Bioactive Properties of Marine Phenolics


Abstract
1. Introduction
2. Marine Phenolics: Sources and Phenolic Composition
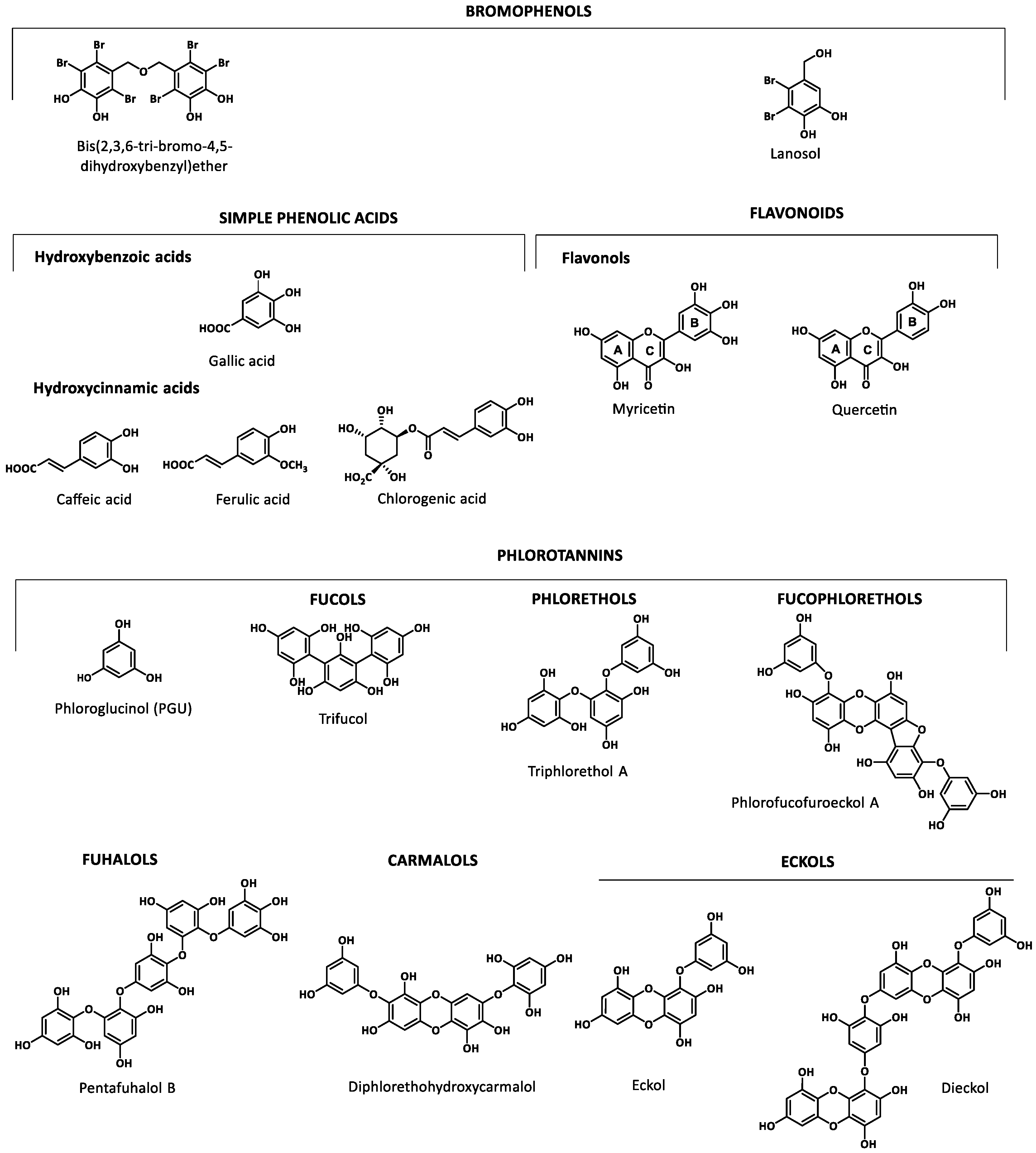
2.1. Families of Phenolic Compounds Identified in Marine Sources
2.2. Sources
2.2.1. Seawater
2.2.2. Microalgae

{kind=link}
{kind=link}
| Marine Organism Extraction Chemical Analysis | Phenolic Compounds Antioxidant Activity (When Provided) | Ref. |
|---|---|---|
| Marine-derived Fungus | ||
| Alternaria sp. SCSIO41014 from sponge AC, EtOAc, US HPLC-UV, HRESIMS, NMR, ECD, XRay | Perylenequinone derivatives; altenusin derivative; phenol derivatives | [62] |
| Arthrinium sp. MeOH, MeCN HPLC, HRESIMS, NMR | 2-(2,3-Dihydroxy-5-methyl benzoyl)-6-hydroxybenzoic acid | [63] |
| Aspergillus sydowii, from the sponge Stelletta sp. EtOAc, BuOH HPLC, UV, IR, HRESIMS, NMR, OP | Diorcinolic acid; β-d-glucopyranosyl aspergillusene A; diphenylethers; chromone; xanthone first glycoside of phenolic bisabolane sesquiterpenes | [64] |
| Aspergillus sp. from the sponge Xestospongia testudinaria EtOAc, AC RP-HPLC, HRESIMS, NMR | Phenolic bisabolane sesquiterpenoid dimers (disydonols A–C), (S)-(+)-sydonol | [65] |
| Aspergillus sp. from the sponge Chondrilla nucula EtOAc HPLC–PDA, UV, HRESIMS, NMR, OP | Phenolic bisabolane sesquiterpenes; asperchondol A; asperchondol B | [66] |
| Aspergillus sp., from the sponge Chondrilla nucula EtOAc, BuOH HPLC, UV, IR, HRESIMS, NMR, OP | Phenolic bisabolane sesquiterpenes; asperchondols A and B; diphenyl ethers | [64] |
| Aspergillus versicolor, deep-sea fungus EtOAc, BuOH HPLC, IR, TLC, HRESIMS, NMR, OP, ECD | Aspergilols A–F; diorcinal; cordyol E; 4-carboxydiorcinal; 4-methoxycarbonyldiorcinol; 4-carbethoxydiorcinal; cordyol C; methylgerfelin; violaceol II; averythrin; averantin, 1’-O-methylaverantin; lecanoric acid; orsellic acid; orcinol; 1- methylpyrogallol; fumaric acid ABTS = 0.1–5.4 mmol Trolox/g | [67] |
| Cladosporium cladosporioides from Sargassum wightii EtOAc LC-MS | N-(2-Iodophenyl)-2-[2-oxo-5-(thiophen-2-yl)-2,3-dihydro-1,3,4-oxadiazol-3-yl] acetamide; 2-[3-chloro-4-(4-chlorophenoxy)phenyl]-1,3-dioxo-2,3-dihydro-1H-isoindole-5-carboxylic acid; 2-(2,4-dichlorophenyl)-2-oxoethyl 3,4-dihydro-2H-1,5-benzodioxepine-7-carboxylate; 4-bromo-N′-(4-fluoro-1-benzothiophene-2-carbonyl)-1H-pyrrole-2-carbohydrazide; (1R,2R,5S)-2-[3-({2-[(2,4-dichlorophenyl)methyl]-2H-1,2,3,4-tetrazol-5-yl}methyl)-4-methyl-5 sulfanylidene-4,5-dihydro-1H-1,2,4-triazol-1-yl]-6,8-dioxabicyclo [3.2.1]octan-4-one; methyl 2-({[5-bromo-2-(4 methoxybenzamido)phenyl] (phenyl)methyl}amino) acetate; 2-[4-(2,4-dichlorophenoxy)phenyl]-5-phenyl-octahydro-1H-isoindole-1,3-dione; N-({2-[(3,4-dichlorophenyl) methoxy]naphthalen-1-yl}methyl)-2,3-dihydro-1,4-benzodioxin- 6-amine; 2-({[(2,4-dichlorophenyl) carbamoyl] methyl}(propyl)amino)-N-(2,2,2-trifluoroethyl) acetamide; N-(4-bromo-2-fluorophenyl)-6-(2-tert-butylhydrazin1-yl)-5-nitropyrimidin-4-amine; N-(2-{3-[(3,4-di chlorophenyl)methyl]-2-oxo-1,3-diazina EC50,DPPH = 50 mg/mL; EC50,DPPH, AS = 18 mg/mL; RP = 0.81 mg/g | [68] |
| Penicillium brevicompactum EtOAc RP-HPLC, UV, HRESIMS, NMR | Anthranilic acid; syringic acid; sinapic acid; acetosyringone IC50, DPPH = 20–30 µg/mL | [69] |
| Penicillium janthinellum EtOAc, MeOH HPLC, UV, IR, HRESIMS, GC-MS, NMR | 6-(2-Acetyl-3,5-dihydroxybenzyl)-4-hydroxy-3-methyl-2H-pyran-2-one; 7-hydroxy-2-(hydroxymethyl)-5-methyl-4H-chromen4-one; 3,5-dihydroxy-2-(2-(2-hydroxy-6-methylphenyl)-2-oxoethyl)-4-methylbenzaldehyde; 3-hydroxy-5-methylphenyl 2,4-dihydroxy-6-methylbenzoate; lecanoric acid; orsellinic acid; orcinol | [70] |
| Penicillium griseofulvum EtOAc, BuOH HPLC, UV, IR, HRESIMS, NMR, OP | 4,6-Dimethylcurvulinic acid | [64] |
| Penicillium expansum 091006 from the mangrove plant Excoecaria agallocha EtOAc, AC HPLC, UV, IR, HRESIMS, TLC, NMR, OP, ECD | Phenolic bisabolane sesquiterpenoid and diphenyl ether units, expansols A and B, (S)-(+)-11-dehydrosydonic acid,(7S,11S)-(+)-12-acetoxysydonic acid, (S)-(+)-sydonic acid, diorcinol,(S)-(+)-2-[3-hydroxy-4-(2-methoxy-6-methylheptan-2-yl)benzyl]-5- (3-hydroxy-5-methylphenoxy)-3-methylphenol, S-(+)-2-[3-hydroxy-4-(2-hydroxy-6- methylheptan-2-yl)benzyl]-5-(3-hydroxy-5-methylphenoxy)-3-methylphenol, (S)-(+)-3-hydroxy-4-(2-hydroxy-6-methylhept-6-en-2-yl)benzoic acid, and 4-[(2S,6S)-7-acetoxy-2-hydroxy-6-methylheptan-2-yl]-3-hydroxybenzoic acid | [71] |
| Scopulariopsis sp. EtOAc, MeOH, W, H HPLC-PDA, RP-HPLC, LC-MS, HRESIMS, TLC, NMR, OP | 12-Dimethoxypinselin; 12-O-acetyl-AGI-B4, 11,12-dihydroxysydonic acid; 1-hydroxyboivinianic acid | [72] |
| ZSDS1-F11 from the sponge Phakellia fusca EtOAc, AC TLC, CC, HRESIMS, NMR | Phenolic bisabolane sesquiterpenoid and diphenyl ether units; expansols A–F; diorcinol | [73] |
| Cyanobacteria | ||
| Anabaena C5 E, US HPLC-MS/MS | Quinic acid; catechin EC50 DPPH = 0.1 mg/mL; FRAP = 11.4 mg ASE/g | [74] |
| Arthrospira S1, S2 EtOH, US HPLC-MS/MS | Catechin EC50, DPPH = 0.1 mg/mL; FRAP = 15.1–21.0 mg ASE/g | [74] |
| Calothrix sp. SI-SV MeOH, W HPLC-UV/VIS | Rutin; tannic acid; orcinol; phloroglucinol; protocatechuic acid EC50, ABTS = 65.79 μg/mL, EC50, DPPH = 69.38 μg/mL | [75] |
| Leptolyngbya sp. MeOH, W HPLC-UV/VIS | Rutin; tannic acid; orcinol; phloroglucinol; protocatechuic acid SI-SM (EC50, ABTS = 63.45, EC50, DPPH = 67.49 μg/mL) | [75] |
| Nostoc commune MeOH, W RP-HPLC-DAD | Gallic and chlorogenic acids | [76] |
| Nostoc sp EtOH, US HPLC-MS/MS | Gallic acid; chlorogenic acid; quinic acid; catechin; epicatechin; kaempferol; rutin; apiin IC50, DPPH = 0.04–9.47 mg/mL; FRAP = 8.4–13.7 mg ASE/g | [74] |
| Microalgae | ||
| Ankistrodesmus sp. MeOH, W RP-HPLC-DAD | Protocatechuic acid DPPH scavenging10 mg/mL = 29% | [76] |
| Euglena cantabrica MeOH, W RP-HPLC-DAD | Gallic acid; protocatechuic acid; chlorogenic acid; (+) catechin; (−)epicatechin DPPH scavenging10 mg/mL = 71% | [76] |
| Nannochloropsis sp. EtOH, MeOH, H RP-HPLC-UV HPLC–ESI–MS/MS | Phenolic acids: chlorogenic; caffeic; gallic; protocatechuic; hydroxybenzoic; syringic; vanillic; ferulic | [27] |
| Spirulina sp. RP-HPLC-UV HPLC-ESI-MS/MS | Protocatechuic; gallic; chlorogenic; vanillic; hydroxybenzoic; syringic; vanillic acids | [27] |
| Spirogyra sp. MeOH, W RP-HPLC-DAD | Gallic acid DPPH scavenging10 mg/mL = 62% | [76] |
| Seagrasses | ||
| Cymodocea nodosa MeOH, CH2CL2 HPLC-DAD, LC/MS-ESI, NMR | Diosmetin 7-sulfate; caftaric acid; coutaric acid | [77] |
| Halodule wrightii, Thalassia testudinum Agar, W HPLC | p-hydroxybenzoic acid; ferulic acid; p-coumaric acid; syringic acid; gallic acid | [78] |
| Halophila stipulacea MeOH, EtOAc, Hexane HR-LC-MS/MS GNPS | Luteolin; apigenin; matairesinol; cirsimarin; spiraeoside; 2,4-dihydroxyheptadec-16-ynyl acetate; 3-hydroxy-4-methoxycinnamic acid, alpha-cyano-4-hydroxycinnamic | [79] |
| Posidonia oceanica (L.) EtOH, W, Formic acid HPLC-ESI-MS/MS | Procyanidin B2; procyanidin C2; isorhamnetin-3-O-glucoside; quercetin-3-O-glucoside; quercetin-3-O-malonylglucoside; isorhamnetin-3-O-malonylglucoside EC50, DPPH = 32 µg/mL | [80] |
| Ruppia cirrhosa (Petagna) Grande, Ruppia maritima L. MeOH, W, EtOAc HPLC-DAD, HR-LCMS-ESI+/TOF, NMR | Chicoric acid; quercetin 3-O-β-d-(6’’-O-malonyl)-glucopyranoside; quercetin 3-O-β-d-galactopyranoside; quercetin 3-O-β-d-glucopyranoside; quercetin 3-O-β-d-(6′’-O-malonyl)galactopyranoside; isorhamnetin 3-O-β-d-galactopyranoside; isorhamnetin 3-O-β-d-glucopyranoside; isorhamnetin 3-O-β-d-(6´´-O-malonyl) galactopyranoside; isorhamnetin 3-O-β-d-(6′’-O-malonyl)-glucopyranoside EC50 DPPH = 23–176 µg/mL | [81] |
| Syringodium isoetifolium MeOH HPLC-EI-MS | Caftaric acid; 2 3-(4-Hydroxyphenyl)lactic acid; caffeic acid; caffeoyl-4′-O-phenyllactate; 3-phenyllactic acid; 4-coumaric acid; chicoric acid DPPH = 5.4 mg TE/g; ABTS = 9.6 mg TE; CUPRAC = 18.7 mg TE/g; FRAP = 9.5 mgTE/g; Chelating ability = 9.17 mg EDTAE/g | [82] |
| Thalassia testudinum AC, W, AA | 3,4-Dihydroxybenzoic acid; p-hydroxybenzoic acid; p-coumaric acid; vanillin | [83] |
| T. testudinum EtOH, W RP-HPLC LC-MS, NMR | 3,4-Dihydroxybenzoic acid, p-hydroxybenzoic acid, p-coumaric acid and vanillin | [84] |
| Zostera asiatica and Z. marina HPLC-MS | Rosmarinic acid; luteolin; 7,3′-disulfate luteolin ROS scavenger; protecting or enhancing endogenous antioxidants; metal chelation | [85] |
| Z. marina Hexane, AC HPLC-MS, NMR | Deoxycymodienol; isotedarene A | [86] |
| Z. marina SPA, IPA HPLC-MS/MS | 3-Hydroxyhexanoic acid; 4-hydroxynonenoic acid; p-coumaric acid; caffeic acid; ferulic acid; zosteric acid; apigenin; luteolin; diosmetin; apigenin-7-sulfate; rosmarinic acid; luteolin-7-sulfate; diosmetin-7-sulfate; kaempferol-7,4′-dimethylether-3-O-sulfate | [5] |
| Z. noltii MeOH HPLC, NMR | Rosmarinic acid; caffeic acid; zosteric acid | [87] |
| Z. noltei MeOH HPLC-PDA-MS-ESI-QTOF, NMR | Rosmarinic acid; apigenin-7-O-glucoside; luteolin; apigenin; diosmetin; acacetin; luteolin-7-sulfate; apigenin-7-sulfate; diosmetin-7-sulfate; acacetin-7-sulfate | [88] |
| Zostera noltei leaves MeOH, W HPLC-DAD, LC-MS, NMR | Apigenin 7-sulfate; diosmetin 7-sulfate | [89] |
| Zostera noltii, Z. marina | Apigenin 7-sulphate; luteolin 7-sulphate; diosmetin 7-sulphate; rosmarinic acid; luteolin 7-glucoside; apigenin 7-glucoside; apigenin; luteolin 7-(6′’-malonyl) glucoside; apigenin 7-(6′’-malonyl) glucoside | [81] |
| Zostera muelleri MeOH, AA RP-HPLC | Proanthocyanidins; gallic acid; rosmarinic acid | [90] |
| Sponges | ||
| Didiscus aceratus MeOH, CH2Cl2, H HRESIMS, NMR | (S)-(+)-Curcuphenol; 10β-hydroxycurcudiol; 10α-hydroxycurcudiol; dicurcuphenols A–E; dicurcuphenol ether F | [91] |
| Hyrtios erectus MeOH, EtOAc HRAPCIMS, HRESIMS, NMR | Phenolic alkenes; erectuseneols A−F | [92] |
| Myrmekioderma sp. MeOH, CH2Cl2, EtOAc, BuOH, hexane HRESIMS, NMR | 1-(2,4-Dihydroxy-5-methylphenyl)ethan-1-one; (1′Z)-2-(1′,5′-dimethylhexa-1′,4′-dieny1)-5- methylbenzene-1,4-diol; 1,8-epoxy-1(6),2,4,7,10-bisaborapentaen-4-ol; 6-(3-hydroxy-6-methyl-1,5- heptadien-2-yl)-3-methylbenzene-1,4-diol; 4-hydroxy-3,7-dimethyl-7-(3-methylbut-2- en-1-yl)benzofuran-15-one; 6-(2-methoxy-6-methylhept-5-en-2-yl)-3-methylbenzene-1,4-diol; 9-(3,3-dimethyloxiran-2-yl)-1,7-dimethyl-7-chromen-4-ol | [93] |
| Myrmekioderma sp. MeOH, EtOAc HRAPCIMS, HRESIMS, NMR | (R)-Biscurcudiol; (S)-biscurcudiol; myrmekiodermaral; myrmekioperoxide A; myrmekioperoxide B (4); myrmekiodermaral; (+)-curcudiol; (+)-dehydrocurcudiol; abolene; abolene epimer at C-5′; (+)-oxoabolene; (+)-curcuphenol; 5′α-hydroxycurcudiol; 5′β- hydroxycurcudiol; curcuepoxide A; curcuepoxide B | [94] |
2.2.3. Macroalgae
Bromophenols
Simple Phenolics
| Seaweed Extraction Chemical Analysis | Compounds Antioxidant Activity (When Provided) | Ref. |
|---|---|---|
| A. nodosum, F. spiralis MeOH, AC, Hexane UPLC, MS, NMR | Phlorotannins (4–6, 9–12 PGU) | [107] |
| A. nodosum, Fucus spiralis, F. vesiculosus, Pelvetia canaliculata, Saccharina longicruris MeOH, W UPLC, HRMS | Phlorotannins (3–50 PGU) | [108] |
| Carpophyllum flexuosum, Carpophyllum plumosum, Ecklonia radiata W, MAE HPLC-DAD-ESI-MS, NMR | Bifuhalol, bifuhalol dimer, bifuhalol trimer, hydroxytrifuhalol, trifuhalol, tetrafuhalol DPPH = 2.7–37.4 mg GAE/g; FRAP = 4.4–62.1 mg GAE/g | [109] |
| Cystoseira barbata TFA, W LC-QTOF-MS | Phloroglucinol, rutin, phlorofucofuroeckol, 3-O-rutinosyl-kaempferol, catechin-catechin-O-gallate, gallocatechin, gallocatechin-O-glucuronide, 1-hydroxy-2-(β-d-glucopyranosyloxy)-9,10 anthraquinone, 2-O-(6,9,12-octadecatrienoyl)-3-O-(nonadecanoyl)glyceryl β-galactopyranoside, chlorogenic acid butyl ester, phloroglucinol, quercetin EC50, DPPH = 11.7 µg/mL; EC50, OH = 11.4 µg/mL; EC50, RP = 51 g/mL; EC50, CA = 40 g/mL | [110] |
| C. barbata AC, MeOH, W UHPLC-DAD-QTOF-MS | Fucophlorethol and eckol derivatives (3–7 PGU) EC50, DPPH = 14 µg/mL; EC50, ABTS = 0.5 µM Trolox; EC50, RP = 16–35 µg/mL | [111] |
| Cystoseira nodicaulis, Cystoseira tamariscifolia, Cystoseira usneoides, F. spiralis AC, Hexane, W HPLC-DAD-ESI-MSn | Fucophloroethol, fucodiphloroethol, fucotriphloroethol, 7-phloroeckol, phlorofucofuroeckol, bieckol, dieckol EC50, SRSA = 0.93–4.02 mg/mL; EC50, LPIA = 2.32–>9.1 mg/mL | [96] |
| Cystoseira nodicaulis, F. serratus, F. vesiculosus, Himanthalia elongata EtOH, W UPLC-ESI-MS | Phlorotannin (3–16 PGU) EC50, DPPH = 4–28 μg/mL; FRAP = 101–307 μg TE/mg | [21] |
| Durvillaea antarctica, Lessonia spicata EtOH, EE, EtOAc, W HPLC-MS-MRM | Phlorotannin (3–8 PGU), flavonoids EC50, DPPH = 0.97–1.24 mg/mL; FRAP = 2.95–6.20 mM TE/kg; ORAC = 4.75–25.9 μM TE/g | [112] |
| Eisenia bicyclis EtOH HPLC-PDA | Eckol, phlorofucofuroeckol-A, dieckol, 6,6′-bieckol, 8,8′-bieckol | [113] |
| Ecklonia cava EtOH UPLC-PDA | Phloroglucinol, eckol, eckstolonol, triphlorethol-A, dieckol | [114] |
| E. cava EtOH, US HPLC-DAD-ESI/MS, NMR | Dieckol, phlorofucofuroeckol-A, 2,7-phloroglucinol-6,6-bieckol, pyrogallol-phloroglucinol-6,6-bieckol | [115] |
| E. cava EtOH, W RP-HPLC | Dieckol ABTS = 1.3 g VCE/g; DPPH = 0.4 g VCE/g | [116] |
| Ecklonia stolonifera EtOH, W HPLC-PDA, NMR | 2-phloroeckol, dioxinodehydroeckol, eckol, phlorofucofuroeckol B, 6,6’-bieckol, dieckol, 974-B, phlorofucofuroeckol A | [117] |
| F. vesiculosus MeOH, W Q-ToF-MS, UPLC-TQD-MS/MS-MRM | Phlorotannins (3–18 PGU) EC50, DPPH = 18.2 μg/mL | [118] |
| F. vesiculosus AC, EtOAc, EtOH, MeOH, W HPLC-DAD-ESI/MSn | Fucodiphlorethol A, trifucodiphlorethol isomers, phlorotannins (3–10 PGU) EC50, DPPH = 2.79–4.23 μg/mL; Fe2+-CA = 25.1–47.6%; RP = 17.8–910.7 mg ASEs/g | [119] |
| F. vesiculosus AC, W UPLC-DAD-ESI/MSn | Fucols, fucophlorethols, fuhalols, phlorotannin derivatives (3–22 PGU), fucofurodiphlorethol, fucofurotriphlorethol, fucofuropentaphlorethol | [120] |
| Halidrys siliquosa AC, W MALDI-TOF-MS, NMR | Diphlorethol, triphlorethol, trifuhalol, tetrafuhalol EC50, DPPH = 0.02–1.00 mg/mL; EC50, RP = 0.06–0.62 mg/mL; EC50, NBT = 0.66–2.44 mg/mL; ORAC = 5.39 μmol TE/mg; BCB = 0.21–1.50 g/mL | [121] |
| H. elongata MeOH, W HPLC-DAD, HPLC-ESI-MS/MS | Phloroglucinol, gallic acid, chlorogenic acid, caffeic acid, ferulic acid, hydroxybenzaldehyde, kaempferol, myricetin, quercetin EC50, DPPH = 14.5 μg/mL | [20] |
| Hydroclathrus clathratus, Padina minor, Padina sp., Sargassum oligosystum, Sargassum aff. bataanens, Sargassum sp. MeOH, W GC-MS-EI-SIM | 2,4,6-tribromophenol; 2,4,6-tribromoanisol; 2′-hydroxy-2,3′,4,5′- tetrabromodiphenyl ether; 2’-methoxy-2,3′,4,5′-tetrabromodiphenyl ether; 6-hydroxy-2,2′,4,4′-tetrabromodiphenyl ether; 6-methoxy-2,2′,4,4′- tetrabromodiphenyl ether; 2′,6-dihydroxy-2,3′,4,5′-tetrabomodiphenyl ether; 2′,6-dimethoxy-2,3′,4,5′-tetrabromodiphenyl ether; 2,2′-dihydroxy- 3,3′,5,5′-tetrabromodiphenyl; 2,2′-dimethoxy-3,3′,5,5′-tetrabromodiphenyl | [122] |
| L. digitata MeOH, W RP-UPLC-UV-MSn, MALDI-TOF-MS, NMR | Di–fuhalols (6–7 PGU), fucols (3–7 PGU), fucophlorethols (3–16 PGU), fuhalols (4–5 PGU), phlorethols (3–18 PGU) | [123] |
| Leathesia nana CH2Cl2, EtOH, W HRESIMS, NMR | 2,2′,3,3′-tetrabromo-4,4′,5,5′-tetrahydroxydiphenylmethane; 3-bromo-4- (2,3-dibromo-4,5-dihydroxybenzyl)-5-methoxymethylpyrocatechol; 2,3,3′-tribromo-4,4′,5,5′-tetrahydroxyl-1′-ethyloxymethyldiphenyl methane; 2,3-dibromo-4,5-dihydroxybenzaldehyde; 2,3-dibromo-4,5- dihydroxybenzyl alcohol; 2,3-dibromo-4,5-dihydroxybenzyl methyl ether; 2,3-dibromo- 4,5-dihydroxybenzyl ethyl ether; 3,5-dibromo-4-hydroxybenzaldehyde; 3,5-dibromo-4-hydroxybenzoic acid; 3-bromo-4,5-dihydroxybenzoic acid methyl ester, 3-bromo-5-hydroxy-4-methoxybenzoic acid, 3-bromo-4-hydroxybenzoic acid EC50, DPPH = 14.5 µg/mL | [124] |
| Lessonia trabeculate MeOH, W, MWE HPLC-DAD-ESI-MS/MS | Phlorotannins derivatives (2–3 PGU), gallocatechin derivative, p-coumaric acid derivative | [103] |
| Padina tetrastromatica PE, MeOH, CHCl3, MEK, Soxhlet HPLC-UV, UPLC-MS/MS | Fucophlorethol (2–18 PGU) EC50, DPPH = 17 µg/mL | [125] |
| Sargassum fusiforme EtOH, W UPLC-DAD-ESI-MS/MS | Eckol, dieckol, dioxinodehydroeckol, fuhalols (2–12 PGU), phlorethols/fucols/fucophlorethols (2–11 PGU), eckols (2–8 PGU) EC50, DPPH = 15–150 µg/mL; FRAP = 1.29 mg TE/mg | [126] |
| Sargassum muticum EtOH, W, PHLE LC × LC-DAD-ESI-MS/MS | Decafuhalol, dihydroxytetrafuhalol, dihydroxypentafuhalol, dihydroxyhexafuhalol, dihydroxyheptafuhalol, dihydroxyoctafuhalol, dihydroxynonafuhalol, heptaphlorethol, hexafuhalol, hexaphlorethol, hydroxytetrafuhalol, hydroxypentafuhalol, hydroxyhexafuhalol, nonafuhalol, octafuhalol, pentafuhalol, pentaphlorethol, tetrafuhalol, trifuhalol, trihydroxyhexafuhalol, trihydroxyheptafuhalol, trihydroxyoctafuhalol ABTS = 0.65–2.29 mmol TE/g | [127] |
| Silvetia compressa EtOH, US HPLC-DAD, HPLC-TOF-MS | Dihydroxytetrafuhalol, dieckol, eckol derivative, eckstolonol, 7-phloroeckol (3 PGU), dihydroxypentafuhalol, phlorofucofuroeckol A, pentafuhalol, trifuhalol | [128] |
Phlorotannins
| Seaweed Extraction Chemical Analysis | Compounds Antioxidant Activity (When Provided) | Ref. |
|---|---|---|
| Green seaweeds | ||
| Caulerpa lentillifera, C. taxifolia, Chaetomorpha crassa, Chara sp., Chlorodesmis sp., Cladophora sp. MeOH, W GC-MS-EI-SIM | 2,4,6-Tribromophenol; 2,4,6-tribromoanisol; 2′-hydroxy-2,3′,4,5′-tetrabromodiphenyl ether; 2’-methoxy-2,3′,4,5′-tetrabromodiphenyl ether; 6-hydroxy-2,2′,4,4′-tetrabromodiphenyl ether; 6-methoxy-2,2′,4,4′-tetrabromodiphenyl ether; 2′,6-dihydroxy-2,3′,4,5′-tetrabromodiphenyl ether; 2′,6-dimethoxy-2,3′,4,5′-tetrabromodiphenyl ether; 2,2′-dihydroxy-3,3′,5,5′-tetrabromodiphenyl; 2,2′-dimethoxy-3,3′,5,5′-tetrabromodiphenyl | [122] |
| Dasycladus vermicularis MeOH UPLC-MS/MS | 4-(Sulfooxy)phenylacetic acid; 4-(sulfooxy)benzoic acid | [133] |
| Red seaweeds | ||
| Acanthophora specifera, Ceratodictyon spongiosum, Gracilaria edulis, Hydropuntia edulis, Halymenia sp., Jania adhaeren, Jania sp., Kappaphycus alvarezii MeOH, W GC-MS-EI-SIM | 2,4,6-Tribromophenol; 2,4,6-tribromoanisol; 2′-hydroxy-2,3′,4,5′-tetrabromodiphenyl ether; 2’-methoxy-2,3′,4,5′-tetrabromodiphenyl ether; 6-hydroxy-2,2′,4,4′-tetrabromodiphenyl ether; 6-methoxy-2,2′,4,4′-tetrabromodiphenyl ether; 2′,6-dihydroxy-2,3′,4,5′-tetrabromodiphenyl ether; 2′,6-dimethoxy-2,3′,4,5′-tetrabromodiphenyl ether; 2,2′-dihydroxy-3,3′,5,5′-tetrabromodiphenyl; 2,2′-dimethoxy-3,3′,5,5′-tetrabomodiphenyl | [122] |
| Asparagopsis taxiformis W, MeOH, CH2Cl2, H GC-MS | Bromoform, dibromochloromethane, bromochloroacetic acid, dibromoacetic acid | [100] |
| Bostrychia radicans MeOH, H, EtOAc GC-MS, NMR | N,4-dihydroxy-N-(2′-hydroxyethyl)-benzamide; N,4-dihydroxy-N-(2′-hydroxyethyl)-benzeneacetamide; methyl 4-hydroxymandelate; methyl 2-hydroxy-3-(4-hydroxyphenyl)-propanoate | [134] |
| C. tenuicorne H + DEt + 2-P GC-MS, ECNI | Phenols, hydroxylated, and methoxylated penta- and hexabrominated diphenyl ethers | [135] |
| C. tenuicorne H + DEt + 2-P GC-MS, ECNI | Hydroxylated polybrominated diphenyl ethers 2′-hydroxy-2,3′,4,5′-tetrabromodiphenyl ether; 6-hydroxy-2,2′,4,4′-tetrabromodiphenyl ether | [136] |
| Laurencia nipponica, Odonthalia corymbifera, Polysiphonia morrowii A, W, MeOH LC-MS, NMR | 3,5-Dibromo-4-hydroxybenzaldehyde; 3-bromo-4,5-dihydroxybenzyl ether; 3-bromo-4,5-dihydroxybenzyl alcohol; 5-((2,3-dibromo-4,5-dihydroxybenzyloxy)methyl)-3,4-dibromobenzene-1,2-diol; 5-(2-bromo-3,4-dihydroxy-6-(hydroxymethyl) benzyl)-3,4-dibromobenzene-1,2-diol | [137] |
| O. corymbifera, Neorhodomela aculeata, Symphyocladia latiuscula A, W, MeOH LC-MS, NMR | n-Butyl 2,3-dibromo-4,5-dihydroxybenzyl ether; 3-bromo-4-(2,3-dibromo-4,5-dihydroxybenzyl)-5-methoxymethylpyrocatechol; 2,3-dibromo-4,5- dihydroxybenzyl alcohol; 2,3-dibromo-4,5-dihydroxybenzyl methyl ether; bis-(2,3,6-tribromo-4,5-dihydroxybenzyl) ether; 2,3,6-tribromo-4,5-dihydroxybenzyl methyl ether; 2,2′,3,3′-tetrabromo-4,4′,5,5′-tetrahydroxydiphenylmethane; 5-(2-bromo-3,4-dihydroxy-6-(hydroxymethyl)benzyl)-3,4-dibromobenzene- 1,2-diol; 5-((2,3-dibromo-4,5-dihydroxybenzyloxy)methyl)-3,4-dibromobenzene-1,2-diol | [138] |
| Odonthalia corymbifera MeOH, EtOAc NMR | Odonthalol, odonthadione | [139] |
| Polysiphonia decipiens 3:1 MeOH:CH2Cl2 NMR | α-O-Methyllanosol; lanosol; 5-(2-bromo-3,4-dihydroxy-6-(hydroxymethyl)benzyl)-3,4-dibromobenzene-1, 2-diol; 5-(2-bromo-3, 4-dihydroxy-6-(methoxymethyl)benzyl)-3, 4-dibromobenzene-1, 2-diol; rhodomelol; polysiphonol | [140] |
| Polysiphonia morrowii W, MeOH, CH2Cl2 NMR | 3-bromo-4,5-dihydroxybenzyl methyl ether; 3-bromo-4,5-dihydroxybenzaldehyde | [141] |
| P. morrowii W, MeOH ESI-MS, NMR | bis (3-Bromo-4,5-dihydroxybenzyl) ether | [142] |
| Rhodomela confervoides EtOH NMR | 3-(2,3-Dibromo-4,5-dihydroxybenzyl) pyrrolidine-2,5-dione; methyl 4-(2,3-dibromo-4,5-dihydroxybenzylamino)-4-oxobutanoate; 4-(2,3-dibromo-4,5-dihydroxybenzylamino)-4-oxobutanoic acid; 3-bromo-5-hydroxy-4-methoxybenzamide; 2-(3-bromo-5-hydroxy-4-methoxyphenyl)acetamide; 3-bromo-4,5-bis(2,3-dibromo-4,5-dihydroxybenzyl) pyrocatechol; methyl 1-(2-(2,3-dibromo-4,5-dihydroxybenzyl)-3-bromo-4,5- dihydroxybenzyl)-5-oxopyrrolidine-2-carboxylate; 5-((2,3-dibromo-4,5-dihydroxybenzyloxy)methyl)-3,4-dibromobenzene-1,2-diol; 5-(2-bromo-3,4-dihydroxy-6-(hydroxymethyl)benzyl)-3,4-dibromobenzene-1,2-diol; 5-(2-bromo-3,4-dihydroxy-6-(methoxymethyl) benzyl)-3,4-dibromobenzene-1,2-diol; 5-(2-bromo-6-(ethoxymethyl)-3,4-dihydroxybenzyl)-3,4-dibromobenzene-1,2-diol; 5-(2,3-dibromo-4,5-dihydroxybenzyl)- 3,4-dibromobenzene-1,2-diol; 1-(2,3-dibromo-4,5-dihydroxybenzyl)-5-oxopyrrolidine-2-carboxylic acid; methyl 1-(2,3-dibromo-4,5-dihydroxybenzyl)-5-oxopyrrolidine-2-carboxylate EC50, DPPH = 5.2–23.6 µmol/L; ABTS = 2.1–3.6 mmol TE/L | [143] |
| Symphyocladia latiuscula EtOH NMR | 2,3-Dibromo-4,5-dihydroxybenzyl methyl ether, 3,5-dibromo-4-hydroxybenzoic acid; 2,3,6-tribromo-4,5-dihydroxymethylbenzene; 2,3,6-tribromo-4,5-dihydroxybenzaldehyde; 2,3,6-tribromo-4,5-dihydroxybenzyl methyl ether; bis(2,3,6-tribromo-4,5-dihydroxyphenyl)methane; 1,2-bis(2,3,6-tribromo-4,5-dihydroxyphenyl)-ethane; 1-(2,3,6-tribromo-4,5-dihydroxybenzyl)-pyrrolidin-2-one | [144] |
| S. latiuscula EtOH, EtOAc, W HRMS, NMR, MS | 1-[2,5-dibromo-3,4-dihydroxy-6-(2,3,6-tribromo-4,5-dihydroxybenzyl)benzyl]pyrrolidin-2-one; methyl 4-{(2,3,6-tribromo-4,5-dihydroxybenzyl)[(2,3,6-tribromo-4,5-dihydroxybenzyl)carbamoyl] amino}butanoate; methyl 4-{(2,5-dibromo-3,4-dihydroxybenzyl)[(2,3,6-tribromo-4,5-dihydroxybenzyl)carbamoyl]amino} butanoate; 2,5-dibromo-3,4-dihydroxy-6-(2,3,6-tribromo-4,5-dihydroxybenzyl)benzyl methyl ether EC50, DPPH = 14.5, 20.5 µg/L | [145,146] |
| S. latiuscula EtOH, EtOAc, W ESIMS, NMR | Bromocatechol conjugates (symphyocladins) | [147,148] |
| S. latiuscula MeOH, CH2Cl2, EtOAc, BuOH, W ESIMS, NMR | 2,3,6-tribromo-4,5-dihydroxybenzyl alcohol; 2,3,6-tribromo-4,5-dihydroxybenzyl methyl ether; bis-(2,3,6-tribromo-4,5-dihydroxybenzyl) ether | [149] |
| Vertebrata lanosa MeOH, EtOAc ESIMS, NMR | 2,3-Dibromo-4,5-dihydroxybenzylaldehyde; 2,2′,3-tribromo-3′,4,4′,5-tetrahydroxy-6′-hydroxymethyldiphenylmethane; bis(2, 3-dibromo-4,5-dihydroxylbenzyl) ether; 5,5″-oxybis(methylene)bis (3-bromo-4-(2′,3′-dibromo-4′,5′-dihydroxylbenzyl)benzene-1,2-diol) ORAC = 0.08–0.33 μg TE/mL; CAAinhib. 10 μg/mL = 68%; CLPAAinhib. 10 μg/mL = 100% | [95] |
| V. lanosa | Methylrhodomelol; lanosol; lanosol methyl ether; 2-amino-5-(3-(2,3-dibromo-4,5-dihydroxybenzyl)ureido)pentanoic acid; 3-bromo-4-(2,3-dibromo-4,5-dihydroxybenzyl)-5-methoxymethylpyrocatechol; 5-((2,3-dibromo-4,5-dihydroxybenzyloxy)methyl)-3,4-dibromobenzene-1,2-diol; 2,2′,3,3′-tetrabromo-4,4′,5,5′-tetrahydroxydiphenylmethane | [150] |
2.2.4. Seagrasses
2.2.5. Sponges
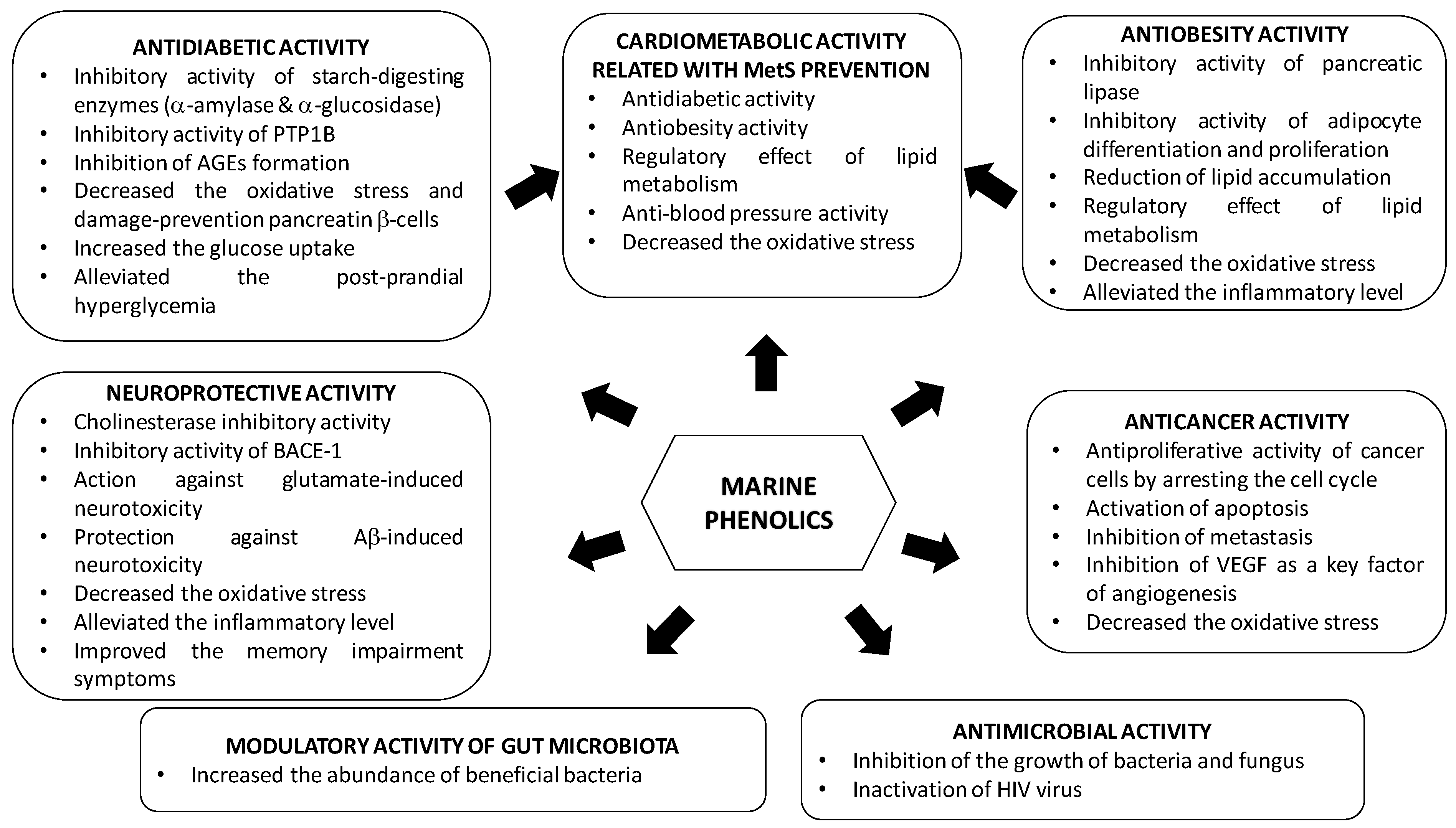
3. Bioactive Properties of Marine Phenolics
3.1. Type 2 Diabetes Mellitus
| Compounds/Marine Source | Test Model | Outcome | Ref. |
|---|---|---|---|
| Five isolated phlorotannins from E. cava (fucodiphloroethol G, dieckol, 6,6′-bieckol, 7-phloroeckol, phlorofucofuroeckol-A) | In vitro assay: α-glucosidase and α-amylase inhibitory activity | Inhibition of α-glucosidase (IC50 values ranged from 10.8 μM for dieckol to 49.5 μM for 7-phloroeckol) and α-amylase (IC50 values ranged from 125 μM for dieckol to <500 μM for the rest of compounds, except 7-phloroeckol with a value of 250 μM) activities | [165] |
| Methanolic extract isolated from A. nodosum rich in phlorotannins | In vitro assay: α-glucosidase and α-amylase inhibitory activity | Inhibition of α- glucosidase (IC50~20 μg/mL GAE) and α-amylase (IC50~0.1 μg/mL GAE) activities | [166] |
| Cold aqueous and ethanolic extracts of A. nodosum and F. vesiculosus rich in phlorotannins | In vitro assay: α-glucosidase and α-amylase inhibitory activity | Inhibition of α- glucosidase (IC50~0.32–0.50 μg/mL GAE for F. vesiculosus) and α-amylase (IC50~44.7–53.6 μg/mL GAE for A. nodosum) activities | [167] |
| Methanolic extract from Alaria marginata and Fucus distichus rich in phlorotannins | In vitro assay: α-glucosidase and α-amylase inhibitory activity | Inhibition of α- glucosidase (IC50~0.89 μg/mL) and α-amylase (IC50~13.9 μg/mL) activities | [168] |
| Polyphenol-rich extracts from L. trabeculate | In vitro assay: α-glucosidase and lipase activity | Inhibition of α-glucosidase and lipase activities (IC50 < 0.25 mg/mL) | [103] |
| Crude extract and semi-purified phlorotannins from F. vesiculosus composed by fucols, fucophlorethols, fuhalols and several other phlorotannin derivatives | In vitro assay: α-glucosidase, α-amylase and pancreatic lipase inhibitory activity | Inhibition of α-amylase (IC50~28.8–2.8 μg/mL), α-glucosidase (IC50~4.5–0.82 μg/mL) and pancreatic lipase (IC50~45.9–19.0 μg/mL) activities | [120] |
| Phlorotannin derivatives from E. cava | In vitro assay: α-glucosidase inhibitory activity | Inhibition of α-glucosidase activity (IC50~2.3–59.8 μM) Kinetic parameters of receptor–ligand binding | [163] |
| Phlorotannin-targeted extracts from four edible Fucus species (F. guiryi, F. serratus, F. spiralis and F. vesiculosus) | In vitro assay: α-glucosidase and α-amylase inhibitory activity | Inhibition of α-glucosidase (IC50~2.48–4.77 μg/mL), α-amylase (IC50~23.31–253.31 μg/mL) and xanthine oxidase (IC50~157.66–800.08 μg/mL) activities | [8] |
| Marine-derived bromophenol compound (3,4-dibromo-5-(2-bromo-3,4-dihydroxy-6-(ethoxymethyl)benzyl)benzene-1,2-diol) isolated from Rhodomela confervoides | In vitro: insulin resistant C2C12 cells treated with bromophenol (0.1–0.5 μM for phenol) | Inhibition of PTP1B activity (IC50~0.84 μM) Activation of insulin signaling and potentiate insulin sensitivity | [172] |
| 3-Bromo-4,5-bis(2,3-dibromo-4,5-dihydroxybenzyl)-1,2-benzenediol isolated from the red alga Rhodomela confervoides | In vitro: palmitate-induced insulin resistance in C2C12 cells treated with bromophenol (0.5–2.0 μM for phenol) | Inhibition of PTP1B activity (IC50~2 μM) Activation of insulin signaling and prevent palmitate-induced insulin resistance | [173] |
| Phlorofucofuroeckol-A, eckol, phloroglucinol, fucofuroeckol A, dieckol and 8,8′-bieckol isolated and crude phlorotannins from Lessoniaceae | In vitro assay: human and bovine serum albumin models | Inhibition of AGEs formation, crude phlorotannins showed IC50~0.43–0.53 mg/mL, and among the purified phlorotannins, phlorofucofuroeckol A was the most active (IC50~4.1–4.8 μM) | [177] |
| Methanolic extract from P. pavonica and Turbinaria ornate rich in phlorotannins | In vitro assay: BSA-glucose assay In vivo: Caenorhabditis elegans with induced hyperglycemia | Inhibition of AGEs formation (IC50~15.16 μg/mL, 35.25 μg/mL and 22.70 μg/mL, respectively) Inhibition of AGEs formation | [178] |
| Phlorofucofuroeckol-A isolated from E. stolonifera | In vitro assay for non-enzymatic insulin glycation | Inhibition of AGEs formation (IC50 29.50–43.55 μM for D-ribose and D-glucose-induced insulin glycation, respectively) | [179] |
| Octaphlorethol A isolated from Ishige foliacea | In vitro: STZ-induced pancreatic β-cell damage (RINm5F pancreatic β-cells) (12.5–50.0 μg/mL for phenol) | Decreased the death of STZ-treated pancreatic β-cells Decreased the TBARS and ROS Increased the activity of antioxidant enzymes | [181] |
| 6,6-Bieckol, phloroeckol, dieckol and phlorofucofuroeckol isolated from E. cava | In vivo: high glucose-stimulated oxidative stress in Zebrafish, a vertebrate model (10–20 μM of phenols) | Inhibition of high glucose-induced ROS and cell death Dieckol reduced the heart rates, ROS, NO and lipid peroxidation Dieckol reduced the overexpression of iNOS and COX-2 | [182] |
| Extract isolated from the red seaweed Polysiphonia japonica | In vitro: palmitate-induced damage in β-cells (Ins-1 cells) (1–10 μg/mL of extract) | Inhibited the palmitate-induced damage in β-cells Preserved the glucose-induced insulin secretion in β-cells | [183] |
| Octaphlorethol A from Ishige foliacea | In vitro: rat myoblast L6 cells (6.25–50 μM of phenol) | Increased the glucose uptake Increased the Glut4 translocation to the plasma membrane, via Akt and AMPK activation | [185] |
| Dieckol isolated from E. cava | In vivo: STZ-induced diabetic mice (acute, 100 mg/kg bw of dieckol administered orally) | Delayed the absorption of dietary carbohydrates | [187] |
| 2,7’’-Phloroglucinol-6,6’-bieckol from E. cava | In vivo: STZ-induced diabetic mice (acute, 10 mg/kg bw of phenol administered orally) | Delayed the absorption of dietary carbohydrates Inhibition of α-glucosidase and α-amylase activities (IC50 23.35 μM and 6.94 μM, respectively) | [188] |
| Polyphenol-rich seaweed extract from F. vesiculosus | In vivo: 38 healthy adults (acute, 500 mg and 2000 mg of phenol) | No change in postprandial blood glucose and insulin levels | [189] |
| Dieckol isolated from brown seaweed E. cava | In vivo: a T2DM mouse model (C57BL/KsJ-db/db) (10 and 20 mg/kg bw of phenol for 14 days administered intraperitoneal injection) | Diminished the fasting blood glucose and insulin levels Diminished the body weight Decreased the TBARS Increased the activity of antioxidant enzymes in liver tissues Increased the levels of AMPK and Akt phosphorylation in muscle tissues | [190] |
| Polyphenol-rich extracts from brown macroalgae L. trabeculata | In vitro assay: α-glucosidase and lipase inhibitory activities --- In vivo: high-fat diet and STZ-induced diabetic rats (200 mg/kg/day bw of phenol for 4 weeks by gavage) | Inhibition of α-glucosidase and lipase activities (IC50 < 0.25 mg/mL) --- Diminished the fasting blood glucose and insulin levels Improved the serum lipid profile Improved the antioxidant stress parameters | [103] |
| Water-ethanolic extract of green macroalgae Enteromorpha prolifera rich in flavonoids | In vivo: STZ-induced diabetic rats (150 mg/kg/day bw of phenol for 4 weeks by gavage) | Diminished the fasting blood glucose and improved oral glucose tolerance Hypoglycemic effect by increasing IRS1/PI3K/Akt and suppressing JNK1/2 in liver | [191] |
| Dieckol-rich extract of brown algae E. cava | In vivo: 8 pre-diabetic adults (1500 mg per day for 12 weeks) | Decreased the postprandial glucose, insulin, and C-peptide levels | [192] |
3.2. Obesity
| Compounds/Marine Source | Test Model | Outcome | Ref. |
|---|---|---|---|
| Methanolic extract of E. bicyclis (eckol, fucofuroeckol A, 7-phloroeckol, dioxindehydroeckol, phlorofucofuroeckol A, and dieckol) | In vitro: assay of pancreatic lipase activity | Inhibition of pancreatic lipase activity; fucofuroeckol A and 7-phloroeckol were the most potent (IC50 values of 37.2 and 12.7 μM, respectively) | [198] |
| Polyphenol-rich extract (crude) from the edible seaweed A. nodosum and phlorotannin-enriched fraction from crude extract | In vitro: assay of pancreatic lipase activity | Inhibition of pancreatic lipase activity Evaluated the interaction between phlorotannins and polysaccharides on inhibitory lipase activity and phlorotannins were more effective | [199] |
| Dieckol isolated from E. cava | In vitro: 3T3-L1 pre-adipocytes cells (25–100 μM of phenol) | Suppression of pre-adipocytes differentiation Down-regulated the expression of PPARγ, C/EBPα, SREBP1 and FABP4 by AMPK activation | [201] |
| Phloroglucinol, eckol, dieckol, dioxinodehydroeckol, and phlorofucofuroeckol A isolated from E. stolonifera | In vitro: 3T3-L1 pre-adipocytes cells (12.5–100 μM of phenol) | Suppression of pre-adipocytes differentiation Down-regulated the expression of PPARγ and C/EBPα | [202] |
| 6,6′-Bieckol, 6,8′-bieckol, 8,8′-bieckol, dieckol and phlorofucofuroeckol A isolated from E. bicyclis | In vitro: 3T3-L1 pre-adipocytes cells (10–50 μg/mL of phenol) | Suppression of pre-adipocytes differentiation Down-regulated the expression of PPARγ, C/EBPα, SREBP-1c, FAS and ACC | [195] |
| Triphlorethol-A, eckol and dieckol from E. cava | In vitro: 3T3-L1 pre-adipocytes cells (5 μM of phenol) | Increased the glycerol secretion and reduced glucose consumption level Down-regulated the expression of PPARγ, C/EBPα, SREBP-1c as well as FABP4, FATP1, FAS, leptin and ACSL1 | [204] |
| Extract from E. cava containing eckol, dieckol and phlorofucofuroeckol-A | In vitro: 3T3-L1 adipocytes cells (50 μg/mL of extract) | Inhibited the glucose utilization and TG accumulation Down-regulated the expression of C/EBPα, SREBP-1c, A-FABP, FAS and adiponectin | [207] |
| Triphlorethol-A, eckol and dieckol isolated from E. cava | In vitro: 3T3-L1 pre-adipocytes cells (1–20 μM of phenol) --- In vitro: MC3T3-E1 cells (1–20 μM of phenol) | Suppressed the lipid accumulation and expression of adipogenic differentiation markers --- Enhanced the osteoblast differentiation by increasing alkaline phosphatase activity and raising intracellular calcification | [208] |
| Dieckol from E. cava | In vitro: 3T3-L1 pre-adipocytes cells (25–100 μM of phenol) --- In vivo: high-fat diet-fed zebrafish (1–4 μM of phenol) --- In vivo: high-fat diet-fed mice (15 mg/kg bw/day or 60 mg/kg bw/day, 11 weeks administered orally) | Suppressed the lipid accumulation in the three models Inhibited the early adipogenic events by suppressing cell cycle progression Regulated the AMPKα, ERK, and Akt signaling to inhibit lipid accumulation | [209] |
| 5-Bromo-3,4-dihydroxybenzaldehyde isolated from Polysiphonia morrow | In vitro: 3T3-L1 pre-adipocytes cells (25–100 μM) | Inhibited the intracellular lipid accumulation and triglyceride levels Down-regulated the expression of PPARγ, C/EBPα, SREBP-1, FABP4, FAS, leptin, and adiponectin by AMPK and ACC activation. | [210] |
| Phlorotannin fraction from Fucus distichus | In vitro: murine macrophage RAW 264.7 cells (12.5–50 μg/mL of extract) --- In vitro: 3T3-L1 adipocytes cells (12.5–50 μg/mL of extract) | Anti-inflammatory activity via TLR attenuation in macrophages -- Decreased the lipid accumulation in 3T3-L1 adipocytes cells | [211] |
| Pyrogallol-phloroglucinol-6,6-bieckol from E. cava | In vivo: mouse model of diet-induced obesity (2.5mg/kg bw/day for 4 weeks administered orally) | Reduced the expression of RAGE and the secretion of ligands Reduced the inflammatory cytokine level (TNFα and IL-1β) | [213] |
| Polyphenol extracts from E. cava and dieckol | In vivo: high-fat diet-induced obese mice (5.0 mg, 2.5 mg and 1.25 mg extract/mouse; 2.0 mg, 1.0 mg and 0.5 mg dieckol/mouse for 4 weeks administered orally) --- In vitro: 3T3-L1 pre-adipocytes cells | Suppressed the body weight gain Reduced the TC, TG and LDL-C levels ---- Inhibited the lipid accumulation Inhibition of HMGCoA reductase activity | [214] |
| Polyphenol-rich fraction of E. cava from Gijang (Korea) | In vivo: high-fat diet-induced obese mice (200 mg/kg bw for 8 weeks by oral intubation) | Reduced the body weight gain, body fat and hyperglycemia Reduced the mRNA expression of inflammatory cytokines (TNF-α and IL-1β) and macrophage marker gene (F4/80) | [215] |
| Polyphenol-rich fraction of E. cava containing dieckol, 2,7”-phloroglucino-6,6′-bieckol, pyrogallo-phloroglucinol-6,6′-bieckol and phlorofucofuro-eckol A | In vivo: high-fat diet-induced obese mice (100mg/kg bw/day or 500 mg/kg bw/day, 5 times a week for 12 weeks by gavage) | Reduced the body weight gain, body fat, plasma lipid levels (TC and TG), insulin resistance and plasma leptin/adiponectin ratio Ameliorated the hepatic protein levels: hepatic lipogenesis (PPARγ, SREBP-1c, FAS and LPL), fatty acid β-oxidation (p-ACC and CPT1A), inflammation (TNF-α, NFkB and IL-1β) and antioxidant defense system | [216] |
| Diphlorethohydroxycarmalol isolated from Ishige okamurae | In vivo: high-fat diet-induced obese mice (25 mg/kg bw/day or 50 mg/kg bw/day for 6 weeks administered orally) | Reduced the body weight gain, body fat and hepatic lipid accumulation, and improved lipid profile Reduced the hepatic lipid accumulation by reduction in expression level of SREBP-1c, FABP4, and FAS Reduced the adipocyte size by down-regulation of enzyme expression (PPARγ, C/EBPα, SREBP-1c, FABP4, and FAS) | [217] |
| Polyphenol-rich extract from E. cava | In vivo: 97 overweight adults (low dose-72 mg/day or high dose-144 mg/day for 12 weeks) | Decreased the body fat and serum lipoid levels (TC and LDL-C) | [218] |
3.3. Metabolic Syndrome.
| Compounds/Marine Source | Test Model | Outcome | Ref. |
|---|---|---|---|
| Dietary iodine and seaweed consumption | In vivo: 2588 postmenopausal women for 2.85 years (between 2 and 4 years) | Inverse association between seaweed consumption with MetS incidence | [222] |
| Bioactive fraction of Sargassum wightii | In vitro: assays of ACE enzyme activity and antioxidant activity (DPPH, ABTS and FRAP) | Inhibition of ACE activity (IC50 56.96 μg/mL) and improved the antioxidant potency determined | [223] |
| An extract l from E. cava and pyrogallol-phloroglucinol-6,6-bieckol | In vivo: two mice models, high-fat diet-induced obese mice and high-cholesterol and saline diet-induced hypertension mice (70 mg extract or 500 mg extract or 2.5 mg pure phenol/kg bw/day for 4 weeks administered orally) In vitro: VSMC cells, an endothelial cell line | Reduced the blood pressure and serum lipoprotein levels in vivo Reduced the adhesion molecule expression, endothelial cell death and excessive migration and proliferation of VSMCs in vitro, as well as in the obese and hypertension mouse models | [224] |
| Ethanolic extract from Ulva lactuca enriched in phlorotannins | In vivo: hypercholesterolemic mice (250 mg/kg body weight for 4 weeks by gavage) | Improved the heart oxidative stress, plasma biochemical parameters and index of atherogenesis Down-regulated the expression of pro-inflammatory cytokines (TNFα, IL-1β and IL-6) in the heart | [225] |
| Food supplement from K. alvarezii | In vivo: rats fed for 8 weeks on high-carbohydrate, high-fat diet, alone or supplemented with 5% (w/w) algae | Reduced the body weight, adiposity, systolic blood pressure and plasma lipid levels Improved the heart and liver structure | [226] |
3.4. Neurodegenerative Diseases
| Compounds/Marine Source | Test Model | Outcome | Ref. |
|---|---|---|---|
| Phlorotannin-rich extract from E. cava (dieckol, 6,6′-bieckol, 8,8′-bieckol, eckol and phlorofucofuroeckol-A) | In vitro: assays of AChE, BChE and BACE-1 activities -- In vitro: Jurkat clone E1–6 cells (GSK3β activity at 50 μM) | Inhibition of AChE and BChE activities (IC50 16.0–96.3 μM and 0.9–29.0 μM, respectively) Inhibition of BACE-1 activity (18.6–58.3% at 1 μM) Inhibition of GSK3β activity (14.4–39.7% at 50 μM) | [234] |
| Phlorotannin-rich extract from E. bicyclis (eckols) | In vitro: assays of AChE and BChE activities | Inhibition of AChE and BChE activities (IC50 2.78 and 3.48 μg/mL, respectively) | [235] |
| Aqueous extracts of Gracilaria beckeri, Gelidium pristoides, U. rigida and E. maxima composed by phloroglucinol, catechin and epicatechin 3-glucoside | In vitro: assays of AChE and BChE activities | High antioxidant potency Inhibition of AChE and BChE activities (IC50 49.41 and 52.11 μg/mL, respectively, for E. maxima) Inhibition of Aβ aggregation | [236] |
| Aqueous-ethanolic extracts from E. maxima, G. pristoides, Gracilaria gracilis, and Ulva lactuca containing phlorotannins, flavonoids and phenolic acids | In vitro: assays of AChE, BChE and BACE-1 activities | Inhibition of AChE and BChE activities (IC50 1.74–2.42 and 1.55–2.04 mg/mL, respectively) Inhibition of BACE-1 activity (IC50 0.052–0.062 mg/mL) Inhibition of Aβ aggregation | [237] |
| Phlorofucofuroeckol isolated from E. cava | In vitro: Glutamate-stimulated PC12 cells (10 μM of phenol) | Increased the cell viability and attenuated glutamate excitotoxicity Inhibited the apoptosis in a caspase-dependent manner Regulated the production of ROS and attenuated mitochondrial dysfunction | [238] |
| Phloroglucinol isolated from E. cava | In vitro: Aβ-induced neurotoxicity in HT-22 cells (10 μg/mL) --- In vivo: 5XFAD mice, model of AD (acute, 1.2 μmol of phenol bilaterally delivery) | Reduced the Aβ-induced ROS accumulation in HT-22 cells Ameliorated the reduction in dendritic spine density --- Attenuated the impairments in cognitive dysfunction | [239] |
| Eckmaxol from E. maxima | In vitro: Aβ oligomer-induced neurotoxicity in SH-SY5Y cells (5–20 μM of phenol) | Prevented the Aβ oligomer-induced neurotoxicity Inhibition of GSK3β and ERK signaling pathway | [240] |
| E. cava rich in phlorotannins (eckol, 8,80-bieckol and dieckol) | In vitro: Aβ 25–35-induced damage in PC12 Cells (1–50 μM of phenol) | Inhibition of pro-inflammatory enzymes preventing Aβ production and neurotoxicity on the brain | [241] |
| Phlorotannin-rich fraction from Ishige foliacea | In vivo: scopolamine-induced amnesic mice (50 and 100 mg/kg bw/day of extract orally administered for 6 weeks) | Inhibition of AChE activity in the brain Improved the status antioxidant Prevented the memory impairment via regulation of ERK–CREB–BDNF pathway | [242] |
3.5. Cancer
| Compounds/Marine Source | Test Model | Outcome | Ref. |
|---|---|---|---|
| Dietary seaweed intake (gim, miyeok, and dashima) | In vivo: 923 colorectal cancer patients and 1846 controls | Association between c-MYC rs6983267 and colorectal cancer risk Inverse association between dietary seaweed intake and colorectal cancer risk | [248] |
| Aqueous extract derived from brown Cystoseira crinita | In vitro: HCT15 and MCF7 cells (25–250 μg/mL for extracts) | Antiproliferative activity (IC50 of 10.5–26.4 μg/mL on HCT15 and 17.9–29.5 μg/mL for 24 h) associated with phenolic content and antioxidant activity | [249] |
| Phlorotannin-rich extract from A. nodosum | In vitro: HT29 cells (100–500 μg/mL for extracts) | Antiproliferative activity | [250] |
| Ethanolic extract from S. muticum rich in phlorotannins | In vitro: HT29 cells (12.5–100 μg/mL for extracts) | Antiproliferative activity (IC50 of ~53.5–57.9, 55.0–57.8 and 59.4–74.0 μg/L for 24, 48 and 72 h of treatment of S. muticum extracts) | [127] |
| Phlorotannins isolated from Ecklonia maxima (phloroglucinol, eckol, 7-phloroeckol and 2-phloroeckol) | In vitro: HeLa, H157 and MCF7 cells (6.25–500 μg/mL for phenol) | Antiproliferative activity: eckol was the most active of all the tested phlorotannins against HeLa and MCF7 cells after 24 of treatment (IC50 < 50 μg/mL) | [251] |
| Alcoholic extract from red (Gracillaria corticata), green (Ulva fasciata) and brown (Sargassum ilicifolium) seaweeds | In vitro: MCF-7, MDA-MB-231, HeLa, HepG2 and HT-29 cells (15–300 μg/mL for extracts) | Antiproliferative activity: G. corticata extract had the greatest activity against MCF-7 cells (IC50 of 30, 37, 53, 102 and 250 μg/mL on MCF-7, HeLa, MDA-MB-231, HepG2 and HT-29 cells, respectively, after 24 h of treatment) G. corticata extract induced the apoptosis in human breast cancer cells | [252] |
| Phloroglucinol | In vitro: HCT116 and HT29 cells (10–300 μM of phenol) | Antiproliferative activity Intensified the 5-fluorouracil activity | [253] |
| 3,4,5-Tribromo-2-(2’,4’-dibromophenoxy)-phenol (1) and 3,5-dibromo-2-(2’,4’-dibromophenoxy)-phenol (2) isolated from marine sponge Dysidea sp. | In vitro: PANC-1 cells under glucose-starved conditions (1–100 μM of phenol) | Antiproliferative activity (IC50 values of 2.1 and 3.8 μM for 1 and 2, respectively, after 12 h) Inhibition of the complex II in the mitochondrial electron transport chain | [254] |
| Different extracts from F. vesiculosus L. rich in phlorotannins | In vitro: Panc89 and PancTU1 cells (0.8–500 μg/mL for crude extracts and 0.16–200 μg/mL for fractions) | Antiproliferative activity (IC50 of 72 μg/mL against Panc89 and of 77 μg/mL against PancTU1 cells after 72 h of treatment for the most active crude extract) | [24] |
| Extract from P. pavonica | In vitro: SaOS-2 and MNNG cells (0.5–2.5 μg/mL for extract) | Antiproliferative (IC50 value of 152.2 and 87.75 μg/mL for SaOS-2 and MNNG cells, respectively, after 24 h) and pro-apoptotic activities | [255] |
| Extracts of three brown marine macroalgae Dictyota dichotoma, Padaina pavonia and Sargassum vulgare | In vitro: LS174, A549, FemX, K562 cells (12.5–200 μg/mL for extract) | Characterization of the cytotoxic activity D. dichotoma showed the strongest cytotoxic activity of all the tested extracts (IC50 values ranging from 9.76 to 50.96 μg/mL after 72 h) | [256] |
| Extracts from detached leaves of Posidonia oceanica and Zostera marina | In vitro: MCF-7, MDA-MB-231, SK-BR-3, HT-29, HeLa, PC-3 and Neuro 2A cells, as well as African green monkey kidney (VERO) (6.25–100 μg/mL for extract) | Characterization of the cytotoxic activity Z. marina extract showed the best IC50 values of 25, 20 and 8 μg/mL after 48 h in neuroblastoma, colon and cervix cancer cell lines, respectively | [155] |
| Phlorotannin-rich fraction from Cystoseira sedoides | In vitro: MCF-7 cells (10–200 μg/mL for extract) | Characterization of the antiproliferative activity (IC50 value of 78 μg/mL after 72 h) | [257] |
| Crude extracts from two Egyptian brown seaweeds, Sargassum linearifolium and Cystoseira crinita | In vitro: a panel of cancer cells such as MCF-7 cells, among others (0.01–2000 μg/mL for extract) | Characterization of the cytotoxic activity. C. crinite cold methanolic extract showed a strong cytotoxic activity against MCF-7 cells (IC50 value of 18 μg/mL after 48 h) Induced the apoptosis and autophagy in MCF-7 cells | [258] |
| Aqueous seaweed extracts of 23 different species in Sri Lanka | In vitro: L929 cells (10–100 μg/mL for extract) | Antiproliferative activity Crude extracts of brown and red seaweeds species have shown high mortality rate compared to green seaweeds Jania adherens showed a remarkable cytotoxic effect on L929 cell line (51% cell viability compared with control after 24 h) | [259] |
| Ethanolic extract from E. cava whose main component was dieckol | In vitro: A2780 and SKOV3 cells | Cytotoxic effects on A2780 and SKOV3 ovarian cancer cells (IC50 ranging from 84 to 100 μg/mL for extract and from 77 to 169 μM for phenols, with dieckol being the most active of all, after 24 h) Induced the apoptosis on SKOV3 cells via Akt and p38 signaling pathways | [260] |
| Phlorotannin-rich extract from E. cava rich in dieckol | In vitro: A2780 and SKOV3 cells (50–100 μg/mL) In vivo: SKOV3-bearing mouse model (75 and 150 mg/kg bw for extract and 50 and 100 mg/kg bw for dieckol was given orally three times/week for 4 weeks) | Phlorotannin-rich extract may improve the efficacy of cisplatin for ovarian cancer by enhancing cancer cell apoptosis via the ROS/Akt/NFkB pathway | [261] |
| Phlorofucofuroeckol A present in E. bicyclis | In vitro: LoVo, HT-29, SW480 and HCT116 cells (25–100 μM of phenol) | Antiproliferative and pro-apoptotic properties Induced the apoptosis on colorectal cancer cells by ATF3 signaling pathway | [262] |
| Ethanolic extract of H. fusiforme | In vitro: B16F10 cells (25–400 μg/mL of extract) | Cytotoxic activity Induced the apoptosis through activation of extrinsic and intrinsic apoptotic pathways and ROS-dependent inhibition of the PI3K/Akt signaling pathway | [263] |
| Phlorotannin-rich extract from E. cava rich in phenolic compounds | In vitro: A549 cells (12.5–50 μg/mL of extract) | Inhibition of metastatic activity including suppression of migration and invasion Down-regulated the MMP-2 activity via PI3K/Akt | [264] |
| Phloroglucinol isolated from E. cava | In vitro: MCF7, SKBR3 and BT549 cells (10–100 μM of phenol) In vivo: MDA-MB231 breast cancer cells implanted into mammary fat pads of NOD-scid gamma (NSG) mice, treated with phloroglucinol 4 times on alternate days (25 mg/kg bw by intratumoral injections) | Antiproliferative effect by KRAS inhibition and its downstream PI3K/Akt and RAF-1/ERK signaling pathways Increased the sensitization of breast cancer cells to conventional therapy | [265] |
| Phloroglucinol isolated from E. cava | In vitro: BT549 and MDA-MB-231 cells (10–100 μM of phenol) In vivo: GFP-labeled metastatic MDA-MB231 cells transplanted into mammary fat pads of NSG mice, treated with phloroglucinol 4 times on alternate days (25 mg/kg bw by intraperitoneal injection) | Inhibited the metastatic ability of breast cancer cells Decreased the expression of SLUG, EMT master regulator through inhibition of PI3K⁄Akt and Ras⁄Raf-1 ⁄ERK Inhibited the in vivo metastatic ability of breast cancer cells | [266] |
| Bis(2,3-dibromo-4,5-dihydroxybenzyl) ether | In vitro: HUVEC cells (12.5–50 μM of phenol) In vivo: Zebrafish embryos model (6.25–25 μM of phenol) | Repressed the angiogenesis in both in vitro and in vivo models by inhibiting the VEGF signal systems | [267] |
| Dieckol from E. cava | In vivo: N-nitrosodiethylamime-induced hepatocarcinogenesis rats (40 mg/kg bw/day for 15 weeks administered orally) | Regulated the xenobiotic-metabolizing enzymes Induced the apoptosis by mitochondrial pathway Inhibited the invasion by decreasing PCNA expression Inhibited the angiogenesis by changing MMP-2 and MMP-9 activity and VEGF expression Anti-inflammatory activity by inhibiting NF-kB and COX2 | [268] |
| Dieckol | In vitro: EA.hy926 cells (10–100 μM of phenol) | Antiangiogenic activity by inhibiting the proliferation and migration of cells through MAPK, ERK and p38 signaling pathways | [269] |
| Eckol | In vitro: on human HaCaT keratinocytes against PM2.5-induced cell damage (30 μM of phenol for 17 days) | Decreased the ROS generation Protected the cells from apoptosis by inhibiting MAPK signaling pathway | [270] |
| Eckol | In vivo: sarcoma 180 (S180) xenograft-bearing animal model supplemented with low dose (0.25 mg/kg bw), middle dose (0.5 mg/kg bw) and high dose (1.0 mg/kg bw) of phenol administered orally | Proapoptotic and antiproliferative activities by improving the immune response | [271] |
| Polyphenol-rich extract from A. nodosum | In vivo: 80 overweight or obese population (100 mg/day of extract for 8 weeks) | Improvements in DNA damage in the obese subset | [30] |
3.6. Human Gut Microbiota
| Compounds/Marine Source | Test Model | Outcome | Ref. |
|---|---|---|---|
| Food supplement from Kappaphycus alvarezii | In vivo: rats fed for 8 weeks on high-carbohydrate, high-fat diet, alone or supplemented with 5% (w/w) algae | Improved the cardiovascular, liver and metabolic biomarkers in obese rats Modulated the balance between Firmicutes and Bacteroidetes in the gut | [226] |
| Polyphenol-rich extracts from brown macroalgae L. trabeculate | In vivo: high-fat diet and STZ-induced diabetic rats (200 mg/kg/day bw of phenol for 4 weeks by gavage) | Attenuated the hyperglycemia in diabetic rats Increased the short-chain fatty acid contents in fecal samples Enhanced the abundance of Bacteroidetes, Odoribacter and Muribaculum Decreased the abundance of Proteobacteria as well as Firmicutes/Bacteroidetes ratio | [103] |
| Water-ethanolic extract of green macroalgae Enteromorpha prolifera rich in flavonoids | In vivo: STZ-induced diabetic rats (150 mg/kg/day bw of phenol for 4 weeks by gavage) | Showed the antidiabetic activity on diabetic mice Modulated the balance between Firmicutes and Bacteroidetes in the gut and increased the abundance of the Lachnospiraceae and Alisties bacteria involved in the prevention of T2DM | [191] |
| Water-soluble compounds from Nitzschia laevis extract | In vivo: high-fat diet obese mice (50 mg/kg/day bw of extract for 8 weeks by gavage) | Prevented obesity in mice Protected the gut epithelium and positively reshaped the gut microbiota | [274] |
3.7. Infectious Diseases
| Compounds/Marine Source | Test Model | Outcome | Ref. |
|---|---|---|---|
| Phlorotannins purified extracts isolated from ten brown algal species (Cystoseira tamariscifolia, C. nodicaulis, C. usneoides, Sargassum vulgare, F. spiralis, Halopteris filicina, Stypocaulon scoparium, Cladostephus spongiosus, P. pavonica and Saccorhiza polyschides) from Portugal | In vitro broth microdilution assay | Less effective against fungi than bacteria Phlorotannin extracts were more effective against Gram-positive than Gram-negative bacteria Cystoseira species and F. spiralis were the most active against Staphylococcus and M. luteus (minimum MIC of 2.0 mg/mL) F. spiralis and C. nodicaulis extracts were the most effective against the studied fungi (MIC = 3.9 mg/mL) | [131] |
| Aqueous methanolic extracts isolated from Irish brown seaweed H. elongata | In vitro broth microdilution assay | High antimicrobial activity against the Gram-positive bacteria, L. monocytogenes and E. faecalis High antimicrobial activity against the Gram-negative bacteria, P. aeruginosa and S. abony | [276] |
| Turtlegrass Thalassia testudinum | Inoculations of healthy turtlegrass blades with Labyrinthula sp. | The emergence of Labyrinthula sp. lesions on turtlegrass blades causes a “pseudo-induction” of plant phenolic acids as carbon resources over-accumulate in tissues located above wound sites | [83] |
| Extracts isolated from Eelgrass Zostera marina, whose leaf surface contained hydroxycinnamic acids, flavones and flavanols | In vitro bioassays against microbial foulers | Involvement of surface-associated phenolic compounds to control yeasts | [5] |
| Free phenolic acid extracts from Nannochloropsis sp. and Spirulina sp., as well as pure compounds | In vitro antifungal activity of phenols | Antifungal activity of phenolic acid extracts of the microalgae Higher activity of the natural free phenolic acid extracts (EC50 values of 49.6 μg/mL and 33.9 μg/mL for Nannochloropsis sp. and Spirulina sp., respectively) than the synthetic mixtures | [27] |
| Ethanolic extracts isolated from nine marine microalgae (Nannochloropsis gaditana, Dunaliella salina, Dunaliella sp., Phaedactylum tricornutum, Isochrysis sp., Navicula sp., Chaeotoceros sp., Chlorella sp. and Tetraselmis sp.) | In vitro broth microdilution assay | Variable inhibitory activity against Escherichia coli, Pseudomonas aeruginosa and Staphylococcus aureus (Tetraselmis sp. was the most active of all those tested with MIC of 2.6 to 3.0 mg/mL of extract) Inhibition of the growth of Candida albicans (N. gaditana showed the highest activity with a MIC of 4.0 mg/mL of extract) Aspergillus niger (fungus) was resistant to the effects of the extracts Activity of the extracts was due to the presence of fatty acids, carotenoids and phenols | [50] |
| Methanol, ethanol and hexane extracts from four marine microalgae (Chaetoceros calcitrans, Skeletonema costatum, Chroococcus turgidus and Nannochloropsis oceanica) | In vitro disc diffusion method | Inhibitory activity against Staphylococcus aureus, Streptococcus pyogenes and Bacillus subtilis Antifungal activity only in Skeletonema costatum and Chroococcus turgidus | [59] |
| Diphlorethohydroxycarmalol isolated from Ishige okamurae | In vitro antiviral enzyme assay | Inhibited the activity of HIV-1 reverse transcriptase and integrase with IC50 values of 9.1 μM and 25.2 μM, respectively | [278] |
| 8,4′’-Dieckol isolated from E. cava | In vitro: H9, H9/HIV-1IIIB, CEM-SS, C8166 cells (1–50 μM of phenol) | Inhibited the activity of HIV-1 reverse transcriptase (RT) enzyme (91% inhibition ratio at 50 μM) and HIV-1 entry Exhibited the inhibitory effects against HIV-1 induced syncytia formation, lytic effects and viral p24 antigen production | [280] |
| 6,6′-Bieckol isolated from E. cava | In vitro: H9, H9/HIV-1IIIB, CEM-SS, C8166 cells (0.1–30 μM of phenol) | Inhibited the activity of HIV-1 RT enzyme (EC50 1.07 μM) as well as HIV-1 entry Exhibited the inhibitory effects against HIV-1 induced syncytia formation (EC50 1.72 μM), lytic effects (EC50 1.23 μM) and viral p24 antigen production (EC50 1.26 μM) | [279] |
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviatures
| A2780 | Human ovarian carcinoma cell line |
| A549 | Adenocarcinomic human alveolar basal epithelial cell line |
| Aβ | Amyloid-beta peptides |
| ABTS | 2, 2′-Azino-bis (3-ethylbenzothiazoline-6-sulphonic acid) |
| ACC | Acetyl-CoA carboxylase |
| ACE | Angiotensin-I converting enzyme |
| AChE | Acetylcholinesterase |
| ACh | Acetylcholine |
| ACSL1 | Adipose acyl-CoA synthetase 1 |
| AD | Alzheimer’s disease |
| A-FABP | Adipocyte fatty acid binding protein |
| AGEs | Advanced glycation end-products |
| AIDS | Acquired immunedeficiency syndrome |
| Akt | Protein kinase B |
| AMPK | AMP-activated protein kinase |
| ASE | Ascorbic acid equivalents |
| ATF3 | Transcription factor 3 |
| B16F10 | Murine melanoma cell line |
| BACE-1 | Beta-site amyloid precursor protein cleaving enzyme 1 |
| BCBM | β-Carotene bleaching |
| BChE | Butyrylcholinesterases |
| BDNF | Brain-derived neurotrophic factor |
| BT549 | Human triple negative breast cancer cell line |
| C2C12 | Mouse myoblast cell line |
| C8166 | Human cancer cell line |
| CA | Chelating ability |
| CAA | Antioxidant assay for cellular antioxidant activity |
| CAT | Catalase |
| CEM-SS | Human lymphoblastic leukemia cell line |
| CLPAA | Cellular lipid peroxidation antioxidant activity assay |
| COSY | Homonuclear correlation spectroscopy |
| GC–MS | Gas chromatography–mass spectrometry |
| Glut4 | Glucose transporter 4 |
| C/EBPα | CCAAT/enhancer-binding protein alpha |
| COX-2 | Cyclooxygenase-2 |
| CPT1A | Carnitine palmitoyltransferase I |
| CVD | Cardiovascular disease |
| DNA | Deoxyribonucleic acid |
| DPPH | 2,2-Diphenyl-1-picrylhydrazyl |
| E1-6 | Human Jurkat clone cell line |
| EA.hy926 | Human umbilical vein endothelial cell line |
| ERK | Extracellular signal-regulated kinase |
| FABP4 | Fatty acid binding protein 4 |
| FAS | Fatty acid synthase |
| FATP1 | Fatty acid transport protein-1 |
| FemX | Human malignant melanoma cell line |
| FFAs | Free fatty acids |
| FRAP | Ferric reducing antioxidant power |
| GAE | Gallic acid equivalents |
| Glut4 | Glucose transporter 4 |
| GPx | Glutathione peroxidase |
| GR | Glutathione reductase |
| GSK3β | Glycogen synthase kinase 3β |
| H157 | Human oral squamous cell carcinoma cell line |
| H9 | Human embryonic stem cell line |
| HCT-15 | Human colon adenocarcinoma cell line |
| HCT-116 | Human colon cancer cell line |
| HDL-C | HDL-cholesterol |
| HeLa | Human cervical cervix cancer cell line |
| HepG2 | Human hepatocellular carcinoma cell line |
| HIV-1 | Human immunodeficiency virus-1 |
| HMGCoA | 3-Hydroxyl-methyl glutaryl coenzyme A |
| HPLC–DAD–ESI/MS | High performance liquid chromatography–Diode array–Electrospray ionization–Mass spectrometry |
| HSQC | Heteronuclear single-quantum coherence spectroscopy |
| HT-22 | Mouse hippocampal neuronal cell line |
| HT29 | Human colon adenocarcinoma cell line |
| HUVEC | Human umbilical vein endothelia cells |
| IL | Interleukin |
| Ins-1 | Rat insulinoma cell line |
| IR | Infrared spectroscopy |
| IRS1 | Insulin receptor substrate 1 |
| JNKs | c-Jun N-terminal kinases |
| K562 | Human myelogenous leukemia cell line |
| L6 | Rat skeletal myoblast cell |
| L929 | Mouse fibroblasts cell line |
| LC–ESI–MS/MS | Liquid chromatography–Electrospray ionization–tandem Mass spectrometry |
| LDL-C | LDL-cholesterol |
| LMW | Low molecular weight |
| LoVo | Human colon cancer cell line derived from supraclavicular lymph node metastasis |
| LPIA | Lipid peroxidation inhibition assay |
| LS174 | Human colon adenocarcinoma cell line |
| MAPK | Mitogen-activated protein kinase |
| MC3T3-E1 | Mouse osteoblastic cell line |
| MetS | Metabolic syndrome |
| MMPs | Matrix metalloproteinase |
| MNNG | Human osteosarcoma cell line |
| NBT | Superoxide anion scavenging test |
| MCF-7 | Human breast adenocarcinoma cell line |
| MDA-MB-231 | Human breast adenocarcinoma cell line |
| MIC | Minimum inhibitory concentration |
| NFkB | Nuclear factor kappa B |
| NMR | Nuclear magnetic resonance |
| Neuro 2A | Mouse neuroblastoma cell line |
| NO | Nitric oxide |
| NOESY | Nuclear overhauser spectroscopy |
| iNOS | Inducible nitric oxide synthase |
| ORAC | Oxygen radical absorbance capacity |
| PANC-1 | Human pancreatic carcinoma cell line |
| Panc89 | Human pancreatic carcinoma cell line |
| PancTU1 | Human pancreatic carcinoma cell line |
| PC12 | Rat neuronal cell line |
| PC3 | Human prostate cancer cell line |
| PCNA | Proliferating cell nuclear antigen |
| PGE2 | Prostaglandin E2 |
| PGU | Phloroglucinol units |
| PI3K | Phosphoinositide 3-kinase |
| PPARγ | Proliferator activated receptor gamma |
| PTP1B | Protein tyrosine phosphatase 1B |
| RAGE | Receptor for advanced glycation end-products |
| RAW 264.7 | Murine macrophage cell line |
| RINm5F | Rat insulinoma cell line (pancreatic β-cells) |
| ROS | Reactive oxygen species |
| RP | Reducing power |
| RRLC-ESI-MS | Rapid resolution liquid chromatography coupled to mass spectrometry detection with negative ion electrospray ionization |
| RT | Reverse transcriptase |
| SaOS-2 | Human osteosarcoma cell line |
| SKBR3 | Human breast cancer cell line |
| SK-N-SH | Human neuroblastoma cell line |
| SKOV3 | Human ovarian carcinoma cell line |
| SOD | Superoxide dismutase |
| SREBP1 | Sterol regulatory element binding protein 1 |
| SREBP-1c | Sterol regulatory element binding protein -1c |
| SRSA | Superoxide radical scavenging assay |
| STZ | Streptozotocin |
| SW480 | Human colon cancer cell line |
| 3T3-L1 | Mouse adipocyte cell line |
| T2DM | Type 2 diabetes mellitus |
| TAA | Total antioxidant capacity |
| TBARs | Thiobarbituric acid reactive substances |
| TC | Total cholesterol |
| TEAC | Trolox equivalent antioxidant capacity |
| TG | Triglycerides |
| TLR | Toll-like receptor |
| TNFα | Tumor necrosis factor alpha |
| TOCSY | Total correlation spectroscopy |
| UCP-1 | Uncoupling protein 1 |
| UPLC | Ultra-performance liquid chromatography |
| UPLC–ESI–QTOF/MS | Ultra-performance liquid chromatography–Electrospray ionization–Quadrupole-time-of-flight high definition–Mass spectrometry |
| UPLC–MS | Ultra-performance liquid chromatography–Mass spectrometry |
| UPLC–MS/MS–TIC | Ultra-performance liquid chromatography–tandem Mass spectrometry–Total ion chromatogram |
| VEGF | Vascular endothelial growth factor |
| VERO | Green monkey kidney cell line |
| VSMC | Human vascular smooth muscle cell line |
References
- Shannon, E.; Abu-Ghannam, N. Seaweeds as nutraceuticals for health and nutrition. Phycologia 2019, 58, 563–577. [Google Scholar] [CrossRef]
- Dahlgren, E.; Enhus, C.; Lindqvist, D.; Eklund, B.; Asplund, L. Induced production of brominated aromatic compounds in the alga Ceramium tenuicorne. Environ. Sci. Pollut. Res. 2015, 22, 18107–18114. [Google Scholar] [CrossRef] [PubMed]
- Maadane, A.; Merghoub, N.; Ainane, T.; El Arroussi, H.; Benhima, R.; Amzazi, S.; Bakri, Y.; Wahby, I. Antioxidant activity of some Moroccan marine microalgae: PUFA profiles, carotenoids and phenolic content. J. Biotechnol. 2015, 215, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Kirke, D.A.; Rai, D.K.; Smyth, T.J.; Stengel, D.B. An assessment of temporal variation in the low molecular weight phlorotannin profiles in four intertidal brown macroalgae. Algal Res. 2019, 41, 101550. [Google Scholar] [CrossRef]
- Papazian, S.; Parrot, D.; Burýšková, B.; Weinberger, F.; Tasdemir, D. Surface chemical defence of the eelgrass Zostera marina against microbial foulers. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Barbosa, M.; Lopes, G.; Ferreres, F.; Andrade, P.B.; Pereira, D.M.; Gil-Izquierdo, Á.; Valentão, P. Phlorotannin extracts from Fucales: Marine polyphenols as bioregulators engaged in inflammation-related mediators and enzymes. Algal Res. 2017, 28, 1–8. [Google Scholar] [CrossRef]
- Lopes, G.; Barbosa, M.; Vallejo, F.; Gil-Izquierdo, Á.; Andrade, P.B.; Valentão, P.; Pereira, D.M.; Ferreres, F. Profiling phlorotannins from Fucus spp. of the Northern Portuguese coastline: Chemical approach by HPLC-DAD-ESI/MSn and UPLC-ESI-QTOF/MS. Algal Res. 2018, 29, 113–120. [Google Scholar] [CrossRef]
- Lopes, G.; Barbosa, M.; Andrade, P.B.; Valentão, P. Phlorotannins from Fucales: Potential to control hyperglycemia and diabetes-related vascular complications. J. Appl. Phycol. 2019, 31, 3143–3152. [Google Scholar] [CrossRef]
- Ravn, H.; Pedersen, M.F.; Borum, J.; Andary, C.; Anthoni, U.; Christophersen, C.; Nielsen, P.H. Seasonal variation and distribution of two phenolic compounds, rosmarinic acid and caffeic acid, in leaves and roots-rhizomes of eelgrass (Zostera marina L.). Ophelia 1994, 40, 51–61. [Google Scholar] [CrossRef]
- Achamlale, S.; Rezzonico, B.; Grignon-Dubois, M. Rosmarinic acid from beach waste: Isolation and HPLC quantification in Zostera detritus from Arcachon lagoon. Food Chem. 2009, 113, 878–883. [Google Scholar] [CrossRef]
- Achamlale, S.; Rezzonico, B.; Grignon-Dubois, M. Evaluation of Zostera detritus as a potential new source of zosteric acid. J. Appl. Phycol. 2009, 21, 347–352. [Google Scholar] [CrossRef]
- Mishra, N.; Prasad, S.M.; Mishra, N. Influence of high light intensity and nitrate deprivation on growth and biochemical composition of the marine microalgae Isochrysis galbana. Brazilian Arch. Biol. Technol. 2019, 62, 19180398. [Google Scholar] [CrossRef]
- Tierney, M.S.; Smyth, T.J.; Hayes, M.; Soler-Vila, A.; Croft, A.K.; Brunton, N. Influence of pressurised liquid extraction and solid-liquid extraction methods on the phenolic content and antioxidant activities of Irish macroalgae. Int. J. Food Sci. Technol. 2013, 48, 860–869. [Google Scholar] [CrossRef]
- Hardouin, K.; Burlot, A.S.; Umami, A.; Tanniou, A.; Stiger-Pouvreau, V.; Widowati, I.; Bedoux, G.; Bourgougnon, N. Biochemical and antiviral activities of enzymatic hydrolysates from different invasive French seaweeds. J. Appl. Phycol. 2014, 26, 1029–1042. [Google Scholar] [CrossRef]
- Casas, M.P.; Conde, E.; Domínguez, H.; Moure, A. Ecofriendly extraction of bioactive fractions from Sargassum muticum. Process Biochem. 2019, 79, 166–173. [Google Scholar] [CrossRef]
- Kadam, S.U.; Tiwari, B.K.; Smyth, T.J.; O’Donnell, C.P. Optimization of ultrasound assisted extraction of bioactive components from brown seaweed Ascophyllum nodosum using response surface methodology. Ultrason. Sonochem. 2015, 23, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Parniakov, O.; Apicella, E.; Koubaa, M.; Barba, F.J.; Grimi, N.; Lebovka, N.; Pataro, G.; Ferrari, G.; Vorobiev, E. Ultrasound-assisted green solvent extraction of high-added value compounds from microalgae Nannochloropsis spp. Bioresour. Technol. 2015, 198, 262–267. [Google Scholar] [CrossRef]
- Huang, J.J.H.; Xu, W.W.; Lin, S.L.; Cheung, P.C.K. Phytochemical profiles of marine phytoplanktons: An evaluation of their: In vitro antioxidant and anti-proliferative activities. Food Funct. 2016, 7, 5002–5017. [Google Scholar] [CrossRef]
- Rajauria, G.; Foley, B.; Abu-Ghannam, N. Identification and characterization of phenolic antioxidant compounds from brown Irish seaweed Himanthalia elongata using LC-DAD–ESI-MS/MS. Innov. Food Sci. Emerg. Technol. 2016, 37, 261–268. [Google Scholar] [CrossRef]
- Rajauria, G. Optimization and validation of reverse phase HPLC method for qualitative and quantitative assessment of polyphenols in seaweed. J. Pharm. Biomed. Anal. 2018, 148, 230–237. [Google Scholar] [CrossRef]
- Heffernan, N.; Brunton, N.P.; FitzGerald, R.J.; Smyth, T.J. Profiling of the molecular weight and structural isomer abundance of macroalgae-derived phlorotannins. Mar. Drugs 2015, 13, 509–528. [Google Scholar] [CrossRef]
- Bogolitsyn, K.; Druzhinina, A.; Kaplitsin, P.; Ovchinnikov, D.; Parshina, A.; Kuznetsova, M. Relationship between radical scavenging activity and polymolecular properties of brown algae polyphenols. Chem. Pap. 2019, 73, 2377–2385. [Google Scholar] [CrossRef]
- Laabir, M.; Grignon-Dubois, M.; Masseret, E.; Rezzonico, B.; Soteras, G.; Rouquette, M.; Rieuvilleneuve, F.; Cecchi, P. Algicidal effects of Zostera marina L. and Zostera noltii Hornem. extracts on the neuro-toxic bloom-forming dinoflagellate Alexandrium catenella. Aquat. Bot. 2013, 111, 16–25. [Google Scholar] [CrossRef]
- Zenthoefer, M.; Geisen, U.; Hofmann-Peiker, K.; Fuhrmann, M.; Kerber, J.; Kirchhöfer, R.; Hennig, S.; Peipp, M.; Geyer, R.; Piker, L.; et al. Isolation of polyphenols with anticancer activity from the Baltic Sea brown seaweed Fucus vesiculosus using bioassay-guided fractionation. J. Appl. Phycol. 2017, 29, 2021–2037. [Google Scholar] [CrossRef]
- Tierney, M.S.; Soler-Vila, A.; Rai, D.K.; Croft, A.K.; Brunton, N.P.; Smyth, T.J. UPLC-MS profiling of low molecular weight phlorotannin polymers in Ascophyllum nodosum, Pelvetia canaliculata and Fucus spiralis. Metabolomics 2014, 10, 524–535. [Google Scholar] [CrossRef]
- Zhong, B.; Robinson, N.A.; Warner, R.D.; Barrow, C.J.; Dunshea, F.R.; Suleria, H.A.R. LC-ESI-QTOF-MS/MS characterization of seaweed phenolics and their antioxidant potential. Mar. Drugs 2020, 18, 331. [Google Scholar] [CrossRef]
- Scaglioni, P.T.; Pagnussatt, F.A.; Lemos, A.C.; Nicolli, C.P.; Del Ponte, E.M.; Badiale-Furlong, E. Nannochloropsis sp. and Spirulina sp. as a source of antifungal compounds to mitigate contamination by Fusarium graminearum sspecies complex. Curr. Microbiol. 2019, 76, 930–938. [Google Scholar] [CrossRef]
- Klejdus, B.; Plaza, M.; Šnóblová, M.; Lojková, L. Development of new efficient method for isolation of phenolics from sea algae prior to their rapid resolution liquid chromatographic–tandem mass spectrometric determination. J. Pharm. Biomed. Anal. 2017, 135, 87–96. [Google Scholar] [CrossRef]
- Pilavtepe, M.; Yucel, M.; Helvaci, S.S.; Demircioglu, M.; Yesil-Celiktas, O. Optimization and mathematical modeling of mass transfer between Zostera marina residues and supercritical CO2 modified with ethanol. J. Supercrit. Fluids 2012, 68, 87–93. [Google Scholar] [CrossRef]
- Baldrick, F.R.; McFadden, K.; Ibars, M.; Sung, C.; Moffatt, T.; Megarry, K.; Thomas, K.; Mitchell, P.; Wallace, J.M.W.; Pourshahidi, L.K.; et al. Impact of a (poly)phenol-rich extract from the brown algae Ascophyllum nodosum on DNA damage and antioxidant activity in an overweight or obese population: A randomized controlled trial. Am. J. Clin. Nutr. 2018, 108, 688–700. [Google Scholar] [CrossRef]
- Akköz, C.; Arslan, D.; Ünver, A.; Özcan, M.M.; Yilmaz, B. Chemical composition, total phenolic and mineral contents of Enteromorpha intestinalis (l.) kütz. and cladophora glomerata (l.) kütz. seaweeds. J. Food Biochem. 2011, 35, 513–523. [Google Scholar] [CrossRef]
- Wijesekara, I.; Kim, S.K.; Li, Y.; Li, Y.X. Phlorotannins as bioactive agents from brown algae. Process Biochem. 2011, 46, 2219–2224. [Google Scholar] [CrossRef]
- Zidorn, C. Secondary metabolites of seagrasses (Alismatales and Potamogetonales; Alismatidae): Chemical diversity, bioactivity, and ecological function. Phytochemistry 2016, 124, 5–28. [Google Scholar] [CrossRef] [PubMed]
- Catarino, M.D.; Silva, A.M.S.; Cardoso, S.M. Fucaceae: A source of bioactive phlorotannins. Int. J. Mol. Sci. 2017, 18, 1327. [Google Scholar] [CrossRef]
- Montero, L.; del Pilar Sánchez-Camargo, A.; Ibáñez, E.; Gilbert-López, B. Phenolic compounds from edible algae: Bioactivity and health benefits. Curr. Med. Chem. 2018, 25, 4808–4826. [Google Scholar] [CrossRef]
- Erpel, F.; Mateos, R.; Pérez-jiménez, J.; Pérez-correa, J.R. Phlorotannins: From isolation and structural characterization, to the evaluation of their antidiabetic and anticancer potential. Food Res. Int. 2020, 137, 109589. [Google Scholar] [CrossRef]
- Bidleman, T.F.; Andersson, A.; Brugel, S.; Ericson, L.; Haglund, P.; Kupryianchyk, D.; Lau, D.C.P.; Liljelind, P.; Lundin, L.; Tysklind, A.; et al. Bromoanisoles and methoxylated bromodiphenyl ethers in macroalgae from Nordic coastal regions. Environ. Sci. Process. Impacts 2019, 21, 881–892. [Google Scholar] [CrossRef]
- Cade, S.E.; Kuo, L.J.; Schultz, I.R. Polybrominated diphenyl ethers and their hydroxylated and methoxylated derivatives in seafood obtained from Puget Sound, WA. Sci. Total Environ. 2018, 630, 1149–1154. [Google Scholar] [CrossRef]
- Malmvärn, A.; Marsh, G.; Kautsky, L.; Athanasiadou, M.; Bergman, Å.; Asplund, L. Hydroxylated and methoxylated brominated diphenyl ethers in the red algae Ceramium tenuicorne and blue mussels from the Baltic Sea. Environ. Sci. Technol. 2005, 39, 2990–2997. [Google Scholar] [CrossRef]
- Koch, C.; Sures, B. Environmental concentrations and toxicology of 2,4,6-tribromophenol (TBP). Environ. Pollut. 2018, 233, 706–713. [Google Scholar] [CrossRef]
- Ragupathi Raja Kannan, R.; Arumugam, R.; Iyapparaj, P.; Thangaradjou, T.; Anantharaman, P. In vitro antibacterial, cytotoxicity and haemolytic activities and phytochemical analysis of seagrasses from the Gulf of Mannar, South India. Food Chem. 2013, 136, 1484–1489. [Google Scholar] [CrossRef]
- Zangrando, R.; Corami, F.; Barbaro, E.; Grosso, A.; Barbante, C.; Turetta, C.; Capodaglio, G.; Gambaro, A. Free phenolic compounds in waters of the Ross Sea. Sci. Total Environ. 2019, 650, 2117–2128. [Google Scholar] [CrossRef]
- Glombitza, K.W.; Gerstberger, G. Phlorotannins with dibenzodioxin structural elements from the brown alga Eisenia arborea. Phytochemistry 1985, 24, 543–551. [Google Scholar] [CrossRef]
- Pal Singh, I.; Bharate, S.B. Phloroglucinol compounds of natural origin. Nat. Prod. Rep. 2006, 23, 558–591. [Google Scholar] [CrossRef]
- Koivikko, R.; Loponen, J.; Pihlaja, K.; Jormalainen, V. High-performance liquid chromatographic analysis of phlorotannins from the brown alga Fucus vesiculosus. Phytochem. Anal. 2007, 18, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Santana-Casiano, J.M.; González-Dávila, M.; González, A.G.; Millero, F.J. Fe(III) reduction in the presence of Catechol in seawater. Aquat. Geochemistry 2010, 16, 467–482. [Google Scholar] [CrossRef]
- Sansone, C.; Brunet, C. Promises and challenges of microalgal antioxidant production. Antioxidants 2019, 8, 199. [Google Scholar] [CrossRef] [PubMed]
- Tibbetts, S.M.; Milley, J.E.; Lall, S.P. Chemical composition and nutritional properties of freshwater and marine microalgal biomass cultured in photobioreactors. J. Appl. Phycol. 2015, 27, 1109–1119. [Google Scholar] [CrossRef]
- Safafar, H.; van Wagenen, J.; Møller, P.; Jacobsen, C. Carotenoids, phenolic compounds and tocopherols contribute to the antioxidative properties of some microalgae species grown on industrial wastewater. Mar. Drugs 2015, 13, 7339–7356. [Google Scholar] [CrossRef] [PubMed]
- Maadane, A.; Merghoub, N.; El Mernissi, N.; Ainane, T.; Amzazi, S.; Wahby, I.; Bakri, Y. Antimicrobial activity of marine microalgae isolated from Moroccan coastlines. J. Microbiol. Biotechnol. Food Sci. 2017, 6, 1257–1260. [Google Scholar] [CrossRef]
- Bhuvana, P.; Sangeetha, P.; Anuradha, V.; Ali, M.S. Spectral characterization of bioactive compounds from microalgae: N. oculata and C. vulgaris. Biocatal. Agric. Biotechnol. 2019, 19, 101094. [Google Scholar] [CrossRef]
- Gomez, A.L.; Lopez, J.A.; Rodriguez, A.; Fortiz, J.; Martinez, L.R.; Apolinar, A.; Enriquez, L.F. Produccion de compuestos fenolicos por cuatro especies de microalgas marinas sometidas a diferentes condiciones de iluminacion. Lat. Am. J. Aquat. Res. 2016, 44, 137–143. [Google Scholar] [CrossRef]
- Fazelian, N.; Movafeghi, A.; Yousefzadi, M.; Rahimzadeh, M. Cytotoxic impacts of CuO nanoparticles on the marine microalga Nannochloropsis oculata. Environ. Sci. Pollut. Res. 2019, 26, 17499–17511. [Google Scholar] [CrossRef] [PubMed]
- Rico, M.; López, A.; Santana-Casiano, J.M.; González, A.G.; González-Dávila, M. Variability of the phenolic profile in the diatom Phaeodactylum tricornutum growing under copper and iron stress. Limnol. Oceanogr. 2013, 58, 144–152. [Google Scholar] [CrossRef]
- Santana-Casiano, J.M.; González-Dávila, M.; González, A.G.; Rico, M.; López, A.; Martel, A. Characterization of phenolic exudates from Phaeodactylum tricornutum and their effects on the chemistry of Fe(II)-Fe(III). Mar. Chem. 2014, 158, 10–16. [Google Scholar] [CrossRef]
- Custódio, L.; Soares, F.; Pereira, H.; Barreira, L.; Vizetto-Duarte, C.; Rodrigues, M.J.; Rauter, A.P.; Alberício, F.; Varela, J. Fatty acid composition and biological activities of Isochrysis galbana T-ISO, Tetraselmis sp. and Scenedesmus sp.: Possible application in the pharmaceutical and functional food industries. J. Appl. Phycol. 2014, 26, 151–161. [Google Scholar] [CrossRef]
- Mekdade, L.; Bey, M.; Hamed, B.; El-kebir, F.Z.; Mohamed, S.; Ayad, E.A.; June, M.; June, M. Evaluation of Antioxidant and Antiproliferative Activities of Nannochloropsis. Res. Journal Pharm., Biol. Chem. Sci. 2016, 7, 904–913. [Google Scholar]
- Hemalatha, A.; Parthiban, C.; Saranya, C.; Girija, K.; Anantharaman, P. Evaluation of antioxidant activities and total phenolic contents of different solvent extracts of selected marine diatoms. Indian J. Geo-Marine Sci. 2015, 44, 1630–1636. [Google Scholar]
- Sushanth, V.R.; Rajashekhar, M. Antioxidant and antimicrobial activities in the four species of marine microalgae isolated from Arabian Sea of Karnataka Coast. Indian J. Geo-Marine Sci. 2015, 44, 69–75. [Google Scholar]
- Niwano, Y.; Sato, E.; Kohno, M.; Matsuyama, Y.; Kim, D.; Oda, T. Antioxidant properties of aqueous extracts from red tide plankton cultures. Biosci. Biotechnol. Biochem. 2007, 71, 1145–1153. [Google Scholar] [CrossRef]
- Custódio, L.; Justo, T.; Silvestre, L.; Barradas, A.; Duarte, C.V.; Pereira, H.; Barreira, L.; Rauter, A.P.; Alberício, F.; Varela, J. Microalgae of different phyla display antioxidant, metal chelating and acetylcholinesterase inhibitory activities. Food Chem. 2012, 131, 134–140. [Google Scholar] [CrossRef]
- Pang, X.; Lin, X.; Wang, P.; Zhou, X.; Yang, B.; Wang, J.; Liu, Y. Perylenequione derivatives with anticancer activities isolated from the marine sponge-derived fungus, Alternaria sp. SCSIO41014. Mar. Drugs 2018, 16, 280. [Google Scholar] [CrossRef]
- Zhao, Z.; Ding, W.; Wang, P.-M.; Zheng, D.; Xu, J. Five polyketides isolated from the marine-derived fungus Arthrinium sp. Nat. Prod. Res. 2019, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Tian, L.; Chen, G.; Zhang, L.; Liu, B.; Zhang, W.; Bai, J.; Hua, H.; Wang, H.; Pei, Y.-H. Two new compounds from a marine-derived Penicillium griseofulvum T21-03. J. Asian Nat. Prod. Res. 2017, 19, 678–683. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.L.; Shao, C.L.; Chen, J.F.; Guo, Z.Y.; Fu, X.M.; Chen, M.; Chen, Y.Y.; Li, R.; De Voogd, N.J.; She, Z.G.; et al. New bisabolane sesquiterpenoids from a marine-derived fungus Aspergillus sp. isolated from the sponge Xestospongia testudinaria. Bioorganic Med. Chem. Lett. 2012, 22, 1326–1329. [Google Scholar] [CrossRef]
- Liu, S.; Dai, H.; Konuklugil, B.; Orfali, R.S.; Lin, W.; Kalscheuer, R.; Liu, Z.; Proksch, P. Phenolic bisabolanes from the sponge-derived fungus Aspergillus sp. Phytochem. Lett. 2016, 18, 187–191. [Google Scholar] [CrossRef]
- Wu, Z.; Wang, Y.; Liu, D.; Proksch, P.; Yu, S.; Lin, W. Antioxidative phenolic compounds from a marine-derived fungus Aspergillus versicolor. Tetrahedron 2016, 72, 50–57. [Google Scholar] [CrossRef]
- Hulikere, M.M.; Joshi, C.G.; Ananda, D.; Poyya, J.; Nivya, T. Antiangiogenic, wound healing and antioxidant activity of Cladosporium cladosporioides (Endophytic Fungus) isolated from seaweed (Sargassum wightii). Mycology 2016, 7, 203–211. [Google Scholar] [CrossRef]
- El-Hawary, S.S.; Sayed, A.M.; Mohammed, R.; Hassan, H.M.; Zaki, M.A.; Rateb, M.E.; Mohammed, T.A.; Amin, E.; Abdelmohsen, U.R. Epigenetic modifiers induce bioactive phenolic metabolites in the marine-derived fungus Penicillium brevicompactum. Mar. Drugs 2018, 16, 253. [Google Scholar] [CrossRef]
- Zheng, Y.; Chen, X.; Chen, L.; Shen, L.; Fu, X.; Chen, Q.; Chen, M.; Wang, C. Isolation and neuroprotective activity of phenolic derivatives from the marine derived fungus Penicillium janthinellum. J. Ocean Univ. China 2020, 19, 700–706. [Google Scholar] [CrossRef]
- Lu, Z.; Zhu, H.; Fu, P.; Wang, Y.; Zhang, Z.; Lin, H.; Liu, P.; Zhuang, Y.; Hong, K.; Zhu, W. Cytotoxic polyphenols from the marine-derived fungus Penicillium expansum. J. Nat. Prod. 2010, 73, 911–914. [Google Scholar] [CrossRef] [PubMed]
- Elnaggar, M.S.; Ebada, S.S.; Ashour, M.L.; Ebrahim, W.; Müller, W.E.G.; Mándi, A.; Kurtán, T.; Singab, A.; Lin, W.; Liu, Z.; et al. Xanthones and sesquiterpene derivatives from a marine-derived fungus Scopulariopsis sp. Tetrahedron 2016, 72, 2411–2419. [Google Scholar] [CrossRef]
- Wang, J.F.; Qin, X.; Xu, F.Q.; Zhang, T.; Liao, S.; Lin, X.; Yang, B.; Liu, J.; Wang, L.; Tu, Z.; et al. Tetramic acid derivatives and polyphenols from sponge-derived fungus and their biological evaluation. Nat. Prod. Res. 2015, 29, 1761–1765. [Google Scholar] [CrossRef] [PubMed]
- Blagojević, D.; Babić, O.; Rašeta, M.; Šibul, F.; Janjušević, L.; Simeunović, J. Antioxidant activity and phenolic profile in filamentous cyanobacteria: The impact of nitrogen. J. Appl. Phycol. 2018, 30, 2337–2346. [Google Scholar] [CrossRef]
- Ijaz, S.; Hasnain, S. Antioxidant potential of indigenous cyanobacterial strains in relation with their phenolic and flavonoid contents. Nat. Prod. Res. 2016, 30, 1297–1300. [Google Scholar] [CrossRef] [PubMed]
- Jerez-Martel, I.; García-Poza, S.; Rodríguez-Martel, G.; Rico, M.; Afonso-Olivares, C.; Gómez-Pinchetti, J.L. Phenolic profile and antioxidant activity of crude extracts from microalgae and cyanobacteria strains. J. Food Qual. 2017, 2017. [Google Scholar] [CrossRef]
- Milović, S.; Stanković, I.; Nikolić, D.; Radović, J.; Kolundžić, M.; Nikolić, V.; Stanojković, T.; Petović, S.; Kundaković-Vasović, T. Chemical analysis of selected seaweeds and seagrass from the Adriatic Coast of Montenegro. Chem. Biodivers. 2019, 16. [Google Scholar] [CrossRef] [PubMed]
- Steele, L.T.; Valentine, J.F. Idiosyncratic responses of seagrass phenolic production following sea urchin grazing. Mar. Ecol. Prog. Ser. 2012, 466, 81–92. [Google Scholar] [CrossRef]
- Mabrouk, S.B.; Reis, M.; Sousa, M.L.; Ribeiro, T.; Almeida, J.R.; Pereira, S.; Antunes, J.; Rosa, F.; Vasconcelos, V.; Achour, L.; et al. The marine seagrass Halophila stipulacea as a source of bioactive metabolites against obesity and biofouling. Mar. Drugs 2020, 18, 88. [Google Scholar] [CrossRef]
- Cornara, L.; Pastorino, G.; Borghesi, B.; Salis, A.; Clericuzio, M.; Marchetti, C.; Damonte, G.; Burlando, B. Posidonia oceanica (L.) delile ethanolic extract modulates cell activities with skin health applications. Mar. Drugs 2018, 16, 21. [Google Scholar] [CrossRef]
- Enerstvedt Hasle, K.; Lundberg, A.; Jordheim, M. Characterization of polyphenolic content in the aquatic plants Ruppia cirrhosa and Ruppia maritima -A source of nutritional natural products. Molecules 2018, 23, 16. [Google Scholar] [CrossRef] [PubMed]
- Rengasamy, K.R.R.; Sadeer, N.B.; Zengin, G.; Mahomoodally, M.F.; Cziáky, Z.; Jekő, J.; Diuzheva, A.; Abdallah, H.H.; Kim, D.H. Biopharmaceutical potential, chemical profile and in silico study of the seagrass—Syringodium isoetifolium (Asch.) Dandy. S. Afr. J. Bot. 2019, 127, 167–175. [Google Scholar] [CrossRef]
- Steele, L.T.; Caldwell, M.; Boettcher, A.; Arnold, T. Seagrass-pathogen interactions: “pseudo-induction” of turtlegrass phenolics near wasting disease lesions. Mar. Ecol. Prog. Ser. 2005, 303, 123–131. [Google Scholar] [CrossRef]
- Trevathan-Tackett, S.M.; Lane, A.L.; Bishop, N.; Ross, C. Metabolites derived from the tropical seagrass Thalassia testudinum are bioactive against pathogenic Labyrinthula sp. Aquat. Bot. 2015, 122, 1–8. [Google Scholar] [CrossRef]
- Styshova, O.N.; Popov, A.M.; Artyukov, A.A.; Klimovich, A.A. Main constituents of polyphenol complex from seagrasses of the genus Zostera, their antidiabetic properties and mechanisms of action. Exp. Ther. Med. 2017, 13, 1651–1659. [Google Scholar] [CrossRef]
- Li, Y.; Mangoni, A.; Shulha, O.; Çiçek, S.S.; Zidorn, C. Cyclic diarylheptanoids deoxycymodienol and isotedarene A from Zostera marina (Zosteraceae). Tetrahedron Lett. 2019, 60, 150930. [Google Scholar] [CrossRef]
- Grignon-Dubois, M.; Rezzonico, B.; Alcoverro, T. Regional scale patterns in seagrass defences: Phenolic acid content in Zostera noltii. Estuar. Coast. Shelf Sci. 2012, 114, 18–22. [Google Scholar] [CrossRef]
- Manck, L.; Quintana, E.; Suárez, R.; Brun, F.G.; Hernández, I.; Ortega, M.J.; Zubía, E. Profiling of phenolic natural products in the seagrass Zostera noltei by UPLC-MS. Nat. Prod. Commun. 2017, 12, 687–690. [Google Scholar] [CrossRef]
- Grignon-Dubois, M.; Rezzonico, B. Phenolic chemistry of the seagrass Zostera noltei Hornem. Part 1: First evidence of three infraspecific flavonoid chemotypes in three distinctive geographical regions. Phytochemistry 2018, 146, 91–101. [Google Scholar] [CrossRef]
- Arnold, T.; Freundlich, G.; Weilnau, T.; Verdi, A.; Tibbetts, I.R. Impacts of groundwater discharge at Myora Springs (North Stradbroke Island, Australia) on the phenolic metabolism of eelgrass, Zostera muelleri, and grazing by the juvenile rabbitfish, Siganus fuscescens. PLoS ONE 2014, 9, e104738. [Google Scholar] [CrossRef]
- Cichewicz, R.H.; Clifford, L.J.; Lassen, P.R.; Cao, X.; Freedman, T.B.; Nafie, L.A.; Deschamps, J.D.; Kenyon, V.A.; Flanary, J.R.; Holman, T.R.; et al. Stereochemical determination and bioactivity assessment of (S)-(+)-curcuphenol dimers isolated from the marine sponge Didiscus aceratus and synthesized through laccase biocatalysis. Bioorganic Med. Chem. 2005, 13, 5600–5612. [Google Scholar] [CrossRef] [PubMed]
- Kaweetripob, W.; Mahidol, C.; Wongbundit, S.; Tuntiwachwuttikul, P.; Ruchirawat, S.; Prawat, H. Sesterterpenes and phenolic alkenes from the Thai sponge Hyrtios erectus. Tetrahedron 2018, 74, 316–323. [Google Scholar] [CrossRef]
- Costa, M.; Coello, L.; Urbatzka, R.; Pérez, M.; Thorsteinsdottir, M. New aromatic bisabolane derivatives with lipid-reducing activity from the marine sponge Myrmekioderma sp. Mar. Drugs 2019, 17, 375. [Google Scholar] [CrossRef] [PubMed]
- Wongbundit, S.; Mahidol, C.; Kaweetripob, W.; Prachyawarakorn, V.; Eurtivong, C.; Sahakitpichan, P.; Tuntiwachwuttikul, P.; Ruchirawat, S.; Prawat, H. Biscurcudiols, myrmekioperoxides, and myrmekiodermaral from the Thai marine sponge Myrmekioderma sp. Tetrahedron 2020, 76, 131162. [Google Scholar] [CrossRef]
- Olsen, E.K.; Hansen, E.; Isaksson, J.; Andersen, J.H. Cellular antioxidant effect of four bromophenols from the red algae, Vertebrata lanosa. Mar. Drugs 2013, 11, 2769–2784. [Google Scholar] [CrossRef]
- Ferreres, F.; Lopes, G.; Gil-Izquierdo, A.; Andrade, P.B.; Sousa, C.; Mouga, T.; Valentão, P. Phlorotannin extracts from fucales characterized by HPLC-DAD-ESI-MS n: Approaches to hyaluronidase inhibitory capacity and antioxidant properties. Mar. Drugs 2012, 10, 2766–2781. [Google Scholar] [CrossRef]
- Prieto, P.; Pineda, M.; Aguilar, M. Spectrophotometric quantitation of antioxidant capacity through the formation of a phosphomolybdenum complex: Specific application to the determination of vitamin E. Anal. Biochem. 1999, 269, 337–341. [Google Scholar] [CrossRef]
- Cabrita, M.T.; Vale, C.; Rauter, A.P. Halogenated compounds from marine algae. Mar. Drugs 2010, 8, 2301–2317. [Google Scholar] [CrossRef]
- Topcu, G.; Aydogmus, Z.; Imre, S.; Gören, A.C.; Pezzuto, J.M.; Clement, J.A.; Kingston, D.G.I. Brominated sesquiterpenes from the red alga Laurencia obtusa. J. Nat. Prod. 2003, 66, 1505–1508. [Google Scholar] [CrossRef]
- Machado, L.; Magnusson, M.; Paul, N.A.; Kinley, R.; de Nys, R.; Tomkins, N. Identification of bioactives from the red seaweed Asparagopsis taxiformis that promote antimethanogenic activity in vitro. J. Appl. Phycol. 2016, 28, 3117–3126. [Google Scholar] [CrossRef]
- Whitfield, F.B.; Last, J.H.; Shaw, K.J.; Tindale, C.R. 2,6-Dibromophenol: The cause of an iodoform-like off-flavour in some Australian crustacea. J. Sci. Food Agric. 1988, 46, 29–42. [Google Scholar] [CrossRef]
- Kim, K.C.; Hyun, Y.J.; Hewage, S.R.K.M.; Piao, M.J.; Kang, K.A.; Kang, H.K.; Koh, Y.S.; Ahn, M.J.; Hyun, J.W. 3-Bromo-4,5-dihydroxybenzaldehyde enhances the level of reduced glutathione via the Nrf2-mediated pathway in human keratinocytes. Mar. Drugs 2017, 15, 291. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Zheng, Y.; Zhou, J.; Geng, Y.; Zou, P.; Li, Y.; Zhang, C. Polyphenol-rich extracts from brown macroalgae Lessonia trabeculate attenuate hyperglycemia and modulate gut microbiota in high-rfat diet and Streptozotocin-Induced diabetic rats. J. Agric. Food Chem. 2019, 67, 12472–12480. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, D.; Freitas, A.C.; Pereira, L.; Rocha-Santos, T.A.P.; Vasconcelos, M.W.; Roriz, M.; Rodríguez-Alcalá, L.M.; Gomes, A.M.P.; Duarte, A.C. Chemical composition of red, brown and green macroalgae from Buarcos bay in Central West Coast of Portugal. Food Chem. 2015, 183, 197–207. [Google Scholar] [CrossRef]
- Uribe, E.; Vega-Gálvez, A.; Heredia, V.; Pastén, A.; Di Scala, K. An edible red seaweed (Pyropia orbicularis): Influence of vacuum drying on physicochemical composition, bioactive compounds, antioxidant capacity, and pigments. J. Appl. Phycol. 2018, 30, 673–683. [Google Scholar] [CrossRef]
- Tanna, B.; Brahmbhatt, H.R.; Mishra, A. Phenolic, flavonoid, and amino acid compositions reveal that selected tropical seaweeds have the potential to be functional food ingredients. J. Food Process. Preserv. 2019, 43, 1–10. [Google Scholar] [CrossRef]
- Ford, L.; Theodoridou, K.; Sheldrake, G.N.; Walsh, P.J. A critical review of analytical methods used for the chemical characterisation and quantification of phlorotannin compounds in brown seaweeds. Phytochem. Anal. 2019, 30, 587–599. [Google Scholar] [CrossRef]
- Steevensz, A.J.; MacKinnon, S.L.; Hankinson, R.; Craft, C.; Connan, S.; Stengel, D.B.; Melanson, J.E. Profiling phlorotannins in brown macroalgae by liquid chromatography-high resolution mass spectrometry. Phytochem. Anal. 2012, 23, 547–553. [Google Scholar] [CrossRef]
- Zhang, R.; Yuen, A.K.L.; Magnusson, M.; Wright, J.T.; de Nys, R.; Masters, A.F.; Maschmeyer, T. A comparative assessment of the activity and structure of phlorotannins from the brown seaweed Carpophyllum flexuosum. Algal Res. 2018, 29, 130–141. [Google Scholar] [CrossRef]
- Sellimi, S.; Benslima, A.; Barragan-Montero, V.; Hajji, M.; Nasri, M. Polyphenolic-protein-polysaccharide ternary conjugates from Cystoseira barbata Tunisian seaweed as potential biopreservatives: Chemical, antioxidant and antimicrobial properties. Int. J. Biol. Macromol. 2017, 105, 1375–1383. [Google Scholar] [CrossRef]
- Trifan, A.; Vasincu, A.; Luca, S.V.; Neophytou, C.; Wolfram, E.; Opitz, S.E.W.; Sava, D.; Bucur, L.; Cioroiu, B.I.; Miron, A.; et al. Unravelling the potential of seaweeds from the Black Sea coast of Romania as bioactive compounds sources. Part I: Cystoseira barbata (Stackhouse) C. Agardh. Food Chem. Toxicol. 2019, 134, 110820. [Google Scholar] [CrossRef]
- Olate-Gallegos, C.; Barriga, A.; Vergara, C.; Fredes, C.; García, P.; Giménez, B.; Robert, P. Identification of polyphenols from Chilean brown seaweeds extracts by LC-DAD-ESI-MS/MS. J. Aquat. Food Prod. Technol. 2019, 28, 375–391. [Google Scholar] [CrossRef]
- Kim, S.M.; Kang, S.W.; Jeon, J.S.; Jung, Y.J.; Kim, W.R.; Kim, C.Y.; Um, B.H. Determination of major phlorotannins in Eisenia bicyclis using hydrophilic interaction chromatography: Seasonal variation and extraction characteristics. Food Chem. 2013, 138, 2399–2406. [Google Scholar] [CrossRef]
- Cho, S.; Yang, H.; Jeon, Y.J.; Lee, C.J.; Jin, Y.H.; Baek, N.I.; Kim, D.; Kang, S.M.; Yoon, M.; Yong, H.; et al. Phlorotannins of the edible brown seaweed Ecklonia cava Kjellman induce sleep via positive allosteric modulation of gamma-aminobutyric acid type A-benzodiazepine receptor: A novel neurological activity of seaweed polyphenols. Food Chem. 2012, 132, 1133–1142. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Ko, J.Y.; Oh, J.Y.; Kim, C.Y.; Lee, H.J.; Kim, J.; Jeon, Y.J. Preparative isolation and purification of phlorotannins from Ecklonia cava using centrifugal partition chromatography by one-step. Food Chem. 2014, 158, 433–437. [Google Scholar] [CrossRef] [PubMed]
- Nho, J.A.; Shin, Y.S.; Jeong, H.-R.; Cho, S.; Heo, H.J.; Kim, G.H.; Kim, D.-O. Neuroprotective effects of phlorotannin-rich extract from brown seaweed Ecklonia cava on neuronal PC-12 and SH-SY5Y Cells with Oxidative Stress. J. Microbiol. Biotechnol. 2020, 30, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Wei, R.; Lee, M.S.; Lee, B.; Oh, C.W.; Choi, C.G.; Kim, H.R. Isolation and identification of anti-inflammatory compounds from ethyl acetate fraction of Ecklonia stolonifera and their anti-inflammatory action. J. Appl. Phycol. 2016, 28, 3535–3545. [Google Scholar] [CrossRef]
- Kirke, D.A.; Smyth, T.J.; Rai, D.K.; Kenny, O.; Stengel, D.B. The chemical and antioxidant stability of isolated low molecular weight phlorotannins. Food Chem. 2017, 221, 1104–1112. [Google Scholar] [CrossRef]
- Wang, T.; Jónsdóttir, R.; Liu, H.; Gu, L.; Kristinsson, H.G.; Raghavan, S.; Ólafsdóttir, G. Antioxidant capacities of phlorotannins extracted from the brown algae Fucus vesiculosus. J. Agric. Food Chem. 2012, 60, 5874–5883. [Google Scholar] [CrossRef]
- Catarino, M.D.; Silva, A.M.S.; Mateus, N.; Cardoso, S.M. Optimization of phlorotannins extraction from Fucus vesiculosus and evaluation of their potential to prevent metabolic disorders. Mar. Drugs 2019, 17, 162. [Google Scholar] [CrossRef]
- Le Lann, K.; Surget, G.; Couteau, C.; Coiffard, L.; Cérantola, S.; Gaillard, F.; Larnicol, M.; Zubia, M.; Guérard, F.; Poupart, N.; et al. Sunscreen, antioxidant, and bactericide capacities of phlorotannins from the brown macroalga Halidrys siliquosa. J. Appl. Phycol. 2016, 28, 3547–3559. [Google Scholar] [CrossRef]
- Haraguchi, K.; Kotaki, Y.; Relox, J.R.; Romero, M.L.J.; Terada, R. Monitoring of naturally produced brominated phenoxyphenols and phenoxyanisoles in aquatic plants from the Philippines. J. Agric. Food Chem. 2010, 58, 12385–12391. [Google Scholar] [CrossRef] [PubMed]
- Vissers, A.M.; Caligiani, A.; Sforza, S.; Vincken, J.P.; Gruppen, H. Phlorotannin Composition of Laminaria digitata. Phytochem. Anal. 2017, 28, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Guo, S.; Jia, X.; Li, X.; Shi, D. Phytochemical and chemotaxonomic study on Leathesia nana (Chordariaceae). Biochem. Syst. Ecol. 2018, 81, 42–44. [Google Scholar] [CrossRef]
- Nair, D.; Vanuopadath, M.; Balasubramanian, A.; Iyer, A.; Ganesh, S.; Anil, A.N.; Vikraman, V.; Pillai, P.; Bose, C.; Nair, B.G.; et al. Phlorotannins from Padina tetrastromatica: Structural characterisation and functional studies. J. Appl. Phycol. 2019, 31, 3131–3141. [Google Scholar] [CrossRef]
- Li, Y.; Fu, X.; Duan, D.; Liu, X.; Xu, J.; Gao, X. Extraction and identification of phlorotannins from the brown aqlga, Sargassum fusiforme (Harvey) Setchell. Mar. Drugs 2017, 15, 49. [Google Scholar] [CrossRef]
- Montero, L.; Sánchez-Camargo, A.P.; García-Cañas, V.; Tanniou, A.; Stiger-Pouvreau, V.; Russo, M.; Rastrelli, L.; Cifuentes, A.; Herrero, M.; Ibáñez, E. Anti-proliferative activity and chemical characterization by comprehensive two-dimensional liquid chromatography coupled to mass spectrometry of phlorotannins from the brown macroalga Sargassum muticum collected on North-Atlantic coasts. J. Chromatogr. A 2016, 1428, 115–125. [Google Scholar] [CrossRef]
- Vázquez-Rodríguez, B.; Gutiérrez-Uribe, J.A.; Antunes-Ricardo, M.; Santos-Zea, L.; Cruz-Suárez, L.E. Ultrasound-assisted extraction of phlorotannins and polysaccharides from Silvetia compressa (Phaeophyceae). J. Appl. Phycol. 2020, 32, 1441–1453. [Google Scholar] [CrossRef]
- Murugan, K.; Iyer, V.V. Differential growth inhibition of cancer cell lines and antioxidant activity of extracts of red, brown, and green marine algae. Vitr. Cell. Dev. Biol. - Anim. 2013, 49, 324–334. [Google Scholar] [CrossRef]
- Aravindan, S.; Delma, C.R.; Thirugnanasambandan, S.S.; Herman, T.S.; Aravindan, N. Anti-pancreatic cancer deliverables from sea: First-hand evidence on the efficacy, molecular targets and mode of action for multifarious polyphenols from five different brown-algae. PLoS ONE 2013, 8, e61977. [Google Scholar] [CrossRef]
- Lopes, G.; Sousa, C.; Silva, L.R.; Pinto, E.; Andrade, P.B.; Bernardo, J.; Mouga, T.; Valentão, P. Can phlorotannins purified extracts constitute a novel pharmacological alternative for microbial infections with associated inflammatory conditions? PLoS ONE 2012, 7, e31145. [Google Scholar] [CrossRef] [PubMed]
- Audibert, L.; Fauchon, M.; Blanc, N.; Hauchard, D.; Ar Gall, E. Phenolic compounds in the brown seaweed Ascophyllum nodosum: Distribution and radical-scavenging activities. Phytochem. Anal. 2010, 21, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Kurth, C.; Welling, M.; Pohnert, G. Sulfated phenolic acids from Dasycladales siphonous green algae. Phytochemistry 2015, 117, 417–423. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, A.L.L.; Da Silva, D.B.; Lopes, N.P.; Debonsi, H.M.; Yokoya, N.S. Chemical constituents from red algae Bostrychia radicans (Rhodomelaceae): New amides and phenolic compounds. Quim. Nova 2012, 35, 2186–2188. [Google Scholar] [CrossRef]
- Dahlgren, E.; Lindqvist, D.; Dahlgren, H.; Asplund, L.; Lehtilä, K. Trophic transfer of naturally produced brominated aromatic compounds in a Baltic Sea food chain. Chemosphere 2016, 144, 1597–1604. [Google Scholar] [CrossRef] [PubMed]
- Lindqvist, D.; Dahlgren, E.; Asplund, L. Biosynthesis of hydroxylated polybrominated diphenyl ethers and the correlation with photosynthetic pigments in the red alga Ceramium tenuicorne. Phytochemistry 2017, 133, 51–58. [Google Scholar] [CrossRef]
- Mikami, D.; Kurihara, H.; Kim, S.M.; Takahashi, K. Red algal bromophenols as glucose 6-phosphate dehydrogenase inhibitors. Mar. Drugs 2013, 11, 4050–4057. [Google Scholar] [CrossRef]
- Mikami, D.; Kurihara, H.; Ono, M.; Kim, S.M.; Takahashi, K. Inhibition of algal bromophenols and their related phenols against glucose 6-phosphate dehydrogenase. Fitoterapia 2016, 108, 20–25. [Google Scholar] [CrossRef]
- Islam, M.R.; Mikami, D.; Kurihara, H. Two new algal bromophenols from Odonthalia corymbifera. Tetrahedron Lett. 2017, 58, 4119–4121. [Google Scholar] [CrossRef]
- Lever, J.; Curtis, G.; Brkljača, R.; Urban, S. Bromophenolics from the reed alga Polysiphonia decipiens. Mar. Drugs 2019, 17, 497. [Google Scholar] [CrossRef]
- Kim, S.Y.; Kim, S.R.; Oh, M.J.; Jung, S.J.; Kang, S.Y. In Vitro antiviral activity of red alga, Polysiphonia morrowii extract and its bromophenols against fish pathogenic infectious hematopoietic necrosis virus and infectious pancreatic necrosis virus. J. Microbiol. 2011, 49, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.K.; Ye, B.R.; Kim, E.A.; Kim, J.; Kim, M.S.; Lee, W.W.; Ahn, G.N.; Kang, N.; Jung, W.K.; Heo, S.J. Bis (3-bromo-4,5-dihydroxybenzyl) ether, a novel bromophenol from the marine red alga Polysiphonia morrowii that suppresses LPS-induced inflammatory response by inhibiting ROS-mediated ERK signaling pathway in RAW 264.7 macrophages. Biomed. Pharmacother. 2018, 103, 1170–1177. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Li, X.M.; Gloer, J.B.; Wang, B.G. New nitrogen-containing bromophenols from the marine red alga Rhodomela confervoides and their radical scavenging activity. Food Chem. 2012, 135, 868–872. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, X.; Gao, L.; Cui, C.; Li, C.; Li, J.; Wang, B. Extraction and PTP1B inhibitory activity of bromophenols from the marine red alga Symphyocladia latiuscula. Chinese J. Oceanol. Limnol. 2011, 29, 686–690. [Google Scholar] [CrossRef]
- Xu, X.; Yin, L.; Gao, L.; Gao, J.; Chen, J.; Li, J.; Song, F. Two new bromophenols with radical scavenging activity from marine red alga Symphyocladia latiuscula. Mar. Drugs 2013, 11, 842–847. [Google Scholar] [CrossRef]
- Xu, X.; Yin, L.; Gao, J.; Gao, L.; Song, F. Antifungal bromophenols from marine red alga Symphyocladia latiuscula. Chem. Biodivers. 2014, 11, 807–811. [Google Scholar] [CrossRef]
- Xu, X.; Yang, H.; Khalil, Z.G.; Yin, L.; Xiao, X.; Neupane, P.; Bernhardt, P.V.; Salim, A.A.; Song, F.; Capon, R.J. Chemical diversity from a Chinese marine red alga, Symphyocladia latiuscula. Mar. Drugs 2017, 15, 374. [Google Scholar] [CrossRef]
- Xu, X.; Yang, H.; Khalil, Z.G.; Yin, L.; Xiao, X.; Salim, A.A.; Song, F.; Capon, R.J. Bromocatechol conjugates from a Chinese marine red alga, Symphyocladia latiuscula. Phytochemistry 2019, 158, 20–25. [Google Scholar] [CrossRef]
- Paudel, P.; Seong, S.H.; Zhou, Y.; Park, H.J.; Jung, H.A.; Choi, J.S. Anti-Alzheimer’s Disease Activity of Bromophenols from a Red Alga, Symphyocladia latiuscula (Harvey) Yamada. ACS Omega 2019, 4, 12259–12270. [Google Scholar] [CrossRef]
- Hofer, S.; Hartmann, A.; Orfanoudaki, M.; Ngoc, H.N.; Nagl, M.; Karsten, U.; Heesch, S.; Ganzera, M. Development and validation of an HPLC method for the quantitative analysis of bromophenolic compounds in the red alga Vertebrata lanosa. Mar. Drugs 2019, 17, 675. [Google Scholar] [CrossRef]
- Huang, D.; Boxin, O.U.; Prior, R.L. The chemistry behind antioxidant capacity assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef] [PubMed]
- Tierney, M.S.; Smyth, T.J.; Rai, D.K.; Soler-Vila, A.; Croft, A.K.; Brunton, N. Enrichment of polyphenol contents and antioxidant activities of Irish brown macroalgae using food-friendly techniques based on polarity and molecular size. Food Chem. 2013, 139, 753–761. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.N.; Cheung, P.C.K.; Ooi, V.E.C.; Ang, P.O. Evaluation of antioxidative activity of extracts from a brown seaweed, Sargassum siliquastrum. J. Agric. Food Chem. 2002, 50, 3862–3866. [Google Scholar] [CrossRef] [PubMed]
- Martins, C.D.L.; Ramlov, F.; Nocchi Carneiro, N.P.; Gestinari, L.M.; dos Santos, B.F.; Bento, L.M.; Lhullier, C.; Gouvea, L.; Bastos, E.; Horta, P.A.; et al. Antioxidant properties and total phenolic contents of some tropical seaweeds of the Brazilian coast. J. Appl. Phycol. 2013, 25, 1179–1187. [Google Scholar] [CrossRef]
- Sevimli-Gur, C.; Yesil-Celiktas, O. Cytotoxicity screening of supercritical fluid extracted seaweeds and phenylpropanoids. Mol. Biol. Rep. 2019, 46, 3691–3699. [Google Scholar] [CrossRef] [PubMed]
- Fraga, C.G.; Croft, K.D.; Kennedy, D.O.; Tomás-Barberán, F.A. The effects of polyphenols and other bioactives on human health. Food Funct. 2019, 10, 514–528. [Google Scholar] [CrossRef]
- Williams, R.; Colagiuri, S.; Chan, J.; Gregg, E.W.; Ke, C.; Lim, L.-L.; Yang, X. IDF Atlas, 9th ed.; International Diabetes Federation: Brussels, Belgium, 2019; ISBN 978-2-930229-87-4. [Google Scholar]
- World Health Organization. Global Report on Diabetes. Available online: https://apps.who.int/iris/handle/10665/204871 (accessed on 7 August 2020).
- Hermans, M.P.; Dath, N. Prevalence and co-prevalence of comorbidities in Belgian patients with type 2 diabetes mellitus: A transversal, descriptive study. Acta Clin. Belgica Int. J. Clin. Lab. Med. 2018, 73, 68–74. [Google Scholar] [CrossRef]
- Lee, S.H.; Jeon, Y.J. Anti-diabetic effects of brown algae derived phlorotannins, marine polyphenols through diverse mechanisms. Fitoterapia 2013, 86, 129–136. [Google Scholar] [CrossRef]
- Gunathilaka, T.L.; Samarakoon, K.; Ranasinghe, P.; Peiris, L.D.C. Antidiabetic potential of marine brown algae - A Mini Review. J. Diabetes Res. 2020, 2020. [Google Scholar] [CrossRef]
- Murugan, A.C.; Karim, M.R.; Yusoff, M.B.M.; Tan, S.H.; Asras, M.F.B.F.; Rashid, S.S. New insights into seaweed polyphenols on glucose homeostasis. Pharm. Biol. 2015, 53, 1087–1097. [Google Scholar] [CrossRef]
- Zhao, C.; Yang, C.; Liu, B.; Lin, L.; Sarker, S.D.; Nahar, L.; Yu, H.; Cao, H.; Xiao, J. Bioactive compounds from marine macroalgae and their hypoglycemic benefits. Trends Food Sci. Technol. 2018, 72, 1–12. [Google Scholar] [CrossRef]
- Benalla, W.; Bellahcen, S.; Bnouham, M. Antidiabetic medicinal plants as a source of alpha glucosidase Inhibitors. Curr. Diabetes Rev. 2010, 6, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Yong-Li; Karadeniz, F.; Kim, M.M.; Kim, S.K. α-Glucosidase and α-amylase inhibitory activities of phloroglucinal derivatives from edible marine brown alga, Ecklonia cava. J. Sci. Food Agric. 2009, 89, 1552–1558. [Google Scholar] [CrossRef]
- Nwosu, F.; Morris, J.; Lund, V.A.; Stewart, D.; Ross, H.A.; McDougall, G.J. Anti-proliferative and potential anti-diabetic effects of phenolic-rich extracts from edible marine algae. Food Chem. 2011, 126, 1006–1012. [Google Scholar] [CrossRef]
- Lordan, S.; Smyth, T.J.; Soler-Vila, A.; Stanton, C.; Paul Ross, R. The α-amylase and α-glucosidase inhibitory effects of Irish seaweed extracts. Food Chem. 2013, 141, 2170–2176. [Google Scholar] [CrossRef]
- Kellogg, J.; Grace, M.H.; Lila, M.A. Phlorotannins from alaskan seaweed inhibit carbolytic enzyme activity. Mar. Drugs 2014, 12, 5277–5294. [Google Scholar] [CrossRef]
- Sharifuddin, Y.; Chin, Y.X.; Lim, P.E.; Phang, S.M. Potential bioactive compounds from seaweed for diabetes management. Mar. Drugs 2015, 13, 5447–5491. [Google Scholar] [CrossRef]
- Park, S.R.; Kim, J.H.; Jang, H.D.; Yang, S.Y.; Kim, Y.H. Inhibitory activity of minor phlorotannins from Ecklonia cava on α-glucosidase. Food Chem. 2018, 257, 128–134. [Google Scholar] [CrossRef]
- Ezzat, S.M.; El Bishbishy, M.H.; Habtemariam, S.; Salehi, B.; Sharifi-Rad, M.; Martins, N.; Sharifi-Rad, J. Looking at marine-derived bioactive molecules as upcoming anti-diabetic agents: A special emphasis on PTP1B inhibitors. Molecules 2018, 23, 3334. [Google Scholar] [CrossRef]
- Xu, Q.; Luo, J.; Wu, N.; Zhang, R.; Shi, D. BPN, a marine-derived PTP1B inhibitor, activates insulin signaling and improves insulin resistance in C2C12 myotubes. Int. J. Biol. Macromol. 2018, 106, 379–386. [Google Scholar] [CrossRef]
- Luo, J.; Hou, Y.; Xie, M.; Ma, W.; Shi, D.; Jiang, B. CYC31, A Natural bromophenol PTP1B inhibitor, activates insulin signaling and improves long chain-fatty acid oxidation in C2C12 Myotubes. Mar. Drugs 2020, 18, 267. [Google Scholar] [CrossRef]
- Boyd, A.C.; Abdel-Wahab, Y.H.A.; McKillop, A.M.; McNulty, H.; Barnett, C.R.; O’Harte, F.P.M.; Flatt, P.R. Impaired ability of glycated insulin to regulate plasma glucose and stimulate glucose transport and metabolism in mouse abdominal muscle. Biochim. Biophys. Acta Gen. Subj. 2000, 1523, 128–134. [Google Scholar] [CrossRef]
- Iannuzzi, C.; Borriello, M.; Carafa, V.; Altucci, L.; Vitiello, M.; Balestrieri, M.L.; Ricci, G.; Irace, G.; Sirangelo, I. D-ribose-glycation of insulin prevents amyloid aggregation and produces cytotoxic adducts. Biochim. Biophys. Acta Mol. Basis Dis. 2016, 1862, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.P.; Bali, A.; Singh, N.; Jaggi, A.S. Advanced glycation end products and diabetic complications. Korean J. Physiol. Pharmacol. 2014, 18, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, S.; Minami, Y.; Taniguchi, R.; Tanaka, R.; Miyake, H.; Mori, T.; Ueda, M.; Shibata, T. Evaluation of anti-glycation activities of phlorotannins in human and bovine serum albumin-methylglyoxal models. Nat. Prod. Commun. 2017, 12, 1793–1796. [Google Scholar] [CrossRef]
- Shakambari, G.; Ashokkumar, B.; Varalakshmi, P. Phlorotannins from Brown Algae: Inhibition of advanced glycation end products formation in high glucose induced Caenorhabditis elegans. Indian J. Exp. Biol. 2015, 53, 371–379. [Google Scholar]
- Seong, S.H.; Paudel, P.; Jung, H.A.; Choi, J.S. Identifying phlorofucofuroeckol-A as a dual inhibitor of amyloid-β25-35 self-aggregation and insulin glycation: Elucidation of the molecular mechanism of action. Mar. Drugs 2019, 17, 600. [Google Scholar] [CrossRef]
- Malin, S.K.; Finnegan, S.; Fealy, C.E.; Filion, J.; Rocco, M.B.; Kirwan, J.P. Β-Cell dysfunction is associated with metabolic syndrome severity in adults. Metab. Syndr. Relat. Disord. 2014, 12, 79–85. [Google Scholar] [CrossRef]
- Lee, S.H.; Kang, S.M.; Ko, S.C.; Kang, M.C.; Jeon, Y.J. Octaphlorethol A, a novel phenolic compound isolated from Ishige foliacea, protects against streptozotocin-induced pancreatic β cell damage by reducing oxidative stress and apoptosis. Food Chem. Toxicol. 2013, 59, 643–649. [Google Scholar] [CrossRef]
- Kim, E.A.; Kang, M.C.; Lee, J.H.; Kang, N.; Lee, W.; Oh, J.Y.; Yang, H.W.; Lee, J.S.; Jeon, Y.J. Protective effect of marine brown algal polyphenols against oxidative stressed zebrafish with high glucose. RSC Adv. 2015, 5, 25738–25746. [Google Scholar] [CrossRef]
- Cha, S.H.; Kim, H.S.; Hwang, Y.; Jeon, Y.J.; Jun, H.S. Polysiphonia japonica extract attenuates palmitate-induced toxicity and enhances insulin secretion in pancreatic beta-cells. Oxid. Med. Cell. Longev. 2018, 2018. [Google Scholar] [CrossRef] [PubMed]
- Sheetz, M.J.; King, G.L. Molecular understanding of hyperglycemia’s adverse effects for diabetic complications. J. Am. Med. Assoc. 2002, 288, 2579–2588. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Kang, S.M.; Ko, S.C.; Lee, D.H.; Jeon, Y.J. Octaphlorethol A, a novel phenolic compound isolated from a brown alga, Ishige foliacea, increases glucose transporter 4-mediated glucose uptake in skeletal muscle cells. Biochem. Biophys. Res. Commun. 2012, 420, 576–581. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Wang, Q.; Ling, Z.; Pipeleers, D.; McDermott, P.; Pende, M.; Heimberg, H.; Van de Casteele, M. Akt activation protects pancreatic beta cells from AMPK-mediated death through stimulation of mTOR. Biochem. Pharmacol. 2008, 75, 1981–1993. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Park, M.H.; Heo, S.J.; Kang, S.M.; Ko, S.C.; Han, J.S.; Jeon, Y.J. Dieckol isolated from Ecklonia cava inhibits α-glucosidase and α-amylase in vitro and alleviates postprandial hyperglycemia in streptozotocin-induced diabetic mice. Food Chem. Toxicol. 2010, 48, 2633–2637. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.A.; Lee, J.H.; Han, J.S. A phlorotannin constituent of Ecklonia cava alleviates postprandial hyperglycemia in diabetic mice. Pharm. Biol. 2017, 55, 1149–1154. [Google Scholar] [CrossRef]
- Murray, M.; Dordevic, A.L.; Ryan, L.; Bonham, M.P. The impact of a single dose of a polyphenol-rich seaweed extract on postprandial glycaemic control in healthy adults: A randomised cross-over trial. Nutrients 2018, 10, 270. [Google Scholar] [CrossRef]
- Kang, M.C.; Wijesinghe, W.A.J.P.; Lee, S.H.; Kang, S.M.; Ko, S.C.; Yang, X.; Kang, N.; Jeon, B.T.; Kim, J.; Lee, D.H.; et al. Dieckol isolated from brown seaweed Ecklonia cava attenuates type II diabetes in db/db mouse model. Food Chem. Toxicol. 2013, 53, 294–298. [Google Scholar] [CrossRef]
- Yan, X.; Yang, C.; Lin, G.; Chen, Y.; Miao, S.; Liu, B.; Zhao, C. Antidiabetic potential of green seaweed Enteromorpha prolifera flavonoids regulating insulin signaling pathway and gut microbiota in Type 2 Diabetic Mice. J. Food Sci. 2019, 84, 165–173. [Google Scholar] [CrossRef]
- Lee, S.H.; Jeon, Y.J. Efficacy and safety of a dieckol-rich extract (AG-dieckol) of brown algae, Ecklonia cava, in pre-diabetic individuals: A double-blind, randomized, placebo-controlled clinical trial. Food Funct. 2015, 6, 853–858. [Google Scholar] [CrossRef]
- Cavalot, F.; Petrelli, A.; Traversa, M.; Bonomo, K.; Fiora, E.; Conti, M.; Anfossi, G.; Costa, G.; Trovati, M. Postprandial blood glucose is a stronger predictor of cardiovascular events than fasting blood glucose in type 2 diabetes mellitus, particularly in women: Lessons from the San Luigi Gonzaga diabetes study. J. Clin. Endocrinol. Metab. 2006, 91, 813–819. [Google Scholar] [CrossRef] [PubMed]
- Abdali, D.; Samson, S.E.; Grover, A.K. How effective are antioxidant supplements in obesity and diabetes? Med. Princ. Pract. 2015, 24, 201–215. [Google Scholar] [CrossRef] [PubMed]
- Gregor, M.F.; Hotamisligil, G.S. Inflammatory mechanisms in obesity. Annu. Rev. Immunol. 2011, 29, 415–445. [Google Scholar] [CrossRef] [PubMed]
- Jung, U.J.; Choi, M.S. Obesity and its metabolic complications: The role of adipokines and the relationship between obesity, inflammation, insulin resistance, dyslipidemia and nonalcoholic fatty liver disease. Int. J. Mol. Sci. 2014, 15, 6184–6223. [Google Scholar] [CrossRef]
- Marseglia, L.; Manti, S.; D’Angelo, G.; Nicotera, A.; Parisi, E.; Di Rosa, G.; Gitto, E.; Arrigo, T. Oxidative stress in obesity: A critical component in human diseases. Int. J. Mol. Sci. 2015, 16, 378–400. [Google Scholar] [CrossRef]
- Eom, S.H.; Lee, M.S.; Lee, E.W.; Kim, Y.M.; Kim, T.H. Pancreatic lipase inhibitory activity of phlorotannins isolated from Eisenia bicyclis. Phyther. Res. 2013, 27, 148–151. [Google Scholar] [CrossRef]
- Austin, C.; Stewart, D.; Allwood, J.W.; McDougall, G.J. Extracts from the edible seaweed Ascophyllum nodosum, inhibit lipase activity in vitro: Contributions of phenolic and polysaccharide components. Food Funct. 2018, 9, 502–510. [Google Scholar] [CrossRef]
- Franssen, R.; Monajemi, H.; Stroes, E.S.G.; Kastelein, J.J.P. Obesity and dyslipidemia. Med. Clin. N. Am. 2011, 95, 893–902. [Google Scholar] [CrossRef]
- Ko, S.C.; Lee, M.; Lee, J.H.; Lee, S.H.; Lim, Y.; Jeon, Y.J. Dieckol, a phlorotannin isolated from a brown seaweed, Ecklonia cava, inhibits adipogenesis through AMP-activated protein kinase (AMPK) activation in 3T3-L1 preadipocytes. Environ. Toxicol. Pharmacol. 2013, 36, 1253–1260. [Google Scholar] [CrossRef]
- Jung, H.A.; Jung, H.J.; Jeong, H.Y.; Kwon, H.J.; Ali, M.Y.; Choi, J.S. Phlorotannins isolated from the edible brown alga Ecklonia stolonifera exert anti-adipogenic activity on 3T3-L1 adipocytes by downregulating C/EBPα and PPARγ. Fitoterapia 2014, 92, 260–269. [Google Scholar] [CrossRef]
- Kwon, T.H.; Wu, Y.X.; Kim, J.S.; Woo, J.H.; Park, K.T.; Kwon, O.J.; Seo, H.J.; Kim, T.; Park, N.H. 6,6′-Bieckol inhibits adipocyte differentiation through downregulation of adipogenesis and lipogenesis in 3T3-L1 cells. J. Sci. Food Agric. 2015, 95, 1830–1837. [Google Scholar] [CrossRef] [PubMed]
- Kong, C.S.; Kim, H.; Seo, Y. Edible brown alga Ecklonia cava derived phlorotannin-induced anti-adipogenic activity in vitro. J. Food Biochem. 2015, 39, 1–10. [Google Scholar] [CrossRef]
- Salas, A.; Noé, V.; Ciudad, C.J.; Romero, M.M.; Remesar, X.; Esteve, M. Short-term oleoyl-estrone treatment affects capacity to manage lipids in rat adipose tissue. BMC Genom. 2007, 8, 292. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo, A.G.; Crujeiras, A.B.; Casanueva, F.F.; Carreira, M.C. Leptin, obesity, and leptin resistance: Where are we 25 years later? Nutrients 2019, 11, 2704. [Google Scholar] [CrossRef]
- Kim, I.H.; Nam, T.J. Enzyme-treated Ecklonia cava extract inhibits adipogenesis through the downregulation of C/EBPα in 3T3-L1 adipocytes. Int. J. Mol. Med. 2017, 39, 636–644. [Google Scholar] [CrossRef]
- Karadeniz, F.; Ahn, B.N.; Kim, J.A.; Seo, Y.; Jang, M.S.; Nam, K.H.; Kim, M.; Lee, S.H.; Kong, C.S. Phlorotannins suppress adipogenesis in pre-adipocytes while enhancing osteoblastogenesis in pre-osteoblasts. Arch. Pharm. Res. 2015, 38, 2172–2182. [Google Scholar] [CrossRef]
- Choi, H.S.; Jeon, H.J.; Lee, O.H.; Lee, B.Y. Dieckol, a major phlorotannin in Ecklonia cava, suppresses lipid accumulation in the adipocytes of high-fat diet-fed zebrafish and mice: Inhibition of early adipogenesis via cell-cycle arrest and AMPKα activation. Mol. Nutr. Food Res. 2015, 59, 1458–1471. [Google Scholar] [CrossRef]
- Ko, S.C.; Ding, Y.; Kim, J.; Ye, B.R.; Kim, E.A.; Jung, W.K.; Heo, S.J.; Lee, S.H. Bromophenol (5-bromo-3,4-dihydroxybenzaldehyde) isolated from red alga Polysiphonia morrowii inhibits adipogenesis by regulating expression of adipogenic transcription factors and AMP-activated protein kinase activation in 3T3-L1 adipocytes. Phyther. Res. 2019, 33, 737–744. [Google Scholar] [CrossRef]
- Kellogg, J.; Esposito, D.; Grace, M.H.; Komarnytsky, S.; Lila, M.A. Alaskan seaweeds lower inflammation in RAW 264.7 macrophages and decrease lipid accumulation in 3T3-L1 adipocytes. J. Funct. Foods 2015, 15, 396–407. [Google Scholar] [CrossRef]
- Song, F.; Del Pozo, C.H.; Rosario, R.; Zou, Y.S.; Ananthakrishnan, R.; Xu, X.; Patel, P.R.; Benoit, V.M.; Yan, S.F.; Li, H.; et al. RAGE regulates the metabolic and inflammatory response to high-fat feeding in mice. Diabetes 2014, 63, 1948–1965. [Google Scholar] [CrossRef]
- Choi, J.; Oh, S.; Son, M.; Byun, K. Pyrogallol-phloroglucinol-6,6-bieckol alleviates obesity and systemic inflammation in a mouse model by reducing expression of RAGE and RAGE ligands. Mar. Drugs 2019, 17, 612. [Google Scholar] [CrossRef] [PubMed]
- Yeo, A.R.; Lee, J.; Tae, I.H.; Park, S.R.; Cho, Y.H.; Lee, B.H.; Cheol Shin, H.; Kim, S.H.; Yoo, Y.C. Anti-hyperlipidemic effect of polyphenol extract (SeapolynolTM) and dieckol isolated from Ecklonia cava in in vivo and in vitro models. Prev. Nutr. Food Sci. 2012, 17, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Park, E.Y.; Kim, E.H.; Kim, M.H.; Seo, Y.W.; Lee, J.I.; Jun, H.S. Polyphenol-rich fraction of brown alga Ecklonia cava collected from Gijang, Korea, reduces obesity and glucose levels in high-fat diet-induced obese mice. Evidence-based Complement. Altern. Med. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Eo, H.; Jeon, Y.J.; Lee, M.; Lim, Y. Brown alga Ecklonia cava polyphenol extract ameliorates hepatic lipogenesis, oxidative stress, and inflammation by activation of AMPK and SIRT1 in high-fat diet-induced obese mice. J. Agric. Food Chem. 2015, 63, 349–359. [Google Scholar] [CrossRef]
- Ding, Y.; Wang, L.; Im, S.; Hwang, O.; Kim, H.S.; Kang, M.C.; Lee, S.H. Anti-obesity effect of diphlorethohydroxycarmalol isolated from brown alga Ishige okamurae in high-fat diet-induced obese mice. Mar. Drugs 2019, 17, 637. [Google Scholar] [CrossRef]
- Shin, H.C.; Kim, S.H.; Park, Y.; Lee, B.H.; Hwang, H.J. Effects of 12-week oral supplementation of Ecklonia cava polyphenols on anthropometric and blood lipid parameters in overweight Korean individuals: A double-blind randomized clinical trial. Phyther. Res. 2012, 26, 363–368. [Google Scholar] [CrossRef]
- Samson, S.L.; Garber, A.J. Metabolic syndrome. Endocrinol. Metab. Clin. N. Am. 2014, 43, 1–23. [Google Scholar] [CrossRef]
- Alberti, K.G.M.M.; Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z.; Cleeman, J.I.; Donato, K.A.; Fruchart, J.C.; James, W.P.T.; Loria, C.M.; Smith, S.C. Harmonizing the metabolic syndrome: A joint interim statement of the international diabetes federation task force on epidemiology and prevention; National Heart, Lung, and Blood Institute; American Heart Association; World Heart Federation; International. Circulation 2009, 120, 1640–1645. [Google Scholar] [CrossRef]
- Stefanska, A.; Bergmann, K.; Sypniewska, G. Metabolic Syndrome and Menopause. In Advances in Clinical Chemistry; Elsevier Inc.: Amsterdam, The Netherlands, 2015; Volume 72, pp. 1–75. ISBN 9780128033142. [Google Scholar]
- Park, J.K.; Woo, H.W.; Kim, M.K.; Shin, J.; Lee, Y.H.; Shin, D.H.; Shin, M.H.; Choi, B.Y. Dietary iodine, seaweed consumption, and incidence risk of metabolic syndrome among postmenopausal women: A prospective analysis of the Korean Multi-Rural Communities Cohort Study (MRCohort). Eur. J. Nutr. 2020. [Google Scholar] [CrossRef]
- Vijayan, R.; Chitra, L.; Penislusshiyan, S.; Palvannan, T. Exploring bioactive fraction of Sargassum wightii: In vitro elucidation of angiotensin-i-converting enzyme inhibition and antioxidant potential. Int. J. Food Prop. 2018, 21, 674–684. [Google Scholar] [CrossRef]
- Son, M.; Oh, S.; Lee, H.S.; Ryu, B.M.; Jiang, Y.; Jang, J.T.; Jeon, Y.J.; Byun, K. Pyrogallol-phloroglucinol-6,6’-bieckol from Ecklonia cava improved blood circulation in diet-induced obese and diet-induced hypertension mouse models. Mar. Drugs 2019, 17, 272. [Google Scholar] [CrossRef]
- Kammoun, I.; Ben Salah, H.; Ben Saad, H.; Cherif, B.; Droguet, M.; Magné, C.; Kallel, C.; Boudawara, O.; Hakim, A.; Gharsallah, N.; et al. Hypolipidemic and cardioprotective effects of Ulva lactuca ethanolic extract in hypercholesterolemic mice. Arch. Physiol. Biochem. 2018, 124, 313–325. [Google Scholar] [CrossRef] [PubMed]
- Wanyonyi, S.; Du Preez, R.; Brown, L.; Paul, N.A.; Panchal, S.K. Kappaphycus alvarezii as a food supplement prevents diet-induced metabolic syndrome in rats. Nutrients 2017, 9, 1261. [Google Scholar] [CrossRef]
- Gómez-Guzmán, M.; Rodríguez-Nogales, A.; Algieri, F.; Gálvez, J. Potential role of seaweed polyphenols in cardiovascular-associated disorders. Mar. Drugs 2018, 16, 250. [Google Scholar] [CrossRef]
- Alam, T.; Khan, S.; Gaba, B.; Haider, M.F.; Baboota, S.; Ali, J. Nanocarriers as treatment modalities for hypertension. Drug Deliv. 2017, 24, 358–369. [Google Scholar] [CrossRef]
- Seca, A.M.L.; Pinto, D.C.G.A. Overview on the antihypertensive and anti-obesity effects of secondary metabolites from seaweeds. Mar. Drugs 2018, 16, 237. [Google Scholar] [CrossRef] [PubMed]
- Olasehinde, T.A.; Olaniran, A.O.; Okoh, A.I. Macroalgae as a valuable source of naturally occurring bioactive compounds for the treatment of Alzheimer’s disease. Mar. Drugs 2019, 17, 609. [Google Scholar] [CrossRef] [PubMed]
- Gouras, G.K.; Olsson, T.T.; Hansson, O. β-amyloid peptides and amyloid plaques in Alzheimer’s disease. Neurotherapeutics 2015, 12, 3–11. [Google Scholar] [CrossRef]
- Frozza, R.L.; Lourenco, M.V.; de Felice, F.G. Challenges for Alzheimer’s disease therapy: Insights from novel mechanisms beyond memory defects. Front. Neurosci. 2018, 12, 1–13. [Google Scholar] [CrossRef]
- Cai, H.; Wang, Y.; McCarthy, D.; Wen, H.; Borchelt, D.R.; Price, D.L.; Wong, P.C. BACE1 is the major β-secretase for generation of Aβ peptides by neurons. Nat. Neurosci. 2001, 4, 233–234. [Google Scholar] [CrossRef]
- Choi, B.W.; Lee, H.S.; Shin, H.C.; Lee, B.H. Multifunctional activity of polyphenolic compounds associated with a potential for Alzheimer’s disease therapy from ecklonia cava. Phyther. Res. 2015, 29, 549–553. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.S.; Haulader, S.; Karki, S.; Jung, H.J.; Kim, H.R.; Jung, H.A. Acetyl- and butyryl-cholinesterase inhibitory activities of the edible brown alga Eisenia bicyclis. Arch. Pharm. Res. 2015, 38, 1477–1487. [Google Scholar] [CrossRef] [PubMed]
- Olasehinde, T.A.; Olaniran, A.O.; Okoh, A.I. Phenolic composition, antioxidant activity, anticholinesterase potential and modulatory effects of aqueous extracts of some seaweeds on β-amyloid aggregation and disaggregation. Pharm. Biol. 2019, 57, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Olasehinde, T.A.; Olaniran, A.O.; Okoh, A.I. Aqueous–ethanol extracts of some South African seaweeds inhibit beta-amyloid aggregation, cholinesterases, and beta-secretase activities in vitro. J. Food Biochem. 2019, 43, 1–10. [Google Scholar] [CrossRef]
- Kim, J.J.; Kang, Y.J.; Shin, S.A.; Bak, D.H.; Lee, J.W.; Lee, K.B.; Yoo, Y.C.; Kim, D.K.; Lee, B.H.; Kim, D.W.; et al. Phlorofucofuroeckol improves glutamate- induced neurotoxicity through modulation of oxidative stress-mediated mitochondrial dysfunction in PC12 cells. PLoS ONE 2016, 11, e0163433. [Google Scholar] [CrossRef]
- Yang, E.J.; Ahn, S.; Ryu, J.; Choi, M.S.; Choi, S.; Chong, Y.H.; Hyun, J.W.; Chang, M.J.; Kim, H.S. Phloroglucinol attenuates the cognitive deficits of the 5XFAD mouse model of Alzheimer’s disease. PLoS ONE 2015, 10, e0135686. [Google Scholar] [CrossRef]
- Wang, J.; Zheng, J.; Huang, C.; Zhao, J.; Lin, J.; Zhou, X.; Naman, C.B.; Wang, N.; Gerwick, W.H.; Wang, Q.; et al. Eckmaxol, a phlorotannin extracted from Ecklonia maxima, produces anti-β-amyloid oligomer neuroprotective effects possibly via directly acting on glycogen synthase kinase 3β. ACS Chem. Neurosci. 2018, 9, 1349–1356. [Google Scholar] [CrossRef]
- Lee, S.; Youn, K.; Kim, D.H.; Ahn, M.R.; Yoon, E.; Kim, O.Y.; Jun, M. Anti-neuroinflammatory property of phlorotannins from Ecklonia cava on Aβ 25-35 -induced damage in PC12 cells. Mar. Drugs 2019, 17, 7. [Google Scholar] [CrossRef]
- Um, M.Y.; Lim, D.W.; Son, H.J.; Cho, S.; Lee, C. Phlorotannin-rich fraction from Ishige foliacea brown seaweed prevents the scopolamine-induced memory impairment via regulation of ERK-CREB-BDNF pathway. J. Funct. Foods 2018, 40, 110–116. [Google Scholar] [CrossRef]
- Lesné, S.; Ali, C.; Gabriel, C.; Croci, N.; MacKenzie, E.T.; Glabe, C.G.; Plotkine, M.; Marchand-Verrecchia, C.; Vivien, D.; Buisson, A. NMDA receptor activation inhibits α-secretase and promotes neuronal amyloid-β production. J. Neurosci. 2005, 25, 9367–9377. [Google Scholar] [CrossRef]
- Figueira, I.; Garcia, G.; Pimpão, R.C.; Terrasso, A.P.; Costa, I.; Almeida, A.F.; Tavares, L.; Pais, T.F.; Pinto, P.; Ventura, M.R.; et al. Polyphenols journey through blood-brain barrier towards neuronal protection. Sci. Rep. 2017, 7, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Schram, F.R.; Ng, P.K.L. What is cancer? J. Crustac. Biol. 2012, 32, 665–672. [Google Scholar] [CrossRef]
- Gutiérrez-Rodríguez, A.G.; Juárez-Portilla, C.; Olivares-Bañuelos, T.; Zepeda, R.C. Anticancer activity of seaweeds. Drug Discov. Today 2018, 23, 434–447. [Google Scholar] [CrossRef]
- Matulja, D.; Wittine, K.; Malatesti, N.; Laclef, S.; Turks, M.; Markovic, M.K.; Ambrožić, G.; Marković, D. marine natural products with high anticancer activities. Curr. Med. Chem. 2020, 27, 1243–1307. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Lee, J.; Oh, J.H.; Chang, H.J.; Sohn, D.K.; Shin, A.; Kim, J. Associations among dietary seaweed intake, c-MYC rs6983267 polymorphism, and risk of colorectal cancer in a Korean population: A case–control study. Eur. J. Nutr. 2020, 59, 1963–1974. [Google Scholar] [CrossRef] [PubMed]
- Mhadhebi, L.; Mhadhebi, A.; Robert, J.; Bouraoui, A. Antioxidant, anti-inflammatory and antiproliferative effects of aqueous extracts of three mediterranean brown seaweeds of the Genus Cystoseira. Iran. J. Pharm. Res. 2014, 13, 207–220. [Google Scholar] [CrossRef]
- Corona, G.; Coman, M.M.; Spencer, J.P.E.; Rowland, I. Digested and fermented seaweed phlorotannins reduce DNA damage and inhibit growth of HT-29 colon cancer cells. Proc. Nutr. Soc. 2014, 73, 262519. [Google Scholar] [CrossRef]
- Mwangi, H.M.; Njue, W.M.; Onani, M.O.; Thovhoghi, N.; Mabusela, W.T. Phlorotannins and a sterol isolated from a brown alga Ecklonia maxima, and their cytotoxic activity against selected cancer cell lines HeLa, H157 and MCF7. Interdiscip. J. Chem. 2017, 2, 1–6. [Google Scholar] [CrossRef]
- Namvar, F.; Baharara, J.; Mahdi, A.A. Antioxidant and anticancer activities of selected Persian Gulf algae. Indian J. Clin. Biochem. 2014, 29, 13–20. [Google Scholar] [CrossRef]
- Lopes-Costa, E.; Abreu, M.; Gargiulo, D.; Rocha, E.; Ramos, A.A. Anticancer effects of seaweed compounds fucoxanthin and phloroglucinol, alone and in combination with 5-fluorouracil in colon cells. J. Toxicol. Environ. Heal. Part A Curr. Issues 2017, 80, 776–787. [Google Scholar] [CrossRef]
- Arai, M.; Shin, D.; Kamiya, K.; Ishida, R.; Setiawan, A.; Kotoku, N.; Kobayashi, M. Marine spongean polybrominated diphenyl ethers, selective growth inhibitors against the cancer cells adapted to glucose starvation, inhibits mitochondrial complex II. J. Nat. Med. 2017, 71, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Bernardini, G.; Minetti, M.; Polizzotto, G.; Biazzo, M.; Santucci, A. Pro-Apoptotic activity of French polynesian Padina pavonica extract on human osteosarcoma cells. Mar. Drugs 2018, 16, 504. [Google Scholar] [CrossRef] [PubMed]
- Kosanić, M.; Ranković, B.; Stanojković, T. Brown macroalgae from the Adriatic Sea as a promising source of bioactive nutrients. J. Food Meas. Charact. 2019, 13, 330–338. [Google Scholar] [CrossRef]
- Abdelhamid, A.; Lajili, S.; Elkaibi, M.A.; Ben Salem, Y.; Abdelhamid, A.; Muller, C.D.; Majdoub, H.; Kraiem, J.; Bouraoui, A. Optimized extraction, preliminary characterization and evaluation of the in vitro anticancer activity of phlorotannin-rich fraction from the brown seaweed, Cystoseira sedoides. J. Aquat. Food Prod. Technol. 2019, 28, 892–909. [Google Scholar] [CrossRef]
- Abu-Khudir, R.; Ismail, G.A.; Diab, T. Antimicrobial, antioxidant, and anti-tumor activities of Sargassum linearifolium and Cystoseira crinita from Egyptian Mediterranean Coast. Nutr. Cancer 2020, 0, 1–16. [Google Scholar] [CrossRef]
- Premarathna, A.D.; Ranahewa, T.H.; Wijesekera, S.K.; Harishchandra, D.L.; Karunathilake, K.J.K.; Waduge, R.N.; Wijesundara, R.R.M.K.K.; Jayasooriya, A.P.; Wijewardana, V.; Rajapakse, R.P.V.J. Preliminary screening of the aqueous extracts of twenty-three different seaweed species in Sri Lanka with in-vitro and in-vivo assays. Heliyon 2020, 6, e03918. [Google Scholar] [CrossRef]
- Ahn, J.H.; Yang, Y.I.; Lee, K.T.; Choi, J.H. Dieckol, isolated from the edible brown algae Ecklonia cava, induces apoptosis of ovarian cancer cells and inhibits tumor xenograft growth. J. Cancer Res. Clin. Oncol. 2015, 141, 255–268. [Google Scholar] [CrossRef]
- Yang, Y.I.; Ahn, J.H.; Choi, Y.S.; Choi, J.H. Brown algae phlorotannins enhance the tumoricidal effect of cisplatin and ameliorate cisplatin nephrotoxicity. Gynecol. Oncol. 2015, 136, 355–364. [Google Scholar] [CrossRef]
- Eo, H.J.; Kwon, T.H.; Park, G.H.; Song, H.M.; Lee, S.J.; Park, N.H.; Jeong, J.B. In vitro anticancer activity of phlorofucofuroeckol a via upregulation of activating transcription factor 3 against human colorectal cancer cells. Mar. Drugs 2016, 14, 69. [Google Scholar] [CrossRef]
- Park, C.; Lee, H.; Hwangbo, H.; Ji, S.Y.; Kim, M.Y.; Kim, S.Y.; Hong, S.H.; Kim, G.Y.; Choi, Y.H. Ethanol extract of Hizikia fusiforme induces apoptosis in B16F10 mouse melanoma cells through ROS-dependent inhibition of the PI3K/Akt signaling pathway. Asian Pacific J. Cancer Prev. 2020, 21, 1275–1282. [Google Scholar] [CrossRef]
- Lee, H.; Kang, C.; Jung, E.S.; Kim, J.S.; Kim, E. Antimetastatic activity of polyphenol-rich extract of Ecklonia cava through the inhibition of the Akt pathway in A549 human lung cancer cells. Food Chem. 2011, 127, 1229–1236. [Google Scholar] [CrossRef] [PubMed]
- Kim, R.K.; Uddin, N.; Hyun, J.W.; Kim, C.; Suh, Y.; Lee, S.J. Novel anticancer activity of phloroglucinol against breast cancer stem-like cells. Toxicol. Appl. Pharmacol. 2015, 286, 143–150. [Google Scholar] [CrossRef]
- Kim, R.K.; Suh, Y.; Yoo, K.C.; Cui, Y.H.; Hwang, E.; Kim, H.J.; Kang, J.S.; Kim, M.J.; Lee, Y.Y.; Lee, S.J. Phloroglucinol suppresses metastatic ability of breast cancer cells by inhibition of epithelial-mesenchymal cell transition. Cancer Sci. 2015, 106, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Liu, G.; Qiu, L.; Lin, X.; Liu, M. Marine bromophenol bis(2,3-dibromo-4,5-dihydroxybenzyl) ether, represses angiogenesis in HUVEC cells and in zebrafish embryos via inhibiting the VEGF signal systems. Biomed. Pharmacother. 2015, 75, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Sadeeshkumar, V.; Duraikannu, A.; Ravichandran, S.; Kodisundaram, P.; Fredrick, W.S.; Gobalakrishnan, R. Modulatory efficacy of dieckol on xenobiotic-metabolizing enzymes, cell proliferation, apoptosis, invasion and angiogenesis during NDEA-induced rat hepatocarcinogenesis. Mol. Cell. Biochem. 2017, 433, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.X.; Li, Y.; Je, J.Y.; Kim, S.K. Dieckol as a novel anti-proliferative and anti-angiogenic agent and computational anti-angiogenic activity evaluation. Environ. Toxicol. Pharmacol. 2015, 39, 259–270. [Google Scholar] [CrossRef]
- Zhen, A.X.; Hyun, Y.J.; Piao, M.J.; Devage, P.; Madushan, S. Eckol inhibits particulate matter 2.5-induced skin damage via MAPK signaling pathway. Mar. Drugs 2019, 17, 444. [Google Scholar] [CrossRef]
- Zhang, M.Y.; Guo, J.; Hu, X.M.; Zhao, S.Q.; Li, S.L.; Wang, J. An in vivo anti-tumor effect of eckol from marine brown algae by improving the immune response. Food Funct. 2019, 10, 4361–4371. [Google Scholar] [CrossRef] [PubMed]
- Ford, J.M.; Kastan, M.B. DNA Damage response pathways and fcancer. In Abeloff’s Clinical Oncology: Fifth Edition; Saunders: Philadelphia, PA, USA, 2013; pp. 142–153.e4. ISBN 9780323222112. [Google Scholar]
- Singh, A.K.; Cabral, C.; Kumar, R.; Ganguly, R.; Rana, H.K.; Gupta, A.; Lauro, M.R.; Carbone, C.; Reis, F.; Pandey, A.K. Beneficial effects of dietary polyphenols on gut microbiota and strategies to improve delivery efficiency. Nutrients 2019, 11, 2216. [Google Scholar] [CrossRef]
- Guo, B.; Liu, B.; Wei, H.; Cheng, K.W.; Chen, F. Extract of the microalga Nitzschia laevis prevents high-fat-diet-induced obesity in mice by modulating the composition of gut microbiota. Mol. Nutr. Food Res. 2019, 63, 1–14. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [PubMed]
- Rajauria, G.; Jaiswal, A.K.; Abu-Gannam, N.; Gupta, S. Antimicrobial, antioxidant and free radical-scavenging capacity of brown seaweed Himanthalia elongata from western coast of Ireland. J. Food Biochem. 2013, 37, 322–335. [Google Scholar] [CrossRef]
- Besednova, N.N.; Zvyagintseva, T.N.; Kuznetsova, T.A.; Makarenkova, I.D.; Smolina, T.P.; Fedyanina, L.N.; Kryzhanovsky, S.P.; Zaporozhets, T.S. Marine algae metabolites as promising therapeutics for the prevention and treatment of HIV/AIDS. Metabolites 2019, 9, 87. [Google Scholar] [CrossRef] [PubMed]
- Ahn, M.-J.; Yoon, K.-D.; Kim, C.Y.; Kim, J.H.; Shin, C.-G.; Kim, J.M. Inhibitory activity on HIV-1 reverse transcriptase and integrase of a carmalol derivative from a brown alga, Ishige okamurae. Phyther. Res. 2006, 20, 711–713. [Google Scholar] [CrossRef] [PubMed]
- Artan, M.; Li, Y.; Karadeniz, F.; Lee, S.H.; Kim, M.M.; Kim, S.K. Anti-HIV-1 activity of phloroglucinol derivative, 6,6′-bieckol, from Ecklonia cava. Bioorganic Med. Chem. 2008, 16, 7921–7926. [Google Scholar] [CrossRef]
- Karadeniz, F.; Kang, K.H.; Park, J.W.; Park, S.J.; Kim, S.K. Anti-HIV-1 activity of phlorotannin derivative 8,4’’’-dieckol from Korean brown alga Ecklonia cava. Biosci. Biotechnol. Biochem. 2014, 78, 1151–1158. [Google Scholar] [CrossRef]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mateos, R.; Pérez-Correa, J.R.; Domínguez, H. Bioactive Properties of Marine Phenolics. Mar. Drugs 2020, 18, 501. https://doi.org/10.3390/md18100501
Mateos R, Pérez-Correa JR, Domínguez H. Bioactive Properties of Marine Phenolics. Marine Drugs. 2020; 18(10):501. https://doi.org/10.3390/md18100501
Chicago/Turabian StyleMateos, Raquel, José Ricardo Pérez-Correa, and Herminia Domínguez. 2020. "Bioactive Properties of Marine Phenolics" Marine Drugs 18, no. 10: 501. https://doi.org/10.3390/md18100501
APA StyleMateos, R., Pérez-Correa, J. R., & Domínguez, H. (2020). Bioactive Properties of Marine Phenolics. Marine Drugs, 18(10), 501. https://doi.org/10.3390/md18100501