Carrageenans as Broad-Spectrum Microbicides: Current Status and Challenges


Abstract
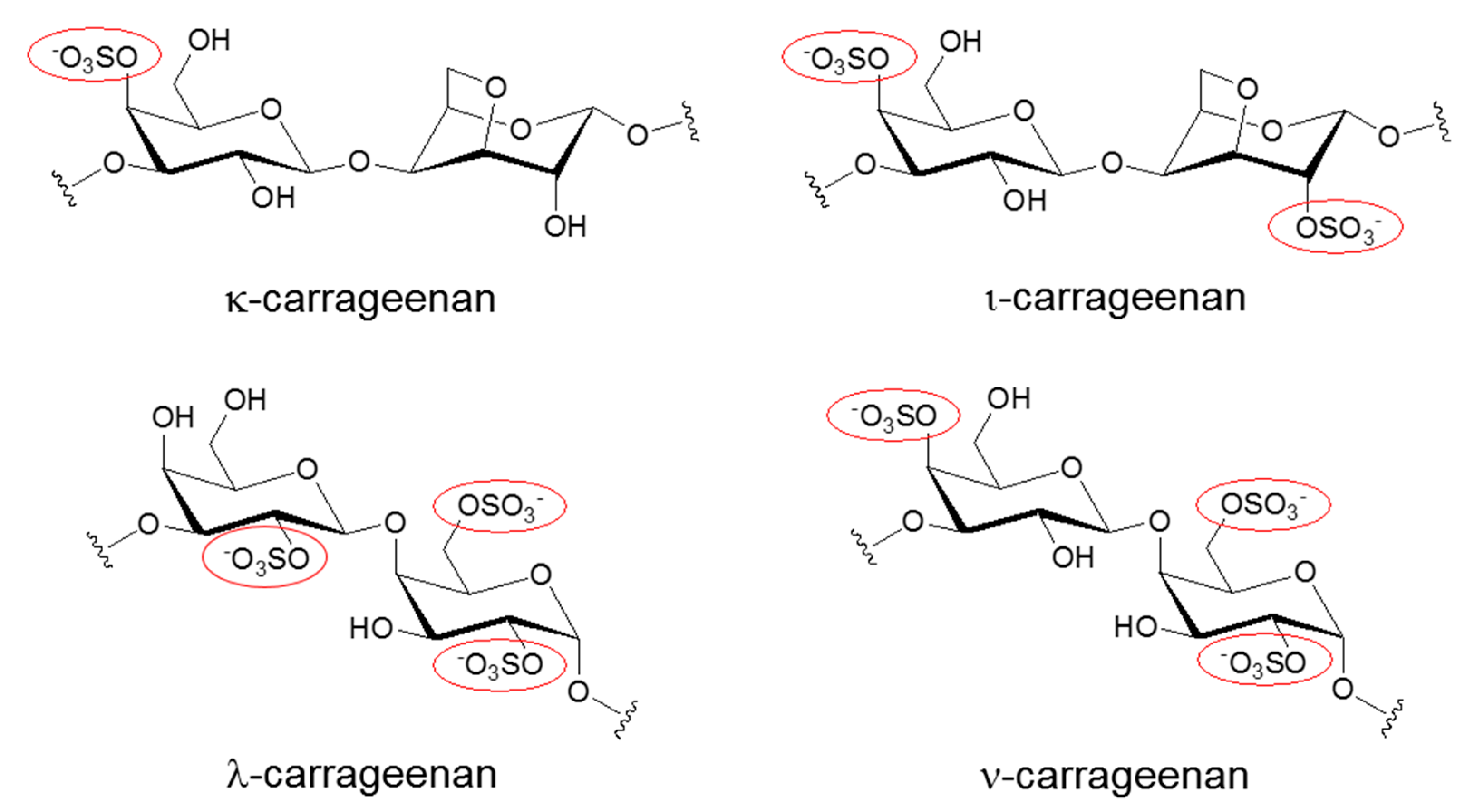
1. Introduction
In Vitro Toxicity and In Vivo Safety
2. Mechanisms of Antiviral Action
3. Human Immunodeficiency Virus (HIV)
3.1. In Vitro Anti-HIV Activity
3.2. Ex Vivo Anti-HIV Activity
3.3. In Vivo Anti-HIV Activity
3.4. Enhancement of HIV Infectivity
4. Herpesviruses
4.1. In Vitro Anti-Herpesviruses Activity
4.2. In Vivo Anti-Herpesviruses Activity
4.3. CG-Resistant HSV Variants
5. Respiratory Viruses
5.1. In Vitro Anti-Respiratory Viruses Activity
5.2. In Vivo Anti-Respiratory Viruses Activity
6. Human Papillomavirus (HPV)
6.1. In Vitro Anti-HPV Activity
6.2. In Vivo Anti-HPV Activity
7. Dengue Virus (DENV)
8. Other Viruses
9. Conclusions and Future Direction
Funding
Conflicts of Interest
References
- Ahmadi, A.; Zorofchian Moghadamtousi, S.; Abubakar, S.; Zandi, K. Antiviral Potential of Algae Polysaccharides Isolated from Marine Sources: A Review. Biomed Res. Int. 2015, 2015, 825203. [Google Scholar] [CrossRef]
- Damonte, E.B.; Matulewicz, M.C.; Cerezo, A.S. Sulfated seaweed polysaccharides as antiviral agents. Curr. Med. Chem. 2004, 11, 2399–2419. [Google Scholar] [CrossRef]
- Witvrouw, M.; De Clercq, E. Sulfated polysaccharides extracted from sea algae as potential antiviral drugs. Gen. Pharmacol. 1997, 29, 497–511. [Google Scholar] [CrossRef]
- Pirrone, V.; Wigdahl, B.; Krebs, F.C. The rise and fall of polyanionic inhibitors of the human immunodeficiency virus type 1. Antivir. Res. 2011, 90, 168–182. [Google Scholar] [CrossRef] [PubMed]
- Tobacman, J.K. Review of harmful gastrointestinal effects of carrageenan in animal experiments. Environ. Health Perspect. 2001, 109, 983–994. [Google Scholar] [CrossRef] [PubMed]
- Van de Wijgert, J.H.; Shattock, R.J. Vaginal microbicides: Moving ahead after an unexpected setback. AIDS 2007, 21, 2369–2376. [Google Scholar] [CrossRef] [PubMed]
- Jean-Pierre, N.; Barnable, P.; Kizima, L.; Rodriguez, A.; Seidor, S.; Cooney, M.L.; Clark, M.R.; Doncel, G.F.; Robbiani, M.; Zydowsky, T.M.; et al. Multipurpose Prevention Approaches with Antiretroviral-Based Formulations. Antimicrob. Agents Chemother. 2016, 60, 1141–1144. [Google Scholar] [CrossRef][Green Version]
- Kenney, J.; Rodriguez, A.; Kizima, L.; Seidor, S.; Menon, R.; Jean-Pierre, N.; Pugach, P.; Levendosky, K.; Derby, N.; Gettie, A.; et al. A modified zinc acetate gel, a potential nonantiretroviral microbicide, is safe and effective against simian-human immunodeficiency virus and herpes simplex virus 2 infection in vivo. Antimicrob. Agents Chemother. 2013, 57, 4001–4009. [Google Scholar] [CrossRef]
- Villegas, G.; Calenda, G.; Zhang, S.; Mizenina, O.; Kleinbeck, K.; Cooney, M.L.; Hoesley, C.J.; Creasy, G.W.; Friedland, B.; Fernandez-Romero, J.A.; et al. In Vitro Exposure to PC-1005 and Cervicovaginal Lavage Fluid from Women Vaginally Administered PC-1005 Inhibits HIV-1 and HSV-2 Infection in Human Cervical Mucosa. Antimicrob. Agents Chemother. 2016, 60, 5459–5466. [Google Scholar] [CrossRef]
- Friend, D.R.; Clark, J.T.; Kiser, P.F.; Clark, M.R. Multipurpose prevention technologies: Products in development. Antivir. Res. 2013, 100, S39–S47. [Google Scholar] [CrossRef]
- Skoler-Karpoff, S.; Ramjee, G.; Ahmed, K.; Altini, L.; Plagianos, M.G.; Friedland, B.; Govender, S.; De Kock, A.; Cassim, N.; Palanee, T.; et al. Efficacy of Carraguard for prevention of HIV infection in women in South Africa: A randomised, double-blind, placebo-controlled trial. Lancet 2008, 372, 1977–1987. [Google Scholar] [CrossRef]
- Turville, S.G.; Aravantinou, M.; Miller, T.; Kenney, J.; Teitelbaum, A.; Hu, L.; Chudolij, A.; Zydowsky, T.M.; Piatak, M., Jr.; Bess, J.W., Jr.; et al. Efficacy of Carraguard-based microbicides in vivo despite variable in vitro activity. PLoS ONE 2008, 3, e3162. [Google Scholar] [CrossRef]
- Pirrone, V.; Passic, S.; Wigdahl, B.; Krebs, F.C. Application and removal of polyanionic microbicide compounds enhances subsequent infection by HIV-1. Virol. J. 2012, 9, 33. [Google Scholar] [CrossRef]
- Tan, S.; Lu, L.; Li, L.; Liu, J.; Oksov, Y.; Lu, H.; Jiang, S.; Liu, S. Polyanionic candidate microbicides accelerate the formation of semen-derived amyloid fibrils to enhance HIV-1 infection. PLoS ONE 2013, 8, e59777. [Google Scholar] [CrossRef] [PubMed]
- Tao, W.; Richards, C.; Hamer, D. Enhancement of HIV infection by cellulose sulfate. AIDS Res. Hum. Retrovir. 2008, 24, 925–929. [Google Scholar] [CrossRef] [PubMed]
- Acosta, E.G.; Piccini, L.E.; Talarico, L.B.; Castilla, V.; Damonte, E.B. Changes in antiviral susceptibility to entry inhibitors and endocytic uptake of dengue-2 virus serially passaged in Vero or C6/36 cells. Virus Res. 2014, 184, 39–43. [Google Scholar] [CrossRef]
- Andrei, G.; De Clercq, E. Inhibitory effect of selected antiviral compounds on arenavirus replication in vitro. Antivir. Res. 1990, 14, 287–299. [Google Scholar] [CrossRef]
- Baba, M.; Snoeck, R.; Pauwels, R.; de Clercq, E. Sulfated polysaccharides are potent and selective inhibitors of various enveloped viruses, including herpes simplex virus, cytomegalovirus, vesicular stomatitis virus, and human immunodeficiency virus. Antimicrob. Agents Chemother. 1988, 32, 1742–1745. [Google Scholar] [CrossRef]
- Briolant, S.; Garin, D.; Scaramozzino, N.; Jouan, A.; Crance, J.M. In vitro inhibition of Chikungunya and Semliki Forest viruses replication by antiviral compounds: Synergistic effect of interferon-alpha and ribavirin combination. Antivir. Res. 2004, 61, 111–117. [Google Scholar] [CrossRef]
- Buck, C.B.; Thompson, C.D.; Roberts, J.N.; Muller, M.; Lowy, D.R.; Schiller, J.T. Carrageenan is a potent inhibitor of papillomavirus infection. PLoS Pathog. 2006, 2, e69. [Google Scholar] [CrossRef]
- Caceres, P.J.; Carlucci, M.J.; Damonte, E.B.; Matsuhiro, B.; Zuniga, E.A. Carrageenans from chilean samples of Stenogramme interrupta (Phyllophoraceae): Structural analysis and biological activity. Phytochemistry 2000, 53, 81–86. [Google Scholar] [CrossRef]
- Carlucci, M.J.; Ciancia, M.; Matulewicz, M.C.; Cerezo, A.S.; Damonte, E.B. Antiherpetic activity and mode of action of natural carrageenans of diverse structural types. Antivir. Res. 1999, 43, 93–102. [Google Scholar] [CrossRef]
- Carlucci, M.J.; Pujol, C.A.; Ciancia, M.; Noseda, M.D.; Matulewicz, M.C.; Damonte, E.B.; Cerezo, A.S. Antiherpetic and anticoagulant properties of carrageenans from the red seaweed Gigartina skottsbergii and their cyclized derivatives: Correlation between structure and biological activity. Int. J. Biol. Macromol. 1997, 20, 97–105. [Google Scholar] [CrossRef]
- Carlucci, M.J.; Scolaro, L.A.; Damonte, E.B. Inhibitory action of natural carrageenans on Herpes simplex virus infection of mouse astrocytes. Chemotherapy 1999, 45, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Cosenza, V.A.; Navarro, D.A.; Pujol, C.A.; Damonte, E.B.; Stortz, C.A. Partial and total C-6 oxidation of gelling carrageenans. Modulation of the antiviral activity with the anionic character. Carbohydr. Polym. 2015, 128, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Diogo, J.V.; Novo, S.G.; Gonzalez, M.J.; Ciancia, M.; Bratanich, A.C. Antiviral activity of lambda-carrageenan prepared from red seaweed (Gigartina skottsbergii) against BoHV-1 and SuHV-1. Res. Vet. Sci. 2015, 98, 142–144. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Villalon, D.; Gil-Fernandez, C. Antiviral activity of sulfated polysaccharides against African swine fever virus. Antivir. Res. 1991, 15, 139–148. [Google Scholar] [CrossRef]
- Girond, S.; Crance, J.M.; Van Cuyck-Gandre, H.; Renaudet, J.; Deloince, R. Antiviral activity of carrageenan on hepatitis A virus replication in cell culture. Res. Virol. 1991, 142, 261–270. [Google Scholar] [CrossRef]
- Huang, S.H.; Lin, Y.S.; Wu, C.W.; Wu, C.J. Assessment of the inhibition of Dengue virus infection by carrageenan via real-time monitoring of cellular oxygen consumption rates within a microfluidic device. Biomicrofluidics 2014, 8, 024110. [Google Scholar] [CrossRef]
- Luo, Z.; Tian, D.; Zhou, M.; Xiao, W.; Zhang, Y.; Li, M.; Sui, B.; Wang, W.; Guan, H.; Chen, H.; et al. lambda-Carrageenan P32 Is a Potent Inhibitor of Rabies Virus Infection. PLoS ONE 2015, 10, e0140586. [Google Scholar] [CrossRef]
- Marchetti, M.; Pisani, S.; Pietropaolo, V.; Seganti, L.; Nicoletti, R.; Orsi, N. Inhibition of herpes simplex virus infection by negatively charged and neutral carbohydrate polymers. J. Chemother. 1995, 7, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Penuela, A.; Robledo, D.; Bourgougnon, N.; Bedoux, G.; Hernandez-Nunez, E.; Freile-Pelegrin, Y. Environmentally Friendly Valorization of Solieria filiformis (Gigartinales, Rhodophyta) from IMTA Using a Biorefinery Concept. Mar. Drugs 2018, 16, 487. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.; Kleinbeck, K.; Mizenina, O.; Kizima, L.; Levendosky, K.; Jean-Pierre, N.; Villegas, G.; Ford, B.E.; Cooney, M.L.; Teleshova, N.; et al. In vitro and in vivo evaluation of two carrageenan-based formulations to prevent HPV acquisition. Antivir. Res. 2014, 108, 88–93. [Google Scholar] [CrossRef]
- Shao, Q.; Guo, Q.; Xu, W.; Li, Z.; Zhao, T. Specific Inhibitory Effect of kappa-Carrageenan Polysaccharide on Swine Pandemic 2009 H1N1 Influenza Virus. PLoS ONE 2015, 10, e0126577. [Google Scholar]
- Sola, A.; Rodriguez, S.; Gil-Fernandez, C.; Alarcon, B.; Gonzalez, M.E.; Carrasco, L. New agents active against African swine fever virus. Antimicrob. Agents Chemother. 1986, 29, 284–288. [Google Scholar] [CrossRef]
- Talarico, L.B.; Damonte, E.B. Interference in dengue virus adsorption and uncoating by carrageenans. Virology 2007, 363, 473–485. [Google Scholar] [CrossRef]
- Talarico, L.B.; Damonte, E.B. Characterization of in vitro dengue virus resistance to carrageenan. J. Med. Virol. 2016, 88, 1120–1129. [Google Scholar] [CrossRef]
- Talarico, L.B.; Noseda, M.D.; Ducatti, D.R.; Duarte, M.E.; Damonte, E.B. Differential inhibition of dengue virus infection in mammalian and mosquito cells by iota-carrageenan. J. Gen. Virol. 2011, 92 Pt 6, 1332–1342. [Google Scholar] [CrossRef]
- Talarico, L.B.; Pujol, C.A.; Zibetti, R.G.; Faria, P.C.; Noseda, M.D.; Duarte, M.E.; Damonte, E.B. The antiviral activity of sulfated polysaccharides against dengue virus is dependent on virus serotype and host cell. Antivir. Res. 2005, 66, 103–110. [Google Scholar] [CrossRef]
- Talarico, L.B.; Zibetti, R.G.; Faria, P.C.; Scolaro, L.A.; Duarte, M.E.; Noseda, M.D.; Pujol, C.A.; Damonte, E.B. Anti-herpes simplex virus activity of sulfated galactans from the red seaweeds Gymnogongrus griffithsiae and Cryptonemia crenulata. Int. J. Biol. Macromol. 2004, 34, 63–71. [Google Scholar] [CrossRef]
- Vlieghe, P.; Clerc, T.; Pannecouque, C.; Witvrouw, M.; De Clercq, E.; Salles, J.P.; Kraus, J.L. Synthesis of new covalently bound kappa-carrageenan-AZT conjugates with improved anti-HIV activities. J. Med. Chem. 2002, 45, 1275–1283. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhang, P.; Hao, C.; Zhang, X.E.; Cui, Z.Q.; Guan, H.S. In vitro inhibitory effect of carrageenan oligosaccharide on influenza A H1N1 virus. Antivir. Res. 2011, 92, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Abu-Galiyun, E.; Huleihel, M.; Levy-Ontman, O. Antiviral bioactivity of renewable polysaccharides against Varicella Zoster. Cell Cycle 2019, 18, 3540–3549. [Google Scholar] [CrossRef] [PubMed]
- Elias, C.J.; Coggins, C.; Alvarez, F.; Brache, V.; Fraser, I.S.; Lacarra, M.; Lahteenmaki, P.; Massai, R.; Mishell, D.R., Jr.; Phillips, D.M.; et al. Colposcopic evaluation of a vaginal gel formulation of iota-carrageenan. Contraception 1997, 56, 387–389. [Google Scholar] [CrossRef]
- Kilmarx, P.H.; Blanchard, K.; Chaikummao, S.; Friedland, B.A.; Srivirojana, N.; Connolly, C.; Witwatwongwana, P.; Supawitkul, S.; Mock, P.A.; Chaowanachan, T.; et al. A randomized, placebo-controlled trial to assess the safety and acceptability of use of carraguard vaginal gel by heterosexual couples in Thailand. Sex. Transm. Dis. 2008, 35, 226–232. [Google Scholar] [CrossRef]
- Van de Wijgert, J.H.; Braunstein, S.L.; Morar, N.S.; Jones, H.E.; Madurai, L.; Strickfaden, T.T.; Moodley, M.; Aboobaker, J.; Ndlovu, G.; Ferguson, T.M.; et al. Carraguard Vaginal Gel Safety in HIV-Positive Women and Men in South Africa. J. Acquir. Immune Defic. Syndr. 2007, 46, 538–546. [Google Scholar] [CrossRef]
- Bollen, L.J.; Blanchard, K.; Kilmarx, P.H.; Chaikummao, S.; Connolly, C.; Wasinrapee, P.; Srivirojana, N.; Achalapong, J.; Tappero, J.W.; McNicholl, J.M. No increase in cervicovaginal proinflammatory cytokines after Carraguard use in a placebo-controlled randomized clinical trial. J. Acquir. Immune Defic. Syndr. 2008, 47, 253–257. [Google Scholar] [CrossRef]
- Whitehead, S.J.; Kilmarx, P.H.; Blanchard, K.; Manopaiboon, C.; Chaikummao, S.; Friedland, B.; Achalapong, J.; Wankrairoj, M.; Mock, P.; Thanprasertsuk, S.; et al. Acceptability of Carraguard vaginal gel use among Thai couples. AIDS 2006, 20, 2141–2148. [Google Scholar] [CrossRef]
- Friedland, B.A.; Hoesley, C.J.; Plagianos, M.; Hoskin, E.; Zhang, S.; Teleshova, N.; Alami, M.; Novak, L.; Kleinbeck, K.R.; Katzen, L.L.; et al. First-in-Human Trial of MIV-150 and Zinc Acetate Coformulated in a Carrageenan Gel: Safety, Pharmacokinetics, Acceptability, Adherence, and Pharmacodynamics. J. Acquir. Immune Defic. Syndr. 2016, 73, 489–496. [Google Scholar] [CrossRef]
- McLean, C.A.; van de Wijgert, J.H.; Jones, H.E.; Karon, J.M.; McNicoll, J.M.; Whitehead, S.J.; Braunstein, S.; Achalapong, J.; Chaikummao, S.; Tappero, J.W.; et al. HIV genital shedding and safety of Carraguard use by HIV-infected women: A crossover trial in Thailand. AIDS 2010, 24, 717–722. [Google Scholar] [CrossRef]
- Kilmarx, P.H.; van de Wijgert, J.H.; Chaikummao, S.; Jones, H.E.; Limpakarnjanarat, K.; Friedland, B.A.; Karon, J.M.; Manopaiboon, C.; Srivirojana, N.; Yanpaisarn, S.; et al. Safety and acceptability of the candidate microbicide Carraguard in Thai Women: Findings from a Phase II Clinical Trial. J. Acquir. Immune Defic. Syndr. 2006, 43, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Ramjee, G.; Morar, N.S.; Braunstein, S.; Friedland, B.; Jones, H.; van de Wijgert, J. Acceptability of Carraguard, a candidate microbicide and methyl cellulose placebo vaginal gels among HIV-positive women and men in Durban, South Africa. AIDS Res. Ther. 2007, 4, 20. [Google Scholar] [CrossRef] [PubMed]
- Carraguard Phase II South Africa Study Team. Expanded safety and acceptability of the candidate vaginal microbicide Carraguard(R) in South Africa. Contraception 2010, 82, 563–571. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, S.J.; McLean, C.; Chaikummao, S.; Braunstein, S.; Utaivoravit, W.; van de Wijgert, J.H.; Mock, P.A.; Siraprapasiri, T.; Friedland, B.A.; Kilmarx, P.H.; et al. Acceptability of Carraguard vaginal microbicide gel among HIV-infected women in Chiang Rai, Thailand. PLoS ONE 2011, 6, e14831. [Google Scholar] [CrossRef]
- Pearce-Pratt, R.; Phillips, D.M. Sulfated polysaccharides inhibit lymphocyte-to-epithelial transmission of human immunodeficiency virus-1. Biol. Reprod. 1996, 54, 173–182. [Google Scholar] [CrossRef]
- Lynch, G.; Low, L.; Li, S.; Sloane, A.; Adams, S.; Parish, C.; Kemp, B.; Cunningham, A.L. Sulfated polyanions prevent HIV infection of lymphocytes by disruption of the CD4-gp120 interaction, but do not inhibit monocyte infection. J. Leukoc. Biol. 1994, 56, 266–272. [Google Scholar] [CrossRef]
- Schaeffer, D.J.; Krylov, V.S. Anti-HIV activity of extracts and compounds from algae and cyanobacteria. Ecotoxicol. Environ. Saf. 2000, 45, 208–227. [Google Scholar] [CrossRef]
- Ghosh, T.; Chattopadhyay, K.; Marschall, M.; Karmakar, P.; Mandal, P.; Ray, B. Focus on antivirally active sulfated polysaccharides: From structure-activity analysis to clinical evaluation. Glycobiology 2009, 19, 2–15. [Google Scholar] [CrossRef]
- Wang, W.; Wang, S.X.; Guan, H.S. The antiviral activities and mechanisms of marine polysaccharides: An overview. Mar. Drugs 2012, 10, 2795–2816. [Google Scholar] [CrossRef]
- Arad, S.M.; Levy-Ontman, O. Red microalgal cell-wall polysaccharides: Biotechnological aspects. Curr. Opin. Biotechnol. 2010, 21, 358–364. [Google Scholar] [CrossRef]
- Nakashima, H.; Kido, Y.; Kobayashi, N.; Motoki, Y.; Neushul, M.; Yamamoto, N. Purification and characterization of an avian myeloblastosis and human immunodeficiency virus reverse transcriptase inhibitor, sulfated polysaccharides extracted from sea algae. Antimicrob. Agents Chemother. 1987, 31, 1524–1528. [Google Scholar] [CrossRef] [PubMed]
- Hamasuna, R.; Eizuru, Y.; Minamishima, Y. Inhibition by iota-carrageenan of the spread of murine cytomegalovirus from the peritoneal cavity to the blood plasma. J. Gen. Virol. 1994, 75 Pt 1, 111–116. [Google Scholar] [CrossRef]
- Hamasuna, R.E.; Shishime, Y.; Minamishima, Y. Protective effect of carrageenan against murine cytomegalovirus infection in mice. Antimicrob. Agents Chemother. 1993, 4, 353–360. [Google Scholar] [CrossRef]
- Abdool Karim, Q.; Abdool Karim, S.S.; Frohlich, J.A.; Grobler, A.C.; Baxter, C.; Mansoor, L.E.; Kharsany, A.B.; Sibeko, S.; Mlisana, K.P.; Omar, Z.; et al. Effectiveness and safety of tenofovir gel, an antiretroviral microbicide, for the prevention of HIV infection in women. Science 2010, 329, 1168–1174. [Google Scholar] [CrossRef]
- Berry, M.; Wirtz, A.L.; Janayeva, A.; Ragoza, V.; Terlikbayeva, A.; Amirov, B.; Baral, S.; Beyrer, C. Risk factors for HIV and unprotected anal intercourse among men who have sex with men (MSM) in Almaty, Kazakhstan. PLoS ONE 2012, 7, e43071. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Ogamo, A.; Saito, T.; Watanabe, J.; Uchiyama, H.; Nakagawa, Y. Preparation and anti-HIV activity of low molecular-weight carrageenans and their sulfated derivatives. Carbohydr. Polym. 1997, 32, 51–55. [Google Scholar] [CrossRef]
- Fernandez-Romero, J.A.; Thorn, M.; Turville, S.G.; Titchen, K.; Sudol, K.; Li, J.; Miller, T.; Robbiani, M.; Maguire, R.A.; Buckheit, R.W., Jr.; et al. Carrageenan/MIV-150 (PC-815), a combination microbicide. Sex. Transm. Dis. 2007, 34, 9–14. [Google Scholar] [CrossRef]
- Ding, J.; Rapista, A.; Teleshova, N.; Lu, W.; Klotman, M.E.; Chang, T.L. Mucosal human defensins 5 and 6 antagonize the anti-HIV activity of candidate polyanion microbicides. J. Innate Immun. 2011, 3, 208–212. [Google Scholar] [CrossRef]
- Barnable, P.; Calenda, G.; Ouattara, L.; Gettie, A.; Blanchard, J.; Jean-Pierre, N.; Kizima, L.; Rodriguez, A.; Abraham, C.; Menon, R.; et al. A MIV-150/zinc acetate gel inhibits SHIV-RT infection in macaque vaginal explants. PLoS ONE 2014, 9, e108109. [Google Scholar] [CrossRef]
- Barnable, P.; Calenda, G.; Bonnaire, T.; Menon, R.; Levendosky, K.; Gettie, A.; Blanchard, J.; Cooney, M.L.; Fernandez-Romero, J.A.; Zydowsky, T.M.; et al. MIV-150/zinc acetate gel inhibits cell-associated simian-human immunodeficiency virus reverse transcriptase infection in a macaque vaginal explant model. Antimicrob. Agents Chemother. 2015, 59, 3829–3837. [Google Scholar] [CrossRef][Green Version]
- Cummins, J.E., Jr.; Guarner, J.; Flowers, L.; Guenthner, P.C.; Bartlett, J.; Morken, T.; Grohskopf, L.A.; Paxton, L.; Dezzutti, C.S. Preclinical testing of candidate topical microbicides for anti-human immunodeficiency virus type 1 activity and tissue toxicity in a human cervical explant culture. Antimicrob. Agents Chemother. 2007, 51, 1770–1779. [Google Scholar] [CrossRef] [PubMed]
- Perotti, M.E.; Pirovano, A.; Phillips, D.M. Carrageenan formulation prevents macrophage trafficking from vagina: Implications for microbicide development. Biol. Reprod. 2003, 69, 933–939. [Google Scholar] [CrossRef] [PubMed]
- Kenney, J.; Aravantinou, M.; Singer, R.; Hsu, M.; Rodriguez, A.; Kizima, L.; Abraham, C.J.; Menon, R.; Seidor, S.; Chudolij, A.; et al. An antiretroviral/zinc combination gel provides 24 hours of complete protection against vaginal SHIV infection in macaques. PLoS ONE 2011, 6, e15835. [Google Scholar] [CrossRef] [PubMed]
- Singer, R.; Derby, N.; Rodriguez, A.; Kizima, L.; Kenney, J.; Aravantinou, M.; Chudolij, A.; Gettie, A.; Blanchard, J.; Lifson, J.D.; et al. The nonnucleoside reverse transcriptase inhibitor MIV-150 in carrageenan gel prevents rectal transmission of simian/human immunodeficiency virus infection in macaques. J. Virol. 2011, 85, 5504–5512. [Google Scholar] [CrossRef] [PubMed]
- Kenney, J.; Singer, R.; Derby, N.; Aravantinou, M.; Abraham, C.J.; Menon, R.; Seidor, S.; Zhang, S.; Gettie, A.; Blanchard, J.; et al. A single dose of a MIV-150/Zinc acetate gel provides 24 h of protection against vaginal simian human immunodeficiency virus reverse transcriptase infection, with more limited protection rectally 8–24 h after gel use. AIDS Res. Hum. Retrovir. 2012, 28, 1476–1484. [Google Scholar] [CrossRef]
- Halpern, V.; Ogunsola, F.; Obunge, O.; Wang, C.H.; Onyejepu, N.; Oduyebo, O.; Taylor, D.; McNeil, L.; Mehta, N.; Umo-Otong, J.; et al. Effectiveness of cellulose sulfate vaginal gel for the prevention of HIV infection: Results of a Phase III trial in Nigeria. PLoS ONE 2008, 3, e3784. [Google Scholar] [CrossRef]
- Van Damme, L.; Govinden, R.; Mirembe, F.M.; Guedou, F.; Solomon, S.; Becker, M.L.; Pradeep, B.S.; Krishnan, A.K.; Alary, M.; Pande, B.; et al. Lack of effectiveness of cellulose sulfate gel for the prevention of vaginal HIV transmission. N. Engl. J. Med. 2008, 359, 463–472. [Google Scholar] [CrossRef]
- Roan, N.R.; Munch, J.; Arhel, N.; Mothes, W.; Neidleman, J.; Kobayashi, A.; Smith-McCune, K.; Kirchhoff, F.; Greene, W.C. The cationic properties of SEVI underlie its ability to enhance human immunodeficiency virus infection. J. Virol. 2009, 83, 73–80. [Google Scholar] [CrossRef]
- Munch, J.; Rucker, E.; Standker, L.; Adermann, K.; Goffinet, C.; Schindler, M.; Wildum, S.; Chinnadurai, R.; Rajan, D.; Specht, A.; et al. Semen-derived amyloid fibrils drastically enhance HIV infection. Cell 2007, 131, 1059–1071. [Google Scholar] [CrossRef]
- Derby, N.; Lal, M.; Aravantinou, M.; Kizima, L.; Barnable, P.; Rodriguez, A.; Lai, M.; Wesenberg, A.; Ugaonkar, S.; Levendosky, K.; et al. Griffithsin carrageenan fast dissolving inserts prevent SHIV HSV-2 and HPV infections in vivo. Nat. Commun. 2018, 9, 3881. [Google Scholar] [CrossRef]
- Zeitlin, L.; Whaley, K.J.; Hegarty, T.A.; Moench, T.R.; Cone, R.A. Tests of vaginal microbicides in the mouse genital herpes model. Contraception 1997, 56, 329–333. [Google Scholar] [CrossRef]
- Bourne, K.Z.; Bourne, N.; Reising, S.F.; Stanberry, L.R. Plant products as topical microbicide candidates: Assessment of in vitro and in vivo activity against herpes simplex virus type 2. Antivir. Res. 1999, 42, 219–226. [Google Scholar] [CrossRef]
- Gonzalez, M.E.; Carrasco, L. Animal viruses promote the entry of polysaccharides with antiviral activity into cells. Biochem. Biophys. Res. Commun. 1987, 146, 1303–1310. [Google Scholar] [CrossRef]
- Carlucci, M.J.; Scolaro, L.A.; Noseda, M.D.; Cerezo, A.S.; Damonte, E.B. Protective effect of a natural carrageenan on genital herpes simplex virus infection in mice. Antivir. Res. 2004, 64, 137–141. [Google Scholar] [CrossRef]
- Fernandez-Romero, J.A.; Abraham, C.J.; Rodriguez, A.; Kizima, L.; Jean-Pierre, N.; Menon, R.; Begay, O.; Seidor, S.; Ford, B.E.; Gil, P.I.; et al. Zinc acetate/carrageenan gels exhibit potent activity in vivo against high-dose herpes simplex virus 2 vaginal and rectal challenge. Antimicrob. Agents Chemother. 2012, 56, 358–368. [Google Scholar] [CrossRef]
- Levendosky, K.; Mizenina, O.; Martinelli, E.; Jean-Pierre, N.; Kizima, L.; Rodriguez, A.; Kleinbeck, K.; Bonnaire, T.; Robbiani, M.; Zydowsky, T.M.; et al. Griffithsin and Carrageenan Combination To Target Herpes Simplex Virus 2 and Human Papillomavirus. Antimicrob. Agents Chemother. 2015, 59, 7290–7298. [Google Scholar] [CrossRef]
- Maguire, R.A.; Bergman, N.; Phillips, D.M. Comparison of microbicides for efficacy in protecting mice against vaginal challenge with herpes simplex virus type 2, cytotoxicity, antibacterial properties, and sperm immobilization. Sex. Transm. Dis. 2001, 28, 259–265. [Google Scholar] [CrossRef]
- Stiles, J.; Guptill-Yoran, L.; Moore, G.E.; Pogranichniy, R.M. Effects of lambda-carrageenan on in vitro replication of feline herpesvirus and on experimentally induced herpetic conjunctivitis in cats. Investig. Ophthalmol. Vis. Sci. 2008, 49, 1496–1501. [Google Scholar] [CrossRef]
- Zacharopoulos, V.R.; Phillips, D.M. Vaginal formulations of carrageenan protect mice from herpes simplex virus infection. Clin. Diagn. Lab. Immunol. 1997, 4, 465–468. [Google Scholar] [CrossRef]
- Maguire, R.A.; Zacharopoulos, V.R.; Phillips, D.M. Carrageenan-based nonoxynol-9 spermicides for prevention of sexually transmitted infections. Sex. Transm. Dis. 1998, 25, 494–500. [Google Scholar] [CrossRef]
- Carlucci, M.J.; Scolaro, L.A.; Damonte, E.B. Herpes simplex virus type 1 variants arising after selection with an antiviral carrageenan: Lack of correlation between drug susceptibility and syn phenotype. J. Med. Virol. 2002, 68, 92–98. [Google Scholar] [CrossRef]
- Mateu, C.G.; Perez Recalde, M.; Artuso, M.C.; Hermida, G.; Linero, F.N.; Scolaro, L.A.; Damonte, E.B.; Pujol, C.A.; Carlucci, M.J. Emergence of Herpes Simplex Virus-1 Syncytial Variants With Altered Virulence for Mice After Selection With a Natural Carrageenan. Sex. Transm. Dis. 2011, 38, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Carolina Artuso, M.; Linero, F.N.; Gazzaniga, S.; Scolaro, L.A.; Pujol, C.A.; Wainstok, R.; Josefina Carlucci, M. Altered expression of cytokines in mice infected intranasally with two syncytial variants of Herpes simplex virus type 1. Microb. Pathog. 2014, 71, 68–72. [Google Scholar] [CrossRef] [PubMed]
- Graf, C.; Bernkop-Schnurch, A.; Egyed, A.; Koller, C.; Prieschl-Grassauer, E.; Morokutti-Kurz, M. Development of a nasal spray containing xylometazoline hydrochloride and iota-carrageenan for the symptomatic relief of nasal congestion caused by rhinitis and sinusitis. Int. J. Gen. Med. 2018, 11, 275–283. [Google Scholar] [CrossRef]
- Grassauer, A.; Weinmuellner, R.; Meier, C.; Pretsch, A.; Prieschl-Grassauer, E.; Unger, H. Iota-Carrageenan is a potent inhibitor of rhinovirus infection. Virol. J. 2008, 5, 107. [Google Scholar] [CrossRef] [PubMed]
- Leibbrandt, A.; Meier, C.; Konig-Schuster, M.; Weinmullner, R.; Kalthoff, D.; Pflugfelder, B.; Graf, P.; Frank-Gehrke, B.; Beer, M.; Fazekas, T.; et al. Iota-carrageenan is a potent inhibitor of influenza A virus infection. PLoS ONE 2010, 5, e14320. [Google Scholar] [CrossRef] [PubMed]
- Morokutti-Kurz, M.; Graf, C.; Prieschl-Grassauer, E. Amylmetacresol/2,4-dichlorobenzyl alcohol, hexylresorcinol, or carrageenan lozenges as active treatments for sore throat. Int. J. Gen. Med. 2017, 10, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Morokutti-Kurz, M.; Konig-Schuster, M.; Koller, C.; Graf, C.; Graf, P.; Kirchoff, N.; Reutterer, B.; Seifert, J.M.; Unger, H.; Grassauer, A.; et al. The Intranasal Application of Zanamivir and Carrageenan Is Synergistically Active against Influenza A Virus in the Murine Model. PLoS ONE 2015, 10, e0128794. [Google Scholar] [CrossRef]
- Yu, G.; Li, M.; Wang, W.; Liu, X.; Zhao, X.; Lv, Y.; Li, G.; Jiao, G.; Zhao, X. Structure and anti-influenza A (H1N1) virus activity of three polysaccharides from Eucheuma denticulatum. J. Ocean Univ. China 2012, 11, 527–532. [Google Scholar] [CrossRef]
- Fujisawa, H.; Tsuru, S.; Taniguchi, M.; Zinnaka, Y.; Nomoto, K. Protective mechanisms against pulmonary infection with influenza virus. I. Relative contribution of polymorphonuclear leukocytes and of alveolar macrophages to protection during the early phase of intranasal infection. J. Gen. Virol. 1987, 68 Pt 2, 425–432. [Google Scholar] [CrossRef]
- Tang, F.; Chen, F.; Li, F. Preparation and potentialin vivoanti-influenza virus activity of low molecular-weight κ-carrageenans and their derivatives. J. Appl. Polym. Sci. 2013, 127, 2110–2115. [Google Scholar] [CrossRef]
- Tsuru, S.; Fujisawa, H.; Taniguchi, M.; Zinnaka, Y.; Nomoto, K. Mechanism of protection during the early phase of a generalized viral infection. II. Contribution of polymorphonuclear leukocytes to protection against intravenous infection with influenza virus. J. Gen. Virol. 1987, 68 Pt 2, 419–424. [Google Scholar] [CrossRef]
- Tsuru, S.; Kitani, H.; Seno, M.; Abe, M.; Zinnaka, Y.; Nomoto, K. Mechanism of protection during the early phase of a generalized viral infection. I. Contribution of phagocytes to protection against ectromelia virus. J. Gen. Virol. 1983, 64 Pt 9, 2021–2026. [Google Scholar] [CrossRef] [PubMed]
- Hebar, A.; Koller, C.; Seifert, J.M.; Chabicovsky, M.; Bodenteich, A.; Bernkop-Schnurch, A.; Grassauer, A.; Prieschl-Grassauer, E. Non-clinical safety evaluation of intranasal iota-carrageenan. PLoS ONE 2015, 10, e0122911. [Google Scholar] [CrossRef] [PubMed]
- Eccles, R.; Meier, C.; Jawad, M.; Weinmullner, R.; Grassauer, A.; Prieschl-Grassauer, E. Efficacy and safety of an antiviral Iota-Carrageenan nasal spray: A randomized, double-blind, placebo-controlled exploratory study in volunteers with early symptoms of the common cold. Respir. Res. 2010, 11, 108. [Google Scholar] [CrossRef] [PubMed]
- Fazekas, T.; Eickhoff, P.; Pruckner, N.; Vollnhofer, G.; Fischmeister, G.; Diakos, C.; Rauch, M.; Verdianz, M.; Zoubek, A.; Gadner, H.; et al. Lessons learned from a double-blind randomised placebo-controlled study with a iota-carrageenan nasal spray as medical device in children with acute symptoms of common cold. BMC Complement. Altern. Med. 2012, 12, 147. [Google Scholar] [CrossRef]
- Ludwig, M.; Enzenhofer, E.; Schneider, S.; Rauch, M.; Bodenteich, A.; Neumann, K.; Prieschl-Grassauer, E.; Grassauer, A.; Lion, T.; Mueller, C.A. Efficacy of a carrageenan nasal spray in patients with common cold: A randomized controlled trial. Respir. Res. 2013, 14, 124. [Google Scholar] [CrossRef]
- Koenighofer, M.; Lion, T.; Bodenteich, A.; Prieschl-Grassauer, E.; Grassauer, A.; Unger, H.; Mueller, C.A.; Fazekas, T. Carrageenan nasal spray in virus confirmed common cold: Individual patient data analysis of two randomized controlled trials. Multidiscip. Respir. Med. 2014, 9, 57. [Google Scholar] [CrossRef]
- Eccles, R.; Winther, B.; Johnston, S.L.; Robinson, P.; Trampisch, M.; Koelsch, S. Efficacy and safety of iota-carrageenan nasal spray versus placebo in early treatment of the common cold in adults: The ICICC trial. Respir. Res. 2015, 16, 121. [Google Scholar] [CrossRef]
- Kines, R.C.; Cerio, R.J.; Roberts, J.N.; Thompson, C.D.; de Los Pinos, E.; Lowy, D.R.; Schiller, J.T. Human papillomavirus capsids preferentially bind and infect tumor cells. Int. J. Cancer 2016, 138, 901–911. [Google Scholar] [CrossRef]
- Wang, J.W.; Jagu, S.; Kwak, K.; Wang, C.; Peng, S.; Kirnbauer, R.; Roden, R.B. Preparation and properties of a papillomavirus infectious intermediate and its utility for neutralization studies. Virology 2014, 449, 304–316. [Google Scholar] [CrossRef] [PubMed]
- Novetsky, A.P.; Keller, M.J.; Gradissimo, A.; Chen, Z.; Morgan, S.L.; Xue, X.; Strickler, H.D.; Fernandez-Romero, J.A.; Burk, R.; Einstein, M.H. In vitro inhibition of human papillomavirus following use of a carrageenan-containing vaginal gel. Gynecol. Oncol. 2016, 143, 313–318. [Google Scholar] [CrossRef]
- Roberts, J.N.; Buck, C.B.; Thompson, C.D.; Kines, R.; Bernardo, M.; Choyke, P.L.; Lowy, D.R.; Schiller, J.T. Genital transmission of HPV in a mouse model is potentiated by nonoxynol-9 and inhibited by carrageenan. Nat. Med. 2007, 13, 857–861. [Google Scholar] [CrossRef] [PubMed]
- Marais, D.; Gawarecki, D.; Allan, B.; Ahmed, K.; Altini, L.; Cassim, N.; Gopolang, F.; Hoffman, M.; Ramjee, G.; Williamson, A.L. The effectiveness of Carraguard, a vaginal microbicide, in protecting women against high-risk human papillomavirus infection. Antivir. Ther. 2011, 16, 1219–1226. [Google Scholar] [CrossRef]
- Roberts, J.N.; Kines, R.C.; Katki, H.A.; Lowy, D.R.; Schiller, J.T. Effect of Pap smear collection and carrageenan on cervicovaginal human papillomavirus-16 infection in a rhesus macaque model. J. Natl. Cancer Inst. 2011, 103, 737–743. [Google Scholar] [CrossRef] [PubMed]
- Magnan, S.; Tota, J.E.; El-Zein, M.; Burchell, A.N.; Schiller, J.T.; Ferenczy, A.; Tellier, P.P.; Coutlee, F.; Franco, E.L.; Group, C.S. Efficacy of a Carrageenan gel Against Transmission of Cervical HPV (CATCH): Interim analysis of a randomized, double-blind, placebo-controlled, phase 2B trial. Clin. Microbiol. Infect. 2019, 25, 210–216. [Google Scholar] [CrossRef]
- Perino, A.; Consiglio, P.; Maranto, M.; De Franciscis, P.; Marci, R.; Restivo, V.; Manzone, M.; Capra, G.; Cucinella, G.; Calagna, G. Impact of a new carrageenan-based vaginal microbicide in a female population with genital HPV-infection: First experimental results. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 6744–6752. [Google Scholar]
- Gonzalez, M.E.; Alarcon, B.; Carrasco, L. Polysaccharides as antiviral agents: Antiviral activity of carrageenan. Antimicrob. Agents Chemother. 1987, 31, 1388–1393. [Google Scholar] [CrossRef]
- Tamhankar, M.; Gerhardt, D.M.; Bennett, R.S.; Murphy, N.; Jahrling, P.B.; Patterson, J.L. Heparan sulfate is an important mediator of Ebola virus infection in polarized epithelial cells. Virol. J. 2018, 15, 135. [Google Scholar] [CrossRef]
- Chiu, Y.H.; Chan, Y.L.; Tsai, L.W.; Li, T.L.; Wu, C.J. Prevention of human enterovirus 71 infection by kappa carrageenan. Antivir. Res. 2012, 95, 128–134. [Google Scholar] [CrossRef]
- Tseligka, E.D.; Sobo, K.; Stoppini, L.; Cagno, V.; Abdul, F.; Piuz, I.; Meylan, P.; Huang, S.; Constant, S.; Tapparel, C. A VP1 mutation acquired during an enterovirus 71 disseminated infection confers heparan sulfate binding ability and modulates ex vivo tropism. PLoS Pathog. 2018, 14, e1007190. [Google Scholar] [CrossRef] [PubMed]
- Pavliga, S.N.; Kompanets, G.G.; Tsygankov, V.Y. The Experimental Research (In Vitro) of Carrageenans and Fucoidans to Decrease Activity of Hantavirus. Food Environ. Virol. 2016, 8, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Klimyte, E.M.; Smith, S.E.; Oreste, P.; Lembo, D.; Dutch, R.E. Inhibition of Human Metapneumovirus Binding to Heparan Sulfate Blocks Infection in Human Lung Cells and Airway Tissues. J. Virol. 2016, 90, 9237–9250. [Google Scholar] [CrossRef] [PubMed]
- Kimberlin, R.H.; Walker, C.A. Suppression of scrapie infection in mice by heteropolyanion 23, dextran sulfate, and some other polyanions. Antimicrob. Agents Chemother. 1986, 30, 409–413. [Google Scholar] [CrossRef]


{kind=link}
| CG Type | Cell Line | CC50 (µg/mL) | Ref |
|---|---|---|---|
| κ-CG | MDCK cells | >250 | [34] |
| κ-CG | Vero cells | >1000 | [43] |
| κ-CG | PRK cells | >100 | [18] |
| κ-CG | Vero cells | 2000 | [27] |
| κ-CG CO-1 | MDCK and A549 cells | 857 | [42] |
| ι-CG | Vero cells | 250 | [35] |
| ι-CG | Vero cells | >1000 | [21] |
| ι-CG | Vero cells | >500 | [19] |
| ι-CG | Vero and C6/36 HT mosquito cells | >1000 | [16] |
| ι-CG | Vero cells | >400 | [32] |
| ι-CG | Vero cells | >5000 | [43] |
| λ-CG | Vero cells | 400 | [17] |
| λ-CG | Vero cells | >10 | [31] |
| λ-CG | Vero cells | >1000 | [23] |
| λ-CG | MDBK cells | >1000 | [26] |
| λ-CG | Vero and C6/36 HT mosquito cells | >1000 | [37] |
| λ-CG | BSR cells | >1000 | [30] |
| λ-CG | MT-4 cells | 300 | [41] |
| λ-CG | Vero cells | >5000 | [43] |
| λ-CG | PRK cells | >100 | [18] |
| λ-CG | Vero cells | 3000 | [27] |
| κ/λ-CG (PC-515) | Hela cells | >16 | [33] |
| Oxidized κ/ι-CG | Vero cells | >1000 | [25] |
| ι/λ-CG | HeLa cells | >100 | [20] |
| κ/ι/λ-CG | PLC/PRF/5 cells | >1000 | [28] |
| κ/ι/λ-CG | Vero, HepG2 and foreskin PH cells | >1000 | [36] |
| κ/ι/λ-CG | Vero and C6/36 HT mosquito cells | >1000 | [38] |
| κ/ι-CG, λ-CG, and υ/ν-CG | Vero and human foreskin fibroblast cells | >1000 | [22] |
| κ/ι-CG, λ-CG, and υ/ν-CG | Mouse astrocytes and Vero cells | >1000 | [24] |
| κ/ι/ν-CG | Vero, HepG2, and foreskin cells | >1000 | [39] |
| κ/ι/ν-CG C2 | Vero cells | >1000 | [40] |
| Unspecified CG | BHK-21 fibroblast cells | >5 | [29] |
| CG Type | Human Subject | Duration and Frequency | Effects | Ref |
|---|---|---|---|---|
| 2% ι-CG (PC-213) | 25 women | Once a day for 1 week | No irritation of the female reproductive tract by colposcopy | [44] |
| 3% κ/λ-CG (PC-515) | 165 women | 4 times per week for 1 year | No abnormal clinical signs or symptoms, no vaginal flora change, and highly acceptable | [51] |
| 3% κ/λ-CG (PC-515) | 55 HIV(−) couples | Before sexual intercourse for 6 months | Highly acceptable and increased sexual pleasure | [48] |
| 3% κ/λ-CG (PC-515) | 60 HIV(+) healthy women and men | Once a day or before sexual intercourse for 2 weeks | Highly acceptable | [52] |
| 3% κ/λ-CG (PC-515) | 60 HIV(+) healthy women and men | Once a day or before sexual intercourse for 2 weeks | No abnormal clinical signs or symptoms, no vaginal flora change, and no genital shedding of HIV RNA | [46] |
| 3% κ/λ-CG (PC-515) | 55 HIV(−) women | Before sexual intercourse for 1 month | Intact epithelium and no increased proinflammatory cytokines in CVL | [47] |
| 3% κ/λ-CG (PC-515) | 55 HIV(−) couples | 2–3 times per week before sexual intercourse for 6 months | Generally acceptable, no epithelial disruption, and no abnormal genital flora | [45] |
| 3% κ/λ-CG (PC-515) | 60 HIV(+) women crossover | Once daily for 1 week followed by 3-week wash-out for 1 month | Reduced HIV viral load in CVL and well tolerated | [50] |
| 3% κ/λ-CG (PC-515) | 400 HIV(−) women | 3 times a week before sexual intercourse for 6 months | No genital irritation or epithelial disruption, no difference in STI rate, and generally acceptable | [53] |
| 3% κ/λ-CG (PC-515) | 60 HIV(+) women crossover | Once-daily for 1 week for 3 months with cross-over | Highly acceptable | [54] |
| 3% κ/λ-CG + MIV-150 + ZA (PC-1005) | 25 HIV(−) women | Once-daily for 2 weeks | Well tolerated and maintenance of anti-HIV and anti-HPV activity in CVL | [49] |
| CG Type | Experimental System | Antiviral Assay | IC50 (µg/mL) | Ref |
|---|---|---|---|---|
| λ-CG | MT-4 cells infected with HIV-HTLVIIIB | Expression of HIV-specific antigens and plaque reduction assays | 9500 (IU/mL) | [61] |
| ι-CG | MT-4 cells infected with HIV | ELISA of p24 antigen | 100 | [56] |
| ι-CG and κ-CG | MT-4 cells and ME-180 cells infected with HIV | ELISA of p24 antigen | 1.6 and 4 | [55] |
| Depolymerized λ-CG | MT-4 cells infected with HIV-HTLVIIIB | Expression of HIV-specific antigens | 3.9 (IC100) | [66] |
| κ-CG and λ-CG | MT-4 cells infected with HIV-HTLVIIIB | Expression of HIV-specific antigens | 12 and 1.9 | [18] |
| κ-CG and AZT conjugates | MT-4 cells infected with HIV (BRU) | Quantitation of virus-induced cytopathicity | 0.1 | [41] |
| λ/κ-CG + MIV-150 | CEM-SS cells infected with HIVMN and HIV-2CDC310342 | Microtiter syncytial assay | 0.1–13.8 | [67] |
| λ/κ-CG (PC-515) | TMZ.bI and DC cells infected with HIV-Bal | Quantitation of β-gal and ELISA of p24 antigen | 0.03–4.17, 1.61 | [12] |
| ι-CG | HeLa-CD4-CCR5 cells infected with HIVJR-FL | Luciferase assay | 1–10 | [68] |
| λ-CG | P4-R5 MAGI cells | ELISA of p24 antigen | 3.7 and 64 | [13] |
| λ-CG | MT-2 cells infected with HIV BaL and IIIB | ELISA of p24 antigen | 0.58 and 0.68 | [14] |
| CG Type | Experimental Systems | Antiviral Assay | Effects | Ref |
|---|---|---|---|---|
| λ/κ-CG (PC-515) | Human cervical explants infected with HIVBaL | ELISA of p24 antigen | 50% inhibition of infection | [71] |
| λ/κ-CG + MIV-150 + ZA (PC-1005) | Human cervical explants incubated with CVLs and then infected with HIVBaL | ELISA of p24 antigen | Inhibition of infection | [9] |
| λ/κ-CG + MIV-150 + ZA (PC-1005) | Macaque vaginal explants infected with SHIV-RT | ELISA of p24 antigen | Inhibition of infection | [7] |
| λ/κ-CG + MIV-150 + ZA (PC-1005) | Macaques vaginal explants challenged with SHIV-RT | Quantitation of viral DNA and ELISA of SIVmac p27 | Inhibition of infection | [69] |
| λ/κ-CG + MIV-150 + ZA (PC-1005) | Macaques vaginal explants challenged with SHIV-RT infected PBMCs | Quantitation of viral DNA and ELISA of SIVmac p27 | Inhibition of infection | [70] |
| CG Type | Efficacy Model | Dose | ADMINISTRATION | Antiviral Assay | Effects | Ref |
|---|---|---|---|---|---|---|
| λ/κ-CG | Inoculation of stained mouse macrophages into the vagina of mice | 20 µL of stock | Single inoculation | Counting the number of macrophages in lymph nodes | 90% inhibition | [72] |
| λ/κ-CG + TFV + Zn + MIV-150 | Macaques vaginally challenged with SHIV-RT | 3 mL of 3% gel | Single vaginal | Plasma viral load quantitation | Inhibition of vaginal transmission and no difference in antiviral activity | [7] |
| λ/κ-CG + MIV-150 (PC-817) | Macaques rectally challenged with SHIV-RT | 3 mL of 3% gel | Single rectal | Plasma viral load quantitation | Inhibition of rectal transmission | [12] |
| λ/κ-CG + MIV-150 + ZA (PC-1005) | Macaques either vaginally or rectally challenged with SHIV-RT | 2 mL of 3% gel | Single vaginal | Plasma viral load quantitation | Inhibition of infection | [74] |
| λ/κ-CG + MIV-150 + ZA (PC-1005) | Macaques either vaginally or rectally challenged with SHIV-RT | 2 mL of 3% gel | Single vaginal or rectal | Plasma viral load quantitation | Complete and limited inhibition of vaginal or rectal transmission | [73] |
| λ/κ-CG + ZA | Macaques either vaginally or rectally challenged with SHIV-RT | 2 mL of 3% gel | Vaginal for 2 weeks | Plasma viral load quantitation | Inhibition of infection | [75] |
| λ/κ-CG + MIV-150 + ZA (PC-1005) | Macaques vaginal explant challenged with either free or cell-associated SHIV-RT | 1:100 and 1:300 dilution | Immersion of explant with diluted gels | Quantitation of viral DNA and ELISA of SIVmac p27 | Inhibition of infection | [8] |
| λ/κ-CG | Preventive effects on 6202 HIV(−) women | 4 mL of 3% gel | Before sexual intercourse for 9–24 months | HIV incidence by seroconversion | No difference in HIV incidence | [11] |
| CG Type | Herpes Virus Type | Cell Line | Antiviral Assay | IC50 (µg/mL) | Ref |
|---|---|---|---|---|---|
| ι-CG | HSV-1 and 2 | Vero cells | PFU | 2 and 10 | [83] |
| λ/κ-CG | HSV-1 and 2 | PPK cells | CPE | 3.7, 1.6, 2, 1.5 | [18] |
| λ/κ-CG | CMV | Hela cells | CPE | 2.8/0.3 | [18] |
| λ-CG | HSV-1 and 2 | Vero cells | CPE and PFU | <100 | [31] |
| λ-CG | HSV-1 and 2 | Vero cells | CPE and PFU | 0.3 | [23] |
| κ-CG | HSV-2 | Human foreskin fibroblast cells | CPE | 0.01 | [81] |
| λ-CG type IV, ι-CG type V | HSV-2 | HeLa cells | CPE | 2.4 and 1.4 | [82] |
| λ-CG 1T1, κ/ι-CG 1C1, µ/ν-CG 1C3 | HSV-1 strain F and HSV-2 strain G | Vero cells and human diploid foreskin fibroblast cell line PH | PFU | 0.4–3.3 | [22] |
| λ-CG 1T1, κ/ι-CG 1C1, µ/ν-CG 1C3 | HSV-1 strain F and HSV-2 strain G | Mouse astrocytes and Vero cells | PFU | 0.4–3.6 | [24] |
| ι-CG | HSV-1 and 2 | Vero cells | PFU | 0.65–9.33 | [21] |
| κ/ι/ν-CG, C2 | HSV-1 strain F and HSV-2 strain G | Vero cells | PFU and survival | 0.5–5.6 | [40] |
| Oxidized k- and ι-CG | HSV-1 and 2 | Vero cells | PFU | 0.98–34.3 | [25] |
| λ-CG | BoHV-1 and SuHV-1 | MDBK cells | PFU | 0.52 and 10.42 | [26] |
| ι-CG | HSV-1 | Vero cells | Neutral red dye | 6.31 | [32] |
| κ/ι/λ-CG | VZV | Vero cells | PFU | 0.5/0.8/1.8 | [43] |
| CG Type | Efficacy Model | Dose | Antiviral Assay | Administration | Effects | Ref |
|---|---|---|---|---|---|---|
| ι-CG type V | ICR mice infected with MCMV | 0.5 mg | PFU | Intraperitoneal | Decreased mortality and titer and increased PFU/LD50 | [63] |
| ι-CG type V | ICR mice infected with MCMV | 0.5 mg | PFU | Intraperitoneal | Inhibition of viral spread from the peritoneal cavity to the plasma | [62] |
| λ/κ/ι-CG | BALB/c mice infected with HSV-2 | 0.05–1% | PFU in vaginal secretion | Vaginal | Inhibition of infection | [89] |
| κ-CG | C57B1/6 mice infected with HSV-2 | 0.1–100 mg/mL | Shedding of virus, visible lesions, and CPE | Vaginal | Inhibition of infection | [81] |
| CG-based nonoxynol-9 | BALB/c mice infected with HSV-2 | 20 ul of 3% | Symptom of infection | Vaginal | Inhibition of infection | [90] |
| λ-CG type IV (1T1), ι-CG type V | Swiss Webster mice infected with HSV-2 | 10 mg/mL | Sign of disease | Vaginal | Inhibition of infection | [82] |
| λ-CG | BALB/c mice infected with HSV-2 | 2 and 3% | Survival | Vaginal | 100% survival | [87] |
| λ-CG 1T1 | BALB/c mice infected with HSV-2 | 10 mg/mL | PFU and survival | Vaginal | 90% survival and no virus shedding | [84] |
| κι/ν-CG C2 | BALB/c mice infected with HSV-2 | 8 mg/mL | PFU and survival | Vaginal | 70% survival | [40] |
| λ-CG type IV | FHV-1 induced conjunctivitis in cats | 250 µg/mL | PFU | Topical | Reduction of virus titers but no alteration in the clinical course | [88] |
| λ/κ-CG + ZA | BALB/c mice infected with HSV-2 | 20 µL of 3% | Survival | Vaginal and rectal | Inhibition of infection | [85] |
| λ/κ-CG + ZA | BALB/c mice infected with HSV-2 | 20 µL of 3% | Survival | Vaginal and rectal | Inhibition of infection | [8] |
| CG + griffithsin | BALB/c mice infected with HSV-2 | 50 µL of griffithsin solution (19.1 mg/mL) | Infection rate | Vaginal | Inhibition of infection | [86] |
| CG + MIV-150 + ZA | BALB/c mice infected with HSV-2 | 10 µL of stock | qPCR | Vaginal | Inhibition of infection | [7] |
| CG + griffithsin | BALB/c mice infected with HSV-2 | 10 µL of 0.1% stock | qPCR | Vaginal | Inhibition of infection | [80] |
| CG Type | Virus Type | Cell Line | Anti-Viral Assay | IC50 (µg/mL) | Ref |
|---|---|---|---|---|---|
| ι-CG | HRV 1A, 2, 8, 14, 16, 83, and 84 | Human nasal epithelial and HeLa cells | TCID50 | 5–10 | [95] |
| ι-CG | Influenza virus strain H1N1 (A/PR8/34) and the formerly pandemic H3N2 (A/Aichi/2/68) | MDCK cells and human nasal epithelial cells | PFU | 0.04–0.2 | [96] |
| κ-CG | Influenza A virus | MDCK and A549 cells | TCID50 assay and RT-PCR | 32.1 | [42] |
| ι/κ/ν-CG hybrid | Influenza A H1N1 virus | MDCK cells | CPE inhibition | 276.5 | [99] |
| ι-CG + zanamivir | Influenza A virus H1N1(09)pdm, H3N2, H5N1, and H7N7 | MDCK cells | Immunostaining | 0.39–11.8 | [98] |
| κ-CG | Swine pandemic influenza A virus H1N1 | MDCK cells | TCID50, CPE experiments, IF, and Western blot | 89.57 | [34] |
| ι-CG (lozenges) | HRV 1a, HRV8, influenza virus A H1N1, coxsackievirus A10, and human coronavirus OC43 | HeLa, MDCK, and Vero cells | TCID50 and PFU, agglutination assay | 234–4524 dilution | [97] |
| ι-CG and xylometazoline hydrochloride (Coldamaris) | HRV 1a, HRV8, and human coronavirus OC43 | HeLa, MDCK, and Vero cells | TCID50 and PFU, agglutination assay | 16.5, 1.66, and 0.024 | [94] |
| CG Type | Virus Type | Experimental Model | Dose | Antiviral Assay | Effects | Ref |
|---|---|---|---|---|---|---|
| Type II CG | Influenza virus H1N1 (A/PR/8/34) | BALB/c mice intranasal infection | 1 mg/mL | Virus titration with MDCK cells | Depleted macrophages and increased PMN in the blood, enhancement of viral titer | [100] |
| Type II CG | Influenza virus H1N1 (A/PR/8/34) | BALB/c mice intranasal infection | 200 mg/kg | Virus titration with MDCK cells | Depleted macrophages, phagocytes, and monocytes | [102] |
| ι-CG | Influenza virus strain H1N1 (A/PR8/34) and the formerly pandemic H3N2 (A/Aichi/2/68) | C57Bl/6 mice intranasal infection | 60 µg BID for 15 days | PFU | Reduced viral titers and increased survival (40%) | [96] |
| κ-CG (low MW) | Mouse adapted influenza virus A/FM/1/47(H1N1) | ICR mice with nasal drip | 40 µL of 1.5 mg/mL for 7 days | Measure pulmonary edema index | Inhibition of pulmonary edema in mice | [101] |
| κ/ι-CG and zanamivir | Influenza A virus H1N1(09)pdm, H3N2, H5N1, and H7N7 | C57BL/6 mice with intranasal injection | 50 µL of 1.2 and 0.4 mg/mL stock BID for 5 days | Survival and weight loss, necropsy, and inflammation | Increased survival, decreased weight loss, reduced necropsy, and inflammation | [98] |
| CG Type | Virus Type | Experimental Model | Dose | Antiviral Assay | Effects | Ref |
|---|---|---|---|---|---|---|
| ι-CG (Coldmaris) | Respiratory viruses (influenza, parainfluenza, coronavirus, rhinoviruses, and human metapneumovirus) | 35 human subjects, nasal spray, symptom scores | 0.12% | Measure viral loads in nasal lavages | Lowered symptom scores and viral loads, lowered proinflammatory cytokines | [105] |
| ι-CG (Coldmaris) | Respiratory viruses (influenza, parainfluenza, coronavirus, RSV, rhinoviruses, and human metapneumovirus) | 213 young human subjects, nasal spray, symptom scores | 0.12% | Measure viral loads in nasal lavages | Lowered viral loads but no effects on symptom scores | [106] |
| ι-CG (Coldmaris) | Respiratory viruses (influenza, parainfluenza, coronavirus, RSV, rhinoviruses, and human metapneumovirus) | 211 patients intranasal spray | 0.12% | Measure viral loads in nasal lavages | Reduced duration of disease, alleviation of symptom, and reduced viral titers | [107] |
| ι-CG | Rhinovirus, coronavirus, and influenza A virus | 254 human subjects with nasal spray | 0.12% TID for 7 days | Nasal lavage sample | Reduced duration of disease, increased viral clearance, and reduced relapses of symptom | [108] |
| ι-CG | Rhinovirus, coronavirus, and influenza A virus | 200 human subjects with nasal spray | 0.12% QID for 4–10 days | Nasal lavage sample | No difference in total symptom scores and more effective in coronavirus | [109] |
| CG Type | Virus Type | Experimental System | Anti-Viral Assay | IC50 (ng/mL) | Ref |
|---|---|---|---|---|---|
| λ/ι-CG | HPV16 | HeLa cells infected with pseudovirus with GFP | GFP assay | 5–44 | [20] |
| κ/λ-CG (PC-515) | HPV16, 18, and 45 PsVs | HeLa cells infected with pseudovirus with luciferase | Luciferase assay | 1–20 | [33] |
| CG | HPV16, 18, 31, 34, 58, and 6 with luciferase reporter | 293T cell infected with furin-cleaved HPV16 pseudovirus | Luciferase assay | 250–1000 | [111] |
| CG | HPV16 | HeLa cells infected with pseudovirus with luciferase | Luciferase assay | 38.6 | [86] |
| 1% ι-CG | HPV 16 VLP and PsV | NCI-60 cells infected with RFP-encoded HPV pseudovirus | GFP assay by FACS | N/D | [110] |
| CG Type | Virus Type | Experimental System | Anti-Viral Assay Readout | Effects | Ref |
|---|---|---|---|---|---|
| ι-CG (1%) | HPV16 | Mouse cervicovaginal mucosa infected with RFP-encoded pseudovirus with GFP capsid | GFP and RFP assay | Inhibition of infection | [113] |
| ι-CG (1%) | HPV16 | 12 rhesus macaques infected with RFP-encoded pseudovirus with GFP capsid | Immunohistochemistry of the infected cervix tissue | Decreased infection that was enhanced by cytologic examination | [115] |
| 3% λ/κ-CG (PC-515) + divine 9 | HPV16 | Mouse cervicovaginal mucosa infected with RFP-encoded pseudovirus with GFP capsid | GFP and RFP assay | Inhibition not affected by seminal plasma | [33] |
| Griffithsin + CG | HPV16 | Mouse cervicovaginal mucosa infected with luciferase-encoded pseudovirus | Luciferase | Inhibition | [86] |
| CG-based lubricant (Divine 9) | HPV16 | Antiviral activity of cervicovaginal lavage (CVL) using 293T cells infected with SEAP-encoded PsV | RFP assay | Inhibition | [112] |
| Griffithsin + CG | HPV | Balb/C mice equipped with FDI infected with luciferase-encoded HPV16 pseudoviruses | Luciferase | Inhibition | [80] |
| CG Type | Virus Type | Experimental System | Anti-Viral Assay Readout | Effects | Ref |
|---|---|---|---|---|---|
| λ/κ-CG (carraguard) | High-risk HPVs | 1718 women used gel plus condoms during each act of vaginal intercourse | Prevalence of high-risk HPV infection by Pap smear | Lower prevalence of high-risk HPV infection | [114] |
| CG-based lubricant (Divine 9) | HPV | 280 women used gel plus condoms during each act of vaginal intercourse | Prevalence of high-risk HPV infection by Pap smear | Lower prevalence of HPV infection | [116] |
| 0.02% CG and Propionibacterium extract (CGP) | HPV | 40 HPV-infected women | Clearance of HPV infection | Accelerated the clearance of HPV infection (26 to 60% clearance rate) | [117] |
| CG Type | Virus Type | Experimental System | Anti-Viral Asssay | IC50 (µg/mL) | Ref |
|---|---|---|---|---|---|
| κ/ι/ν-CG | DENV2 | Vero cells, human hepatoma HepG2, and foreskin PH cells | Plaque formation | 1 | [39] |
| ι/λ/κ-CG | DENV1, 2, 3, 4 | Vero cells, human hepatoma HepG2, and foreskin PH cells | Plaque formation | 0.1–40.7 | [36] |
| ι/λ/κ-CG | DENV2 | Vero cells and C6/36 HT mosquito cells | Plaque formation | 0.4/7 | [38] |
| ι-CG | DENV2 | Vero cells and C6/36 HT mosquito cells | Plaque formation | 22.5/0.64 | [16] |
| CG | DENV2 | BHK-21 fibroblast cells | Measuring cellular oxygen consumption rate | 10 | [29] |
| λ-CG | DENV2 | Vero cells and C6/36 HT mosquito cells | Plaque formation | >50 | [37] |
| Virus Type | CG Type | Cell Line | IC50 or CPE50 (µg/mL) | Ref |
|---|---|---|---|---|
| Adenovirus type 2 | κ and λ | Hela cells | >200/>200 | [18] |
| Adenovirus type 5 | ι | Hela cells | >200 | [118] |
| African swine fever virus | ι | Vero cell | 10 | [118] |
| African swine fever virus | ι | Vero cell | 50 | [35] |
| African swine fever virus | κ and λ | Vero cell | 25/150 | [27] |
| Arenavirus | λ | Vero cells | 0.2–0.3 | [17] |
| Chikungunya virus | ι | Vero cells | 3.8 | [19] |
| Cosackiesvirus type B4 | κ and λ | Hela cells | >400/>400 | [18] |
| Ebola virus | ι | Caco-2 cells | N/D | [119] |
| Encephalomyocarditis virus | ι | Not specified | 10 | [118] |
| Enterovirus 71 | κ | Vero cells | 10–100 | [120] |
| Enterovirus 71 | ι | Vero cells | 17.8 | [121] |
| Hantavirus | ι | Vero cells and mouse macrophages | [122] | |
| Hepatitis A virus | κ, ι and λ | PLC/PRF/5 cells | 2.5, 4.5, and 100.3 | [28] |
| Measles | ι | Not specified | >200 | [118] |
| Metapneumoniavirus | ι | Vero and human bronchial epithelial cells (BEAS-2B) | 0.1–1 | [123] |
| Parainfluenza virus type 3 | κ and λ | Vero cells | >40/>4 | [18] |
| Polio type 1 | ι | Hela cells | >200 | [118] |
| Polio type 1 | κ and λ | Hela cells | >400/>400 | [18] |
| Rabies | λ P32 | BSR cells | 15–57 | [30] |
| Reovirus type 1 | κ and λ | Vero cells | >40/>4 | [18] |
| Semliki Forest virus | ι | Vero cells | 0.7 | [19] |
| Semliki Forest virus | ι | BHK-21 cells | 10 | [118] |
| Sindbis virus | κ and λ | Vero cells | 7/2 | [18] |
| Scrapie | λ type IV | Female compton mouse infected with scrapie | inhibition of infection | [124] |
| Vaccinia | ι | PPK cells | 10 | [118] |
| Vaccinia | κ and λ | PPK cells | 36/16 | [18] |
| Vesicular stomatitis virus | ι | PRK cells | >200 | [118] |
| Vesicular stomatitis virus | κ and λ | PRK cells | 0.3/0.2 | [18] |
| Vesicular stomatitis virus | κ and λ | Hela cells | 7/4 | [18] |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, C. Carrageenans as Broad-Spectrum Microbicides: Current Status and Challenges. Mar. Drugs 2020, 18, 435. https://doi.org/10.3390/md18090435
Lee C. Carrageenans as Broad-Spectrum Microbicides: Current Status and Challenges. Marine Drugs. 2020; 18(9):435. https://doi.org/10.3390/md18090435
Chicago/Turabian StyleLee, Choongho. 2020. "Carrageenans as Broad-Spectrum Microbicides: Current Status and Challenges" Marine Drugs 18, no. 9: 435. https://doi.org/10.3390/md18090435
APA StyleLee, C. (2020). Carrageenans as Broad-Spectrum Microbicides: Current Status and Challenges. Marine Drugs, 18(9), 435. https://doi.org/10.3390/md18090435