, S.; Antony, A.; Purayil, A.K. Present status and future perspectives of marine actinobacterial metabolites. In Recent Developments in Applied Microbiology and Biochemistry; Buddolla, V., Ed.; Academic Press: Cambridge, CA, USA, 2019; pp. 307–319. [Google Scholar]
- Santos, J.D.; Vitorino, I.; De La Cruz, M.; Díaz, C.; Cautain, B.; Annang, F.; Pérez-Moreno, G.; Martinez, I.G.; Tormo, J.R.; Martín, J.M.; et al. Bioactivities and extract dereplication of Actinomycetales isolated from marine sponges. Front. Microbiol. 2019, 10, 1–11. [Google Scholar] [CrossRef]
- Baig, U.; Dahanukar, N.; Shintre, N.; Holkar, K.; Pund, A.; Lele, U.; Gujarathi, T.; Patel, K.; Jakati, A.; Singh, R.; et al. Phylogenetic diversity and activity screening of cultivable actinobacteria isolated from marine sponges and associated environments from the western coast of India. bioRxiv Prepr. Serv. 2020, 1–36. [Google Scholar] [CrossRef]
- Gohain, A.; Manpoong, C.; Saikia, R.; De Mandal, S. Actinobacteria: Diversity and biotechnological applications. In Recent Advancements in Microbial Diversity; De Mandal, S., Bhatt, P., Eds.; Elsevier Inc.: Berlin/Heidelberg, Germany, 2020; pp. 217–231. [Google Scholar]
- Kamala, K.; Sivaperumal, P.; Kamath, S.M.; Thilagaraj, W.R.; Rajaram, R. Marine actinobacteria as a source for emerging biopharmaceuticals. In Encyclopedia of Marine Biotechnology; Se-Kwon, K., Ed.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2020; pp. 2095–2105. [Google Scholar]
- Jiang, S.; Sun, W.; Chen, M.; Dai, S.; Zhang, L.; Liu, Y.; Lee, K.J.; Li, X. Diversity of culturable actinobacteria isolated from marine sponge Haliclona sp. Antonie Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2007, 92, 405–416. [Google Scholar] [CrossRef]
- Jiang, S.; Li, X.; Zhang, L.; Sun, W.; Dai, S.; Xie, L.; Liu, Y.; Lee, K.J. Culturable actinobacteria isolated from marine sponge Iotrochota sp. Mar. Biol. 2008, 153, 945–952. [Google Scholar] [CrossRef]
- Schneemann, I.; Nagel, K.; Kajahn, I.; Labes, A.; Wiese, J.; Imhoff, J.F. Comprehensive investigation of marine actinobacteria associated with the sponge Halichondria panicea. Appl. Environ. Microbiol. 2010, 76, 3702–3714. [Google Scholar] [CrossRef] [PubMed]
- Selvin, J.; Sathiyanarayanan, G.; Lipton, A.N.; Al-Dhabi, N.A.; Arasu, M.V.; Kiran, G.S. Ketide Synthase (KS) domain prediction and analysis of iterative type II PKS gene in marine sponge-associated actinobacteria producing biosurfactants and antimicrobial agents. Front. Microbiol. 2016, 7, 1–12. [Google Scholar] [CrossRef]
- Huang, X.; Kong, F.; Zhou, S.; Huang, D.; Zheng, J.; Zhu, W. Streptomyces tirandamycinicus sp. nov., a novel marine sponge-derived actinobacterium with antibacterial potential against Streptococcus Agalactiae. Front. Microbiol. 2019, 10, 1–11. [Google Scholar] [CrossRef]
- Madhusoodanan, J. Probing predatory bacteria as an antibacterial remedy. Proc. Natl. Acad. Sci. USA 2019, 116, 22887–22890. [Google Scholar] [CrossRef]
- Williams, L.E.; Cullen, N.; Degiorgis, J.A.; Martinez, K.J.; Mellone, J.; Oser, M.; Wang, J.; Zhang, Y. Variation in genome content and predatory phenotypes between Bdellovibrio sp. NC01 isolated from soil and B. bacteriovorus type strain HD100. Microbiology 2019, 165, 1315–1330. [Google Scholar] [CrossRef]
- Bratanis, E.; Andersson, T.; Lood, R.; Bukowska-Faniband, E. Biotechnological potential of Bdellovibrio and like organisms and their secreted enzymes. Front. Microbiol. 2020, 11, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.L.; Zhao, F.; Shi, M.; Zhang, X.Y.; Zhou, B.C.; Zhang, Y.Z.; Chen, X.L. Characterization and biotechnological potential analysis of a new exopolysaccharide from the Arctic marine bacterium Polaribacter sp. SMSci. Rep. 2015, 5, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.L.; Zhao, F.; Zhang, X.K.; Zhang, X.Y.; Zhang, Y.Z.; Song, X.Y.; Chen, X.L. Improvement of the production of an Arctic bacterial exopolysaccharide with protective effect on human skin cells against UV-induced oxidative stress. Appl. Microbiol. Biotechnol. 2020, 104, 4863–4875. [Google Scholar] [CrossRef] [PubMed]
- Rützler, K. Sponges in coral reefs. In Coral reefs: Research Methods, Monographs on Oceanographic Methodology; Stoddart, D.R., Johannes, R.E., Eds.; UNESCO: Paris, France, 1978; pp. 299–313. [Google Scholar]
- Hooper, J.N.A. ‘Sponguide’. Guide to sponge collection and identification. Available online: https://www.academia.edu/34258606/SPONGE_GUIDE_GUIDE_TO_SPONGE_COLLECTION_AND_IDENTIFICATION_Version_August_2000 (accessed on 17 March 2021).
- Van Soest, R.W.M.; Boury-Esnault, N.; Hooper, J.N.A.; Rützler, K.; de Voogd, N.J.; Alvarez, B.; Hajdu, E.; Pisera, A.B.; Manconi, R.; Schönberg, C.; et al. World Porifera Database. Available online: http://www.marinespecies.org/porifera/ (accessed on 17 March 2021).
- Schmitt, S.; Hentschel, U.; Zea, S.; Dandekar, T.; Wolf, M. ITS-2 and 18S rRNA gene phylogeny of Aplysinidae (Verongida, Demospongiae). J. Mol. Evol. 2005, 60, 327–336. [Google Scholar] [CrossRef]
- Chombard, C.; Boury-Esnault, N.; Tillier, S. Reassessment of homology of morphological characters in Tetractinellid sponges based on molecular data. Syst. Biol. 1998, 47, 351–366. [Google Scholar] [CrossRef]
- Dohrmann, M.; Janussen, D.; Reitner, J.; Collins, A.G.; Wörheide, G. Phylogeny and evolution of glass sponges (Porifera, Hexactinellida). Syst. Biol. 2008, 57, 388–405. [Google Scholar] [CrossRef]
- Meyer, C.P.; Geller, J.B.; Paulay, G. Fine scale endemism on coral reefs: Archipelagic differentiation in turbinid gastropods. Evolution (N. Y). 2005, 59, 113–125. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Corpet, F. Multiple sequence alignment with hierarchical clustering. Nucleic Acids Res. 1988, 16, 10881–10890. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Babicki, S.; Arndt, D.; Marcu, A.; Liang, Y.; Grant, J.R.; Maciejewski, A.; Wishart, D.S. Heatmapper: Web-enabled heat mapping for all. Nucleic Acids Res. 2016, 44, W147–W153. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Site | Sample ID | Sponge Taxonomy | MNA code | Sampling Method | Sampling Depth (m) | Coordinates |
|---|---|---|---|---|---|---|
| 1 | B4 | Mycale (Oxymycale) acerata (Kirkpatrick, 1907) | 13264 | SCUBA | 26 | 74°42.067’S 164°02.518’E |
| 2 | C6 | Haliclona (Rhizoniera) dancoi (Topsent, 1901) | 13265 | SCUBA | 28 | 74°40.537’S 164°04.169’E |
| 2 | D4 | Hemigellius pilosus (Kirkpatrick, 1907) | 13266 | SCUBA | 28 | 74°40.537’S 164°04.169’E |
| 2 | D6 | Microxina sarai (Calcinai & Pansini, 2000) | 13267 | SCUBA | 28 | 74°40.537’S 164°04.169’E |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruocco, N.; Esposito, R.; Bertolino, M.; Zazo, G.; Sonnessa, M.; Andreani, F.; Coppola, D.; Giordano, D.; Nuzzo, G.; Lauritano, C.; et al. A Metataxonomic Approach Reveals Diversified Bacterial Communities in Antarctic Sponges. Mar. Drugs 2021, 19, 173. https://doi.org/10.3390/md19030173
Ruocco N, Esposito R, Bertolino M, Zazo G, Sonnessa M, Andreani F, Coppola D, Giordano D, Nuzzo G, Lauritano C, et al. A Metataxonomic Approach Reveals Diversified Bacterial Communities in Antarctic Sponges. Marine Drugs. 2021; 19(3):173. https://doi.org/10.3390/md19030173
Chicago/Turabian StyleRuocco, Nadia, Roberta Esposito, Marco Bertolino, Gianluca Zazo, Michele Sonnessa, Federico Andreani, Daniela Coppola, Daniela Giordano, Genoveffa Nuzzo, Chiara Lauritano, and et al. 2021. "A Metataxonomic Approach Reveals Diversified Bacterial Communities in Antarctic Sponges" Marine Drugs 19, no. 3: 173. https://doi.org/10.3390/md19030173
APA StyleRuocco, N., Esposito, R., Bertolino, M., Zazo, G., Sonnessa, M., Andreani, F., Coppola, D., Giordano, D., Nuzzo, G., Lauritano, C., Fontana, A., Ianora, A., Verde, C., & Costantini, M. (2021). A Metataxonomic Approach Reveals Diversified Bacterial Communities in Antarctic Sponges. Marine Drugs, 19(3), 173. https://doi.org/10.3390/md19030173