Characterization of the First Conotoxin from Conus ateralbus, a Vermivorous Cone Snail from the Cabo Verde Archipelago


 ,
,  and
and
Abstract
1. Introduction
2. Results
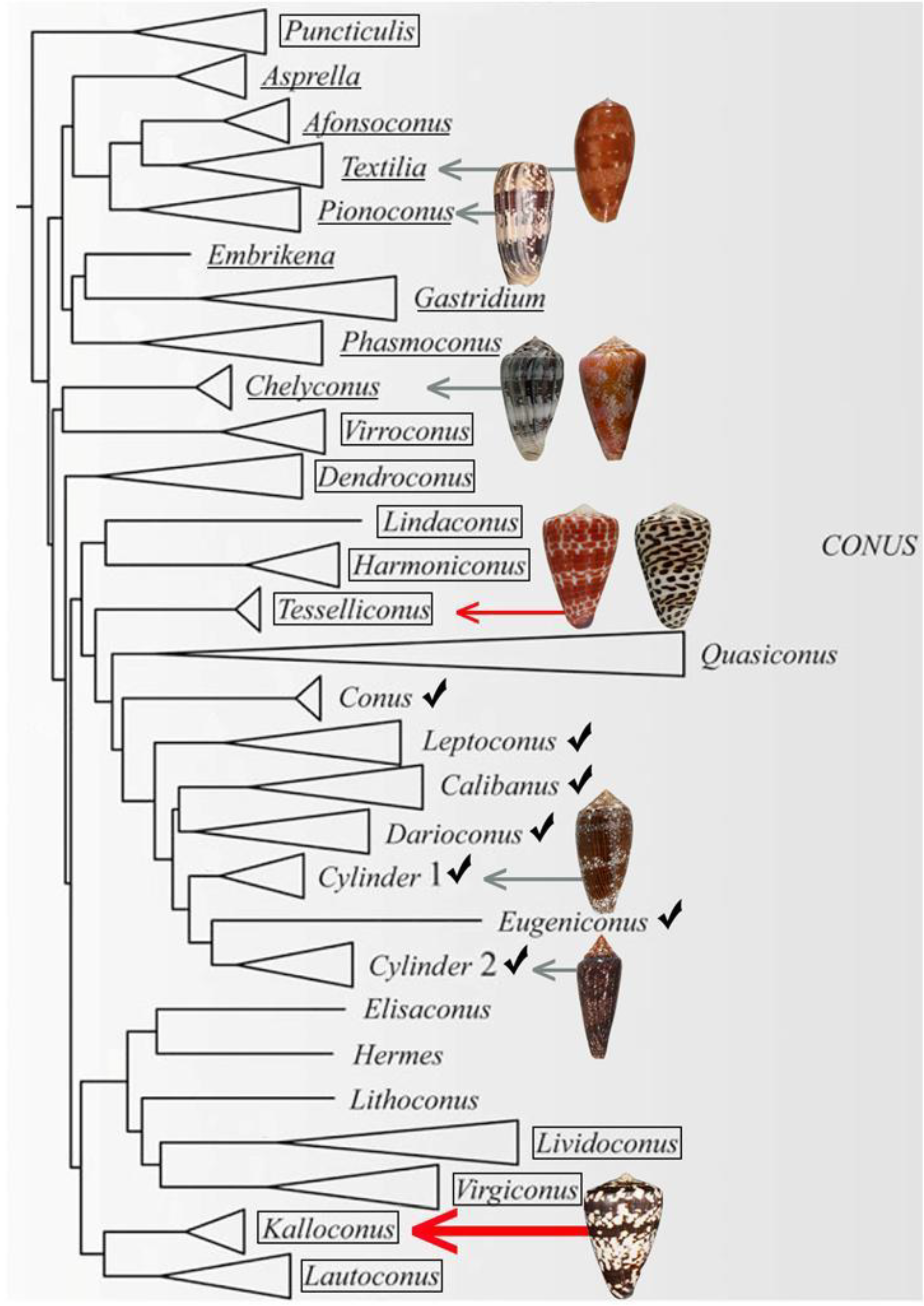
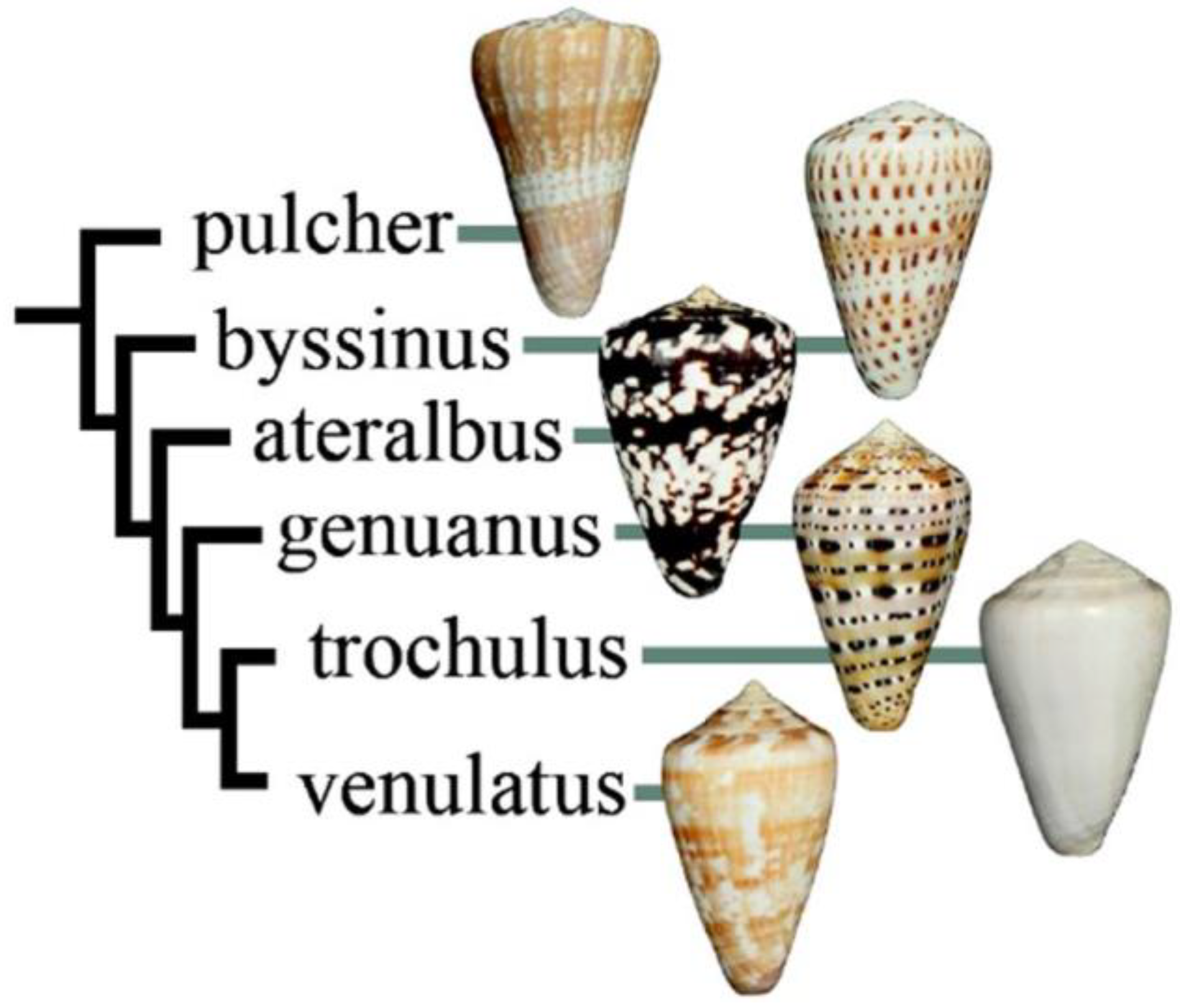
2.1. Collection of Conus Ateralbus and Venom Fractionation
2.2. Conus Ateralbus Venom: Biological Activity
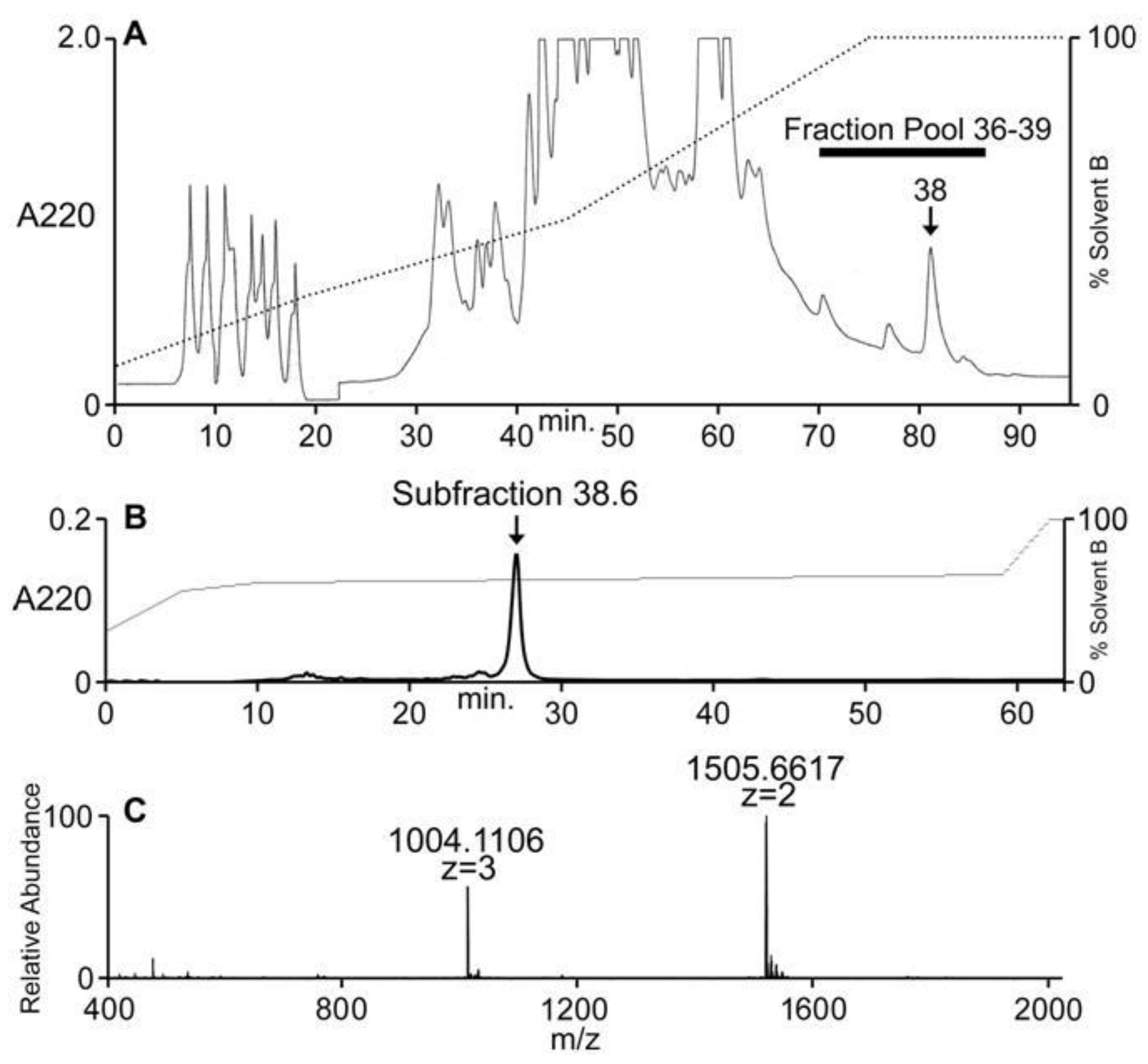
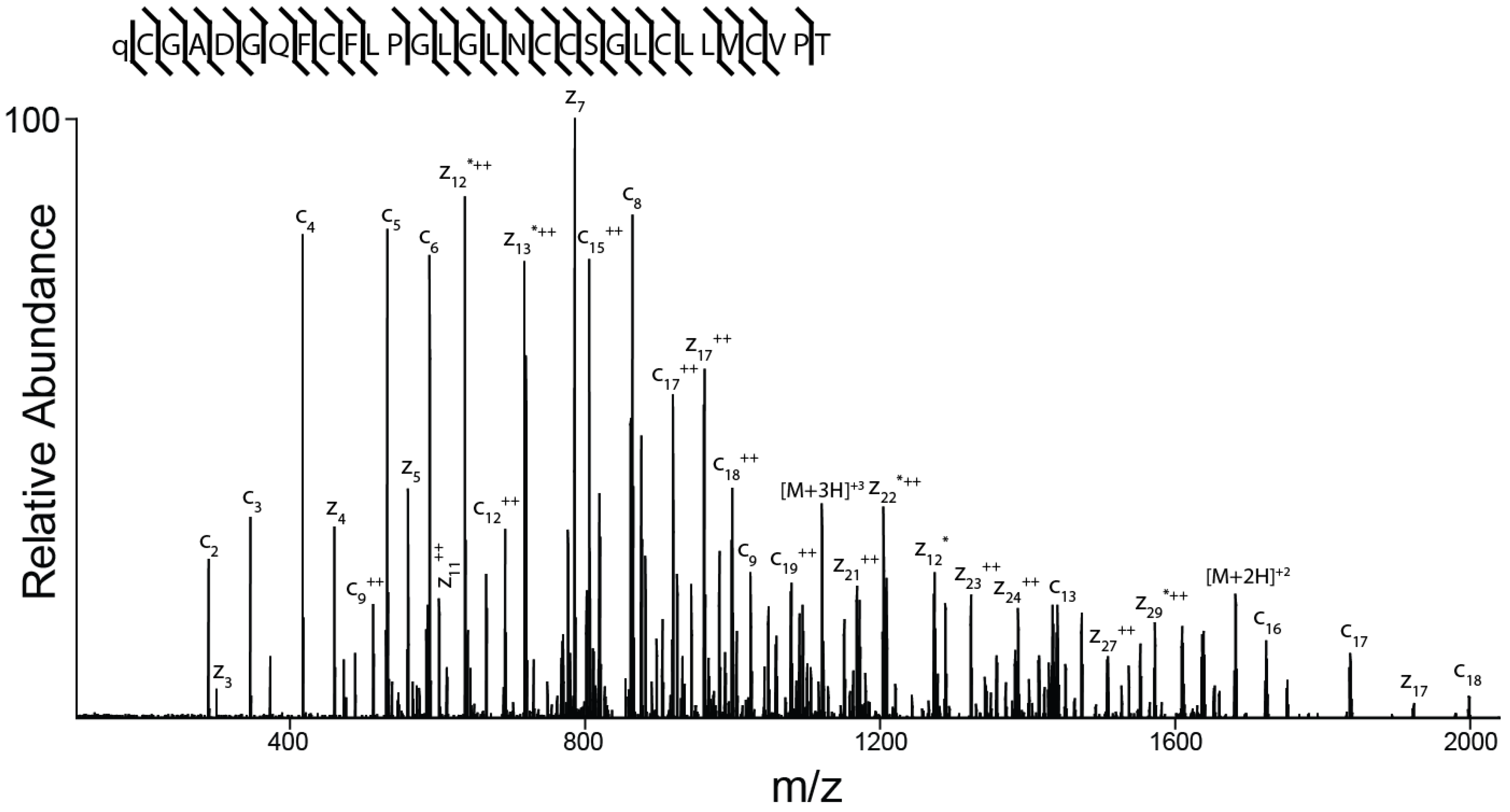
2.3. Peptide Sequence Determination
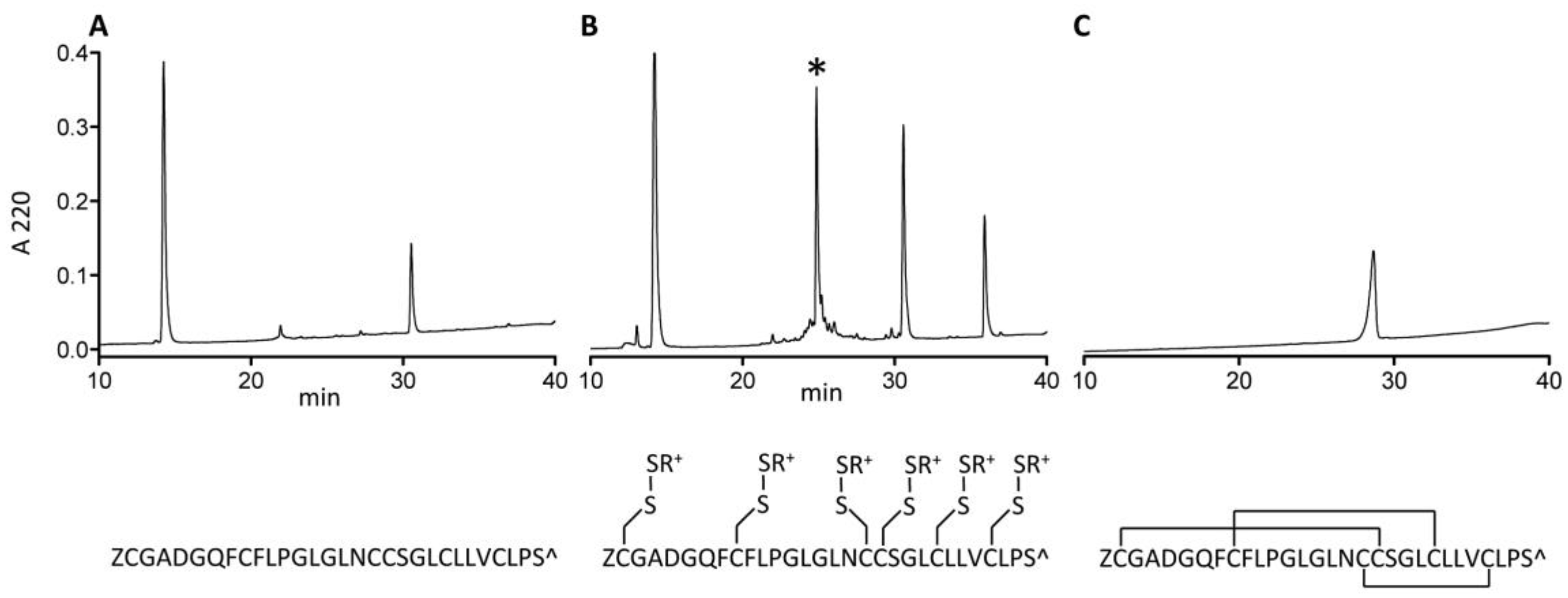
2.4. Synthesis and Folding of AtVIA[I25L;V28L;T30S]
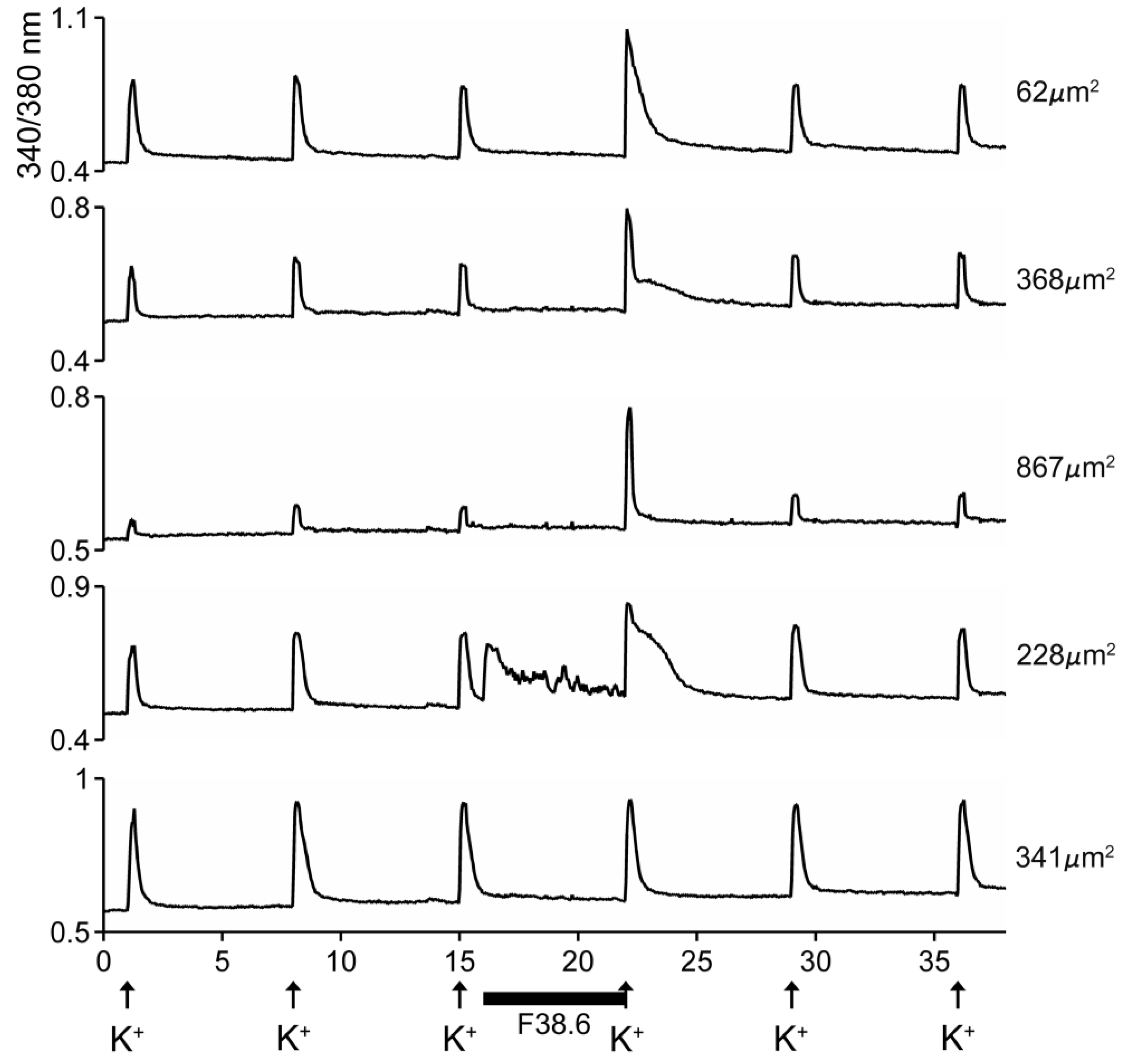
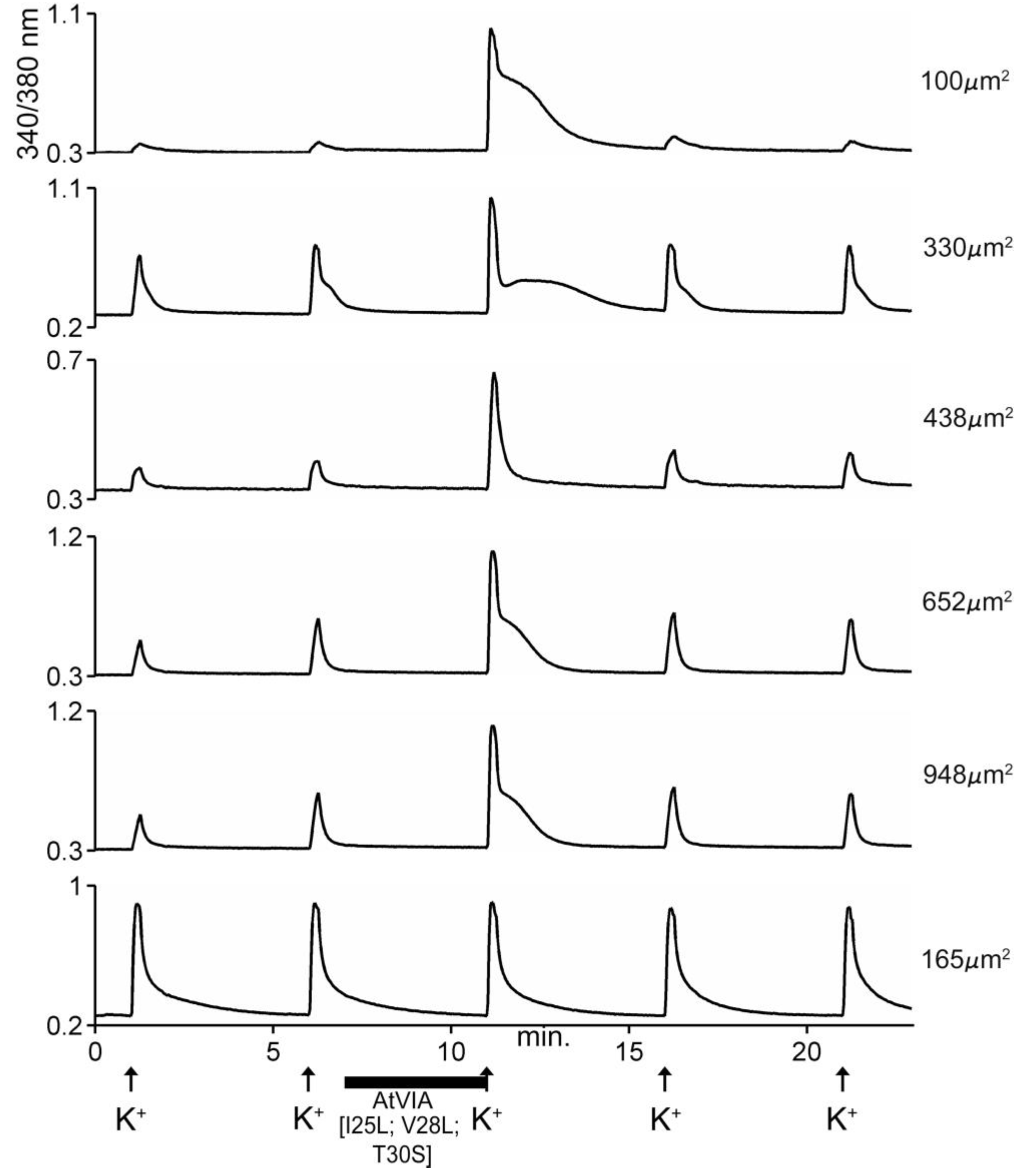
Biological Activity of AtVIA[I25L;V28L;T30S]
3. Discussion
4. Materials and Methods
4.1. Field Collection and Venom Extraction
4.2. Venom Fractionation
4.3. Mass Spectrometry and Sequence Determination
Preparation of Genomic DNA and Characterization of Clones Encoding AtVIA
4.4. In Vivo Assay and Calcium Imaging Assay on DRG
4.5. Peptide Synthesis
4.5.1. AtVIA[I25L;V28L; T30S] Cleavage, Derivatization and Purification
4.5.2. Oxidative Folding of At6A[I25L;V28L;T30S]
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Endean, R.; Rudkin, C. Further Studies of the Venoms of Conidae. Toxicon 1965, 2, 225–249. [Google Scholar] [CrossRef]
- Fainzilber, M.; Lodder, J.C.; Kits, K.S.; Kofman, O.; Vinnitsky, I.; Van Rietschoten, J.; Zlotkin, E.; Gordon, D. A new conotoxin affecting sodium current inactivation interacts with the d-conotoxin receptor site. J. Biol. Chem. 1995, 270, 1123–1129. [Google Scholar] [CrossRef] [PubMed]
- Shon, K.J.; Hasson, A.; Spira, M.E.; Cruz, L.J.; Gray, W.R.; Olivera, B.M. Delta-Conotoxin GmVIA, a novel peptide from the venom of Conus gloriamaris. Biochemistry 1994, 33, 11420–11425. [Google Scholar] [CrossRef] [PubMed]
- Shon, K.-J.; Grilley, M.M.; Marsh, M.; Yoshikami, D.; Hall, A.R.; Kurz, B.; Gray, W.R.; Imperial, J.S.; Hillyard, D.R.; Olivera, B.M. Purification, Characterization, Synthesis, and Cloning of the Lockjaw Peptide from Conus purpurascens Venom. Biochemistry 1995, 34, 4913–4918. [Google Scholar] [CrossRef] [PubMed]
- West, P.J.; Bulaj, G.; Yoshikami, D. Effects of delta-conotoxins PVIA and SVIE on sodium channels in the amphibian sympathetic nervous system. J. Neurophysiol 2005, 94, 3916–3924. [Google Scholar] [CrossRef] [PubMed]
- Aman, J.W.; Imperial, J.; Ueberheide, B.; Zhang, M.-M.; Aguilar, M.; Taylor, D.; Watkins, M.; Yoshikami, D.; Showers-Corneli, P.; Safavi-Hermami, H.; et al. Insights into the origins of fish-hunting in venomous cone snails from studies on Conus tessulatus. Proc. Natl. Acad. Sci. USA 2015, 112, 5087–5092. [Google Scholar] [CrossRef]
- Jin, A.-H.; Israel, M.R.; Inserra, M.C.; Smith, J.J.; Lewis, R.J.; Alewood, P.F.; Vetter, I.; Dutertre, S. δ-Conotoxin SuVIA suggests an evolutionary link between ancestral predator defence and the origin of fish-hunting behaviour in carnivorous cone snails. Proc. R. Soc. B Biol. Sci. 2015, 282, 201508170. [Google Scholar] [CrossRef] [PubMed]
- Puillandre, N.; Bouchet, P.; Duda Jr, T.F.; Kauferstein, S.; Kohn, A.J.; Olivera, B.M.; Watkins, M.; Meyer, C. Molecular phylogeny and evolution of the cone snails (Gastropoda conoidea). Mol. Phylogenet. Evol. 2014, 78, 290–303. [Google Scholar] [CrossRef] [PubMed]
- Cunha, R.L.; Castilho, R.; Ruber, L.; Zardoya, R. Patterns of cladogenesis in the venomous marine gastropod genus Conus from the Cape Verde islands. Syst. Biol. 2005, 54, 634–650. [Google Scholar] [CrossRef] [PubMed]
- Imperial, J.; Cabang, A.; Song, J.; Raghuraman, S.; Gajewiak, J.; Watkins, M.; Showers-Corneli, P.; Fedosov, A.; Concepcion, G.P.; Terlau, H.; et al. A Family of Excitatory Peptide Toxins from Venomous Crassispirine Snails: Using Constellation Pharmacology to Assess Bioactivity. Toxicon 2014, 89, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Teichert, R.W.; Raghuraman, S.; Memon, T.; Cox, J.L.; Foulkes, T.; Rivier, J.E.; Olivera, B.M. Characterization of two neuronal subclasses through constellation pharmacology. Proc. Natl. Acad. Sci. USA 2012, 109, 12758–12763. [Google Scholar] [CrossRef]
- Chen, P.; Garrett, J.E.; Watkins, M.; Olivera, B.M. Purification and characterization of a novel excitatory peptide from Conus distans venom that defines a novel gene superfamily of conotoxins. Toxicon 2008, 52, 139–145. [Google Scholar] [CrossRef]
- Bulaj, G.; DeLaCruz, R.; Azimi-Zonooz, A.; West, P.; Watkins, M.; Yoshikami, D.; Olivera, B.M. δ-Conotoxin Structure/Function through a Cladistic Analysis. Biochemistry 2001, 40, 13201–13208. [Google Scholar] [CrossRef]
- Hasson, A.; Fainzilber, M.; Gordon, D.; Zlotkin, E.; Spira, M.E. Alteration of sodium currents by new peptide toxins from the venom of a molluscivorous Conus snail. Eur. J. Neurosci. 1993, 5, 56–64. [Google Scholar] [CrossRef]
- Tritsch, G.I.; Moore, G.F. Spontaneous decomposition of glutamine in cell culture media. Exp. Cell. Res. 1962, 28, 360–364. [Google Scholar] [CrossRef]
- DeLa Cruz, R.; Whitby, F.G.; Buczek, O.; Bulaj, G. Detergent-assisted oxidative folding of delta-conotoxins. J. Pept. Res. 2003, 61, 202–212. [Google Scholar] [CrossRef]
- Kenyon, G.L.; Bruice, T.W. Novel sulfhydryl reagents. Methods Enzymol. 1977, 47, 407–430. [Google Scholar]
- Luo, S.; Zhangsun, S.D.; Hu, Y.; Zhu, X.; Wu, Y.; McIntosh, J.M. Alpha-Conotoxin LvIA/LVD21, Its Drugs Combination and Application. Patent CN201210347966.3A, 19 September 2012. [Google Scholar]
- Peigneur, S.; Tytgat, J. When cone snails and spiders meet: Design of selective and potent sodium channel inhibitors. Toxicon 2014, 91, 170. [Google Scholar] [CrossRef]
- Duda, T.F., Jr.; Kohn, A.J. Species-level phylogeography and evolutionary history of the hyperdiverse marine gastropod genus Conus. Mol. Phylogenet. Evol. 2005, 34, 257–272. [Google Scholar] [CrossRef]
- Cunha, R.L.; Tenorio, M.J.; Afonso, C.; Castilho, R.; Zardoya, R. Replaying the tape: Recurring biogeographical patterns in Cape Verde Conus after 12 million years. Mol. Ecol. 2008, 17, 885–901. [Google Scholar] [CrossRef]
- Dutertre, S.; Jin, A.H.; Vetter, I.; Hamilton, B.; Sunagar, K.; Lavergne, V.; Dutertre, V.; Fry, B.G.; Antunes, A.; Venter, D.J.; et al. Evolution of separate predation-and defence-evoked venoms in carnivorous cone snails. Nat. Commun. 2014, 5, 3521. [Google Scholar] [CrossRef]
- Imperial, J.; Silverton, N.; Olivera, B.M.; Bandyopadhyay, P.; Sporning, A.; Ferber, M.; Terlau, H. Using chemistry to reconstruct evolution: On the origins of fish-hunting in venomous cone snails. Proc. Am. Philos. Soc. 2007, 151, 185–200. [Google Scholar]
- Olivera, B.M.; Fedosov, A.; Imperial, J.S.; Kantor, Y. Physiology and Pharmacology of Conoidean Venoms. In Physiology of Molluscs; Saleuddin, A.S., Mukai, S.T., Eds.; Apple Academic Press: Palm Bay, FL, USA, 2015. [Google Scholar]
- Cotto-Rios, X.M.; Bekes, M.; Chapman, J.; Ueberheide, B.; Huang, T.T. Deubiquitinases as a signaling target of oxidative stress. Cell Rep. 2012, 2, 1475–1484. [Google Scholar] [CrossRef]
- Olivera, B.M.; Walker, C.; Cartier, G.E.; Hooper, D.; Santos, A.D.; Schoenfeld, R.; Shetty, R.; Watkins, M.; Bandyopadhyay, P.; Hillyard, D.R. Speciation of cone snails and interspecific hyperdivergence of their venom peptides. Potential evolutionary significance of introns. Ann. N. Y. Acad. Sci. 1999, 870, 223–237. [Google Scholar] [CrossRef]
- Olivera, B.M.; Cruz, L.J.; Yoskikami, D. Effects of Conus peptides on the behavior of mice. Curr. Opin. Neurobiol. 1999, 9, 772–777. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subgenus (Prey) | -Conotoxin | Conus Species | Sequence | Ref. |
|---|---|---|---|---|
| A. | ||||
| Tesseliconus (worm) | -TsVIA | C. tessulatus | CAAFGSFCGLPGLVD--CCSGRCFIVCLL | [6] |
| Tesseliconus (worm) | -ErVIA | C. eburneus | CAGIGSFCGLPGLVD--CCSGRCFIVCLP | [6] |
| Tesseliconus (worm) | SuVIA | C. suturatus | CAGIGSFCGLPGLVD--CCSDRCFIVCLP | [7] |
| Kalloconus (worm) | -AtVIA | C. ateralbus | ZCGADGQFCFL-PGLGLNCCSGLCLIVCVPT | This work |
| B. | ||||
| Chelyconus (fish) | PVIA | C. purpurascens | EACYAOGTFCGIKOGL---CCSEFCLPGVCFG | [4] |
| Pionoconus (fish) | SVIE | C. striatus | DGCSSGGTFCGIHOGL---CCSEFCF-LWCITFID | [5] |
| Textilia (fish) | -BuVIA | C. bullatus | DECSAOGAFCLIROGL---CCSEFCF-FACF | [13] |
| Kalloconus (worm) | AtVIA | C. ateralbus | ZCGADGQFCFL-PGLGLNCCSGLCL-IVCVPT | This work |
| Tesseliconus (worm) | TsVIA | C. tessulatus | CAAFGSFCGL-PGLVD-CCSGRCF-IVCLL | [6] |
| Tesseliconus (worm) | ErVIA | C. eburneus | CAGIGSFCGL-PGLVD-CCSGRCF-IVCLP | [6] |
| Tesseliconus (worm) | SuVIA | C. suturatus | CAGIGSFCGL-PGLVD-CCSDRCF-IVCLP | [7] |
| Cylinder (snail) | TxVIA | C. textile | WCKQSGEMCNLLDQN---CCDGYCIVLV-CT | [14] |
| Cylinder (snail) | GmVIA | C. gloriamaris | VKPCRKEGQLCDPIFQN---CCRGWNCVLF-CV | [3] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neves, J.L.B.; Imperial, J.S.; Morgenstern, D.; Ueberheide, B.; Gajewiak, J.; Antunes, A.; Robinson, S.D.; Espino, S.; Watkins, M.; Vasconcelos, V.; et al. Characterization of the First Conotoxin from Conus ateralbus, a Vermivorous Cone Snail from the Cabo Verde Archipelago. Mar. Drugs 2019, 17, 432. https://doi.org/10.3390/md17080432
Neves JLB, Imperial JS, Morgenstern D, Ueberheide B, Gajewiak J, Antunes A, Robinson SD, Espino S, Watkins M, Vasconcelos V, et al. Characterization of the First Conotoxin from Conus ateralbus, a Vermivorous Cone Snail from the Cabo Verde Archipelago. Marine Drugs. 2019; 17(8):432. https://doi.org/10.3390/md17080432
Chicago/Turabian StyleNeves, Jorge L. B., Julita S. Imperial, David Morgenstern, Beatrix Ueberheide, Joanna Gajewiak, Agostinho Antunes, Samuel D. Robinson, Samuel Espino, Maren Watkins, Vitor Vasconcelos, and et al. 2019. "Characterization of the First Conotoxin from Conus ateralbus, a Vermivorous Cone Snail from the Cabo Verde Archipelago" Marine Drugs 17, no. 8: 432. https://doi.org/10.3390/md17080432
APA StyleNeves, J. L. B., Imperial, J. S., Morgenstern, D., Ueberheide, B., Gajewiak, J., Antunes, A., Robinson, S. D., Espino, S., Watkins, M., Vasconcelos, V., & Olivera, B. M. (2019). Characterization of the First Conotoxin from Conus ateralbus, a Vermivorous Cone Snail from the Cabo Verde Archipelago. Marine Drugs, 17(8), 432. https://doi.org/10.3390/md17080432