Transcriptomic and Proteomic Analysis of the Tentacles and Mucus of Anthopleura dowii Verrill, 1869

 , ,
, ,  , , , ,
, , , ,
Abstract
1. Introduction
2. Results and Discussion
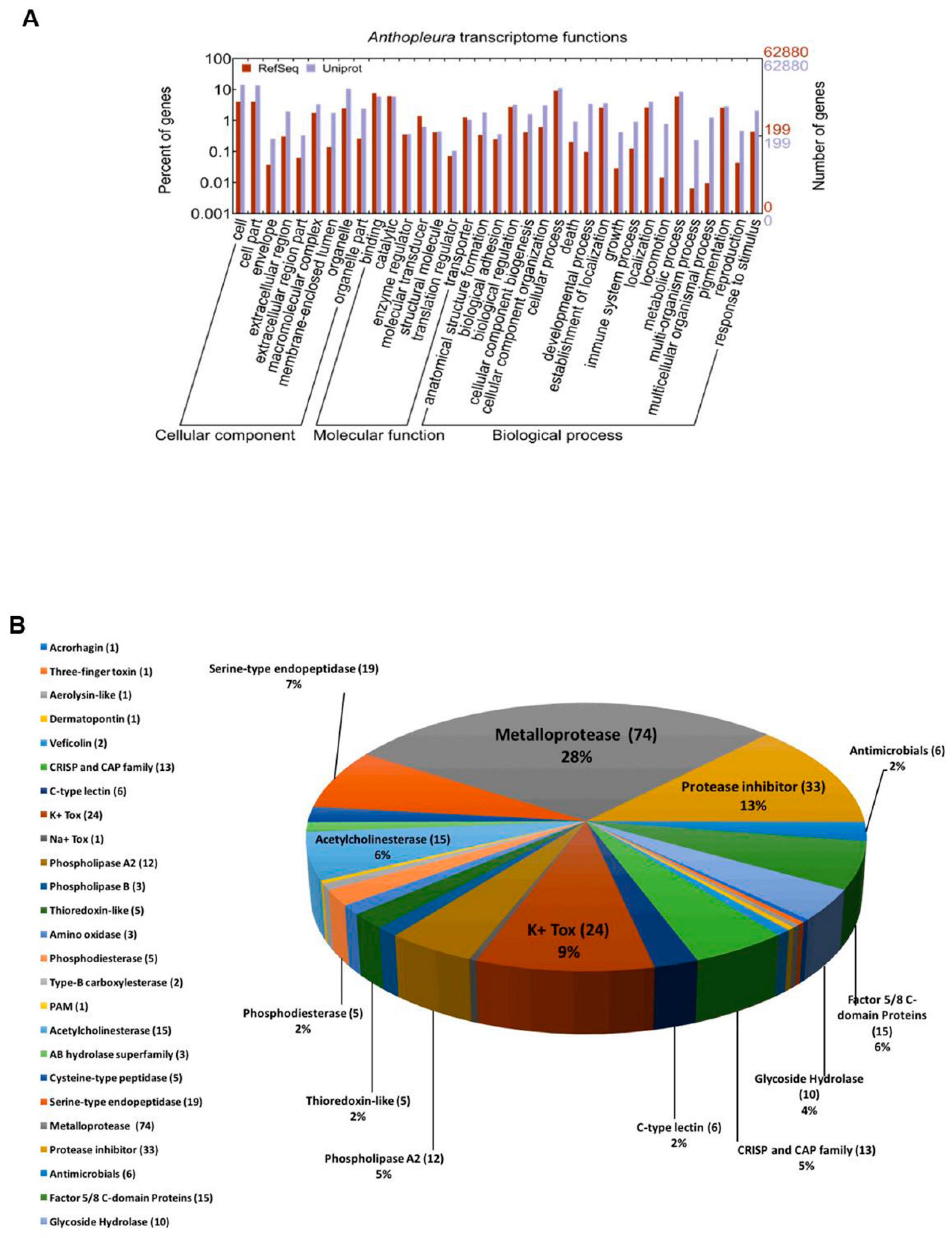
2.1. Venom Components Identified in the Transcriptome of the Tentacles of A. dowii
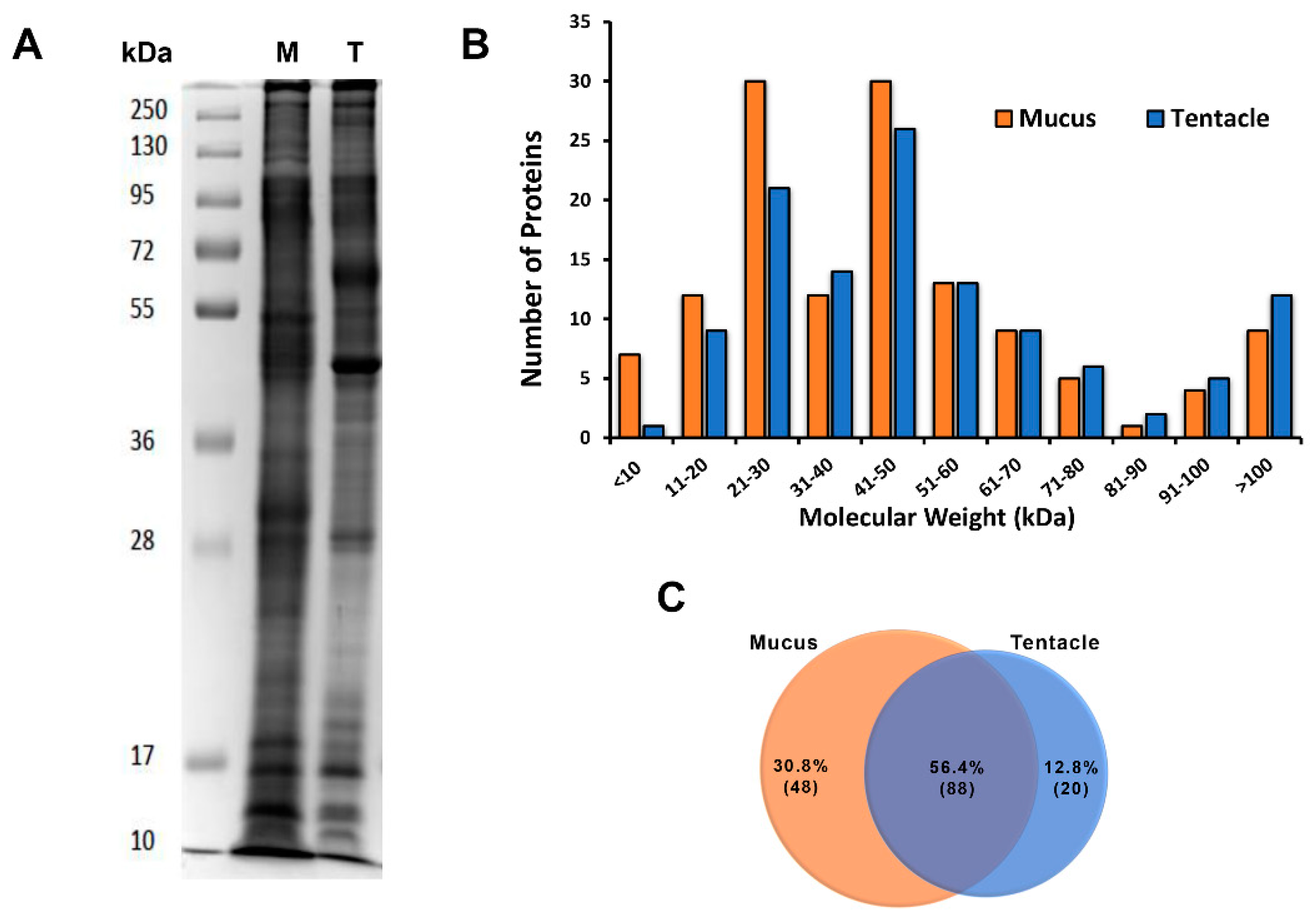
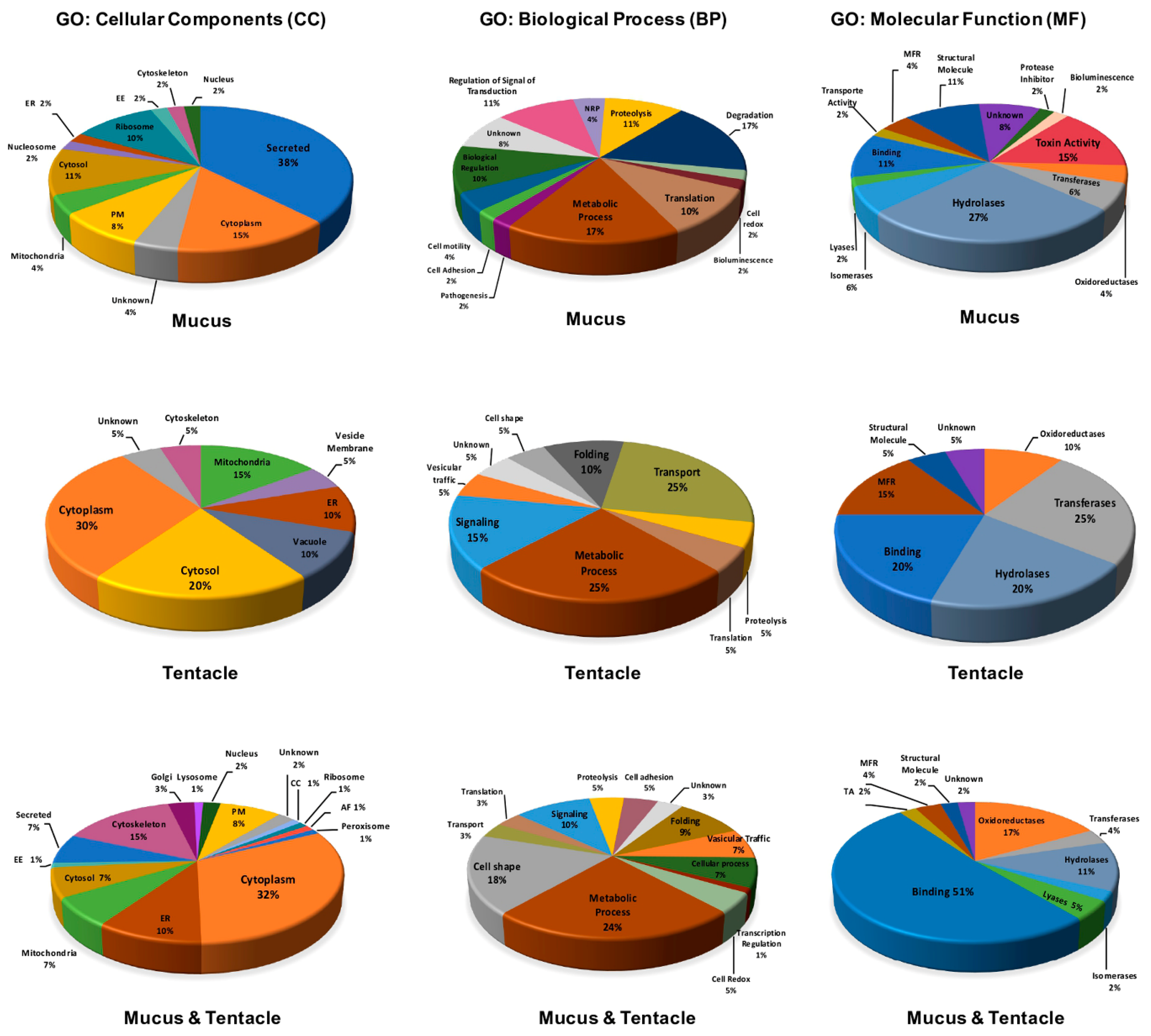
2.2. Venom Components Identified in the Proteome of the Tentacle and Mucus of A. dowii
2.3. Polypeptides Related to the Venom and its Identification in the Proteome and Transcriptome of A. dowii
2.3.1. Sodium Voltage-Gated (NaV) Channel Toxins
2.3.2. Type 2 Potassium Voltage-Gated (Kv) Channel Toxins Subfamily
2.4. Protease Inhibitors
2.5. Proteases
2.6. Glycosyl-Hydrolases
2.7. Putative Venom Components Identified Exclusively in the Transcriptome of A. dowii.
2.7.1. Potassium Channel Toxins (KTx)
Type 1 KTx Family
Type 3 KTx Family
Type 5 Ktx Families
2.7.2. Phospholipases A2 (PLA2)
2.7.3. Other Proteins Identified in the Transcriptome
3. Materials and Methods
3.1. Venom and Tentacle Sample Preparation for Proteomic Analysis
3.2. In solution Protein Digestion and LC-MS/MS Analysis
3.3. Peptide Identification and Functional Annotation of Proteomics Data
3.4. Functional Annotation of Assembled Transcriptome and Identification of Genes of Putative Toxins
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Frazão, B.; Vasconcelos, V.; Antunes, A. Sea Anemone (Cnidaria, Anthozoa, Actiniaria) Toxins: An Overview. Mar. Drugs 2012, 10, 1812–1851. [Google Scholar] [CrossRef] [PubMed]
- Arbuckle, K. Evolutionary Context of Venom in Animals. In Evolution of Venomous Animals and Their Toxins; Springer: Dordrecht, The Netherlands, 2015; Volume 24. [Google Scholar]
- Madio, B.; Peigneur, S.; Chin, Y.K.Y.; Hamilton, B.R.; Henriques, S.T.; Smith, J.J.; Cristofori-Armstrong, B.; Dekan, Z.; Boughton, B.A.; Alewood, P.F.; et al. PHAB toxins: A unique family of predatory sea anemone toxins evolving via intra-gene concerted evolution defines a new peptide fold. Cell. Mol. Life Sci. 2018, 75, 4511–4524. [Google Scholar] [CrossRef] [PubMed]
- Prentis, P.J.; Pavasovic, A.; Norton, R.S. Sea Anemones: Quiet Achievers in the Field of Peptide Toxins. Toxins 2018, 10, 36. [Google Scholar] [CrossRef] [PubMed]
- Fautin, D.G. Structural diversity, systematics, and evolution of cnidae. Toxicon 2009, 54, 1054–1064. [Google Scholar] [CrossRef]
- Jouiaei, M.; Yanagihara, A.; Madio, B.; Nevalainen, T.; Alewood, P.; Fry, B. Ancient Venom Systems: A Review on Cnidaria Toxins. Toxins 2015, 7, 2251–2271. [Google Scholar] [CrossRef]
- Mariottini, G.; Pane, L. Cytotoxic and Cytolytic Cnidarian Venoms. A Review on Health Implications and Possible Therapeutic Applications. Toxins 2013, 6, 108–151. [Google Scholar] [CrossRef]
- King, G.F. Venoms as a platform for human drugs: Translating toxins into therapeutics. Expert Opin. Biol. Ther. 2011, 11, 1469–1484. [Google Scholar] [CrossRef]
- Liao, Q.; Feng, Y.; Yang, B.; Lee, S.M.-Y. Cnidarian peptide neurotoxins: A new source of various ion channel modulators or blockers against central nervous systems disease. Drug Discov. Today 2018, 24, 189–197. [Google Scholar] [CrossRef]
- Tudor, J.E.; Pallaghy, P.K.; Pennington, M.W.; Norton, R.S. Solution structure of ShK toxin, a novel potassium channel inhibitor from a sea anemone. Nat. Struct. Biol. 1996, 3, 317–320. [Google Scholar] [CrossRef]
- Chandy, K.G.; Wulff, H.; Beeton, C.; Pennington, M.; Gutman, G.A.; Cahalan, M.D. K+ channels as targets for specific immunomodulation. Trends Pharmacol. Sci. 2004, 25, 280–289. [Google Scholar] [CrossRef]
- Chandy, K.G.; Norton, R.S. Peptide blockers of Kv1.3 channels in T cells as therapeutics for autoimmune disease. Curr. Opin. Chem. Biol. 2017, 38, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Liu, X.-S.; Diochot, S.; Lazdunski, M.; Tseng, G.-N. APETx1 from Sea Anemone Anthopleura elegantissima Is a Gating Modifier Peptide Toxin of the Human Ether-a-go-go- Related Potassium Channel. Mol. Pharmacol. 2007, 72, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Peigneur, S.; Béress, L.; Möller, C.; Marí, F.; Forssmann, W.-G.; Tytgat, J. A natural point mutation changes both target selectivity and mechanism of action of sea anemone toxins. FASEB J. 2012, 26, 5141–5151. [Google Scholar] [CrossRef]
- Andreev, Y.A.; Kozlov, S.A.; Koshelev, S.G.; Ivanova, E.A.; Monastyrnaya, M.M.; Kozlovskaya, E.P.; Grishin, E.V. Analgesic Compound from Sea Anemone Heteractis crispa Is the First Polypeptide Inhibitor of Vanilloid Receptor 1 (TRPV1). J. Biol. Chem. 2008, 283, 23914–23921. [Google Scholar] [CrossRef] [PubMed]
- Philyppov, I.B.; Paduraru, O.N.; Andreev, Y.A.; Grishin, E.V.; Shuba, Y.M. Modulation of TRPV1-dependent contractility of normal and diabetic bladder smooth muscle by analgesic toxins from sea anemone Heteractis crispa. Life Sci. 2012, 91, 912–920. [Google Scholar] [CrossRef] [PubMed]
- Kozlov, S.A.; Andreev, I.A.; Murashev, A.N.; Skobtsov, D.I.; D’iachenko, I.A.; Grishin, E.V. New polypeptide components from the Heteractis crispa sea anemone with analgesic activity. Bioorganicheskaia Khimiia 2009, 35, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Schaloske, R.H.; Dennis, E.A. The phospholipase A2 superfamily and its group numbering system. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2006, 1761, 1246–1259. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, A.R. Chemopreventive Agents: Protease Inhibitors. Pharmacol. Ther. 1998, 78, 167–209. [Google Scholar] [CrossRef]
- Abbenante, G.; Fairlie, D. Protease Inhibitors in the Clinic. Med. Chem. 2005, 1, 71–104. [Google Scholar] [CrossRef]
- Mourão, C.; Schwartz, E. Protease Inhibitors from Marine Venomous Animals and Their Counterparts in Terrestrial Venomous Animals. Mar. Drugs 2013, 11, 2069–2112. [Google Scholar] [CrossRef]
- Macrander, J.; Broe, M.; Daly, M. Tissue-Specific Venom Composition and Differential Gene Expression in Sea Anemones. Genome Biol. Evol. 2016, 8, 2358–2375. [Google Scholar] [CrossRef] [PubMed]
- Madio, B.; Undheim, E.A.B.; King, G.F. Revisiting venom of the sea anemone Stichodactyla haddoni: Omics techniques reveal the complete toxin arsenal of a well-studied sea anemone genus. J. Proteom. 2017, 166, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Sintsova, O.; Gladkikh, I.; Chausova, V.; Monastyrnaya, M.; Anastyuk, S.; Chernikov, O.; Yurchenko, E.; Aminin, D.; Isaeva, M.; Leychenko, E.; et al. Peptide fingerprinting of the sea anemone Heteractis magnifica mucus revealed neurotoxins, Kunitz-type proteinase inhibitors and a new β-defensin α-amylase inhibitor. J. Proteom. 2018, 173, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Rivera-de-Torre, E.; Martínez-Del-Pozo, Á.; Garb, J.E. Stichodactyla helianthus’ de novo transcriptome assembly: Discovery of a new actinoporin isoform. Toxicon 2018, 150, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Surm, J.M.; Smith, H.L.; Madio, B.; Undheim, E.A.B.; King, G.F.; Hamilton, B.R.; van der Burg, C.A.; Pavasovic, A.; Prentis, P.J. A process of convergent amplification and tissue-specific expression dominates the evolution of toxin and toxin-like genes in sea anemones. Mol. Ecol. 2019, 28, 2272–2289. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Pérez, D.; Campos, A.; Alexei Rodríguez, A.; Turkina, M.V.; Ribeiro, T.; Osorio, H.; Vasconcelos, V.; Antunes, A. Proteomic Analyses of the Unexplored Sea Anemone Bunodactis verrucosa. Mar. Drugs 2018, 16, 42. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Rahman, M.A.; Quintero-Hernandez, V.; Possani, L.D. Venom proteomic and venomous glands transcriptomic analysis of the Egyptian scorpion Scorpio maurus palmatus (Arachnida: Scorpionidae). Toxicon 2013, 74, 193–207. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Zhang, D.; Zhang, Y.; Peng, L.; Chen, J.; Liang, S. Venomics of the spider Ornithoctonus huwena based on transcriptomic versus proteomic analysis. Comp. Biochem. Physiol. Part D Genom. Proteom. 2010, 5, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Himaya, S.W.A.; Jin, A.-H.; Dutertre, S.; Giacomotto, J.; Mohialdeen, H.; Vetter, I.; Alewood, P.F.; Lewis, R.J. Comparative Venomics Reveals the Complex Prey Capture Strategy of the Piscivorous Cone Snail Conus catus. J. Proteome Res. 2015, 14, 4372–4381. [Google Scholar] [CrossRef]
- Campos, P.F.; Andrade-Silva, D.; Zelanis, A.; Paes Leme, A.F.; Rocha, M.M.T.; Menezes, M.C.; Serrano, S.M.T.; Junqueira-de-Azevedo, I. de L.M. Trends in the Evolution of Snake Toxins Underscored by an Integrative Omics Approach to Profile the Venom of the Colubrid Phalotris mertensi. Genome Biol. Evol. 2016, 8, 2266–2287. [Google Scholar] [CrossRef]
- Columbus-Shenkar, Y.Y.; Sachkova, M.Y.; Macrander, J.; Fridrich, A.; Modepalli, V.; Reitzel, A.M.; Sunagar, K.; Moran, Y. Dynamics of venom composition across a complex life cycle. Elife 2018, 7, e35014. [Google Scholar] [CrossRef] [PubMed]
- Sebé-Pedrós, A.; Saudemont, B.; Chomsky, E.; Plessier, F.; Mailhé, M.-P.; Renno, J.; Loe-Mie, Y.; Lifshitz, A.; Mukamel, Z.; Schmutz, S.; et al. Cnidarian Cell Type Diversity and Regulation Revealed by Whole-Organism Single-Cell RNA-Seq. Cell 2018, 173, 1520–1534. [Google Scholar] [CrossRef]
- Honma, T.; Minagawa, S.; Nagai, H.; Ishida, M.; Nagashima, Y.; Shiomi, K. Novel peptide toxins from acrorhagi, aggressive organs of the sea anemone Actinia equina. Toxicon 2005, 46, 768–774. [Google Scholar] [CrossRef] [PubMed]
- Moran, Y.; Praher, D.; Schlesinger, A.; Ayalon, A.; Tal, Y.; Technau, U. Analysis of Soluble Protein Contents from the Nematocysts of a Model Sea Anemone Sheds Light on Venom Evolution. Mar. Biotechnol. 2013, 15, 329–339. [Google Scholar] [CrossRef]
- Tang, P.-C.; Watson, G.M. Proteomic identification of hair cell repair proteins in the model sea anemone Nematostella vectensis. Hear. Res. 2015, 327, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Evans, T.G. Considerations for the use of transcriptomics in identifying the “genes that matter” for environmental adaptation. J. Exp. Biol. 2015, 218, 1925–1935. [Google Scholar] [CrossRef]
- Hamdan, M.; Righetti, P.G. Modern strategies for protein quantification in proteome analysis: Advantages and limitations. Mass Spectrom. Rev. 2002, 21, 287–302. [Google Scholar] [CrossRef]
- Rodríguez, A.A.; Cassoli, J.S.; Sa, F.; Dong, Z.Q.; de Freitas, J.C.; Pimenta, A.M.C.; de Lima, M.E.; Konno, K.; Lee, S.M.Y.; Garateix, A.; et al. Peptide fingerprinting of the neurotoxic fractions isolated from the secretions of sea anemones Stichodactyla helianthus and Bunodosoma granulifera. New members of the APETx-like family identified by a 454 pyrosequencing approach. Peptides 2012, 34, 26–38. [Google Scholar] [CrossRef]
- Ramírez-Carreto, S.; Pérez-García, E.I.; Salazar-García, S.I.; Bernáldez-Sarabia, J.; Licea-Navarro, A.; Rudiño-Piñera, E.; Pérez-Martínez, L.; Pedraza-Alva, G.; Rodríguez-Almazán, C. Identification of a pore-forming protein from sea anemone Anthopleura dowii Verrill (1869) venom by mass spectrometry. J. Venom. Anim. Toxins Incl. Trop. Dis. 2019, 25, 25. [Google Scholar] [CrossRef]
- Alvarado-Mesén, J.; Solano-Campos, F.; Canet, L.; Pedrera, L.; Hervis, Y.P.; Soto, C.; Borbón, H.; Lanio, M.E.; Lomonte, B.; Valle, A.; et al. Cloning, purification and characterization of nigrelysin, a novel actinoporin from the sea anemone Anthopleura nigrescens. Biochimie 2019, 156, 206–223. [Google Scholar] [CrossRef]
- Moreels, L.; Peigneur, S.; Galan, D.T.; De Pauw, E.; Béress, L.; Waelkens, E.; Pardo, L.A.; Quinton, L.; Tytgat, J. APETx4, a Novel Sea Anemone Toxin and a Modulator of the Cancer-Relevant Potassium Channel KV10.1. Mar. Drugs 2017, 15, 287. [Google Scholar] [CrossRef]
- Minagawa, S.; Sugiyama, M.; Ishida, M.; Nagashima, Y.; Shiomi, K. Kunitz-type protease inhibitors from acrorhagi of three species of sea anemones. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2008, 150, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Ayala-Sumuano, J.-T.; Licea-Navarro, A.; Rudiño-Piñera, E.; Rodríguez, E.; Rodríguez-Almazán, C. Sequencing and de novo transcriptome assembly of Anthopleura dowii Verrill (1869), from Mexico. Genom. Data 2017, 11, 92–94. [Google Scholar] [CrossRef]
- Ponce, D.; Brinkman, D.L.; Potriquet, J.; Mulvenna, J. Tentacle Transcriptome and Venom Proteome of the Pacific Sea Nettle, Chrysaora fuscescens (Cnidaria: Scyphozoa). Toxins 2016, 8, 102. [Google Scholar] [CrossRef]
- De Oliveira, U.C.; Nishiyama, M.Y., Jr.; dos Santos, M.B.V.; de Paula Santos-da-Silva, A.; de Menezes Chalkidis, H.; Souza-Imberg, A.; Candido, D.M.; Yamanouye, N.; Dorce, V.A.C.; Junqueira-de, I.D.L.M. Proteomic endorsed transcriptomic profiles of venom glands from Tityus obscurus and T. serrulatus scorpions. PLoS ONE 2018, 13, e0193739. [Google Scholar] [CrossRef] [PubMed]
- Von Reumont, B.; Undheim, E.; Jauss, R.-T.; Jenner, R. Venomics of Remipede Crustaceans Reveals Novel Peptide Diversity and Illuminates the Venom’s Biological Role. Toxins 2017, 9, 234. [Google Scholar] [CrossRef] [PubMed]
- Whitelaw, B.L.; Strugnell, J.M.; Faou, P.; da Fonseca, R.R.; Hall, N.E.; Norman, M.; Finn, J.; Cooke, I.R. Combined Transcriptomic and Proteomic Analysis of the Posterior Salivary Gland from the Southern Blue-Ringed Octopus and the Southern Sand Octopus. J. Proteome Res. 2016, 15, 3284–3297. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Hinz, U.; Consortium, T.U. From protein sequences to 3D-structures and beyond: The example of the UniProt Knowledgebase. Cell. Mol. Life Sci. 2010, 67, 1049–1064. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, I.; Lee, H.; Pyo, M.-J.; Heo, Y.; Bae, S.K.; Kwon, Y.C.; Yoon, W.D.; Kang, C.; Kim, E. Proteomics approach to examine the cardiotoxic effects of Nemopilema nomurai Jellyfish venom. J. Proteom. 2015, 128, 123–131. [Google Scholar] [CrossRef]
- Augusto-de-Oliveira, C.; Stuginski, D.R.; Kitano, E.S.; Andrade-Silva, D.; Liberato, T.; Fukushima, I.; Serrano, S.M.T.; Zelanis, A. Dynamic Rearrangement in Snake Venom Gland Proteome: Insights into Bothrops jararaca Intraspecific Venom Variation. J. Proteom. 2016, 15, 3752–3762. [Google Scholar] [CrossRef] [PubMed]
- Mathé-Hubert, H.; Colinet, D.; Deleury, E.; Belghazi, M.; Ravallec, M.; Poulain, J.; Dossat, C.; Poirié, M.; Gatti, J.-L. Comparative venomics of Psyttalia lounsburyi and P. concolor, two olive fruit fly parasitoids: A hypothetical role for a GH1 β-glucosidase. Sci. Rep. 2016, 6, 35873. [Google Scholar] [CrossRef] [PubMed]
- Gundlach, K.A.; Watson, G.M. Self/Non-Self Recognition Affects Cnida Discharge and Tentacle Contraction in the Sea Anemone Haliplanella luciae. Biol. Bull. 2018, 235, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Graves, P.R.; Haystead, T.A.J. Molecular biologist’s guide to proteomics. Microbiol. Mol. Biol. Rev. 2002, 66, 39–63. [Google Scholar] [CrossRef] [PubMed]
- Benzinger, G.R.; Drum, C.L.; Chen, L.-Q.; Kallen, R.G.; Hanck, D.A.; Hanck, D. Differences in the binding sites of two site-3 sodium channel toxins. Pflügers Arch. Eur. J. Physiol. 1997, 434, 742–749. [Google Scholar] [CrossRef] [PubMed]
- Moran, Y.; Gordon, D.; Gurevitz, M. Sea anemone toxins affecting voltage-gated sodium channels – molecular and evolutionary features. Toxicon 2009, 54, 1089–1101. [Google Scholar] [CrossRef] [PubMed]
- Peigneur, S.; Billen, B.; Derua, R.; Waelkens, E.; Debaveye, S.; Béress, L.; Tytgat, J. A bifunctional sea anemone peptide with Kunitz type protease and potassium channel inhibiting properties. Biochem. Pharm. 2011, 82, 81–90. [Google Scholar] [CrossRef]
- Minagawa, S.; Ishida, M.; Shimakura, K.; Nagashima, Y.; Shiomi, K. Isolation and Amino Acid Sequences of Two Kunitz-Type Protease Inhibitors From the Sea Anemone Anthopleura aff. xanthogrammica. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1997, 118, 381–386. [Google Scholar] [CrossRef]
- Strukelj, B.; Lenarcic, B.; Gruden, K.; Pungercar, J.; Rogelj, B.; Turk, V.; Bosch, D.; Jongsma, M.A. Equistatin, a protease inhibitor from the sea anemone actinia equina, is composed of three structural and functional domains. Biochem. Biophys. Res. Commun. 2000, 269, 732–736. [Google Scholar] [CrossRef]
- Galea, K.; trukelj, B.; Bavec, S.; Turk, V.; Lenarcic, B. Cloning and Expression of Functional Equistatin. Biol. Chem. 2005, 381, 85–88. [Google Scholar] [CrossRef]
- Lenarcic, B.; Ritonja, A.; Strukelj, B.; Turk, B.; Turk, V. Equistatin, a new inhibitor of cysteine proteinases from Actinia equina, is structurally related to thyroglobulin type-1 domain. J. Biol. Chem. 1997, 272, 13899–13903. [Google Scholar] [CrossRef] [PubMed]
- Lenarcic, B.; Turk, V. Thyroglobulin type-1 domains in equistatin inhibit both papain-like cysteine proteinases and cathepsin D. J. Biol. Chem. 1999, 274, 563–566. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, C.; Sheng, Z.; Horan, T.P.; Kitayama, H.; Maki, M.; Hitomi, K.; Kitaura, Y.; Takai, S.; Sasahara, R.M.; Horimoto, A.; et al. Regulation of matrix metalloproteinase-9 and inhibition of tumor invasion by the membrane-anchored glycoprotein RECK. Proc. Natl. Acad. Sci. USA 1998, 95, 13221–13226. [Google Scholar] [CrossRef] [PubMed]
- Mahl, C.; Egea, V.; Megens, R.T.A.; Pitsch, T.; Santovito, D.; Weber, C.; Ries, C. RECK (reversion-inducing cysteine-rich protein with Kazal motifs) regulates migration, differentiation and Wnt/β-catenin signaling in human mesenchymal stem cells. Cell. Mol. Life Sci. 2016, 73, 1489–1501. [Google Scholar] [CrossRef] [PubMed]
- Markland, F.S.; Swenson, S. Snake venom metalloproteinases. Toxicon 2013, 62, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Takeda, S.; Takeya, H.; Iwanaga, S. Snake venom metalloproteinases: Structure, function and relevance to the mammalian ADAM/ADAMTS family proteins. Biochim. Biophys. Acta Proteins Proteom. 2012, 1824, 164–176. [Google Scholar] [CrossRef] [PubMed]
- Trevisan-Silva, D.; Gremski, L.H.; Chaim, O.M.; da Silveira, R.B.; Meissner, G.O.; Mangili, O.C.; Barbaro, K.C.; Gremski, W.; Veiga, S.S.; Senff-Ribeiro, A. Astacin-like metalloproteases are a gene family of toxins present in the venom of different species of the brown spider (genus Loxosceles). Biochimie 2010, 92, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Yu, H.; Xue, W.; Yue, Y.; Liu, S.; Xing, R.; Li, P. Jellyfish venomics and venom gland transcriptomics analysis of Stomolophus meleagris to reveal the toxins associated with sting. J. Proteom. 2014, 106, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Jouiaei, M.; Casewell, N.; Yanagihara, A.; Nouwens, A.; Cribb, B.; Whitehead, D.; Jackson, T.; Ali, S.; Wagstaff, S.; Koludarov, I.; et al. Firing the Sting: Chemically Induced Discharge of Cnidae Reveals Novel Proteins and Peptides from Box Jellyfish (Chironex fleckeri) Venom. Toxins 2015, 7, 936–950. [Google Scholar] [CrossRef]
- Wenger, Y.; Buzgariu, W.; Reiter, S.; Galliot, B. Injury-induced immune responses in Hydra. Semin. Immunol. 2014, 26, 277–294. [Google Scholar] [CrossRef]
- Skejić, J.; Hodgson, W.C. Population Divergence in Venom Bioactivities of Elapid Snake Pseudonaja textilis: Role of Procoagulant Proteins in Rapid Rodent Prey Incapacitation. PLoS ONE 2013, 8, e63988. [Google Scholar] [CrossRef] [PubMed]
- Teng, Z.-W.; Xiong, S.-J.; Xu, G.; Gan, S.-Y.; Chen, X.; Stanley, D.; Yan, Z.-C.; Ye, G.-Y.; Fang, Q. Protein Discovery: Combined Transcriptomic and Proteomic Analyses of Venom from the Endoparasitoid Cotesia chilonis (Hymenoptera: Braconidae). Toxins 2017, 9, 135. [Google Scholar] [CrossRef] [PubMed]
- Key, L.N.; Boyle, P.R.; Jaspars, M. Novel activities of saliva from the octopus Eledone cirrhosa (Mollusca; Cephalopoda). Toxicon 2002, 40, 677–683. [Google Scholar] [CrossRef]
- Xie, B.; Li, X.; Lin, Z.; Ruan, Z.; Wang, M.; Liu, J.; Tong, T.; Li, J.; Huang, Y.; Wen, B.; et al. Prediction of Toxin Genes from Chinese Yellow Catfish Based on Transcriptomic and Proteomic Sequencing. Int. J. Mol. Sci. 2016, 17, 556. [Google Scholar] [CrossRef] [PubMed]
- Quintero-Hernández, V.; Ramírez-Carreto, S.; Romero-Gutiérrez, M.T.; Valdez-Velázquez, L.L.; Becerril, B.; Possani, L.D.; Ortiz, E. Transcriptome Analysis of Scorpion Species Belonging to the Vaejovis Genus. PLoS ONE 2015, 10, e0117188. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.D.; Safavi-Hemami, H.; McIntosh, L.D.; Purcell, A.W.; Norton, R.S.; Papenfuss, A.T. Diversity of Conotoxin Gene Superfamilies in the Venomous Snail, Conus victoriae. PLoS ONE 2014, 9, e87648. [Google Scholar] [CrossRef] [PubMed]
- Beeton, C.; Pennington, M.W.; Wulff, H.; Singh, S.; Nugent, D.; Crossley, G.; Khaytin, I.; Calabresi, P.A.; Chen, C.Y.; Gutman, G.A.; et al. Targeting effector memory T cells with a selective peptide inhibitor of Kv1.3 channels for therapy of autoimmune diseases. Mol. Pharmacol. 2005, 67, 1369–1381. [Google Scholar] [CrossRef] [PubMed]
- Pennington, M.W.; Byrnes, M.E.; Zaydenberg, I.; Khaytin, I.; De, J.C.; Krafte, D.S.; Hill, R.; Mahnir, V.M.; Volberg, W.A.; Gorczyca, W. Chemical synthesis and characterization of ShK toxin: A potent potassium channel inhibitor from a sea anemone. Int. J. Pept. Protein. Res. 1995, 46, 354–358. [Google Scholar] [CrossRef] [PubMed]
- Minagawa, S.; Ishida, M.; Nagashima, Y.; Shiomi, K. Primary structure of a potassium channel toxin from the sea anemone Actinia equina. FEBS Lett. 1998, 427, 149–151. [Google Scholar] [CrossRef]
- Schweitz, H.; Bruhn, T.; Guillemare, E.; Moinier, D.; Lancelin, J.M.; Béress, L.; Lazdunski, M. Kalicludines and kaliseptine. Two different classes of sea anemone toxins for voltage sensitive K+ channels. J. Biol. Chem. 1995, 270, 25121–25126. [Google Scholar] [CrossRef] [PubMed]
- Diochot, S.; Schweitz, H.; Béress, L.; Lazdunski, M. Sea Anemone Peptides with a Specific Blocking Activity against the Fast Inactivating Potassium Channel Kv3.4. J. Biol. Chem. 1998, 273, 6744–6749. [Google Scholar] [CrossRef] [PubMed]
- Jouiaei, M.; Sunagar, K.; Federman Gross, A.; Scheib, H.; Alewood, P.F.; Moran, Y.; Fry, B.G. Evolution of an Ancient Venom: Recognition of a Novel Family of Cnidarian Toxins and the Common Evolutionary Origin of Sodium and Potassium Neurotoxins in Sea Anemone. Mol. Biol. Evol. 2015, 32, 1598–1610. [Google Scholar] [CrossRef] [PubMed]
- Yeung, S.Y.; Thompson, D.; Wang, Z.; Fedida, D.; Robertson, B. Modulation of Kv3 subfamily potassium currents by the sea anemone toxin BDS: Significance for CNS and biophysical studies. J. Neurosci. 2005, 25, 8735–8745. [Google Scholar] [CrossRef] [PubMed]
- Jensen, J.E.; Mobli, M.; Brust, A.; Alewood, P.F.; King, G.F.; Rash, L.D. Cyclisation Increases the Stability of the Sea Anemone Peptide APETx2 but Decreases Its Activity at Acid-Sensing Ion Channel 3. Mar. Drugs 2012, 10, 1511–1527. [Google Scholar] [CrossRef] [PubMed]
- Orts, D.J.; Moran, Y.; Cologna, C.T.; Peigneur, S.; Madio, B.; Praher, D.; Quinton, L.; De, E.P.; Bicudo, J.E.; Tytgat, J.; et al. BcsTx3 is a founder of a novel sea anemone toxin family of potassium channel blocker. FEBS J. 2013, 280, 4839–4852. [Google Scholar] [CrossRef]
- Razpotnik, A.; Krizaj, I.; Sribar, J.; Kordis, D.; Macek, P.; Frangez, R.; Kem, W.R.; Turk, T. A new phospholipase A2 isolated from the sea anemone Urticina crassicornis - its primary structure and phylogenetic classification. FEBS J. 2010, 277, 2641–2653. [Google Scholar] [CrossRef] [PubMed]
- Talvinen, K.A.; Nevalainen, T.J. Cloning of a novel phospholipase A2 from the cnidarian Adamsia carciniopados. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2002, 132, 571–578. [Google Scholar] [CrossRef]
- Miyoshi, S.; Tu, A.T. Phospholipase A2fromNaja naja sputatrixVenom Is a Muscarinic Acetylcholine Receptor Inhibitor. Arch. Biochem. Biophys. 1996, 328, 17–25. [Google Scholar] [CrossRef]
- Shayman, J.A.; Tesmer, J.J.G. Lysosomal phospholipase A2. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2018, 1864, 932–940. [Google Scholar] [CrossRef]
- Hiraoka, M.; Abe, A.; Shayman, J.A. Cloning and characterization of a lysosomal phospholipase A2, 1-O-acylceramide synthase. J. Biol. Chem. 2002, 277, 10090–10099. [Google Scholar] [CrossRef]
- Ramírez-Carreto, S.; Jiménez-Vargas, J.M.; Rivas-Santiago, B.; Corzo, G.; Possani, L.D.; Becerril, B.; Ortiz, E. Peptides from the scorpion Vaejovis punctatus with broad antimicrobial activity. Peptides 2015, 73, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Mariottini, G.; Grice, I. Antimicrobials from Cnidarians. A New Perspective for Anti-Infective Therapy? Mar. Drugs 2016, 14, 48. [Google Scholar] [CrossRef] [PubMed]
- Perumal Samy, R.; Stiles, B.G.; Franco, O.L.; Sethi, G.; Lim, L.H.K. Animal venoms as antimicrobial agents. Biochem. Pharmacol. 2017, 134, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Bingle, C.D.; Craven, C.J. PLUNC: A novel family of candidate host defence proteins expressed in the upper airways and nasopharynx. Hum. Mol. Genet. 2002, 11, 937–943. [Google Scholar] [CrossRef] [PubMed]
- Bingle, C.D.; Craven, C.J. Meet the relatives: A family of BPI- and LBP-related proteins. Trends Immunol. 2004, 25, 53–55. [Google Scholar] [CrossRef] [PubMed]
- Bingle, C.D.; Craven, C.J. Comparative analysis of the PLUNC (palate, lung and nasal epithelium clone) protein families. Biochem. Soc. Trans. 2003, 31, 806–809. [Google Scholar] [CrossRef] [PubMed]
- Wilde, C.G.; Seilhamer, J.J.; McGrogan, M.; Ashton, N.; Snable, J.L.; Lane, J.C.; Leong, S.R.; Thornton, M.B.; Miller, K.L.; Scott, R.W. Bactericidal/permeability-increasing protein and lipopolysaccharide (LPS)-binding protein. LPS binding properties and effects on LPS-mediated cell activation. J. Biol. Chem. 1994, 269, 17411–17416. [Google Scholar]
- Li, B.; Calvo, E.; Marinotti, O.; James, A.A.; Paskewitz, S.M. Characterization of the c-type lysozyme gene family in Anopheles gambiae. Gene 2005, 360, 131–139. [Google Scholar] [CrossRef]
- Ramos-Silva, P.; Kaandorp, J.; Huisman, L.; Marie, B.; Zanella-Cléon, I.; Guichard, N.; Miller, D.J.; Marin, F. The skeletal proteome of the coral Acropora millepora: The evolution of calcification by co-option and domain shuffling. Mol. Biol. Evol. 2013, 30, 2099–2112. [Google Scholar] [CrossRef]
- Bommer, U.-A.; Thiele, B.-J. The translationally controlled tumour protein (TCTP). Int. J. Biochem. Cell Biol. 2004, 36, 379–385. [Google Scholar] [CrossRef]
- Mitchell, M.L.; Hamilton, B.R.; Madio, B.; Morales, R.A.V.; Tonkin-Hill, G.Q.; Papenfuss, A.T.; Purcell, A.W.; King, G.F.; Undheim, E.A.B.; Norton, R.S. The Use of Imaging Mass Spectrometry to Study Peptide Toxin Distribution in Australian Sea Anemones. Aust. J. Chem. 2017, 70, 1235–1237. [Google Scholar] [CrossRef]
- Lauretta, D.M.; Häussermann, V.; Brugler, M.R.; Rodríguez, E. Isoparactis fionae sp. nov. (Cnidaria: Anthozoa: Actiniaria) from Southern Patagonia, with a discussion of the family Isanthidae. Org. Divers. Evol. 2014, 14, 31–42. [Google Scholar]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Nesvizhskii, A.I.; Keller, A.; Kolker, E.; Aebersold, R. A Statistical Model for Identifying Proteins by Tandem Mass Spectrometry. Anal. Chem. 2003, 75, 4646–4658. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Huang, X. A Time-Efficient, Linear-Space Local Similarity Algorithm. Adv. Appl. Math. 1991, 12, 337–357. [Google Scholar] [CrossRef]
- Moran, Y.; Genikhovich, G.; Gordon, D.; Wienkoop, S.; Zenkert, C.; Özbek, S.; Technau, U.; Gurevitz, M. Neurotoxin localization to ectodermal gland cells uncovers an alternative mechanism of venom delivery in sea anemones. Proc. Biol. Sci. 2012, 279, 1351–1358. [Google Scholar] [CrossRef] [PubMed]
- Macrander, J.; Brugler, M.R.; Daly, M. A RNA-seq approach to identify putative toxins from acrorhagi in aggressive and non-aggressive Anthopleura elegantissima polyps. BMC Genom. 2015, 16, 221. [Google Scholar] [CrossRef]
- Sachkova, M.Y.; Singer, S.A.; Macrander, J.; Reitzel, A.M.; Peigneur, S.; Tytgat, J.; Moran, Y. The birth and death of toxins with distinct functions: A case study in the sea anemone Nematostella. Mol. Biol. Evol. 2019, 1–12. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Putative Conserved Domain Detected a | Uniprot Accession b | Protein Name c | ID-Transcript | Sample M/T d | Organism | % Coverage M/T e |
|---|---|---|---|---|---|---|
| Sea anemone sodium channel inhibitory toxin family. Type I subfamily | P0DL52 | Delta-actitoxin-Avd1e 1 | Unidentified | M/- | Anemonia viridis | 17/- |
| P0C1F0 | Delta-actitoxin-Ael1b | Unidentified | M/- | Anthopleura elegantissima | 100/- | |
| P0C5G1 | Delta-actitoxin-Axm1f | Unidentified | M/- | Anthopleura xanthogrammica | 45/- | |
| P0C1F1 | Delta-actitoxin-Ael1c | Unidentified | M/- | Anthopleura elegantissima | 100/- | |
| P01530 | Delta-actitoxin-Axm1a | c22149_g1 | M/- | Anthopleura xanthogrammica | 29/- | |
| Venom Kunitz-type family. Sea anemone type 2 potassium channel toxin subfamily | P86862 | KappaPI-actitoxin-Ael3a | c14874_g1 | M/- | Anthopleura elegantissima | 26/- |
| Serpin family. Ov-serpin subfamily | Q52L45 | Leukocyte elastase inhibitor | c27265_g1 | M/- | Xenopus laevis | 3/- |
| Thyroglobulin_1 | P81439 | Equistatin | Unidentified | M/- | Actinia equina | 9/- |
| Aerolysin family | P09167 | Aerolysin-like | Unidentified | M/- | Aeromonas hydrophila | 1/- |
| Peptidase S1 family | A7RW61 | Predicted protein | Unidentified | M/- | Nematostella vectensis | 5/- |
| A7S5Y0 | Predicted protein | c28983_g1 | M/- | Nematostella vectensis | 7/- | |
| B8V7S0 | CUB and peptidase domain-containing protein 1 | Unidentified | M/- | Acropora millepora | 3/- | |
| Peptidase_M12A | A7SQR7 | Metallo- endopeptidase | Unidentified | M/- | Nematostella vectensis | 3/- |
| Peptidase_ M14 | T2M3L7 | Carboxypeptidase A4 | c39288_g1 | M/- | Hydra vulgaris | 7/- |
| Glycosyl hydrolase 18 family | A0A1T4JGY1 | Chitinase-C enzyme | Unidentified | M/- | Nematostella vectensis | 8/- |
| A0A1T4JH12 | Chitinase-A enzyme | Unidentified | M/- | Nematostella vectensis | 15/- | |
| A0A1T4JH12 | Chitinase-A enzyme | Unidentified | M/- | Nematostella vectensis | 15/- | |
| Glycosyl hydrolase 20 family | A7RSQ4 | Predicted protein | c30078_g1_i1 | M/- | Nematostella vectensis | 3/- |
| Factor 5/8 C-Domain Proteins | A7RK24 | Predicted protein | Unidentified | M/- | Nematostella vectensis | 6/- |
| Peptidase_M1 | A7RUV9 | Aminopeptidase | Unidentified | M/T | Nematostella vectensis | 3/3 |
| Peptidase_M17 | A7SGM8 | Predicted protein | c27931_g1_i1 | M/T | Nematostella vectensis | 8/6 |
| Peptidase_M20 | A7RZC0 | Predicted protein | c31795_g1_i2 | M/T | Nematostella vectensis | 11/9 |
| Peptidase C2 family | T1E719 | Calpain-1 catalytic subunit-like protein. | c27362_g1_i1 | M/T | Crotalus horridus | 5/5 |
| ID-Transcript | Aminoacid SEQUENCE a | E-value PSI-BLAST | Protein Identity (%) b | Protein Family | UniProtKB Accession |
|---|---|---|---|---|---|
| c23125_g1_i1 | MNSKLVIVFLLCAILVVSVTSRRVRTWDDFERDQDYEEEPAPYGKRACKDNYSAATCKDVKKNNNCGSEKYATNCAKTCGKC | 2e-14 | Kappa-actitoxin-Avd6a (83.3) | Sea anemone type 1 potassium channel toxin family. Type 1b subfamily. | Q9TWG1 |
| c29930_g1_i1 | MSYQRFLFLVVVASLIATSLAVPKDLEERGTTCSCGNTKGIYWFFLKTCPSDRGYTGSCNYFFGICCYPVD | 1e-17 | Pi-AITX-Ael2b (71.4) | Sea anemone type 3 (BDS) potassium channel toxin family. | P61542 |
| c30503_g2_i5 | MAAKSVLMMLAIFMALLLLANGEEAQGEVRIKARALSCNCGKEDNAPSGDWWLWRSSCPGGYGYTSSCNAGFGNICCLPRG | 7e-06 | DeltaKappa-AITX-Avd4b (52.1) | Sea anemone type 3 (BDS) potassium channel toxin family. | P59084 |
| c33344_g1 | MKTLVVFLVVAVIVVNAYRIKEEYEDEMAPELERRACKKKWNECTRDSDCCDEKGWANQKLQCLQQCDEGGCLEYRQCLFHSGLQRK | 3e-20 | Kappa-actitoxin-Bcs4a (72.0) | Sea anemone type 5 potassium channel toxin family. | C0HJC4 |
| ID-Transcript | Aminoacid Sequence a | E-value PSI-BLAST | Protein Identity (%) b | Protein Family | UniProtKB Accession |
|---|---|---|---|---|---|
| c54261_g1 | MMMMKKKSTTTLIVLLGMAFLVEGLSLNNLEDDKRMNVKTGDGRAEKRNLWQFGNMIKCATGRDAGDYNGYGNYCGWGGSGVPVDGVDRCCQAHDRCYDNHDSCNPKTNYYSYSKSGKHPSCTISCGDSTQNDQCERNVCSCDKVAAECFARNNYNNANKH | 6e-57 | A2-AITX-Ucs2a (59.0) | Phospholipase A2 family. | A7LCJ2 |
| c3686_g1_i1 | MGALKLLVLLAVVACVACTSLDLGKLKKKSLSKALKTQVHTRARRSLYEFYKMITCETGRSWQDYNLYGCFCGKGGTGTPVDALDQCCFDHDECYSQAAASVCPWPLQIYLDSYWHKNCSECDASKNSACEQALCECDSKAARCFKNNKWDPQYDDYPQEKCA | 3e-22 | Neutral phospholipase A2 muscarinic inhibitor (41.9) | Phospholipase A2 family. Group I subfamily. | Q92084 |
| c26312_g1 | MDSYVSKIFVILAVILHASLCQAMYDWKTKTFIRKDNSKLVVPGTKWCGKGNNAMSFDDLGEHRETDLCCREHDHCPTYILPFQRRFGILNLYPSHLSLCSCEMKLYNCLWNVTSHVAVAVGRMYFNVLRVPCFHLVEKKVCKERSFDWWKFKYVCKKYGVEVKGQTFMPKRFHKQLQVQPSNWNATANGTM | 1e-30 | Phospholipase A2 isozymes PA3A/PA3B/PA5 (34.4) | Phospholipase A2 family. Group III subfamily. | P16354 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramírez-Carreto, S.; Vera-Estrella, R.; Portillo-Bobadilla, T.; Licea-Navarro, A.; Bernaldez-Sarabia, J.; Rudiño-Piñera, E.; Verleyen, J.J.; Rodríguez, E.; Rodríguez-Almazán, C. Transcriptomic and Proteomic Analysis of the Tentacles and Mucus of Anthopleura dowii Verrill, 1869. Mar. Drugs 2019, 17, 436. https://doi.org/10.3390/md17080436
Ramírez-Carreto S, Vera-Estrella R, Portillo-Bobadilla T, Licea-Navarro A, Bernaldez-Sarabia J, Rudiño-Piñera E, Verleyen JJ, Rodríguez E, Rodríguez-Almazán C. Transcriptomic and Proteomic Analysis of the Tentacles and Mucus of Anthopleura dowii Verrill, 1869. Marine Drugs. 2019; 17(8):436. https://doi.org/10.3390/md17080436
Chicago/Turabian StyleRamírez-Carreto, Santos, Rosario Vera-Estrella, Tobías Portillo-Bobadilla, Alexei Licea-Navarro, Johanna Bernaldez-Sarabia, Enrique Rudiño-Piñera, Jerome J. Verleyen, Estefanía Rodríguez, and Claudia Rodríguez-Almazán. 2019. "Transcriptomic and Proteomic Analysis of the Tentacles and Mucus of Anthopleura dowii Verrill, 1869" Marine Drugs 17, no. 8: 436. https://doi.org/10.3390/md17080436
APA StyleRamírez-Carreto, S., Vera-Estrella, R., Portillo-Bobadilla, T., Licea-Navarro, A., Bernaldez-Sarabia, J., Rudiño-Piñera, E., Verleyen, J. J., Rodríguez, E., & Rodríguez-Almazán, C. (2019). Transcriptomic and Proteomic Analysis of the Tentacles and Mucus of Anthopleura dowii Verrill, 1869. Marine Drugs, 17(8), 436. https://doi.org/10.3390/md17080436