The Marine Seagrass Halophila stipulacea as a Source of Bioactive Metabolites against Obesity and Biofouling

 , , ,
, , ,  , , ,
, , ,  , and
, and 
Abstract
1. Introduction
2. Results
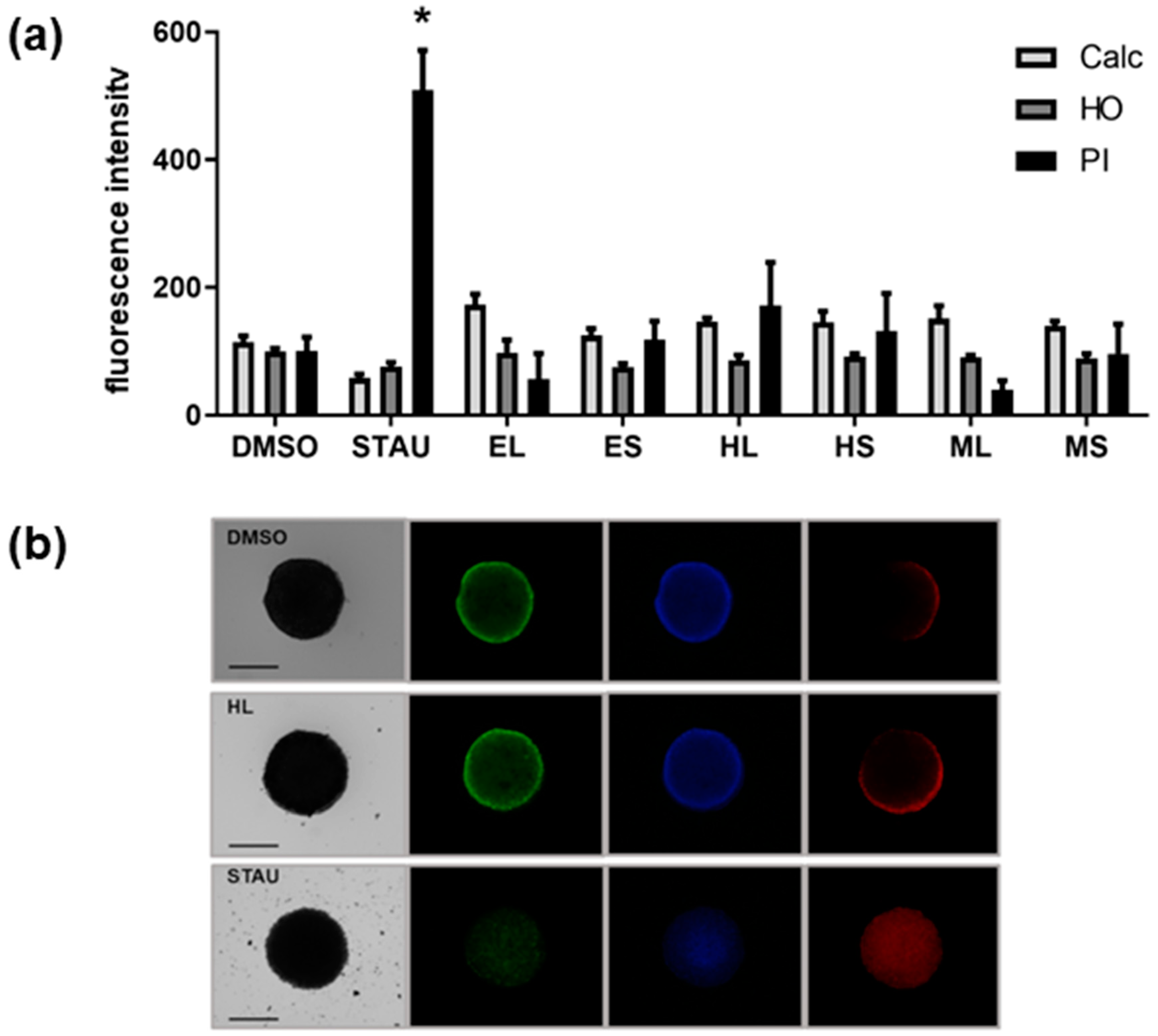
2.1. Cytotoxic Activity
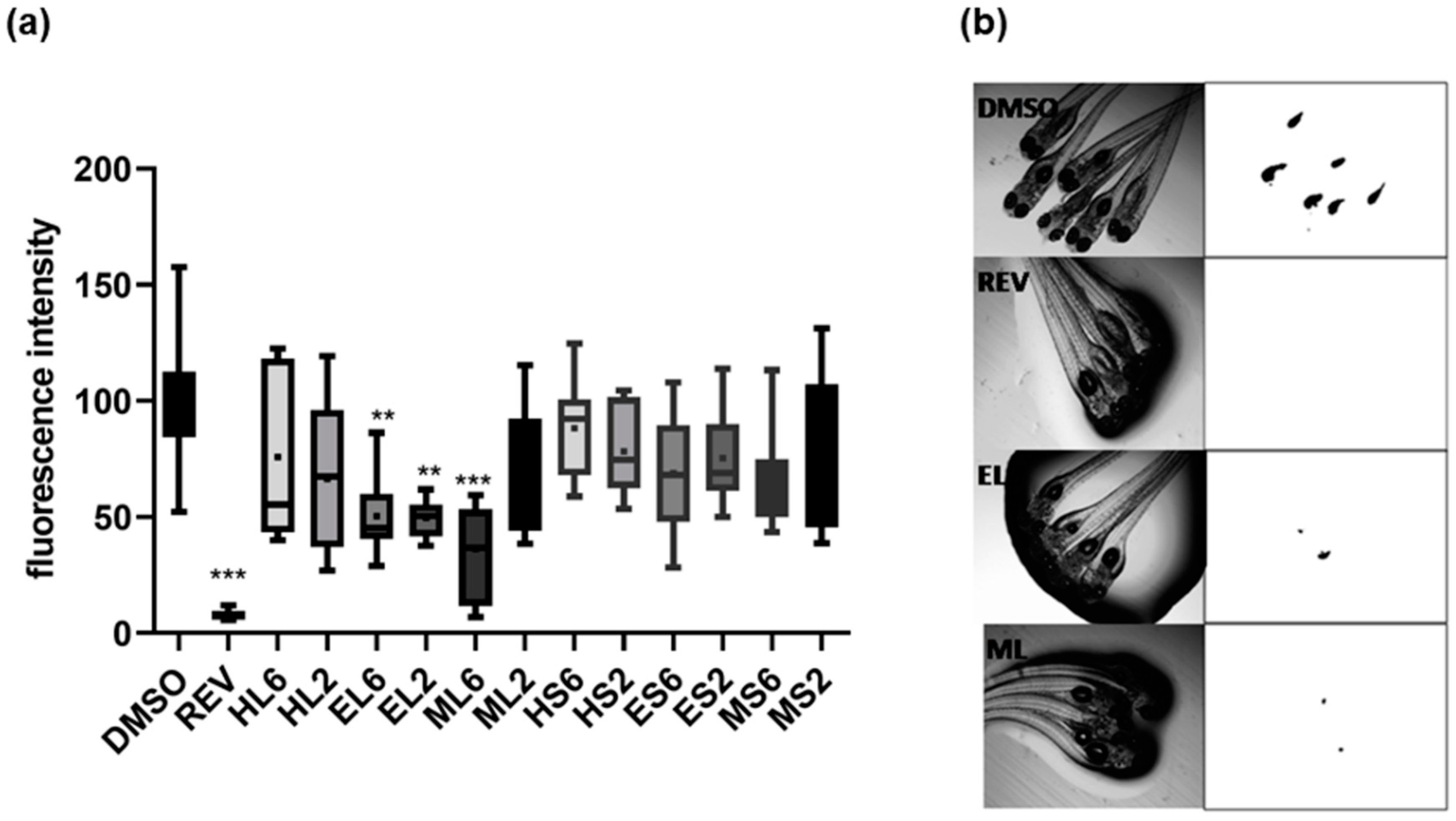
2.2. Lipid-Reducing Activity
2.3. Anti-Steatosis and Anti-Diabetes Activities
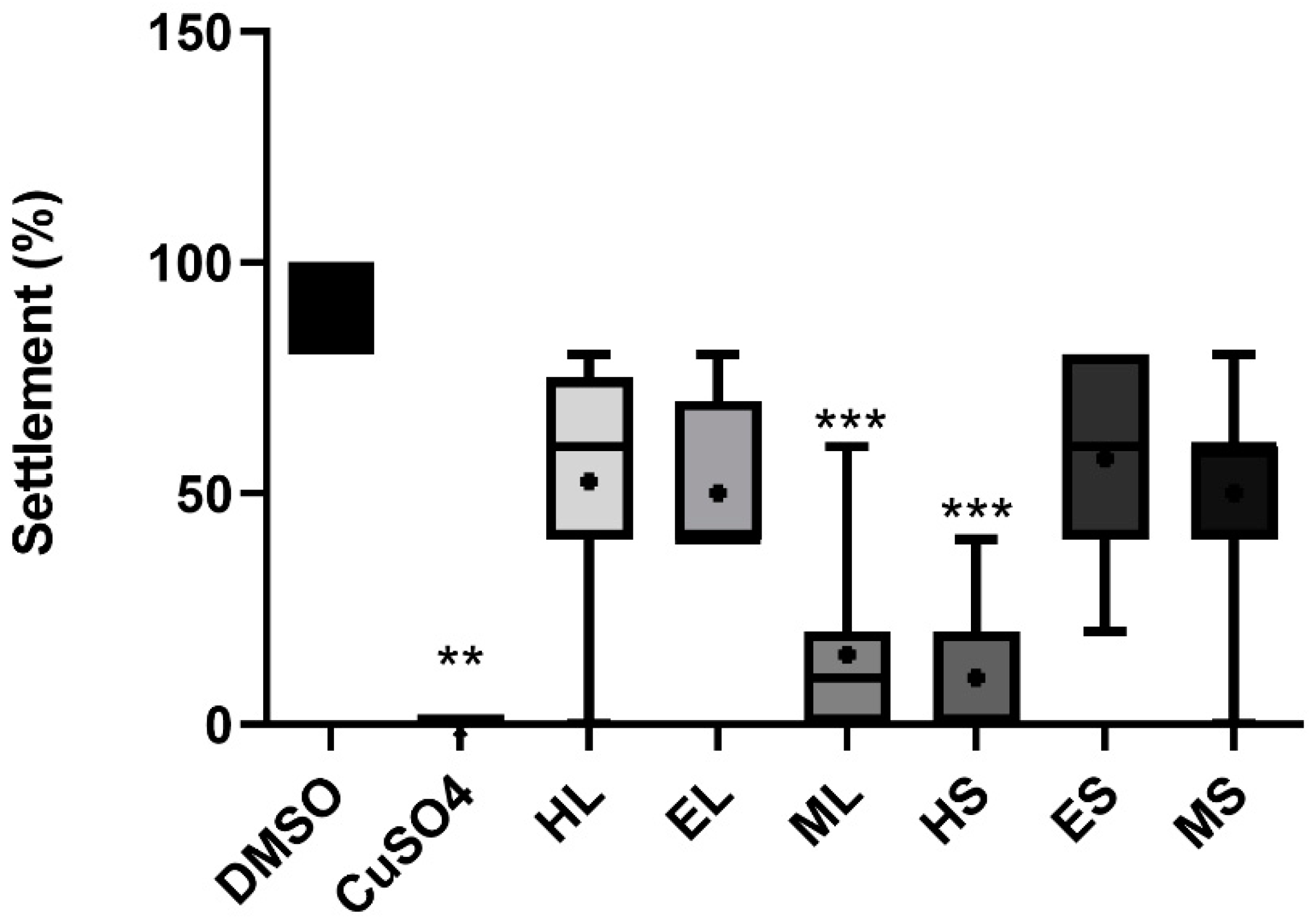
2.4. Anti-Fouling Activity
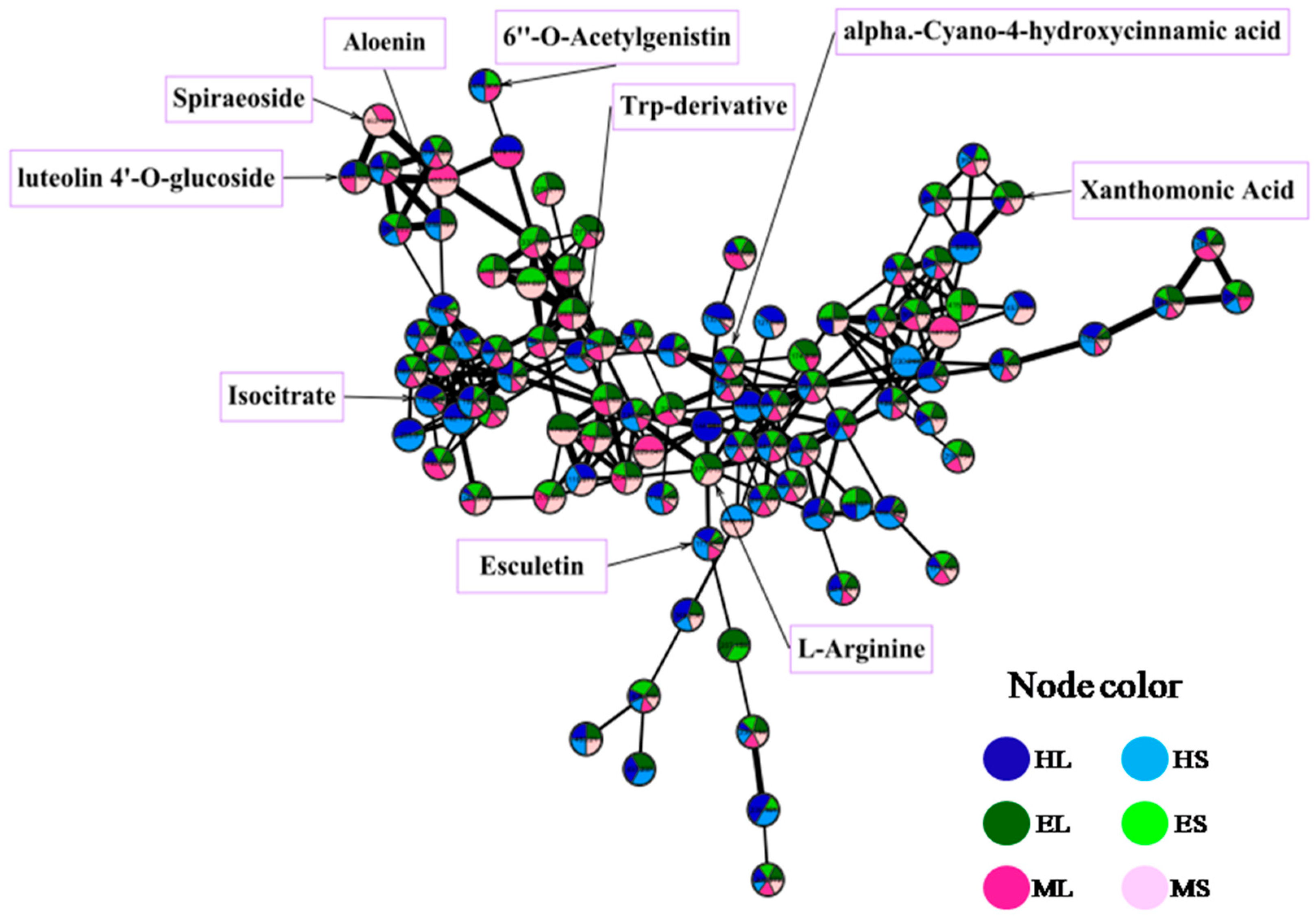
2.5. Metabolite Profiling
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Preparation of Seagrass Extracts
4.3. Cell Culture
4.4. Cytotoxicity on Cancer Cell Lines
4.5. Cytotoxicity in 3D Cell Culture of HCT-116 Cell Line
4.6. Zebrafish Nile Red Fat Metabolism Assay
4.7. Glucose Uptake Assay
4.8. Anti-Steatosis Assay
4.9. Bioassay with Marine Fouling Bacteria
4.10. Mussel Larvae Anti-Biofouling Bioassays
4.11. UPLC-HRMS/MS Analysis
4.12. Molecular Networking
4.13. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Carte, B.K. Biomedical Potential of Marine Natural Products. Bioscience 1996, 46, 271–286. [Google Scholar]
- Cornara, L.; Pastorino, G.; Borghesi, B.; Salis, A.; Clericuzio, M.; Marchetti, C.; Damonte, G.; Burlando, B. Posidonia oceanica (L.) Delile Ethanolic Extract Modulates Cell Activities with Skin Health Applications. Mar. Drugs 2018, 16, 21. [Google Scholar] [CrossRef] [PubMed]
- Missaoui, H. Sur la présence de la phanérogame marine Halophila stipulacea (Forsskål) dans le golfe de Gabes. Bull. Inst. Natl. Sci. Technol. Mer Salammbô 2003, 30, 111–114. [Google Scholar]
- Sghaier, Y.R.; Zakhama-Sraieb, R.; Benamer, I.; Charfi-Cheikhrouha, F. Occurrence of the seagrass Halophila stipulacea (Hydrocharitaceae) in the southern Mediterranean Sea. Bot. Mar. 2011, 54, 575–582. [Google Scholar] [CrossRef]
- Mollo, E.; Gavagnin, M.; Carbone, M.; Castelluccio, F.; Pozone, F.; Roussis, V.; Templado, J.; Ghiselin, M.T.; Cimino, G. Factors promoting marine invasions: A chemoecological approach. Proc. Natl. Acad. Sci. USA 2008, 105, 4582–4586. [Google Scholar] [CrossRef] [PubMed]
- Kannan Rengasamy, R.R.; Rajasekaran, A.; Micheline, G.-D.; Perumal, A. Antioxidant activity of seagrasses of the Mandapam coast, India. Pharm. Biol. 2012, 50, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Bonanno, G.; Raccuia, S.A. Seagrass Halophila stipulacea: Capacity of accumulation and biomonitoring of trace elements. Sci. Total Environ. 2018, 633, 257–263. [Google Scholar] [CrossRef]
- Papenbrock, J. Highlights in Seagrasses’ Phylogeny, Physiology, and Metabolism: What Makes Them Special? ISRN Bot. 2012, 2012, 1–15. [Google Scholar] [CrossRef]
- Reddy, L.; Odhav, B.; Bhoola, K.D. Natural products for cancer prevention: A global perspective. Pharmacol. Ther. 2003, 99, 1–13. [Google Scholar] [CrossRef]
- Wang, Y.; Kong, D.; Wang, X.; Dong, X.; Tao, Y.; Gong, H. Molecular Mechanisms of Luteolin Induced Growth Inhibition and Apoptosis of Human Osteosarcoma Cells. Iran. J. Pharm. Res. 2015, 14, 531. [Google Scholar]
- Das, A.; Banik, N.L.; Ray, S.K. Mechanism of apoptosis with the involvement of calpain and caspase cascades in human malignant neuroblastoma SH-SY5Y cells exposed to flavonoids. Int. J. Cancer 2006, 119, 2575–2585. [Google Scholar] [CrossRef] [PubMed]
- Bitam, F.; Ciavatta, M.L.; Carbone, M.; Manzo, E.; Mollo, E.; Gavagnin, M. Chemical analysis of flavonoid constituents of the seagrass Halophila stipulacea: First finding of malonylated derivatives in marine phanerogams. Biochem. Syst. Ecol. 2010, 38, 686–690. [Google Scholar] [CrossRef]
- Klangprapun, S.; Buranrat, B.; Caichompoo, W.; Nualkaew, S. Pharmacognostical and Physicochemical Studies of Enhalus acoroides (L.F.) Royle (Rhizome). Pharmacogn. J. 2018, 10, 89–94. [Google Scholar] [CrossRef]
- Su, S.; Cheng, X.; Wink, M. Cytotoxicity of arctigenin and matairesinol against the T-cell lymphoma cell line CCRF-CEM: Cytotoxicity of lignans against CCRF-CEM cells. J. Pharm. Pharmacol. 2015, 67, 1316–1323. [Google Scholar] [CrossRef]
- Choi, H.; Mevers, E.; Byrum, T.; Valeriote, F.A.; Gerwick, W.H. Lyngbyabellins K-N from Two Palmyra Atoll Collections of the Marine Cyanobacterium Moorea bouillonii. Eur. J. Org. Chem. 2012, 27, 5141–5150. [Google Scholar] [CrossRef]
- Luesch, H.; Yoshida, W.Y.; Moore, R.E.; Paul, V.J.; Mooberry, S.L. Isolation, Structure Determination, and Biological Activity of Lyngbyabellin A from the Marine Cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2000, 63, 611–615. [Google Scholar] [CrossRef]
- Gavagnin, M.; Carbone, M.; Amodeo, P.; Mollo, E.; Vitale, R.M.; Roussis, V.; Cimino, G. Structure and Absolute Stereochemistry of Syphonoside, a Unique Macrocyclic Glycoterpenoid from Marine Organisms. J. Org. Chem. 2007, 72, 5625–5630. [Google Scholar] [CrossRef]
- Hamdy, A.-H.A.; Mettwally, W.S.A.; Fotouh, M.A.E.; Rodriguez, B.; El-Dewany, A.I.; El-Toumy, S.A.A.; Hussein, A.A. Bioactive Phenolic Compounds from the Egyptian Red Sea Seagrass Thalassodendron ciliatum. Z. Für Naturforschung 2012, 67, 291–296. [Google Scholar] [CrossRef]
- Caro, M.; Sansone, A.; Amezaga, J.; Navarro, V.; Ferreri, C.; Tueros, I. Wine lees modulate lipid metabolism and induce fatty acid remodelling in Zebrafish. Food Funct. 2017, 8, 1652–1659. [Google Scholar] [CrossRef]
- Urbatzka, R.; Freitas, S.; Palmeira, A.; Almeida, T.; Moreira, J.; Azevedo, C.; Afonso, C.; Correia-da-Silva, M.; Sousa, E.; Pinto, M.; et al. Lipid reducing activity and toxicity profiles of a library of polyphenol derivatives. Eur. J. Med. Chem. 2018, 151, 272–284. [Google Scholar] [CrossRef]
- Santos, J.D.; Vitorino, I.; De la Cruz, M.; Díaz, C.; Cautain, B.; Annang, F.; Pérez-Moreno, G.; Gonzalez Martinez, I.; Tormo, J.R.; Martín, J.M.; et al. Bioactivities and Extract Dereplication of Actinomycetales Isolated From Marine Sponges. Front. Microbiol. 2019, 10, 727. [Google Scholar] [CrossRef] [PubMed]
- Freitas, S.; Silva, N.G.; Sousa, M.L.; Ribeiro, T.; Rosa, F.; Leão, P.N.; Vasconcelos, V.; Reis, M.A.; Urbatzka, R. Chlorophyll Derivatives from Marine Cyanobacteria with Lipid-Reducing Activities. Mar. Drugs 2019, 17, 229. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.; Coello, L.; Urbatzka, R.; Pérez, M.; Thorsteinsdottir, M. New Aromatic Bisabolane Derivatives with Lipid-Reducing Activity from the Marine Sponge Myrmekioderma sp. Mar. Drugs 2019, 17, 375. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.S.; Alimov, A.P.; Rilo, H.L.; Jandacek, R.J.; Woollett, L.A.; Penberthy, W.T. A high throughput live transparent animal bioassay to identify non-toxic small molecules or genes that regulate vertebrate fat metabolism for obesity drug development. Nutr. Metab. 2008, 5, 23. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.; Park, J.; Kim, H.-L.; Kang, J.; Jeong, M.-Y.; Youn, D.-H.; Jung, Y.; Kim, Y.-I.; Kim, H.-J.; Ahn, K.S.; et al. Chrysophanic Acid Suppresses Adipogenesis and Induces Thermogenesis by Activating AMP-Activated Protein Kinase Alpha In vivo and In vitro. Front. Pharmacol. 2016, 7, 476. [Google Scholar] [CrossRef]
- Seo, M.-J.; Lee, Y.-J.; Hwang, J.-H.; Kim, K.-J.; Lee, B.-Y. The inhibitory effects of quercetin on obesity and obesity-induced inflammation by regulation of MAPK signaling. J. Nutr. Biochem. 2015, 26, 1308–1316. [Google Scholar] [CrossRef]
- Castro, M.; Preto, M.; Vasconcelos, V.; Urbatzka, R. Obesity: The Metabolic Disease, Advances on Drug Discovery and Natural Product Research. Curr. Top. Med. Chem. 2016, 16, 2577–2604. [Google Scholar] [CrossRef]
- Lee, H.; Kang, R.; Yoon, Y. Shikonin inhibits fat accumulation in 3T3-L1 adipocytes. Phytother. Res. 2010, 24, 344–351. [Google Scholar] [CrossRef]
- Zarrouki, B.; Pillon, N.J.; Kalbacher, E.; Soula, H.A.; Nia N’Jomen, G.; Grand, L.; Chambert, S.; Geloen, A.; Soulage, C.O. Cirsimarin, a potent antilipogenic flavonoid, decreases fat deposition in mice intra-abdominal adipose tissue. Int. J. Obes. 2010, 34, 1566–1575. [Google Scholar] [CrossRef]
- Zhao, L.; Cen, F.; Tian, F.; Li, M.-J.; Zhang, Q.; Shen, H.; Shen, X.-C.; Zhou, M.; Du, J. Combination treatment with quercetin and resveratrol attenuates high fat diet-induced obesity and associated inflammation in rats via the AMPKα1/SIRT1 signaling pathway. Exp. Ther. Med. 2017, 14, 5942–5948. [Google Scholar] [CrossRef]
- Hashimura, H.; Ueda, C.; Kawabata, J.; Kasai, T. Acetyl-CoA Carboxylase Inhibitors from Avocado (Persea americana Mill) Fruits. Biosci. Biotechnol. Biochem. 2001, 65, 1656–1658. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-H.; Chang, M.-Y.; Lin, Y.-S.; Lin, D.-G.; Chen, S.-W.; Chao, P.-M. A herbal extract with acetyl–coenzyme A carboxylase inhibitory activity and its potential for treating metabolic syndrome. Metabolism 2009, 58, 1297–1305. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Ramachandran, U.; Raichur, S.; Chakrabarti, R.; Jain, R. Synthesis and evaluation of N-acetyl-l-tyrosine based compounds as PPARα selective activators. Eur. J. Med. Chem. 2007, 42, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Iyapparaj, P.; Revathi, P.; Ramasubburayan, R.; Prakash, S.; Palavesam, A.; Immanuel, G.; Anantharaman, P.; Sautreau, A.; Hellio, C. Antifouling and toxic properties of the bioactive metabolites from the seagrasses Syringodium isoetifolium and Cymodocea serrulata. Ecotoxicol. Environ. Saf. 2014, 103, 54–60. [Google Scholar] [CrossRef]
- Getachew, P.; Getachew, M.; Joo, J.; Choi, Y.S.; Hwang, D.S.; Hong, Y.-K. The slip agents oleamide and erucamide reduce biofouling by marine benthic organisms (diatoms, biofilms and abalones). Toxicol. Environ. Health Sci. 2016, 8, 341–348. [Google Scholar] [CrossRef]
- Almeida, J.R.; Correia-da-Silva, M.; Sousa, E.; Antunes, J.; Pinto, M.; Vasconcelos, V.; Cunha, I. Antifouling potential of Nature-inspired sulfated compounds. Sci. Rep. 2017, 7, 42424. [Google Scholar] [CrossRef]
- Bragadeeswaran, S.; Prabhu, K.; Thangaraj, S.; Ganesan, K.; Rani, S.S. Biological activity of Seaweed extracts from Cladophora clavuligera (Kutzing, 1843) and Sargassum wightii (Greville, 1995) against marine fouling bacteria. Indian J. Mar. Sci. 2011, 40, 398–402. [Google Scholar]
- Newby, B.Z.; Cutright, T.; Barrios, C.A.; Xu, Q. Zosteric acid—An effective antifoulant for reducing fresh water bacterial attachment on coatings. J. Coat. Technol. Res. 2006, 3, 69–76. [Google Scholar] [CrossRef]
- Sghaier, Y.R.; Zakhama-Sraieb, R.; Mouelhi, S.; Vazquez, M.; Valle, C.; Ramos-Espla, A.A.; Astier, J.M.; Verlaque, M.; Charfi-Cheikhrouha, F. Review of alien marine macrophytes in Tunisia. Mediterr. Mar. Sci. 2015, 17, 109–123. [Google Scholar] [CrossRef]
- Sirenko, O.; Mitlo, T.; Hesley, J.; Luke, S.; Owens, W.; Cromwell, E.F. High-Content Assays for Characterizing the Viability and Morphology of 3D Cancer Spheroid Cultures. ASSAY Drug Dev. Technol. 2015, 13, 402–414. [Google Scholar] [CrossRef]
- Kamentsky, L.; Jones, T.R.; Fraser, A.; Bray, M.-A.; Logan, D.J.; Madden, K.L.; Ljosa, V.; Rueden, C.; Eliceiri, K.W.; Carpenter, A.E. Improved structure, function and compatibility for CellProfiler: Modular high-throughput image analysis software. Bioinformatics 2011, 27, 1179–1180. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.; Rosa, F.; Ribeiro, T.; Hernandez-Bautista, R.; Bonaldo, M.; Gonçalves Silva, N.; Eiríksson, F.; Thorsteinsdóttir, M.; Ussar, S.; Urbatzka, R. Identification of Cyanobacterial Strains with Potential for the Treatment of Obesity-Related Co-Morbidities by Bioactivity, Toxicity Evaluation and Metabolite Profiling. Mar. Drugs 2019, 17, 280. [Google Scholar] [CrossRef] [PubMed]
- Quinn, R.A.; Nothias, L.-F.; Vining, O.; Meehan, M.; Esquenazi, E.; Dorrestein, P.C. Molecular Networking As a Drug Discovery, Drug Metabolism, and Precision Medicine Strategy. Trends Pharmacol. Sci. 2017, 38, 143–154. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell line | MG63 | SHSY5Y | HCT116 | hCMEC | ||||
|---|---|---|---|---|---|---|---|---|
| Extracts | 24 h | 48 h | 24 h | 48 h | 24 h | 48 h | 24 h | 48 h |
| HL | 19.5 ± 5.8 | 29.1 ± 7.5 | >30 | >30 | ||||
| EL | >30 | 29.4 ± 6.3 | 10.6 ± 7.0 | 15.2 ± 1.9 | 11.3 ± 1.8 | 24.5 ± 15.6 | ||
| HS | 7.6 ± 5.4 | 25.4 ± 4.2 | >30 | >30 | ||||
| ES | >30 | 19.1 ± 9.0 | 23.4 ± 1.1 | 18.7 ± 3.1 | 9.2 ± 0.2 | 15.4 ± 1.1 | ||
| Class | Compound Name | m/z | HL | EL | ML | HS | ES | MS |
|---|---|---|---|---|---|---|---|---|
| Apigenin | 271.228 | x | x | x | x | x | x | |
| Luteolin | 287.055 | x | x | |||||
| Matairesinol | 377.142 | x | x | |||||
| Polyphenols | 2,4-dihydroxyheptadec-16-ynyl acetate | 325.184 | x | |||||
| Spiraeoside | 463.124 | x | x | |||||
| 5-hydroxy-6,7-dimethoxy-2-[4-[(2S,3R,4S,5S,6R)-3,4,5-trihydroxy-6-(hydroxymethyl)oxan-2-yl]oxyphenyl]chromen-4-one | 521.13 | x | x | x | ||||
| alpha-Cyano-4-hydroxycinnamic acid | 189.052 | x | x | x | x | x | x | |
| 3-Hydroxy-4-methoxycinnamic acid | 194.127 | x | x | x | x | x | x | |
| Fatty Acids | 13-Docosenamide, (Z) | 338.342 | x | x | x | x | ||
| Amino acids and peptides | Lyngbyabellin A | 713.473 | x | x | ||||
| N-Acetyl-L-tyrosine | 225.926 | x | x |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bel Mabrouk, S.; Reis, M.; Sousa, M.L.; Ribeiro, T.; Almeida, J.R.; Pereira, S.; Antunes, J.; Rosa, F.; Vasconcelos, V.; Achour, L.; et al. The Marine Seagrass Halophila stipulacea as a Source of Bioactive Metabolites against Obesity and Biofouling. Mar. Drugs 2020, 18, 88. https://doi.org/10.3390/md18020088
Bel Mabrouk S, Reis M, Sousa ML, Ribeiro T, Almeida JR, Pereira S, Antunes J, Rosa F, Vasconcelos V, Achour L, et al. The Marine Seagrass Halophila stipulacea as a Source of Bioactive Metabolites against Obesity and Biofouling. Marine Drugs. 2020; 18(2):88. https://doi.org/10.3390/md18020088
Chicago/Turabian StyleBel Mabrouk, Sawssen, Mariana Reis, Maria Lígia Sousa, Tiago Ribeiro, Joana R. Almeida, Sandra Pereira, Jorge Antunes, Filipa Rosa, Vitor Vasconcelos, Lotfi Achour, and et al. 2020. "The Marine Seagrass Halophila stipulacea as a Source of Bioactive Metabolites against Obesity and Biofouling" Marine Drugs 18, no. 2: 88. https://doi.org/10.3390/md18020088
APA StyleBel Mabrouk, S., Reis, M., Sousa, M. L., Ribeiro, T., Almeida, J. R., Pereira, S., Antunes, J., Rosa, F., Vasconcelos, V., Achour, L., Kacem, A., & Urbatzka, R. (2020). The Marine Seagrass Halophila stipulacea as a Source of Bioactive Metabolites against Obesity and Biofouling. Marine Drugs, 18(2), 88. https://doi.org/10.3390/md18020088