
Adding Zooplankton to the OSMAC Toolkit: Effect of Grazing Stress on the Metabolic Profile and Bioactivity of a Diatom

 ,
,  , , ,
, , ,
Abstract
1. Introduction
2. Results
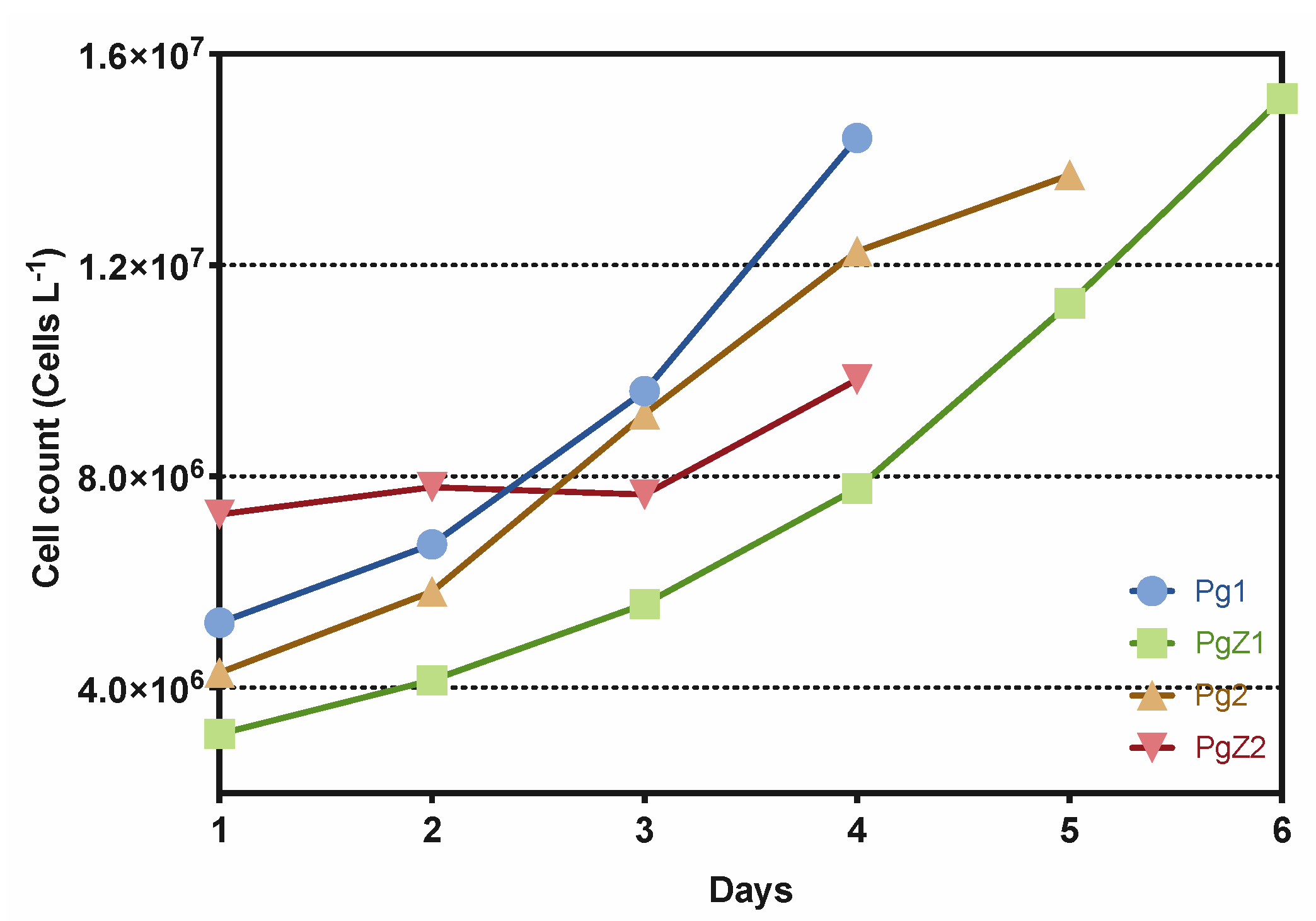
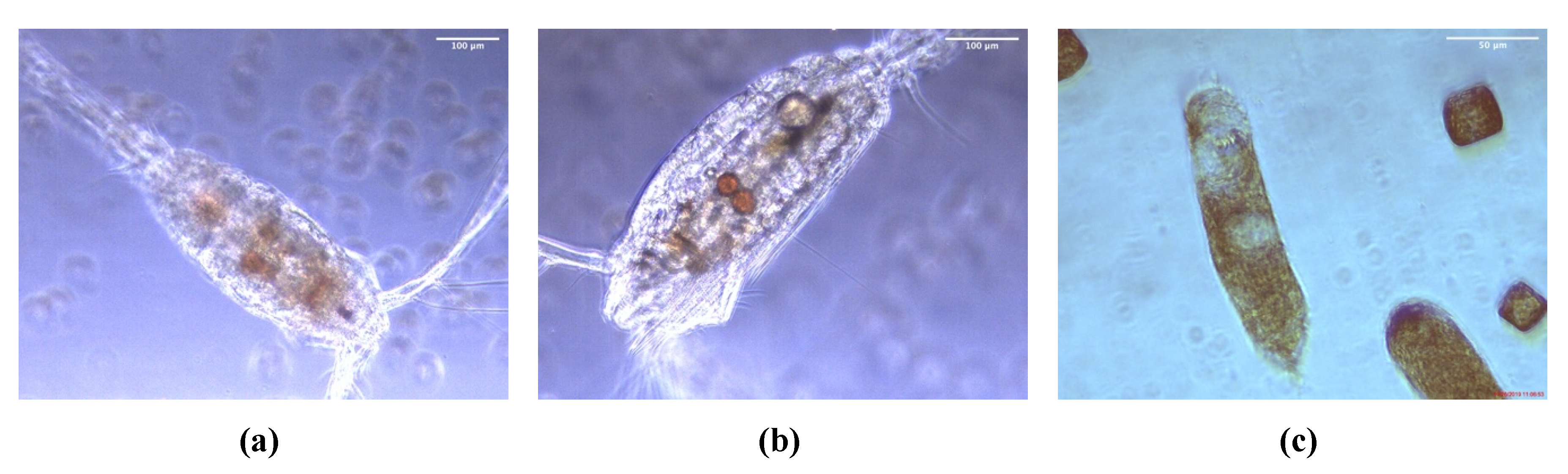
2.1. Cultivation of P. glacialis and Cocultivation with Zooplankton
2.2. Extraction and Fractionation
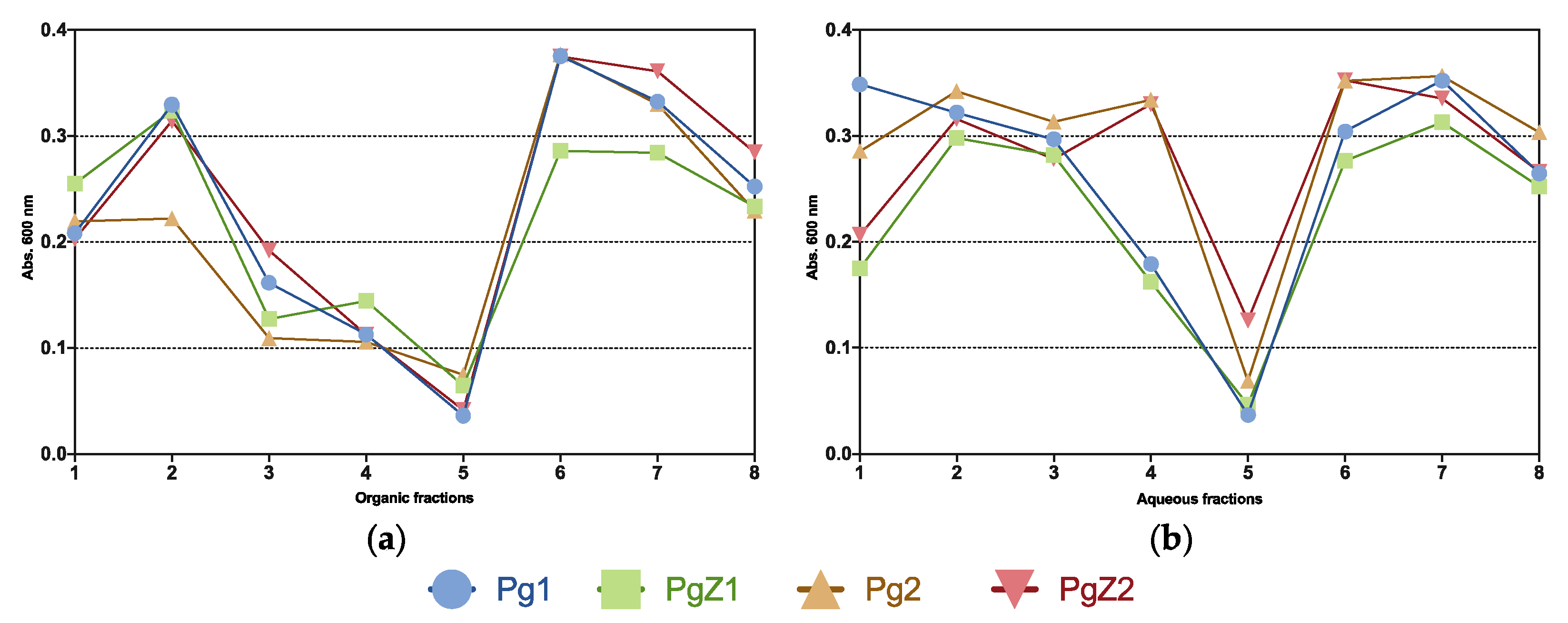
2.3. Inhibition of Biofilm Formation
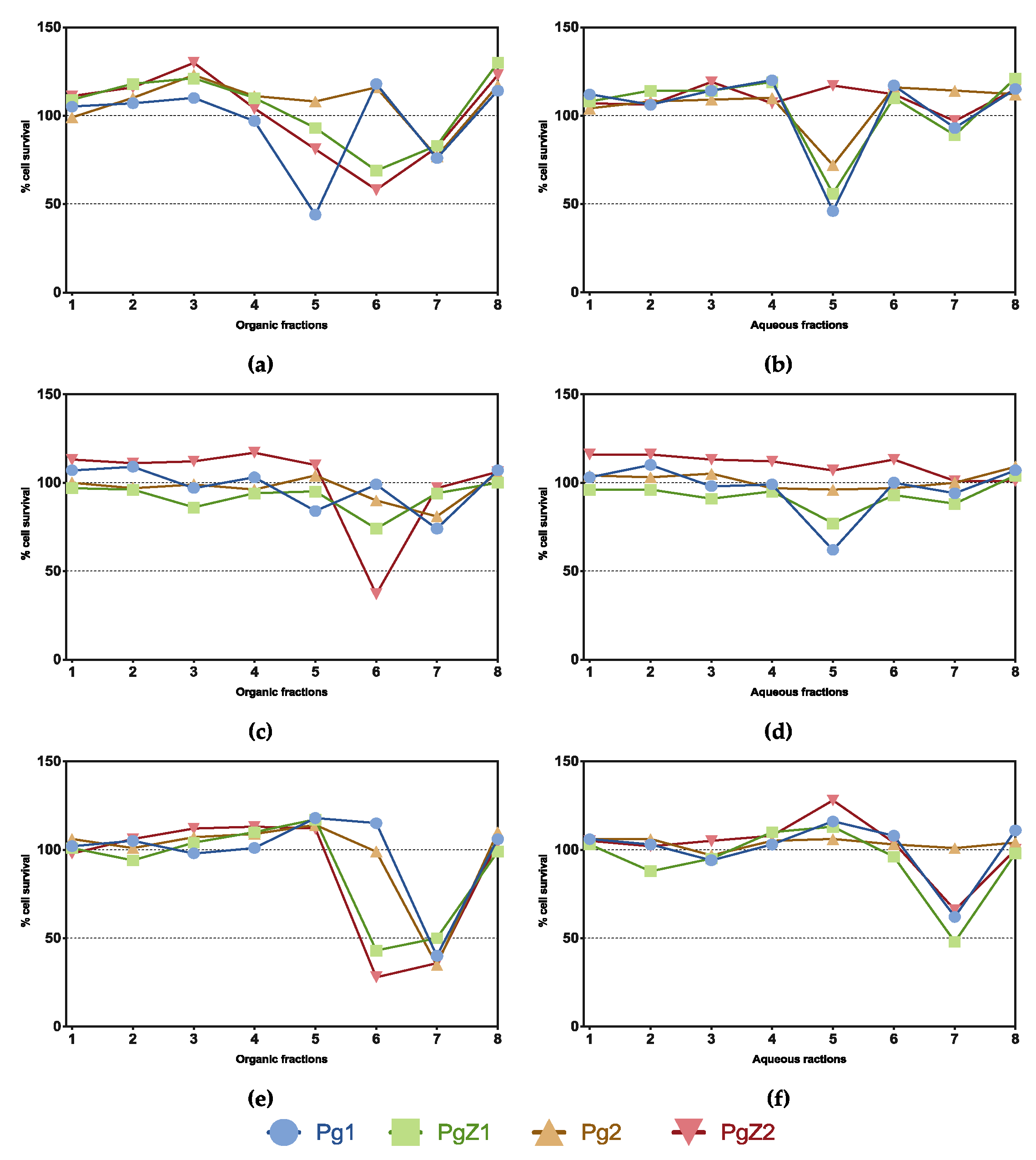
2.4. Cell Viability
2.5. Bacterial Growth Inhibition Assay
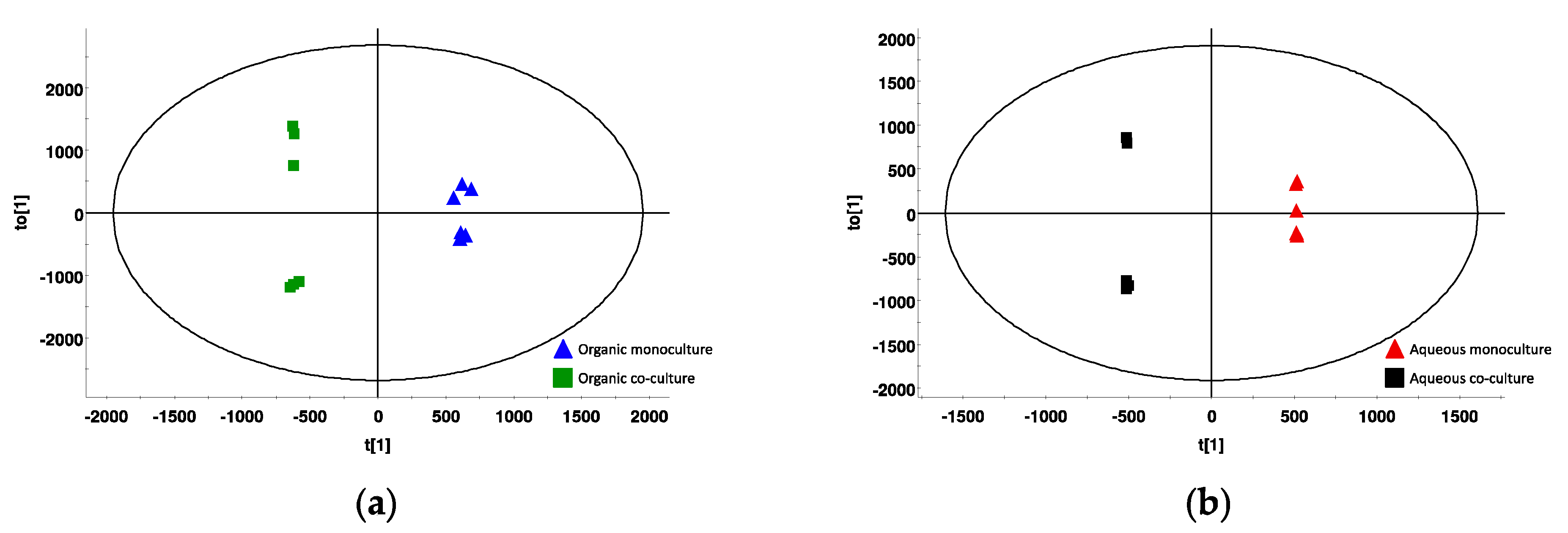
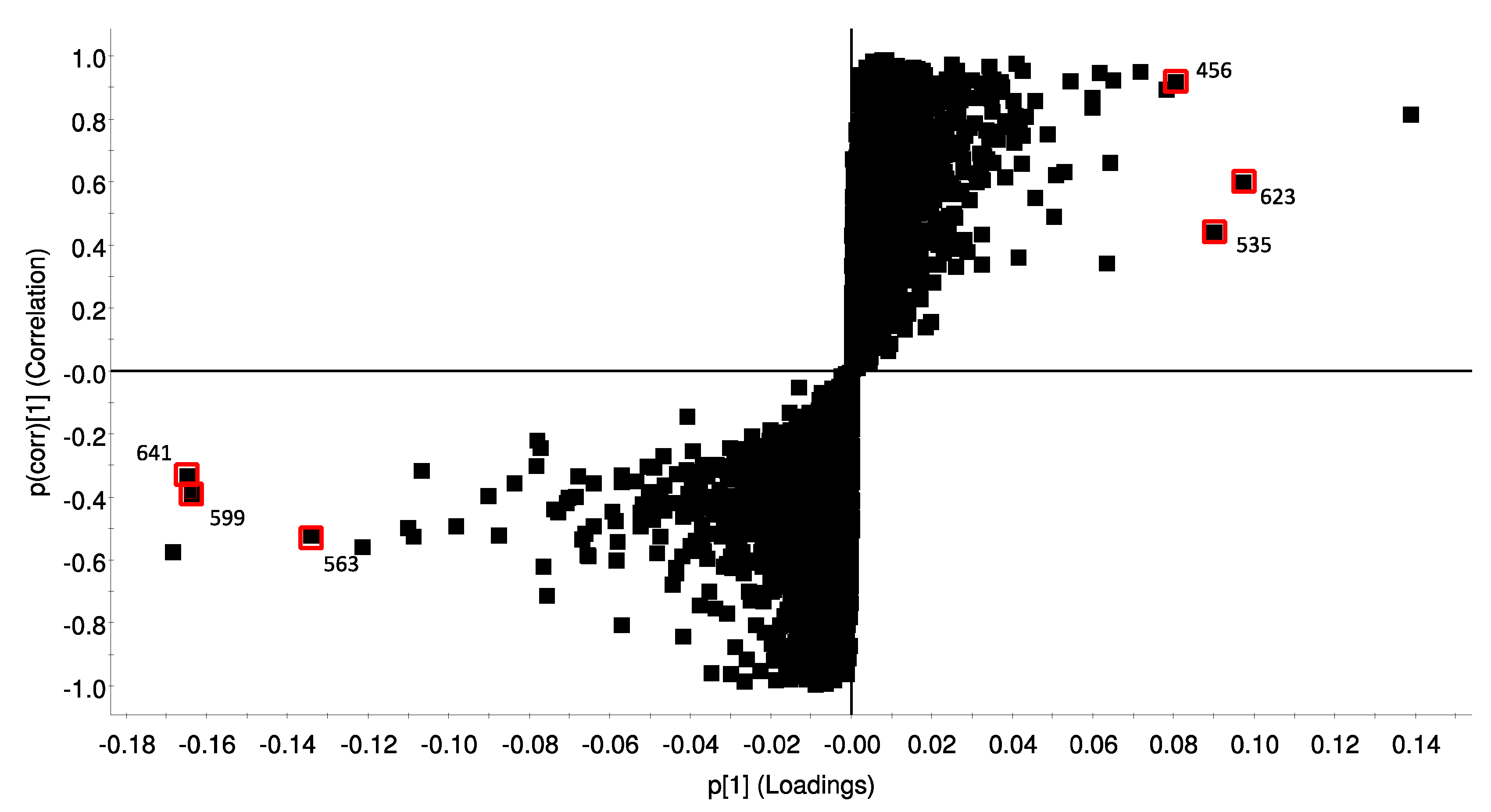
2.6. Metabolic Profile
3. Discussion
4. Materials and Methods
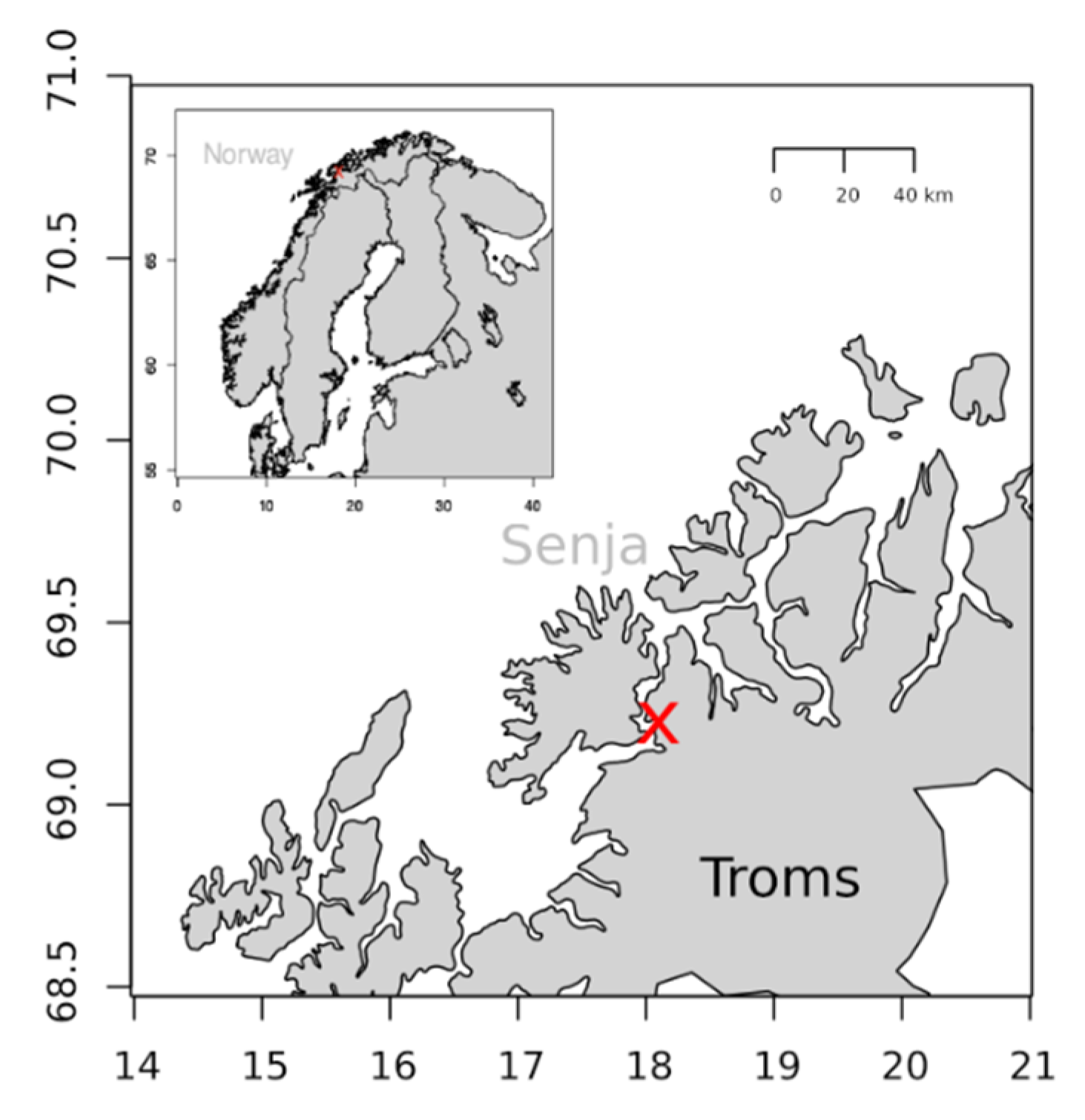
4.1. Mass Cultivation of Porosira Glacialis
4.2. Zooplankton Collection
4.3. Cocultivaion
4.4. Extraction and Flash-Fractionation
4.5. Biofilm Assay
4.6. Growth Inhibtion Assay
4.7. Cell Viability Assay
4.8. UHPLC-ESI-HR-MS Analysis and Data Processing for Metabolomic Profile
4.9. Statistics and Software
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Demain, A.L.; Sanchez, S. Microbial drug discovery: 80 years of progress. J. Antibiot. 2009, 62, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Romano, S.; Jackson, S.A.; Patry, S.; Dobson, A.D. Extending the “one strain many compounds” (OSMAC) principle to marine microorganisms. Mar. Drugs 2018, 16, 244. [Google Scholar] [CrossRef] [PubMed]
- Bode, H.B.; Bethe, B.; Höfs, R.; Zeeck, A.J.C. Big effects from small changes: Possible ways to explore nature’s chemical diversity. ChemBioChem 2002, 3, 619–627. [Google Scholar] [CrossRef]
- Uchoa, P.K.S.; Pimenta, A.T.A.; Braz-Filho, R.; Oliveira, M.D.C.F.D.; Saraiva, N.N.; Rodrigues, B.S.F.; Pfenning, L.H.; Abreu, L.M.; Wilke, D.V.; Florêncio, K.G.D.; et al. New cytotoxic furan from the marine sediment-derived fungi Aspergillus niger. Nat. Prod. Res. 2017, 31, 2599–2603. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Chen, G.; Chen, X.; Huang, M.; Wan, X. Aspergicin, a new antibacterial alkaloid produced by mixed fermentation of two marine-derived mangrove epiphytic fungi. Chem. Nat. Compd. 2011, 47, 767–769. [Google Scholar] [CrossRef]
- Okami, Y.; Okazaki, T.; Kitahara, T.; Umezawa, H. Studies on marine microorganisms. V. A new antibiotic, aplasmomycin, produced by a streptomycete isolated from shallow sea mud. J. Antibiot. 1976, 29, 1019–1025. [Google Scholar] [CrossRef]
- Pan, R.; Bai, X.; Chen, J.; Zhang, H.; Wang, H. Exploring Structural Diversity of Microbe Secondary Metabolites Using OSMAC Strategy: A Literature Review. Front. Microbiol. 2019, 10, 294. [Google Scholar] [CrossRef]
- Si, Y.; Tang, M.; Lin, S.; Chen, G.; Feng, Q.; Wang, Y.; Hua, H.; Bai, J.; Wang, H.; Pei, Y.-H. Cytotoxic cytochalasans from Aspergillus flavipes PJ03-11 by OSMAC method. Tetrahedron Lett. 2018, 59, 1767–1771. [Google Scholar] [CrossRef]
- Sung, A.A.; Gromek, S.M.; Balunas, M.J. Upregulation and Identification of Antibiotic Activity of a Marine-Derived Streptomyces sp. via Co-Cultures with Human Pathogens. Mar. Drugs 2017, 15, 250. [Google Scholar] [CrossRef]
- Jensen, P.R.; Fenical, W. Marine bacterial diversity as a resource for novel microbial products. J. Ind. Microbiol. Biotechnol. 1996, 17, 346–351. [Google Scholar] [CrossRef]
- Mann, D.G. The species concept in diatoms. Phycologia 1999, 38, 437–495. [Google Scholar] [CrossRef]
- Lauritano, C.; Andersen, J.H.; Hansen, E.; Albrigtsen, M.; Escalera, L.; Esposito, F.; Helland, K.; Hanssen, K.Ø.; Romano, G.; Ianora, A. Bioactivity Screening of Microalgae for Antioxidant, Anti-Inflammatory, Anticancer, Anti-Diabetes, and Antibacterial Activities. Front. Mar. Sci. 2016, 3, 68. [Google Scholar] [CrossRef]
- Ingebrigtsen, R.A.; Hansen, E.; Andersen, J.H.; Eilertsen, H.C. Light and temperature effects on bioactivity in diatoms. Environ. Boil. Fishes 2016, 28, 939–950. [Google Scholar] [CrossRef] [PubMed]
- Degerlund, M.; Eilertsen, H.C. Main Species Characteristics of Phytoplankton Spring Blooms in NE Atlantic and Arctic Waters (68–80° N). Chesap. Sci. 2009, 33, 242–269. [Google Scholar] [CrossRef]
- Braarud, T.; Gaarder, K.R.; Nordli, O. Seasonal changes in the phytoplankton at various points off the Norwegian West Coast:(Observations at the permanent oceanographic stations, 1945–1946). Rep. Nor. Fish. Mar. Investig. 1958, 12, 1–77. [Google Scholar]
- Wiborg, K.F. Investigations on zooplankton in coastal and offshore waters of western and northwestern Norway—With special reference to the copepods. Rep. Nor. Fish. Mar. Investig. 1954, 11, 1–246. [Google Scholar]
- Cushing, D. A difference in structure between ecosystems in strongly stratified waters and in those that are only weakly stratified. J. Plankton Res. 1989, 11, 1–13. [Google Scholar] [CrossRef]
- Legendre, L. The significance of microalgal blooms for fisheries and for the export of particulate organic carbon in oceans. J. Plankton Res. 1990, 12, 681–699. [Google Scholar] [CrossRef]
- Caldwell, G. The Influence of Bioactive Oxylipins from Marine Diatoms on Invertebrate Reproduction and Development. Mar. Drugs 2009, 7, 367–400. [Google Scholar] [CrossRef]
- Gudimova, E.; Eilertsen, H.C.; Jørgensen, T.Ø.; Hansen, E. In vivo exposure to northern diatoms arrests sea urchin embryonic development. Toxicon 2016, 109, 63–69. [Google Scholar] [CrossRef]
- Harðardóttir, S.; Pančić, M.; Tammilehto, A.; Krock, B.; Møller, E.F.; Nielsen, T.G.; Lundholm, N. Dangerous Relations in the Arctic Marine Food Web: Interactions between Toxin Producing Pseudo-nitzschia Diatoms and Calanus Copepodites. Mar. Drugs 2015, 13, 3809–3835. [Google Scholar] [CrossRef] [PubMed]
- Ianora, A.; Miralto, A. Toxigenic effects of diatoms on grazers, phytoplankton and other microbes: A review. Ecotoxicology 2009, 19, 493–511. [Google Scholar] [CrossRef] [PubMed]
- Ianora, A.; Poulet, S.A.; Miralto, A. A comparative study of the inhibitory effect of diatoms on the reproductive biology of the copepod Temora stylifera. Mar. Biol. 1995, 121, 533–539. [Google Scholar] [CrossRef]
- Ianora, A.; Poulet, S.A.; Miralto, A. The effects of diatoms on copepod reproduction: A review. Phycologia 2003, 42, 351–363. [Google Scholar] [CrossRef]
- Lauritano, C.; Borra, M.; Carotenuto, Y.; Biffali, E.; Miralto, A.; Procaccini, G.; Ianora, A. First molecular evidence of diatom effects in the copepod Calanus helgolandicus. J. Exp. Mar. Biol. Ecol. 2011, 404, 79–86. [Google Scholar] [CrossRef]
- Pohnert, G. Diatom/Copepod Interactions in Plankton: The Indirect Chemical Defense of Unicellular Algae. ChemBioChem 2005, 6, 946–959. [Google Scholar] [CrossRef]
- Pohnert, G.; Lumineau, O.; Cueff, A.; Adolph, S.; Cordevant, C.; Lange, M.; Poulet, S. Are volatile unsaturated aldehydes from diatoms the main line of chemical defence against copepods? Mar. Ecol. Prog. Ser. 2002, 245, 33–45. [Google Scholar] [CrossRef]
- Wichard, T.; Poulet, S.A.; Halsband-Lenk, C.; Albaina, A.; Harris, R.; Liu, D.; Pohnert, G.J. Survey of the chemical defence potential of diatoms: Screening of fifty species for α, β, γ, δ-unsaturated aldehydes. J. Chem. Ecol. 2005, 31, 949–958. [Google Scholar] [CrossRef]
- Ianora, A.; Poulet, S.A. Egg viability in the copepod Temora stylifera. Limnol. Oceanogr. 1993, 38, 1615–1626. [Google Scholar] [CrossRef]
- Miralto, A.; Barone, G.; Romano, G.; Poulet, S.A.; Ianora, A.; Russo, G.L.; Buttino, I.; Mazzarella, G.; Laabir, M.; Cabrini, M.; et al. The insidious effect of diatoms on copepod reproduction. Nat. Cell Biol. 1999, 402, 173–176. [Google Scholar] [CrossRef]
- Chaudron, Y.; Poulet, S.A.; Laabir, M.; Ianora, A.; Miralto, A. Is hatching success of copepod eggs diatom density-dependent? Mar. Ecol. Prog. Ser. 1996, 144, 185–193. [Google Scholar] [CrossRef][Green Version]
- Poulet, S.; Laabir, M.; Ianora, A.; Miralto, A. Reproductive response of Calanus helgolandicus. I. Abnormal embryonic and naupliar development. Mar. Ecol. Prog. Ser. 1995, 129, 85–95. [Google Scholar] [CrossRef]
- Pohnert, G. Wound-Activated Chemical Defense in Unicellular Planktonic Algae. Angew. Chem. Int. Ed. 2000, 39, 4352–4354. [Google Scholar] [CrossRef]
- Selander, E.; Kubanek, J.; Hamberg, M.; Andersson, M.X.; Cervin, G.; Pavia, H. Predator lipids induce paralytic shellfish toxins in bloom-forming algae. Proc. Natl. Acad. Sci. USA 2015, 112, 6395–6400. [Google Scholar] [CrossRef] [PubMed]
- Selander, E.; Thor, P.; Toth, G.; Pavia, H. Copepods induce paralytic shellfish toxin production in marine dinoflagellates. Proc. R. Soc. B Boil. Sci. 2006, 273, 1673–1680. [Google Scholar] [CrossRef] [PubMed]
- Bergkvist, J.; Thor, P.; Jakobsen, H.H.; Wängberg, S.Å.; Selander, E. Grazer-induced chain length plasticity reduces grazing risk in a marine diatom. Limnol. Oceanogr. 2012, 57, 318–324. [Google Scholar] [CrossRef]
- Selander, E.; Berglund, E.C.; Engström, P.; Berggren, F.; Eklund, J.; Harðardóttir, S.; Lundholm, N.; Grebner, W.; Andersson, M.X. Copepods drive large-scale trait-mediated effects in marine plankton. Sci. Adv. 2019, 5, eaat5096. [Google Scholar] [CrossRef]
- Lundholm, N.; Krock, B.; John, U.; Skov, J.; Cheng, J.; Pančić, M.; Wohlrab, S.; Rigby, K.; Nielsen, T.G.; Selander, E.; et al. Induction of domoic acid production in diatoms—Types of grazers and diatoms are important. Harmful Algae 2018, 79, 64–73. [Google Scholar] [CrossRef]
- Krug, D.; Müller, R. Secondary metabolomics: The impact of mass spectrometry-based approaches on the discovery and characterization of microbial natural products. Nat. Prod. Rep. 2014, 31, 768–783. [Google Scholar] [CrossRef]
- Wishart, D.S. Emerging applications of metabolomics in drug discovery and precision medicine. Nat. Rev. Drug Discov. 2016, 15, 473–484. [Google Scholar] [CrossRef]
- Matsuno, T.; Ookubo, M. A new carotenoid, halocynthiaxanthin from the sea squirt. Tetrahedron Lett. 1981, 22, 4659–4660. [Google Scholar] [CrossRef]
- Liaaen-Jensen, S.; Renstrøm, B.; Ramdahl, T.; Hallenstvet, M.; Bergquist, P. Carotenoids of Marine Sponges. Biochem. Syst. Ecol. 1982, 10, 167–174. [Google Scholar] [CrossRef]
- Litchfield, C.; Liaaen-Jensen, S. Carotenoids of the marine sponge Microciona prolifera. Comp. Biochem. Physiol. Part B Comp. Biochem. 1980, 66, 359–365. [Google Scholar] [CrossRef]
- Mosadeghzad, Z.; Zuriati, Z.; Asmat, A.; Gires, U.; Wickneswari, R.; Pittayakhajonwut, P.; Farahani, G. Chemical components and bioactivity of the marine-derived fungus Paecilomyces sp. collected from Tinggi Island, Malaysia. Chem. Nat. Compd. 2013, 49, 621–625. [Google Scholar] [CrossRef]
- Huseby, S.; Degerlund, M.; Eriksen, G.K.; Ingebrigtsen, R.A.; Eilertsen, H.C.; Hansen, E. Chemical Diversity as a Function of Temperature in Six Northern Diatom Species. Mar. Drugs 2013, 11, 4232–4245. [Google Scholar] [CrossRef] [PubMed]
- Søreide, J.E.; Falk-Petersen, S.; Hegseth, E.N.; Hop, H.; Carroll, M.L.; Hobson, K.A.; Blachowiak-Samolyk, K. Seasonal feeding strategies of Calanus in the high-Arctic Svalbard region. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2008, 55, 2225–2244. [Google Scholar] [CrossRef]
- Barth-Jensen, C.; Koski, M.; Varpe, Ø.; Glad, P.; Wangensteen, O.S.; Præbel, K.; Svensen, C. Temperature-dependent egg production and egg hatching rates of small egg-carrying and broadcast-spawning copepods Oithona similis, Microsetella norvegica and Microcalanus pusillus. J. Plankton Res. 2020, 42, 564–580. [Google Scholar] [CrossRef]
- Castellani, C.; Irigoien, X.; Harris, R.P.; Holliday, N.P. Regional and temporal variation of Oithona spp. biomass, stage structure and productivity in the Irminger Sea, North Atlantic. J. Plankton Res. 2007, 29, 1051–1070. [Google Scholar] [CrossRef][Green Version]
- Norrbin, M.F. Seasonal patterns in gonad maturation, sex ratio and size in some small, high-latitude copepods: Implications for overwintering tactics. J. Plankton Res. 1994, 16, 115–131. [Google Scholar] [CrossRef]
- Durbin, A.G.; Durbin, E.G. Standing stock and estimated production rates of phytoplankton and zooplankton in Narragansett Bay, Rhode Island. Estuaries 1981, 4, 24–41. [Google Scholar] [CrossRef]
- Preisser, E.L.; Bolnick, D.I.; Benard, M.F. Scared to Death? The Effects of Intimidation and Consumption in Predator–Prey Interactions. Ecology 2005, 86, 501–509. [Google Scholar] [CrossRef]
- Amin, S.A.; Parker, M.S.; Armbrust, E.V. Interactions between Diatoms and Bacteria. Microbiol. Mol. Biol. Rev. 2012, 76, 667–684. [Google Scholar] [CrossRef] [PubMed]
- Sansone, C.; Braca, A.; Ercolesi, E.; Romano, G.; Palumbo, A.; Casotti, R.; Francone, M.; Ianora, A. Diatom-derived polyunsaturated aldehydes activate cell death in human cancer cell lines but not normal cells. PLoS ONE 2014, 9, e101220. [Google Scholar] [CrossRef] [PubMed]
- Ingebrigtsen, R.A.; Hansen, E.; Andersen, J.H.; Eilertsen, H.C. Field sampling marine plankton for biodiscovery. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.-H.; Wang, H.-K.; Ito, J.; Bastow, K.F.; Tachibana, Y.; Nakanishi, Y.; Xu, Z.; Luo, T.-Y.; Lee, K.-H. Cytotoxic Pheophorbide-Related Compounds from Clerodendrum calamitosum and C. cyrtophyllum. J. Nat. Prod. 2001, 64, 915–919. [Google Scholar] [CrossRef] [PubMed]
- Andrade, K.A.M.; Lauritano, C.; Romano, G.; Ianora, A. Marine Microalgae with Anti-Cancer Properties. Mar. Drugs 2018, 16, 165. [Google Scholar] [CrossRef]
- Peng, J.; Yuan, J.-P.; Wu, C.-F.; Wang, J.-H. Fucoxanthin, a Marine Carotenoid Present in Brown Seaweeds and Diatoms: Metabolism and Bioactivities Relevant to Human Health. Mar. Drugs 2011, 9, 1806–1828. [Google Scholar] [CrossRef]
- Cha, K.H.; Koo, S.Y.; Lee, D.-U. Antiproliferative Effects of Carotenoids Extracted from Chlorella ellipsoidea and Chlorella vulgaris on Human Colon Cancer Cells. J. Agric. Food Chem. 2008, 56, 10521–10526. [Google Scholar] [CrossRef]
- Bjaerke, O.; Jonsson, P.R.; Alam, A.; Selander, E. Is chain length in phytoplankton regulated to evade predation? J. Plankton Res. 2015, 37, 1110–1119. [Google Scholar] [CrossRef]
- Amato, A.; Sabatino, V.; Nylund, G.M.; Bergkvist, J.; Basu, S.; Andersson, M.X.; Sanges, R.; Godhe, A.; Kiørboe, T.; Selander, E.; et al. Grazer-induced transcriptomic and metabolomic response of the chain-forming diatom Skeletonema marinoi. ISME J. 2018, 12, 1594–1604. [Google Scholar] [CrossRef]
- Gieskes, W.W.C.; Engelkes, M.M.; Grakaay, G.W. Degradation of diatom chlorophyll to colourless, non-fluorescing compounds during copepoo grazing. Aquat. Ecol. 1991, 25, 65–72. [Google Scholar] [CrossRef]
- Shuman, F.R.; Lorenzen, C.J. Quantitative degradation of chlorophyll by a marine herbivore1. Limnol. Oceanogr. 1975, 20, 580–586. [Google Scholar] [CrossRef]
- Chuyen, H.V.; Eun, J.-B. Marine carotenoids: Bioactivities and potential benefits to human health. Crit. Rev. Food Sci. Nutr. 2017, 57, 2600–2610. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, M. Carotenoid biosynthesis in diatoms. Photosynth. Res. 2010, 106, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Krinsky, N.I.; Johnson, E.J. Carotenoid actions and their relation to health and disease. Mol. Asp. Med. 2005, 26, 459–516. [Google Scholar] [CrossRef] [PubMed]
- Rmiki, N.-E.; Schoefs, B.; Lemoine, Y. Carotenoids and Stress in Higher Plants and Algae. In Pesticides in Agriculture and the Environment; Prakash, A., Rao, J., Eds.; CRC Press: Boca Raton, FL, USA, 1999; pp. 465–482. [Google Scholar]
- Steinbrenner, J.; Linden, H. Regulation of Two Carotenoid Biosynthesis Genes Coding for Phytoene Synthase and Carotenoid Hydroxylase during Stress-Induced Astaxanthin Formation in the Green Alga Haematococcus pluvialis. Plant Physiol. 2001, 125, 810–817. [Google Scholar] [CrossRef]
- Vidhyavathi, R.; Venkatachalam, L.; Ravi, S.; Ravishankar, G.A. Regulation of carotenoid biosynthetic genes expression and carotenoid accumulation in the green alga Haematococcus pluvialis under nutrient stress conditions. J. Exp. Bot. 2008, 59, 1409–1418. [Google Scholar] [CrossRef] [PubMed]
- Utermöhl, H. Neue Wege in der quantitativen Erfassung des Plankton. (Mit besonderer Berücksichtigung des Ultraplanktons). SIL Proc. 1922–2010 1931, 5, 567–596. [Google Scholar] [CrossRef]
- RStudio Team. RStudio: Inegrated Development Environment for R. RStudio, PBC, Boston, MA. 2021. Available online: http://www.rstudio (accessed on 18 December 2020).
- Becker, R.; Wilks, A.; Brownrigg, R. Maps: Draw Graphical Maps R Package Version 3.1.1. 2016. Available online: https://cran.r-project.org/package=maps (accessed on 18 December 2020).










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Quantity (%) | |
|---|---|---|
| PgZ1 | PgZ2 | |
| Acartia longiremis | 10.0 | - |
| Calanus finmarchicus | 2.6 | - |
| Calanus nauplius | - | 2.3 |
| Centopages typicus | 1.3 | 2.3 |
| Metridia longa | 0.3 | - |
| Microcalanus sp. | 2.4 | - |
| Oithona sp. | 65.1 | 53.5 |
| Pseudocalanus acuspes/sp. | 16.4 | 39.5 |
| Temora longa | 0.2 | - |
| Calanoida, uid juvenile | 1.0 | - |
| Paraeuchaeta norwegica | - | 2.3 |
| Bryozoa/cyphonautes | 0.8 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Osvik, R.D.; Ingebrigtsen, R.A.; Norrbin, M.F.; Andersen, J.H.; Eilertsen, H.C.; Hansen, E.H. Adding Zooplankton to the OSMAC Toolkit: Effect of Grazing Stress on the Metabolic Profile and Bioactivity of a Diatom. Mar. Drugs 2021, 19, 87. https://doi.org/10.3390/md19020087
Osvik RD, Ingebrigtsen RA, Norrbin MF, Andersen JH, Eilertsen HC, Hansen EH. Adding Zooplankton to the OSMAC Toolkit: Effect of Grazing Stress on the Metabolic Profile and Bioactivity of a Diatom. Marine Drugs. 2021; 19(2):87. https://doi.org/10.3390/md19020087
Chicago/Turabian StyleOsvik, Renate Døving, Richard Andre Ingebrigtsen, Maria Fredrika Norrbin, Jeanette Hammer Andersen, Hans Christian Eilertsen, and Espen Holst Hansen. 2021. "Adding Zooplankton to the OSMAC Toolkit: Effect of Grazing Stress on the Metabolic Profile and Bioactivity of a Diatom" Marine Drugs 19, no. 2: 87. https://doi.org/10.3390/md19020087
APA StyleOsvik, R. D., Ingebrigtsen, R. A., Norrbin, M. F., Andersen, J. H., Eilertsen, H. C., & Hansen, E. H. (2021). Adding Zooplankton to the OSMAC Toolkit: Effect of Grazing Stress on the Metabolic Profile and Bioactivity of a Diatom. Marine Drugs, 19(2), 87. https://doi.org/10.3390/md19020087