
From Natural Xanthones to Synthetic C-1 Aminated 3,4-Dioxygenated Xanthones as Optimized Antifouling Agents

 ,
,  ,
,  ,
,  , ,
, ,  ,
,  ,
,  and
and 
Abstract
:
1. Introduction
2. Results and Discussion
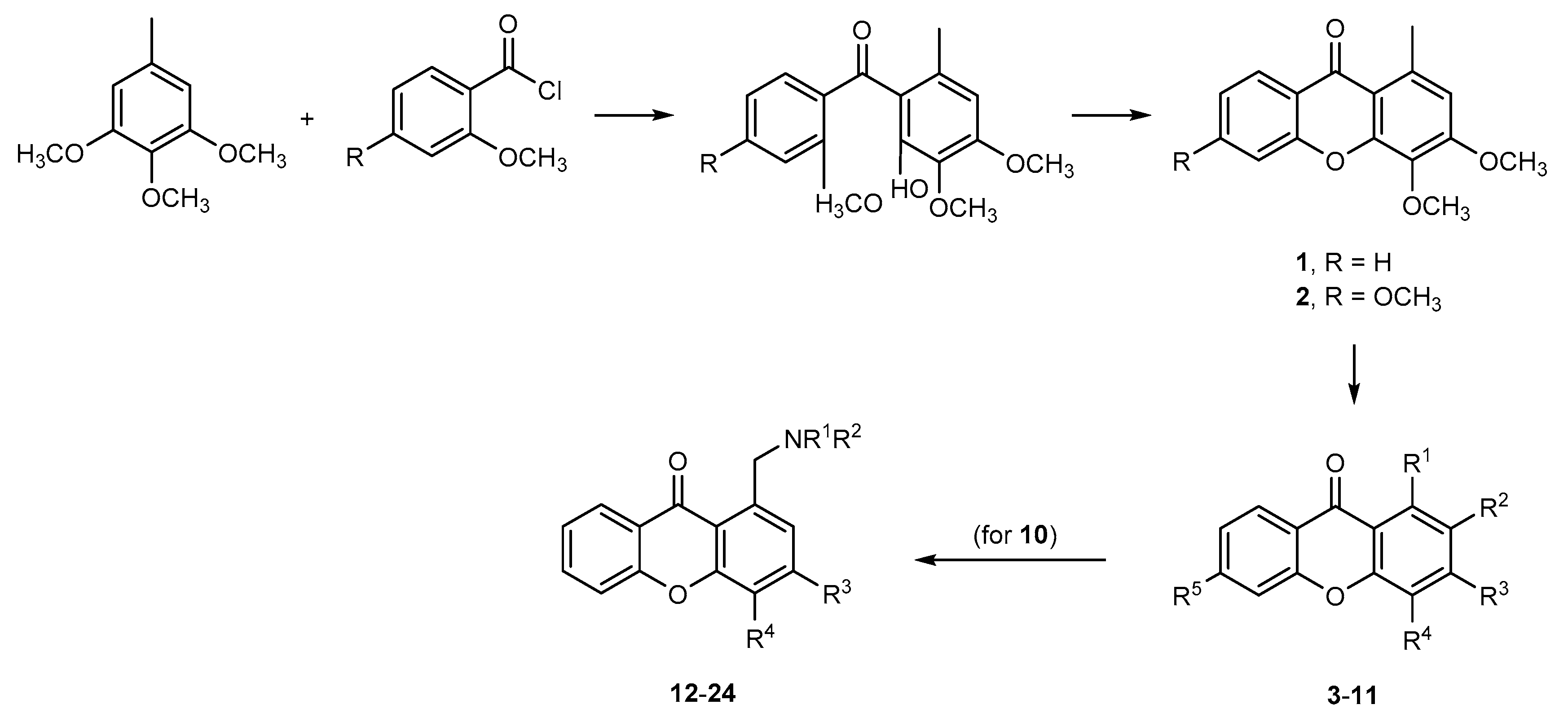
2.1. Synthesis
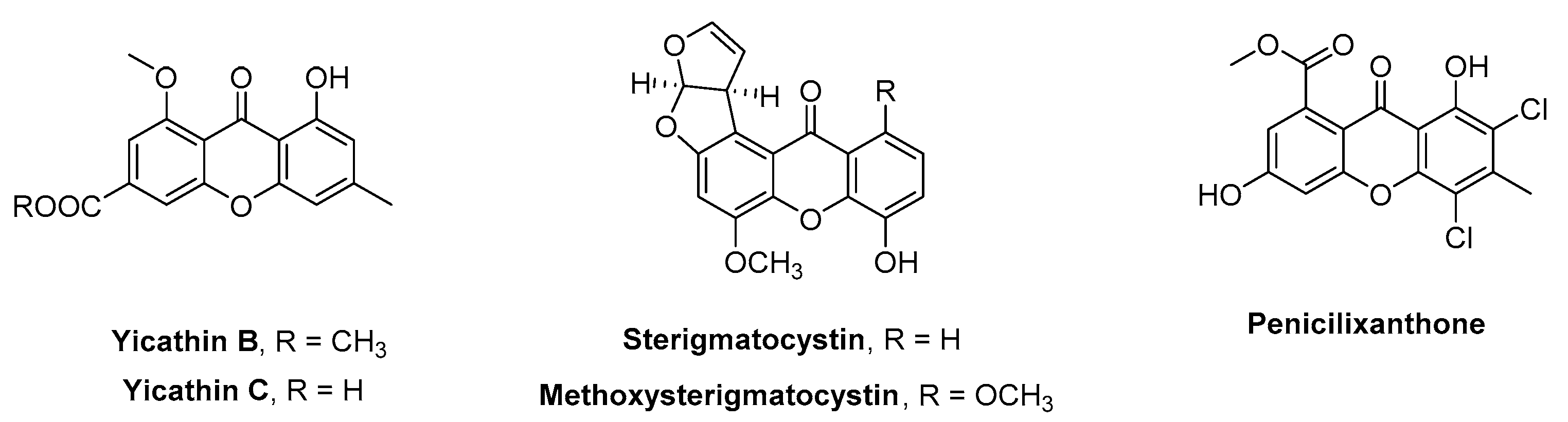
2.2. Structure-Anti-Settlement Activity Relationship of Xanthone Derivatives
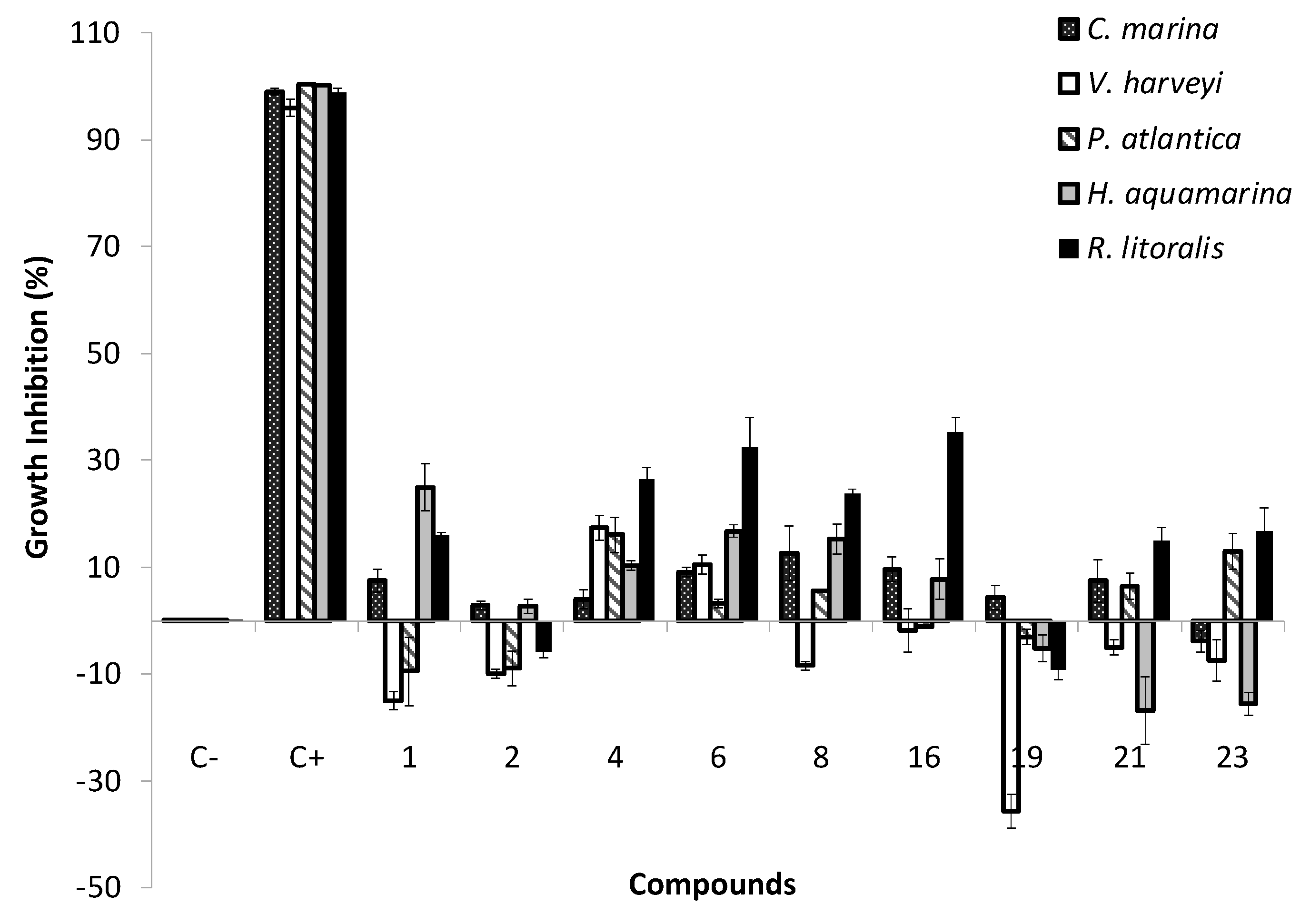
2.3. Antibacterial Activity
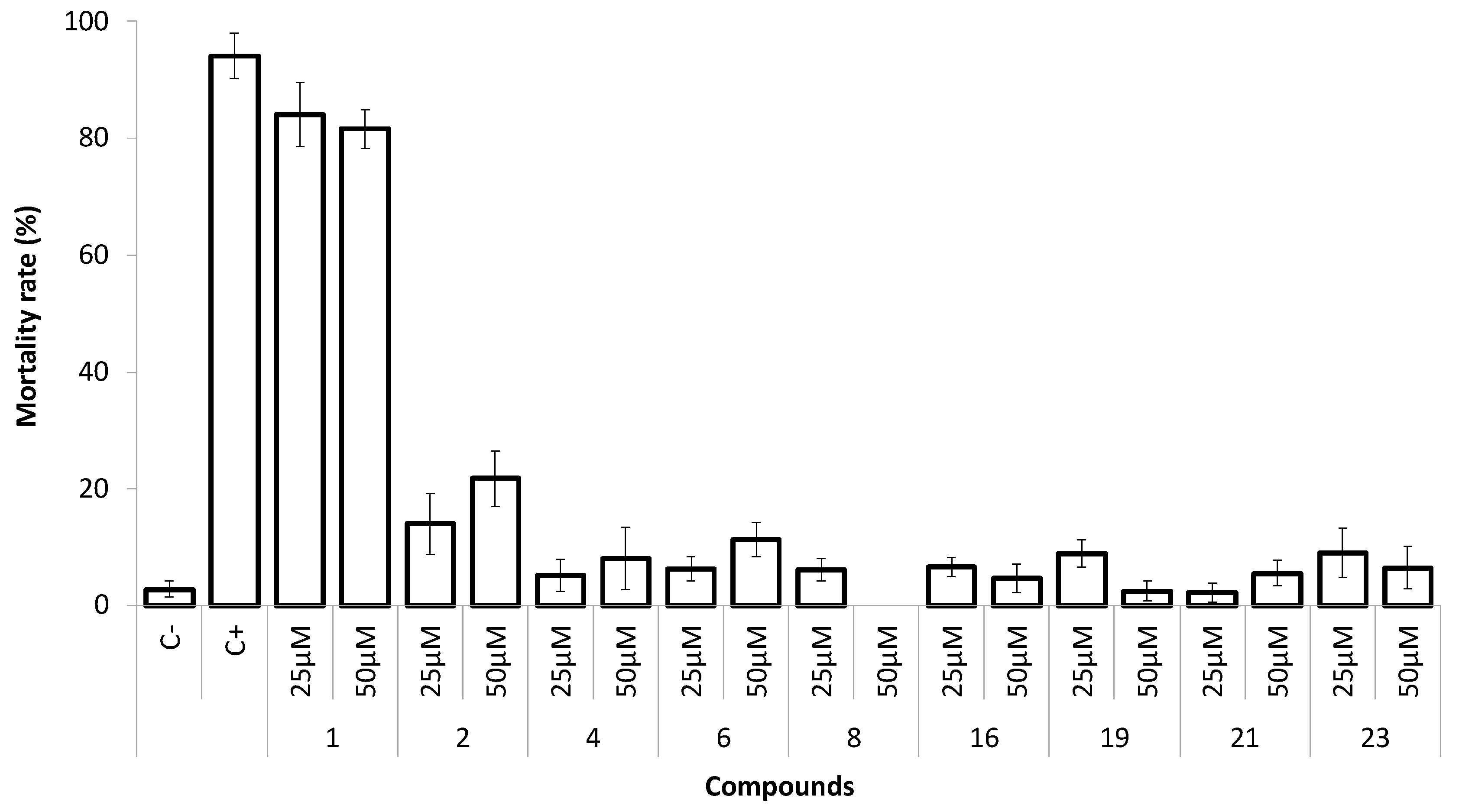
2.4. Marine Ecotoxicity
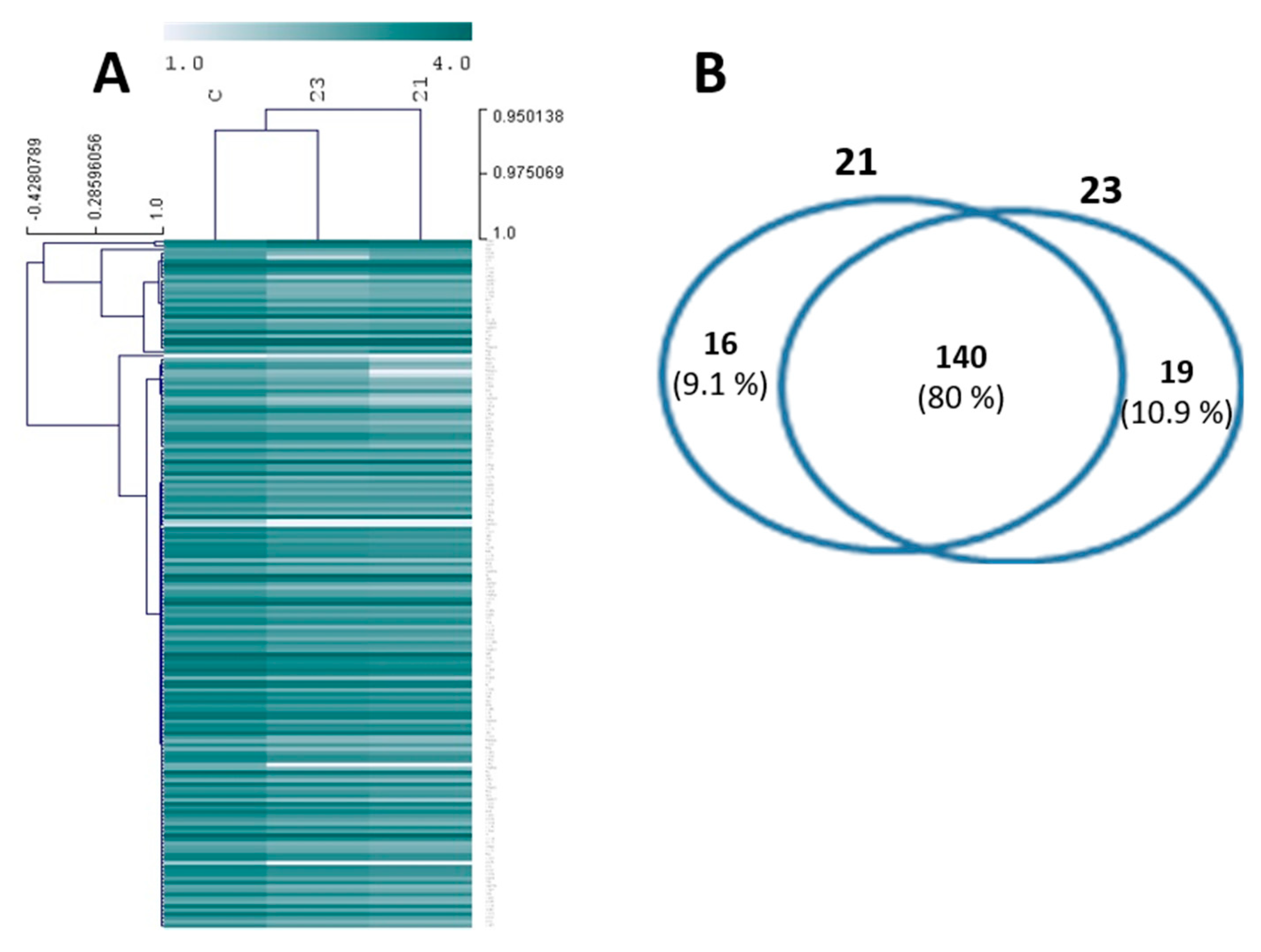
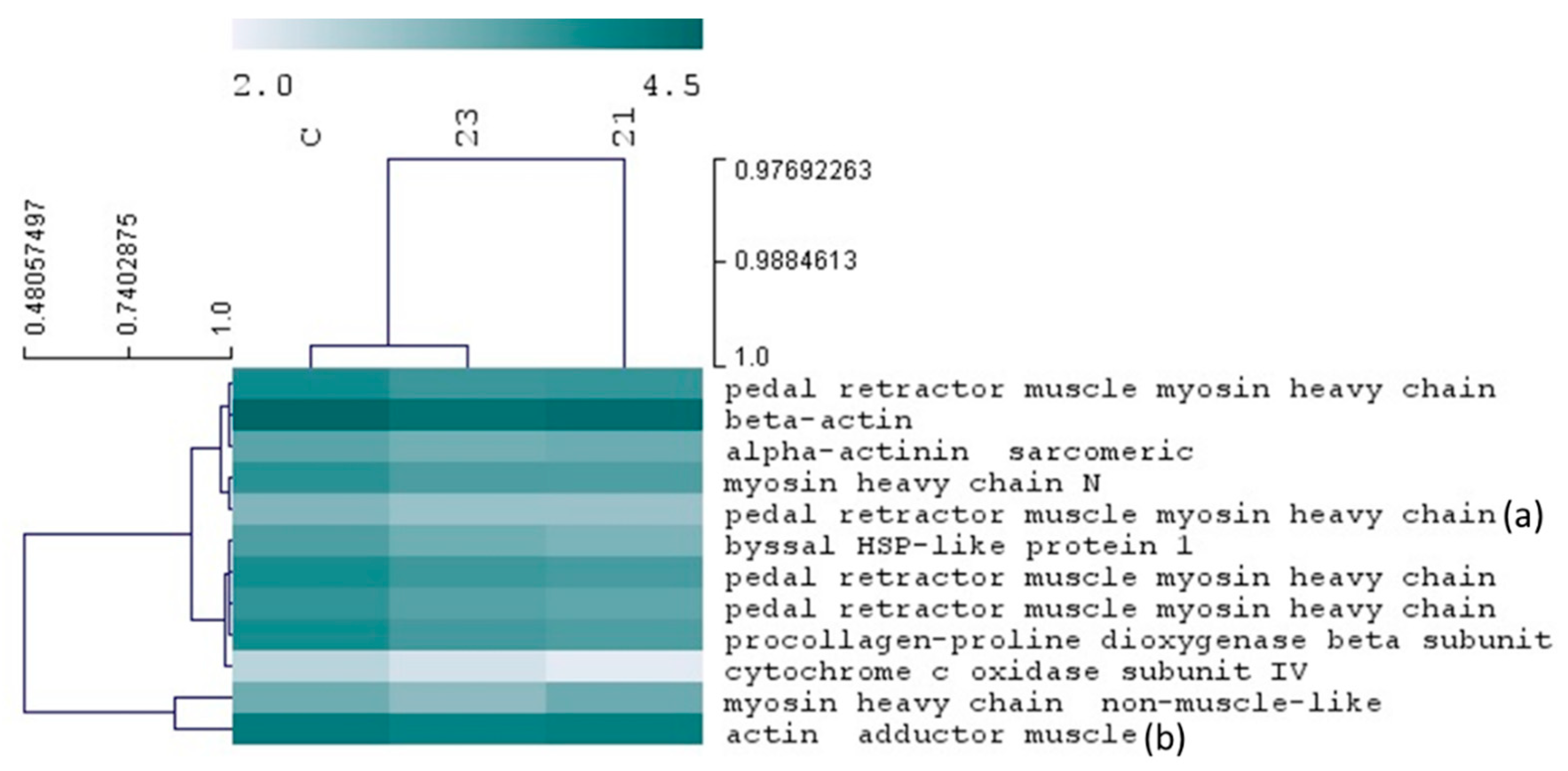
2.5. Insights on the Antifouling Mechanism of Action of the Hit Xanthones 21 and 23
Antifouling Targets by Proteomics
3. Materials and Methods
3.1. Chemicals
3.2. Antifouling Screening
3.2.1. Mytilus galloprovincialis Larvae Anti-Settlement Bioassays
3.2.2. Biofilm-Forming Marine Bacteria Growth Bioassays
3.3. Antifouling Molecular Targets Assessment in M. galloprovincialis Larvae by Differential Proteome Analysis
3.3.1. Sample Preparation
3.3.2. LC-MS/MS Analysis
3.3.3. Protein Identification and Quantification
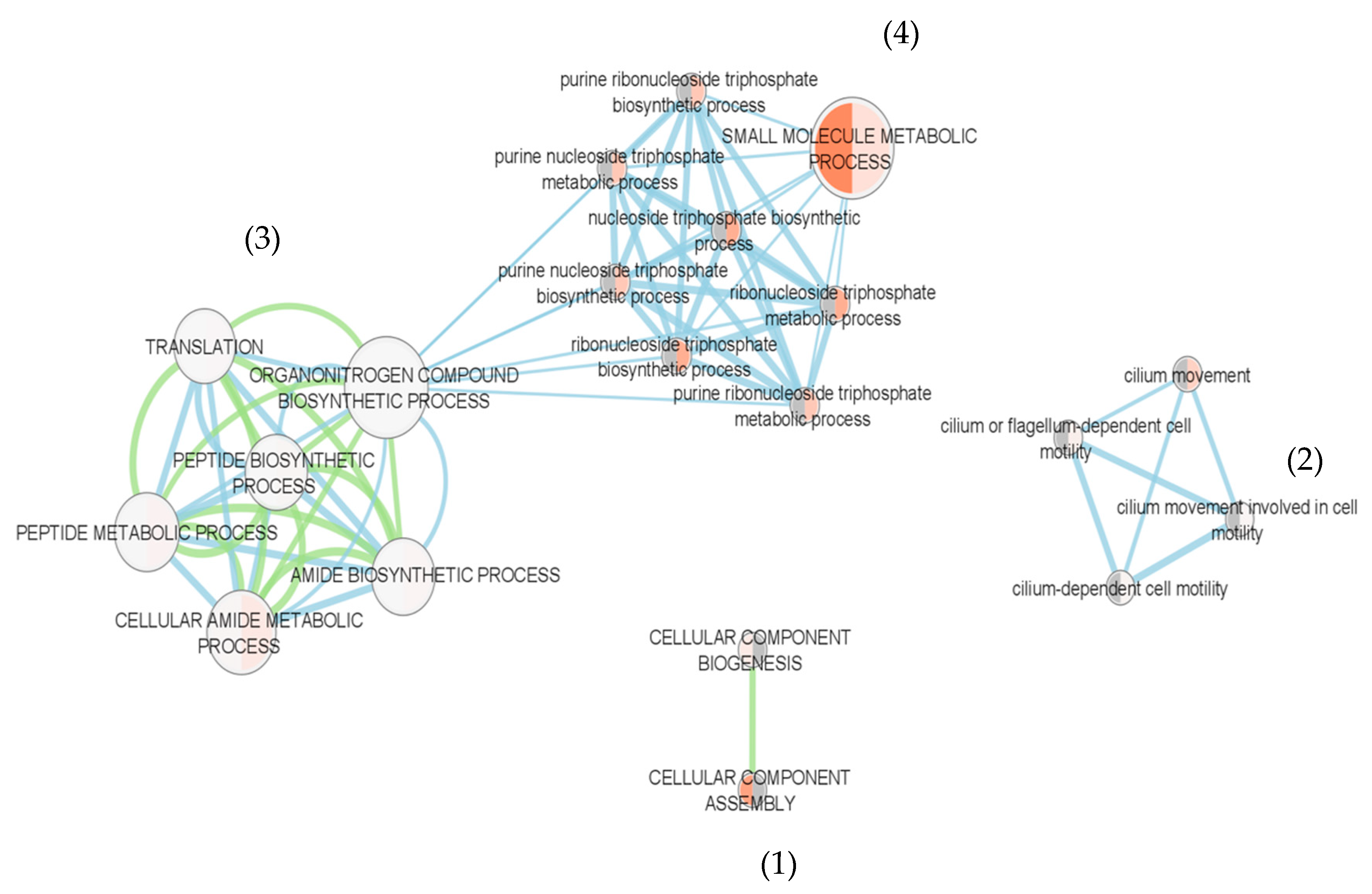
3.3.4. Protein Functional Analysis
3.4. Ecotoxicity Assessment
3.5. Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Vinagre, P.A.; Simas, T.; Cruz, E.; Pinori, E.; Svenson, J. Marine Biofouling: A European Database for the Marine Renewable Energy Sector. J. Mar. Sci. Eng. 2020, 8, 495. [Google Scholar] [CrossRef]
- Demirel, Y.K.; Uzun, D.; Zhang, Y.; Fang, H.-C.; Day, A.H.; Turan, O. Effect of barnacle fouling on ship resistance and powering. Biofouling 2017, 33, 819–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, J.R.; Vasconcelos, V. Natural antifouling compounds: Effectiveness in preventing invertebrate settlement and adhesion. Biotechnol. Adv. 2015, 33, 343–357. [Google Scholar] [CrossRef]
- Qian, P.-Y.; Li, Z.; Xu, Y.; Li, Y.; Fusetani, N. Mini-review: Marine natural products and their synthetic analogs as antifouling compounds: 2009–2014. Biofouling 2015, 31, 101–122. [Google Scholar] [CrossRef]
- Wang, K.-L.; Wu, Z.-H.; Wang, Y.; Wang, C.-Y.; Xu, Y. Mini-Review: Antifouling Natural Products from Marine Microorganisms and Their Synthetic Analogs. Mar. Drugs 2017, 15, 266. [Google Scholar] [CrossRef] [Green Version]
- Dahms, H.-U.; Ying, X.; Pfeiffer, C. Antifouling potential of cyanobacteria: A mini-review. Biofouling 2006, 22, 317–327. [Google Scholar] [CrossRef]
- Fusetani, N. Antifouling marine natural products. Nat. Prod. Rep. 2011, 28, 400–410. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Chen, J.; Hu, G.; Yu, J.; Zhu, X.; Lin, Y.; Chen, S.; Yuan, J. Statistical Research on the Bioactivity of New Marine Natural Products Discovered during the 28 Years from 1985 to 2012. Mar. Drugs 2015, 13, 202–221. [Google Scholar] [CrossRef]
- Liu, L.-L.; Wu, C.-H.; Qian, P.-Y. Marine natural products as antifouling molecules—A mini-review (2014–2020). Biofouling 2020, 36, 1210–1226. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.-R.; Miao, F.-P.; Zhang, J.; Wang, G.; Yin, X.-L.; Ji, N.-Y. Three new xanthone derivatives from an algicolous isolate of Aspergillus wentii. Magn. Reson. Chem. 2013, 51, 65–68. [Google Scholar] [CrossRef]
- Li, Y.-X.; Wu, H.-X.; Xu, Y.; Shao, C.-L.; Wang, C.-Y.; Qian, P.-Y. Antifouling Activity of Secondary Metabolites Isolated from Chinese Marine Organisms. Mar. Biotechnol. 2013, 15, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Nong, X.-H.; Zhang, X.-Y.; Xu, X.-Y.; Qi, S.-H. Antifouling Compounds from the Marine-Derived Fungus Aspergillus terreus SCSGAF0162. Nat. Prod. Commun. 2015, 10, 1934578X1501000659. [Google Scholar] [CrossRef] [Green Version]
- Almeida, J.R.; Palmeira, A.; Campos, A.; Cunha, I.; Freitas, M.; Felpeto, A.B.; Turkina, M.V.; Vasconcelos, V.; Pinto, M.; Correia-da-Silva, M.; et al. Structure-Antifouling Activity Relationship and Molecular Targets of Bio-Inspired(thio)xanthones. Biomolecules 2020, 10, 1126. [Google Scholar] [CrossRef]
- Vilas-Boas, C.; Neves, A.R.; Carvalhal, F.; Pereira, S.; Calhorda, M.J.; Vasconcelos, V.; Pinto, M.; Sousa, E.; Almeida, J.R.; Silva, E.R.; et al. Multidimensional characterization of a new antifouling xanthone:Structure-activity relationship, environmental compatibility, and immobilization in marine coatings. Ecotoxicol. Environ. Saf. 2021, 228, 112970. [Google Scholar] [CrossRef]
- Pinto, M.M.M.; Palmeira, A.; Fernandes, C.; Resende, D.I.S.P.; Sousa, E.; Cidade, H.; Tiritan, M.E.; Correia-da-Silva, M.; Cravo, S. From Natural Products to New Synthetic Small Molecules: A Journey through the World of Xanthones. Molecules 2021, 26, 431. [Google Scholar] [CrossRef]
- Resende, D.I.S.P.; Durães, F.; Maia, M.; Sousa, E.; Pinto, M.M.M. Recent advances in the synthesis of xanthones and azaxanthones. Org. Chem. Front. 2020, 7, 3027–3066. [Google Scholar] [CrossRef]
- Resende, D.I.S.P.; Pereira-Terra, P.; Inácio, Â.S.; Costa, P.M.d.; Pinto, E.; Sousa, E.; Pinto, M.M.M. Lichen Xanthones as Models for New Antifungal Agents. Molecules 2018, 23, 2617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Resende, D.I.S.P.; Pereira-Terra, P.; Moreira, J.; Freitas-Silva, J.; Lemos, A.; Gales, L.; Pinto, E.; de Sousa, M.E.; da Costa, P.M.; Pinto, M.M.M. Synthesis of a Small Library of Nature-Inspired Xanthones and Study of Their Antimicrobial Activity. Molecules 2020, 25, 2405. [Google Scholar] [CrossRef]
- Lemos, A.; Gomes, A.S.; Loureiro, J.B.; Brandão, P.; Palmeira, A.; Pinto, M.M.M.; Saraiva, L.; Sousa, M.E. Synthesis, Biological Evaluation, and In Silico Studies of Novel Aminated Xanthones as Potential p53-Activating Agents. Molecules 2019, 24, 1975. [Google Scholar] [CrossRef] [Green Version]
- Qian, P.Y.; Lau, S.C.K.; Dahms, H.U.; Dobretsov, S.; Harder, T. Marine Biofilms as Mediators of Colonization by Marine Macroorganisms: Implications for Antifouling and Aquaculture. Mar. Biotechnol. 2007, 9, 399–410. [Google Scholar] [CrossRef]
- Almeida, J.R.; Correia-da-Silva, M.; Sousa, E.; Antunes, J.; Pinto, M.; Vasconcelos, V.; Cunha, I. Antifouling potential of Nature-inspired sulfated compounds. Sci. Rep. 2017, 7, 42424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antunes, J.; Leão, P.; Vasconcelos, V. Marine biofilms: Diversity of communities and of chemical cues. Environ. Microbiol. Rep. 2019, 11, 287–305. [Google Scholar] [CrossRef] [PubMed]
- Antunes, J.; Pereira, S.; Ribeiro, T.; Plowman, J.E.; Thomas, A.; Clerens, S.; Campos, A.; Vasconcelos, V.; Almeida, J.R. A Multi-Bioassay Integrated Approach to Assess the Antifouling Potential of the Cyanobacterial Metabolites Portoamides. Mar. Drugs 2019, 17, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neves, A.R.; Almeida, J.R.; Carvalhal, F.; Câmara, A.; Pereira, S.; Antunes, J.; Vasconcelos, V.; Pinto, M.; Silva, E.R.; Sousa, E.; et al. Overcoming environmental problems of biocides: Synthetic bile acid derivatives as a sustainable alternative. Ecotoxicol. Environ. Saf. 2020, 187, 109812. [Google Scholar] [CrossRef] [PubMed]
- Dral, A.D.G. Movements and Co-ordination of the Latero-frontal Cilia of the Gill Filaments of Mytilus. Nature 1966, 210, 1170–1171. [Google Scholar] [CrossRef]
- Marumo, K.; Waite, J.H. Prolyl 4-hydroxylase in the foot of the marine mussel Mytilus edulis L.: Purification and characterization. J. Exp. Zool. 1987, 244, 365–374. [Google Scholar] [CrossRef] [PubMed]
- DeMartini, D.G.; Errico, J.M.; Sjoestroem, S.; Fenster, A.; Waite, J.H. A cohort of new adhesive proteins identified from transcriptomic analysis of mussel foot glands. J. R. Soc. Interface 2017, 14, 20170151. [Google Scholar] [CrossRef]
- Cruz, I.; Puthongking, P.; Cravo, S.; Palmeira, A.; Cidade, H.; Pinto, M.; Sousa, E. Xanthone and Flavone Derivatives as Dual Agents with Acetylcholinesterase Inhibition and Antioxidant Activity as Potential Anti-Alzheimer Agents. J. Chem. 2017, 2017, 8587260. [Google Scholar] [CrossRef]
- Almeida, J.R.; Moreira, J.; Pereira, D.; Pereira, S.; Antunes, J.; Palmeira, A.; Vasconcelos, V.; Pinto, M.; Correia-da-Silva, M.; Cidade, H. Potential of synthetic chalcone derivatives to prevent marine biofouling. Sci. Total Environ. 2018, 643, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Antunes, J.T.; Sousa, A.G.G.; Azevedo, J.; Rego, A.; Leão, P.N.; Vasconcelos, V. Distinct Temporal Succession of Bacterial Communities in Early Marine Biofilms in a Portuguese Atlantic Port. Front. Microbiol. 2020, 11, 1938. [Google Scholar] [CrossRef]
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods. 2009, 6, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. g: Profiler: A web server for functional enrichment analysis and conversions of gene lists (2019 update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, B.N.; Ferrigni, N.R.; Putnam, J.E.; Jacobsen, L.B.; Nichols, D.E.; McLaughlin, J.L. Brine Shrimp: A Convenient General Bioassay for Active Plant Constituents. Planta Med. 1982, 45, 31–34. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | EC50 (μM) | EC50 (μg.mL−1) | LC50 (μM) | LC50/EC50 |
|---|---|---|---|---|
| 1 | 11.81 (95% CI: 4.41–23.22) | 3.19 | >100 | >8.47 |
| 2 | 27.15 | 8.15 | >100 | >3.69 |
| 4 | 16.15 | 5.41 | >100 | >6.19 |
| 6 | 22.78 (95% CI: 14.50–38.06) | 9.75 | >100 | >4.39 |
| 8 | 21.83 (95% CI: 11.25–48.52) | 6.25 | >100 | >4.59 |
| 16 | 17.52 (95% CI: 11.47–26.87) | 6.98 | >100 | >5.71 |
| 19 | 53.85 (95% CI: 31.13–153.26) | 21.46 | >100 | >1.86 |
| 21 | 7.28 (95% CI: 0.91–15.83) | 3.03 | >100 | >13.74 |
| 23 | 3.57 (95% CI: 0.68–6.9) | 1.26 | >100 | >28.01 |
| ECONEA * | 4.01 (95% CI: 0.38–9.54) | 1.40 | 107.78 | 27 |
| Adjusted p Value | |||
|---|---|---|---|
| GO Name | GO id | 21 | 23 |
| organonitrogen compound biosynthetic process | GO:1901566 | 1.18 × 10−5 | 8.94 × 10−6 |
| translation | GO:0006412 | 7.41 × 10−4 | 8.80 × 10−5 |
| peptide biosynthetic process | GO:0043043 | 9.82 × 10−4 | 1.20 × 10−4 |
| amide biosynthetic process | GO:0043604 | 1.47 × 10−3 | 1.88 × 10−4 |
| peptide metabolic process | GO:0006518 | 1.85 × 10−3 | 2.42 × 10−4 |
| cellular amide metabolic process | GO:0043603 | 5.96 × 10−3 | 8.84 × 10−4 |
| small molecule metabolic process | GO:0044281 | 6.69 × 10−4 | 2.69 × 10−3 |
| cellular component biogenesis | GO:0044085 | 2.43 × 10−2 * | 3.94 × 10−3 |
| cilium movement involved in cell motility | GO:0060294 | 2.32 × 10−4 | 1.68 × 10−2 * |
| cilium-dependent cell motility | GO:0060285 | 3.62 × 10−4 | 2.30 × 10−2 * |
| cilium or flagellum-dependent cell motility | GO:0001539 | 7.76 × 10−4 | 3.94 × 10−2 * |
| purine ribonucleoside triphosphate metabolic process | GO:0009205 | 2.52 × 10−3 | 9.17 × 10−2 * |
| purine ribonucleoside triphosphate biosynthetic process | GO:0009206 | 2.52 × 10−3 | 9.17 × 10−2 * |
| purine nucleoside triphosphate metabolic process | GO:0009144 | 2.52 × 10−3 | 9.17 × 10−2 * |
| purine nucleoside triphosphate biosynthetic process | GO:0009145 | 2.52 × 10−3 | 9.17 × 10−2 * |
| cilium movement | GO:0003341 | 3.22 × 10−3 | 1.09 × 10−1 * |
| ribonucleoside triphosphate metabolic process | GO:0009199 | 4.06 × 10−3 | 1.29 × 10−1 * |
| nucleoside triphosphate biosynthetic process | GO:0009142 | 4.06 × 10−3 | 1.29 × 10−1 * |
| ribonucleoside triphosphate biosynthetic process | GO:0009201 | 4.06 × 10−3 | 1.29 × 10−1 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Resende, D.I.S.P.; Almeida, J.R.; Pereira, S.; Campos, A.; Lemos, A.; Plowman, J.E.; Thomas, A.; Clerens, S.; Vasconcelos, V.; Pinto, M.; et al. From Natural Xanthones to Synthetic C-1 Aminated 3,4-Dioxygenated Xanthones as Optimized Antifouling Agents. Mar. Drugs 2021, 19, 638. https://doi.org/10.3390/md19110638
Resende DISP, Almeida JR, Pereira S, Campos A, Lemos A, Plowman JE, Thomas A, Clerens S, Vasconcelos V, Pinto M, et al. From Natural Xanthones to Synthetic C-1 Aminated 3,4-Dioxygenated Xanthones as Optimized Antifouling Agents. Marine Drugs. 2021; 19(11):638. https://doi.org/10.3390/md19110638
Chicago/Turabian StyleResende, Diana I. S. P., Joana R. Almeida, Sandra Pereira, Alexandre Campos, Agostinho Lemos, Jeffrey E. Plowman, Ancy Thomas, Stefan Clerens, Vitor Vasconcelos, Madalena Pinto, and et al. 2021. "From Natural Xanthones to Synthetic C-1 Aminated 3,4-Dioxygenated Xanthones as Optimized Antifouling Agents" Marine Drugs 19, no. 11: 638. https://doi.org/10.3390/md19110638
APA StyleResende, D. I. S. P., Almeida, J. R., Pereira, S., Campos, A., Lemos, A., Plowman, J. E., Thomas, A., Clerens, S., Vasconcelos, V., Pinto, M., Correia-da-Silva, M., & Sousa, E. (2021). From Natural Xanthones to Synthetic C-1 Aminated 3,4-Dioxygenated Xanthones as Optimized Antifouling Agents. Marine Drugs, 19(11), 638. https://doi.org/10.3390/md19110638