The Protective Effect of Mycosporine-Like Amino Acids (MAAs) from Porphyra yezoensis in a Mouse Model of UV Irradiation-Induced Photoaging

Abstract
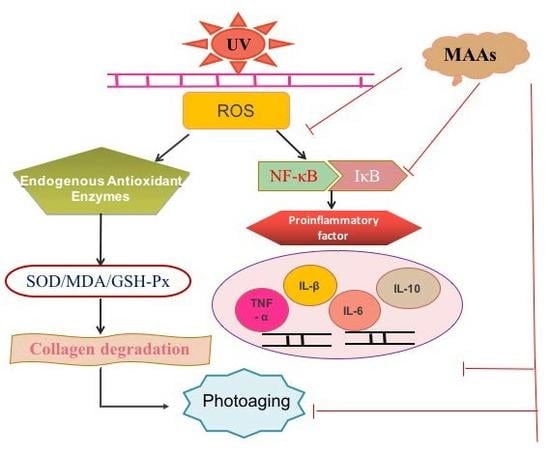
:
1. Introduction
2. Results
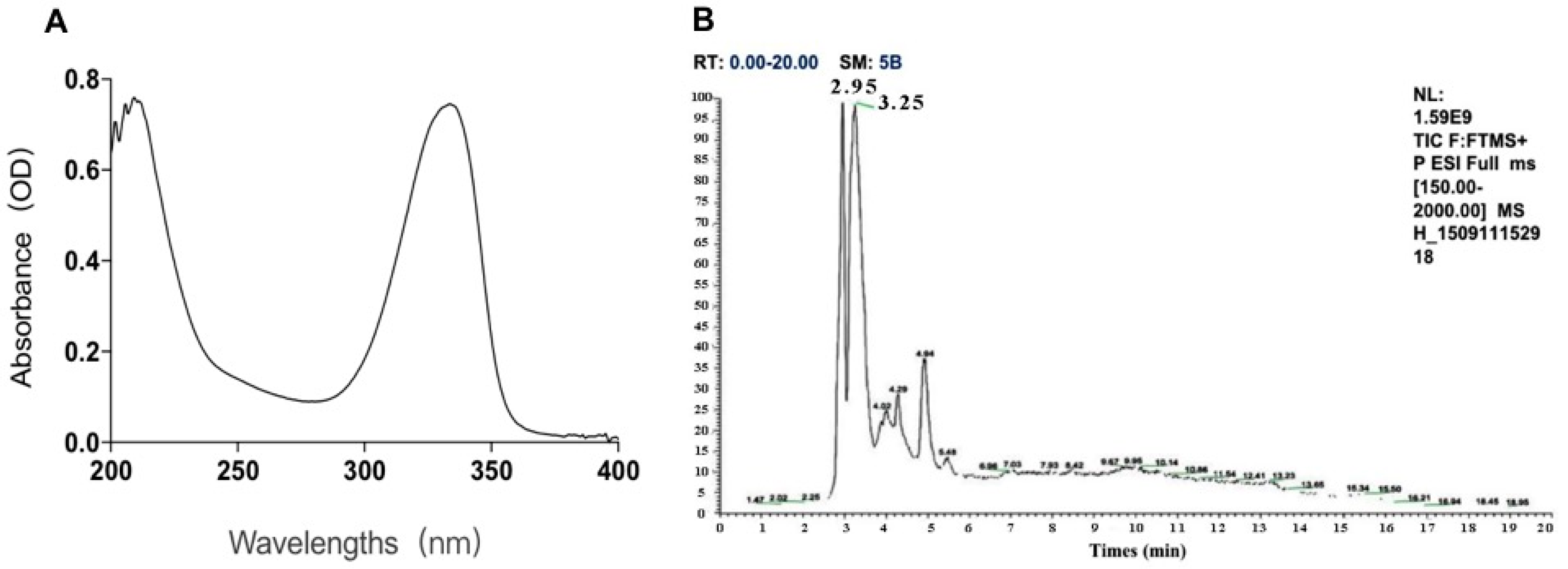
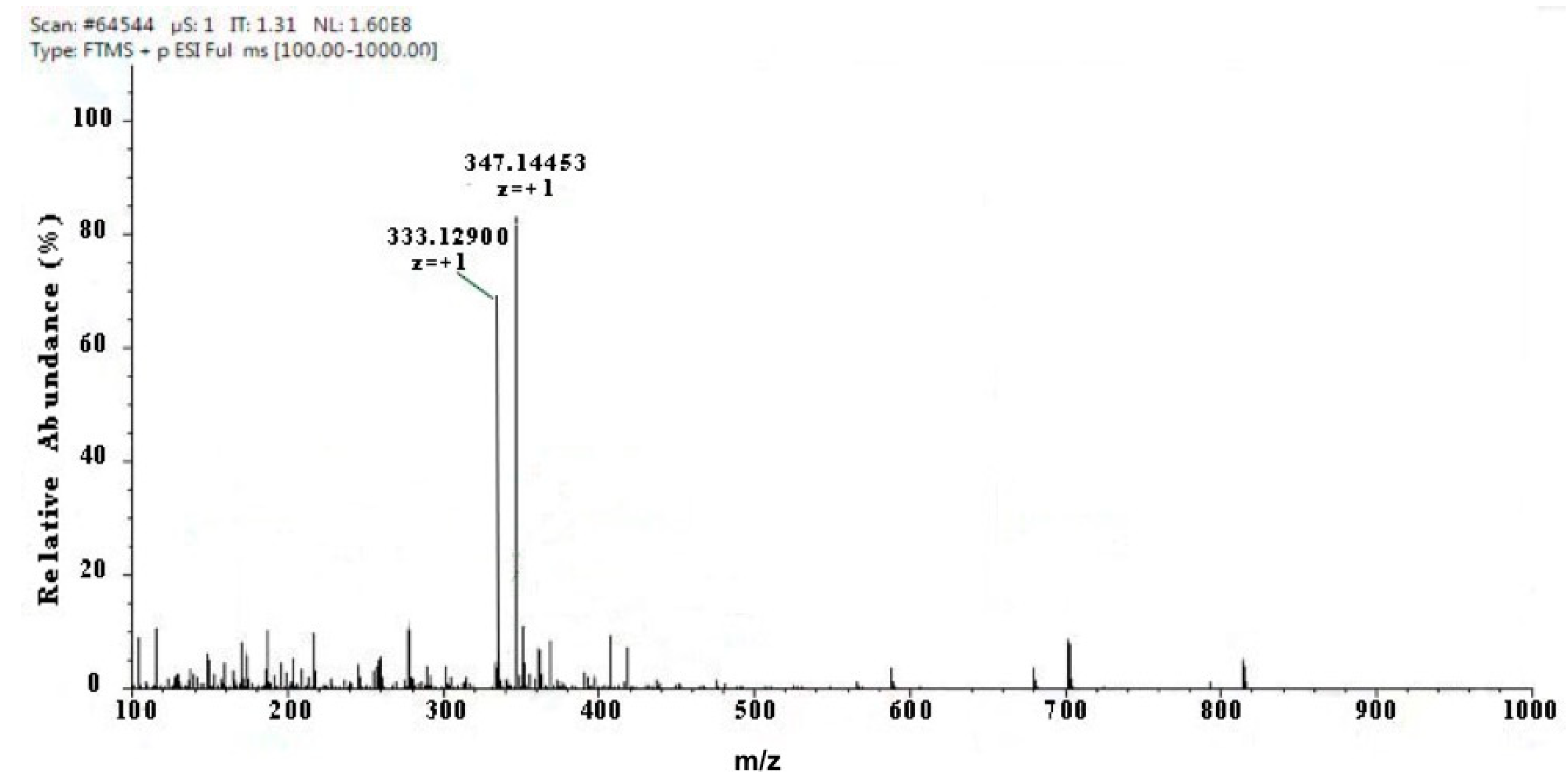
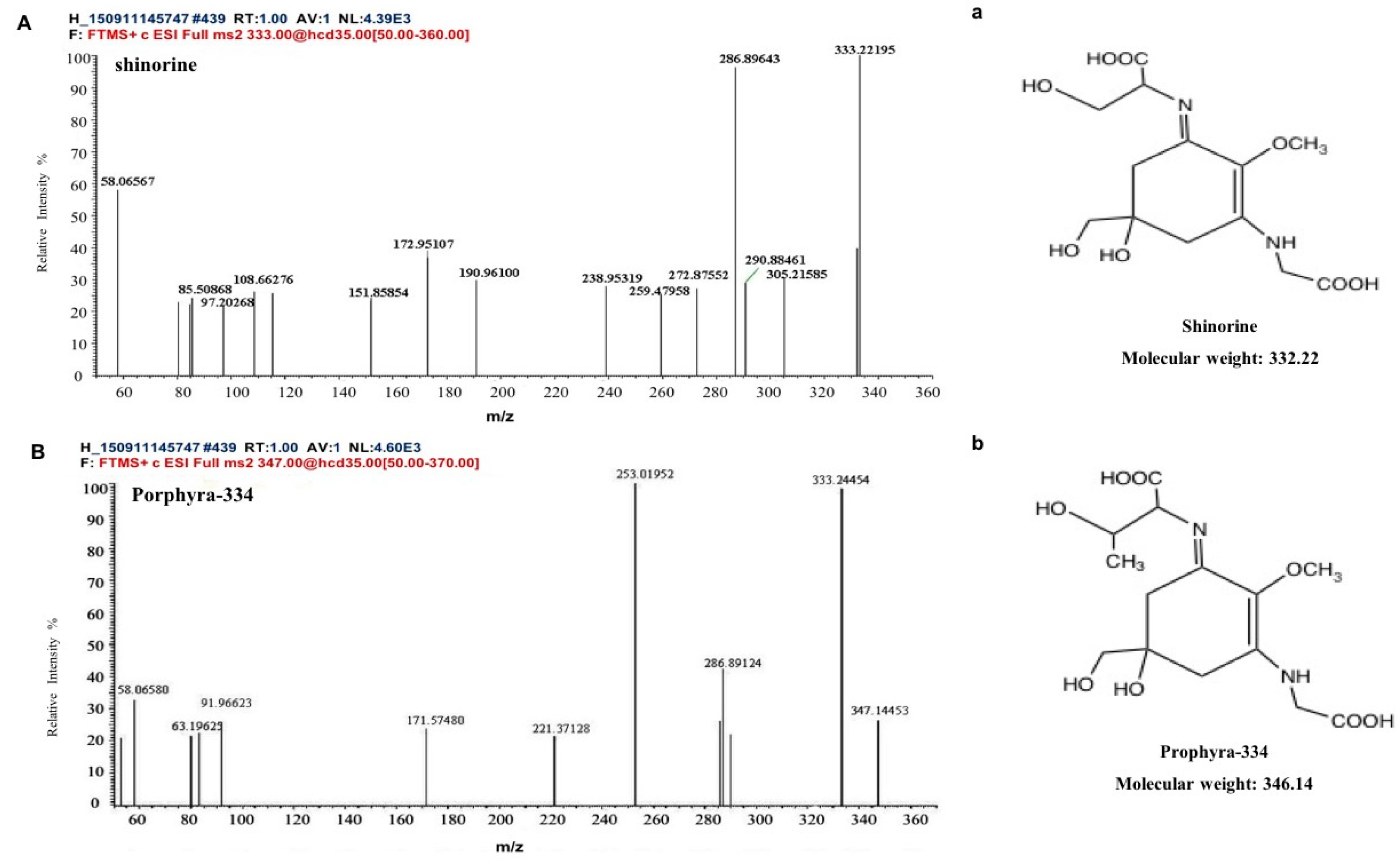
2.1. The Components of MAAs Extracted from Porphyra yezoensis
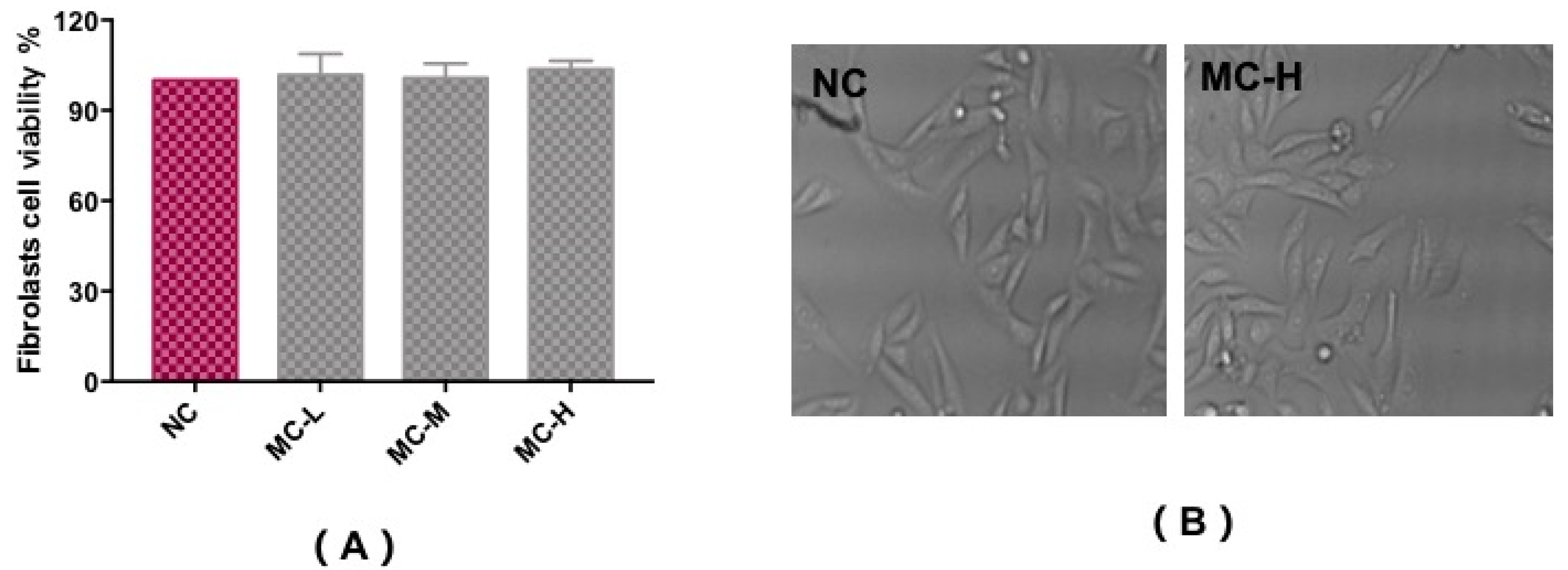
2.2. Effect of MAAs on Fibroblast Cytotoxicity
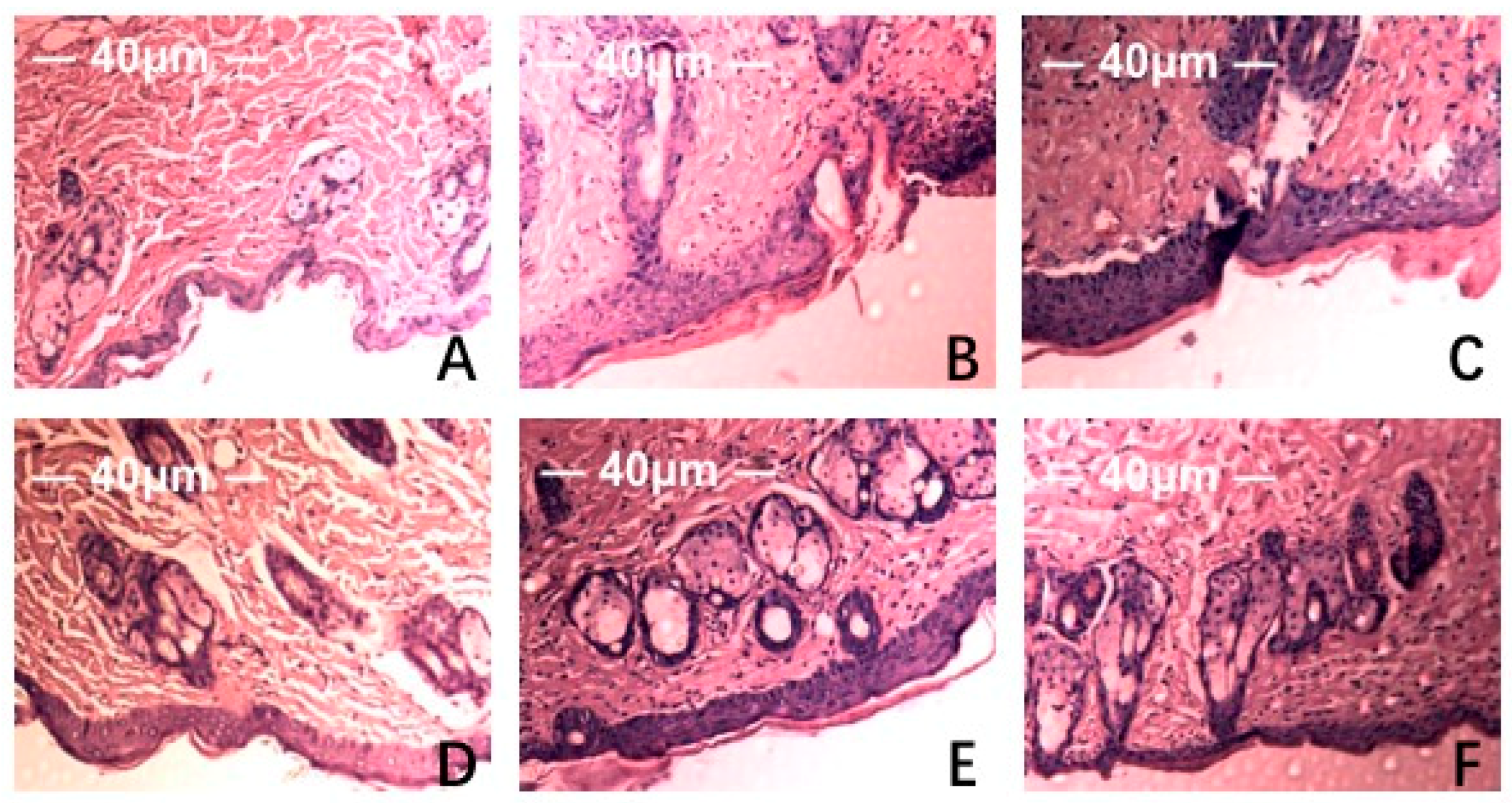
2.3. Pathological Histology Analysis
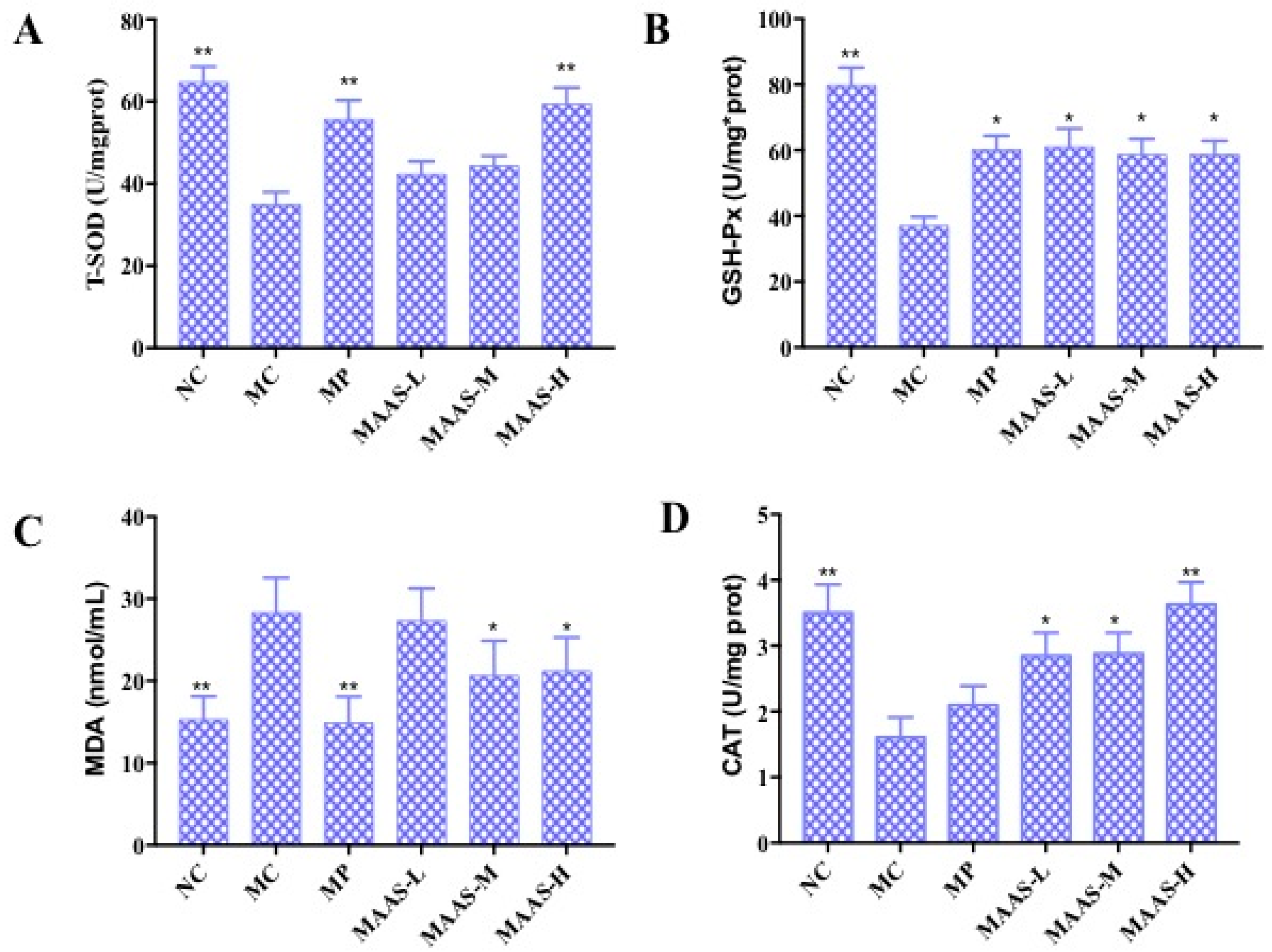
2.4. Effect of MAAs on UV Irradiation-Induced Depletion of Endogenous Antioxidant Enzymes
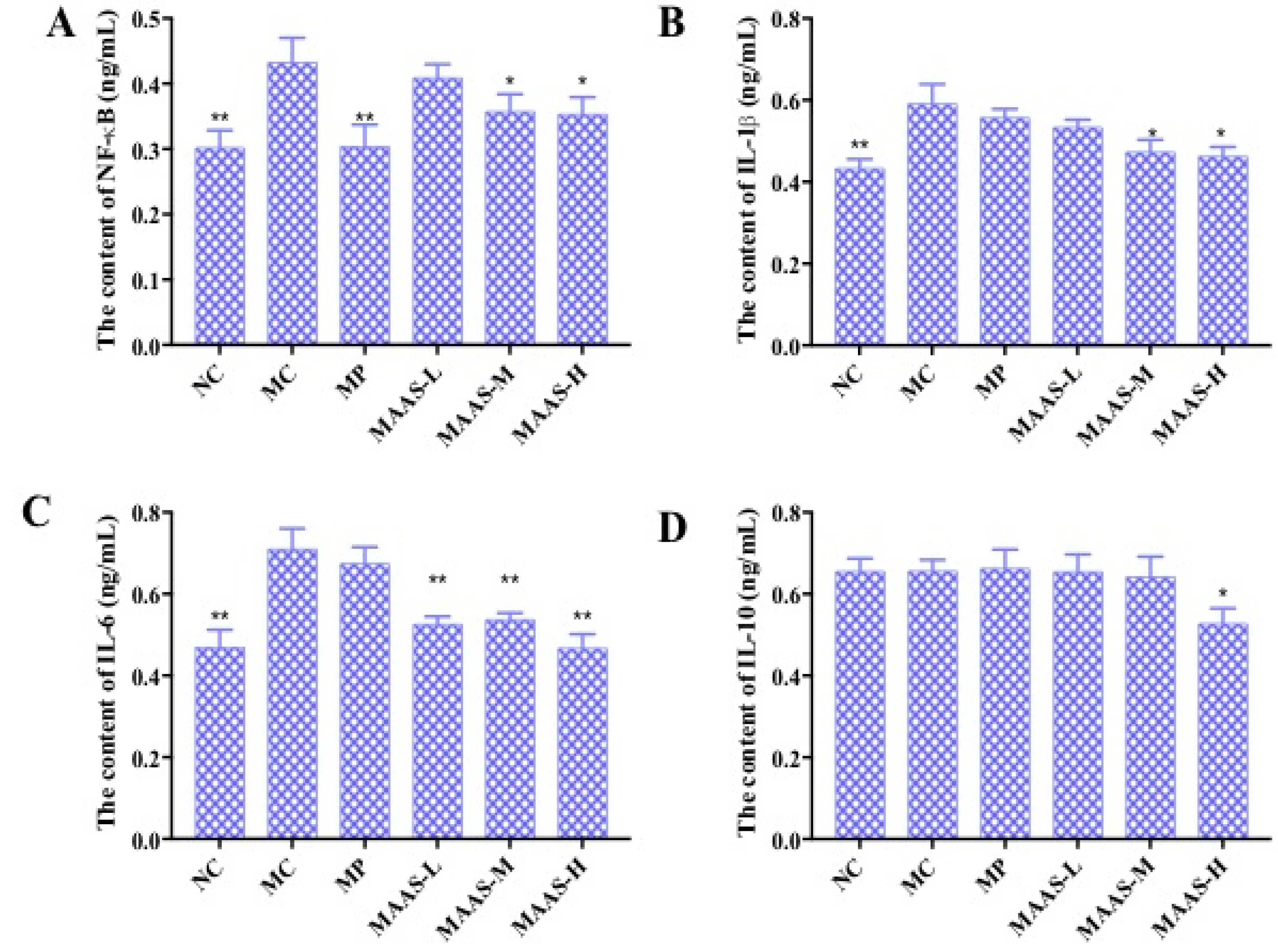
2.5. Effect of MAAs on Inflammatory Cytokine Levels in Photoaging Skin
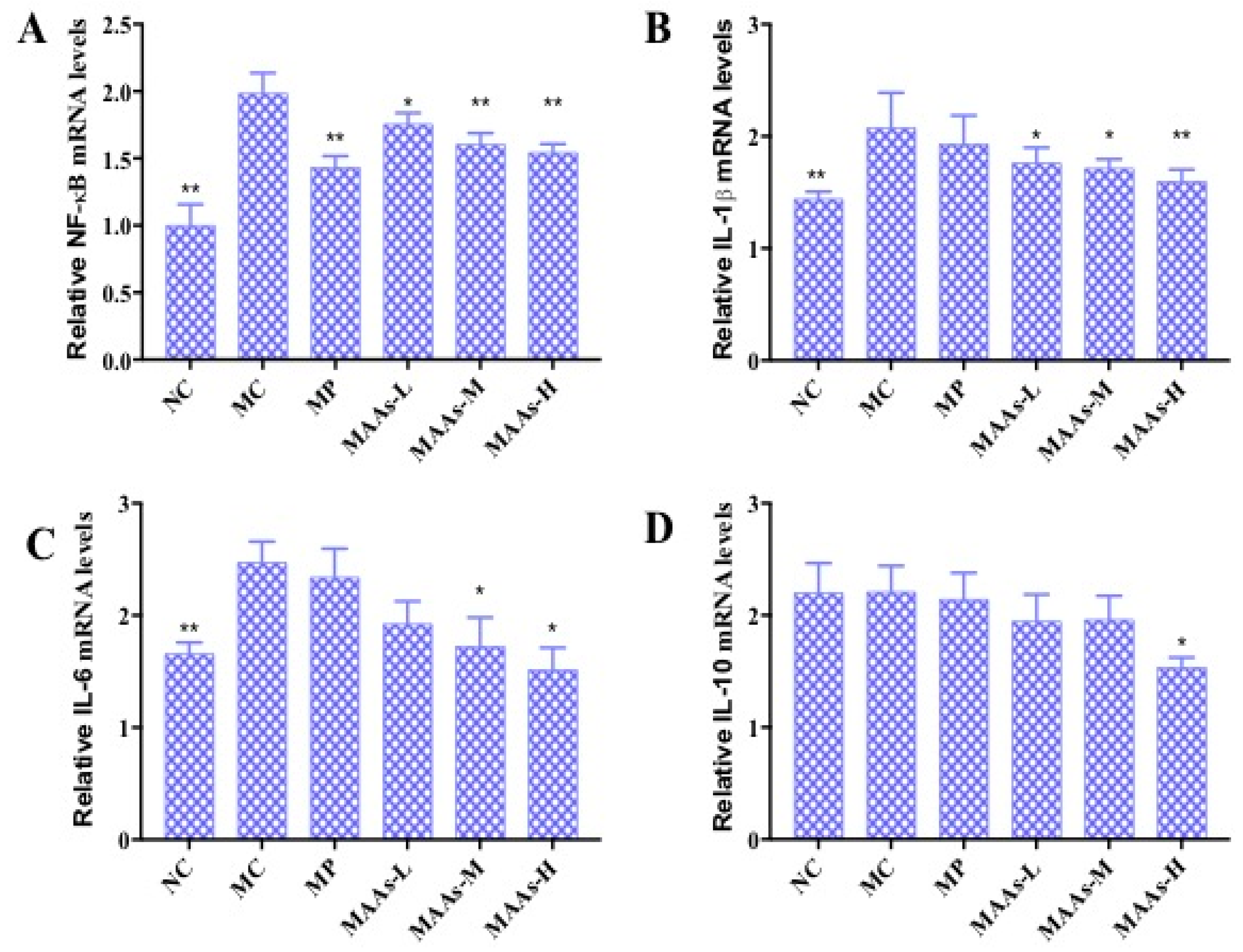
2.6. Effect of MAAs on the mRNA Expression of Inflammatory Cytokines in Mouse Skin
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Preparation and Analysis of Mycosporine-Like Amino Acids
4.3. MTT Cytotoxicity Assay
4.4. Experimental Groups
4.5. Establishment of the UV Irradiation-Induced Skin Photoaging Model
4.6. Tissue Morphology and Histopathological Analysis
4.7. Tissue Histopathological Analysis by Scanning Electron Microscope (SEM)
4.8. The Effect of MAAs on Endogenous Antioxidant Enzymes
4.9. Effect of MAAs on the Expression of Inflammatory Cytokines
4.10. RNA Isolation and Quantitative Real-Time PCR
4.11. Statistical ANALYSIS
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References and Note
- Bravo, K.; Duque, L.; Ferreres, F.; Moreno, D.A.; Osorio, E. Passiflora tarminiana, fruits reduce uvb-induced photoaging in human skin fibroblasts. J. Photochem. Photobiol. B Biol. 2017, 168, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Scharffetter–Kochanek, K.; Brenneisen, P.; Wenk, J.; Herrmann, G.; Ma, W.; Kuhr, L.; Meewes, C.; Wlaschek, M. Photoaging of the skin from phenotype to mechanisms. Exp. Gerontol. 2000, 35, 307–316. [Google Scholar] [CrossRef]
- Grether-Beck, S.; Marini, A.; Jaenicke, T.; Krutmann, J. Photoprotection of human skin beyond ultraviolet radiation. Photodermatol. Photoimmunol. Photomed. 2014, 30, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.J.; Kim, K.B.; Heo, J.; Cho, D.H.; Kim, H.S.; Han, S.H.; Ahn, K.J.; An, I.S.; An, S.; Bae, S. Protective effect of arthrospira platensis extracts against ultraviolet b-induced cellular senescence through inhibition of dna damage and matrix metalloproteinase-1 expression in human dermal fibroblasts. J. Photochem. Photobiol. B Biol. 2017, 173, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Bossi, O.; Gartsbein, M.; Leitges, M.; Kuroki, T.; Tennenbaum, T. Uv irradiation increases ros production via pkcδ signaling in primary murine fibroblasts. J. Cell. Biochem. 2008, 105, 194–207. [Google Scholar] [CrossRef] [PubMed]
- Salminen, A.; Huuskonen, J.; Ojala, J.; Kauppinen, A.; Kaarniranta, K.; Suuronen, T. Activation of innate immunity system during aging: Nf-kb signaling is the molecular culprit of inflamm-aging. Ageing Res. Rev. 2008, 7, 83–105. [Google Scholar] [CrossRef] [PubMed]
- Eda, H.; Shimada, H.; Beidler, D.R.; Monahan, J.B. Proinflammatory cytokines, il-1β and tnf-α, induce expression of interleukin-34 mrna via jnk- and p44/42 mapk-nf-κb pathway but not p38 pathway in osteoblasts. Rheumatol. Int. 2011, 31, 1525–1530. [Google Scholar] [CrossRef] [PubMed]
- Jobin, C.; Bradham, C.A.; Russo, M.P.; Juma, B.; Narula, A.S.; Brenner, D.A.; Sartor, R.B. Curcumin blocks cytokine-mediated nf-kappa b activation and proinflammatory gene expression by inhibiting inhibitory factor i-kappa b kinase activity. J. Immunol. 1999, 163, 3474–3483. [Google Scholar]
- Chen, T.; Hou, H. Protective effect of gelatin polypeptides from pacific cod (gadus macrocephalus) against uv irradiation-induced damages by inhibiting inflammation and improving transforming growth factor-β/smad signaling pathway. J. Photochem. Photobiol. B Biol. 2016, 162, 633–640. [Google Scholar] [CrossRef]
- Han, A.R.; Nam, M.H.; Lee, K.W. Plantamajoside inhibits uvb and advanced glycation end products-induced mmp-1 expression by suppressing the mapk and nf-\r, κ\r, b pathways in hacat cells. Photochem. Photobiol. 2016, 92, 708–719. [Google Scholar] [CrossRef]
- Zhuang, Y.; Hou, H.; Zhao, X.; Zhang, Z.; Li, B. Effects of collagen and collagen hydrolysate from jellyfish (rhopilema esculentum) on mice skin photoaging induced by uv irradiation. J. Food Sci. 2010, 74, H183–H188. [Google Scholar] [CrossRef] [PubMed]
- Rui, Y.; Zhaohui, Z.; Wenshan, S.; Bafang, L.; Hu, H. Protective effect of MAAs extracted from Porphyra tenera against UV irradiation-induced photoaging in mouse skin. J. Photochem. Photobiol. B Biol. 2019, 192, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.P.; Kumari, S.; Rastogi, R.P.; Singh, K.L.; Sinha, R.P. Mycosporine-like amino acids (maas): Chemical structure, biosynthesis and significance as uv-absorbing/screening compounds. Indian J. Exp. Biol. 2008, 46, 7. [Google Scholar] [PubMed]
- Chen, T.; Hou, H.; Fan, Y.; Wang, S.; Chen, Q.; Si, L.; Li, B. Protective effect of gelatin peptides from pacific cod skin against photoaging by inhibiting the expression of mmps via mapk signaling pathway. J. Photochem. Photobiol. B Biol. 2016, 165, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Suh, S.S.; Hwang, J.; Park, M.; Seo, H.; Kim, H.S.; Lee, J.; Moh, S.; Lee, T.K. Anti-inflammation activities of mycosporine-like amino acids (maas) in response to uv radiation suggest potential anti-skin aging activity. Mar. Drugs 2014, 12, 5174–5187. [Google Scholar] [CrossRef]
- Farris, P.K. Topical vitamin c: A useful agent for treating photoaging and other dermatologic conditions. Dermatol. Surg. 2010, 31, 814–818. [Google Scholar] [CrossRef]
- Ewelina, C.; Ariel, K.; Kornelia, D.; Beata, B.; Michal, A.; Jan, B. Mycosporine-like amino acids: Potential health and beauty ingredients. Mar. Drugs 2017, 15, 326. [Google Scholar]
- Sample, A.; He, Y.Y. Mechanisms and prevention of uv-induced melanoma. Photodermatol. Photoimmunol. Photomed. 2017, 34, 13–24. [Google Scholar] [CrossRef]
- Hou, H.; Li, B.; Zhao, X.; Zhuang, Y.; Ren, G.; Yan, M.; Cai, Y.; Zhang, X.; Chen, L. The effect of pacific cod (gadus macrocephalus) skin gelatin polypeptides on uv radiation-induced skin photoaging in icr mice. Food Chem. 2009, 115, 945–950. [Google Scholar] [CrossRef]
- Fisher, G.J.; Kang, S.; Varani, J.; Bata-Csorgo, Z.; Wan, Y.; Datta, S.; Voorhees, J.J. Mechanisms of photoaging and chronological skin aging. Arch. Dermatol. 2002, 138, 1462–1470. [Google Scholar] [CrossRef]
- Cavinato, M.; Jansen-Dürr, P. Molecular mechanisms of uvb-induced senescence of dermal fibroblasts and its relevance for photoaging of the human skin. Exp. Gerontol. 2017, 94, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Ha, S.J.; Lee, J.; Kim, H.; Song, K.M.; Lee, N.H.; Kim, Y.E.; Lee, H.; Kim, Y.H.; Jung, S.K. Preventive effect of rhus javanica, extract on uvb-induced skin inflammation and photoaging. J. Funct. Foods 2016, 27, 589–599. [Google Scholar] [CrossRef]
- Feng, X.X.; Yu, X.T.; Li, W.J.; Kong, S.Z.; Liu, Y.H.; Zhang, X.; Xian, Y.F.; Zhang, X.J.; Su, Z.R.; Lin, Z.X. Effects of topical application of patchouli alcohol on the uv-induced skin photoaging in mice. Eur. J. Pharm. Sci. 2014, 63, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.J.; Lee, C.J.; Lin, T.B.; Liu, H.J.; Huang, S.Y.; Chen, J.Z.; Tseng, K.W. Inhibition of ultraviolet b-induced expression of the proinflammatory cytokines tnf-α and vegf in the cornea by fucoxanthin treatment in a rat model. Mar. Drugs 2016, 14, 13. [Google Scholar] [CrossRef] [PubMed]
- Vogel, H.G.; Vogel, W.H.; Bernward, A.S.; et al. Guidelines for the care and use of laboratory animals [J].
- Singh, S.P.; Häder, D.P.; Sinha, R.P. Cyanobacteria and ultraviolet radiation (uvr) stress: Mitigation strategies. Ageing Res. Rev. 2010, 9, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative pcr and the 2 (-delta delta c (t)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequences |
|---|---|
| GADPH | F:5’-CGTGTTCCTACCCCCAATGA-3’ |
| R:5’-ATGTCATCATACTTGGCAGGTTTCT-3’ | |
| NF-κB | F:5’-CACTGAGGAGACCACCCAAG-3’ |
| R:5’-GTAAACGCCGAAGATGATGG-3’ | |
| IL-β | F:5’-CTCCATGAGCTTTGTACAAGG-3’ |
| R:5’-TGCTGATGTACCAGTTGGGG-3’ | |
| IL-6 | F:5’-GTGGCTAAGGACCAAGACCA-3’ |
| R:5’-TTCCAAGAAACCATCTGGCTA-3’ | |
| IL-10 | F:5’-TCCTTGGAAAACCTCGTTTG-3’ |
| R:5’-CTTCAATTGCTTCCCAAGGA-3’ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ying, R.; Zhang, Z.; Zhu, H.; Li, B.; Hou, H. The Protective Effect of Mycosporine-Like Amino Acids (MAAs) from Porphyra yezoensis in a Mouse Model of UV Irradiation-Induced Photoaging. Mar. Drugs 2019, 17, 470. https://doi.org/10.3390/md17080470
Ying R, Zhang Z, Zhu H, Li B, Hou H. The Protective Effect of Mycosporine-Like Amino Acids (MAAs) from Porphyra yezoensis in a Mouse Model of UV Irradiation-Induced Photoaging. Marine Drugs. 2019; 17(8):470. https://doi.org/10.3390/md17080470
Chicago/Turabian StyleYing, Rui, Zhaohui Zhang, Huiying Zhu, Bafang Li, and Hu Hou. 2019. "The Protective Effect of Mycosporine-Like Amino Acids (MAAs) from Porphyra yezoensis in a Mouse Model of UV Irradiation-Induced Photoaging" Marine Drugs 17, no. 8: 470. https://doi.org/10.3390/md17080470
APA StyleYing, R., Zhang, Z., Zhu, H., Li, B., & Hou, H. (2019). The Protective Effect of Mycosporine-Like Amino Acids (MAAs) from Porphyra yezoensis in a Mouse Model of UV Irradiation-Induced Photoaging. Marine Drugs, 17(8), 470. https://doi.org/10.3390/md17080470